
Long-Term Effects of Very Low Dose Particle Radiation on Gene Expression in the Heart: Degenerative Disease Risks

 ,
,  , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Procedures
2.2. Irradiation Procedures
2.3. Tissue Collection and Sample Processing
2.4. RNA Extraction
2.5. Microarray Analysis
2.6. Batch Examination and Correction
2.7. Microarray Data Preprocessing
2.8. Analysis of Differential Gene Expression
2.9. Analysis of Dose-Dependent Changes in Gene Expression
2.10. Portrayal of Transcriptome Landscapes
2.11. qRT-PCR
2.12. Gene Enrichment Analyses
2.13. Statistical Analysis of qPCR Results
3. Results
3.1. Data Collection, Pre-Processing, and Mapping
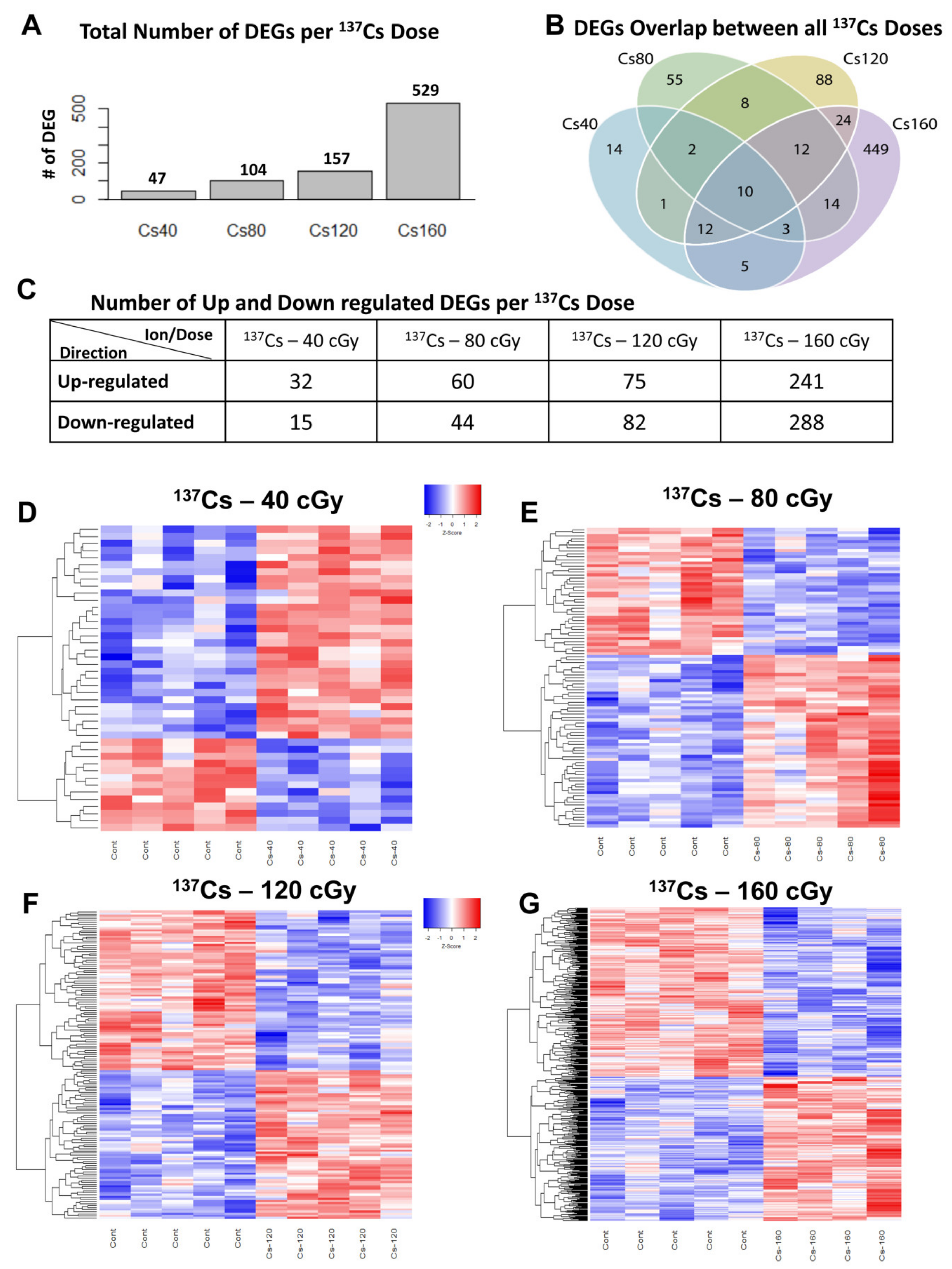
3.2. Different Gene Expression in Mouse Heart Tissue Following 137Cs Irradiation
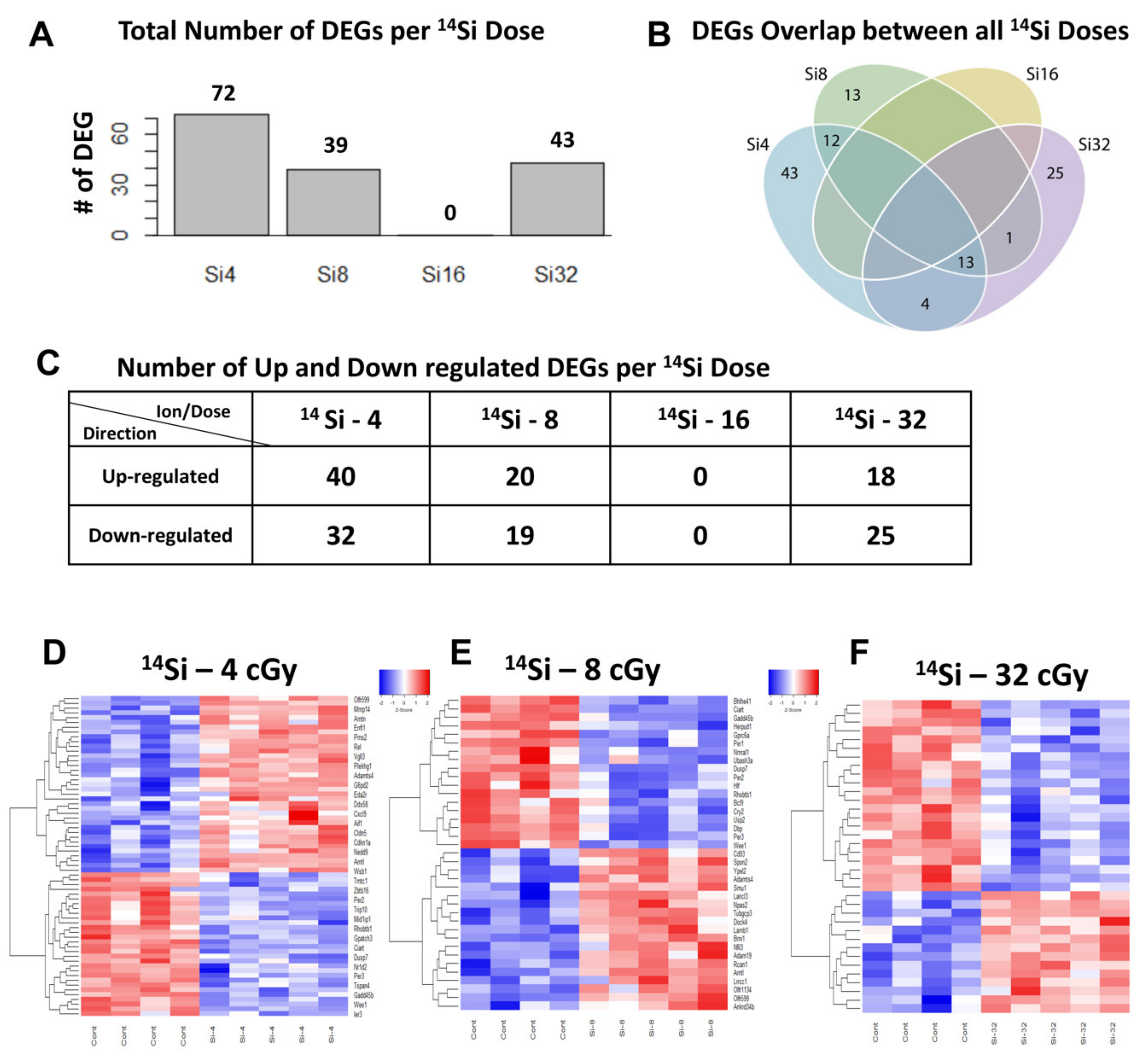
3.3. Different Gene Expression in Mouse Heart Tissue Following 14Si Irradiation
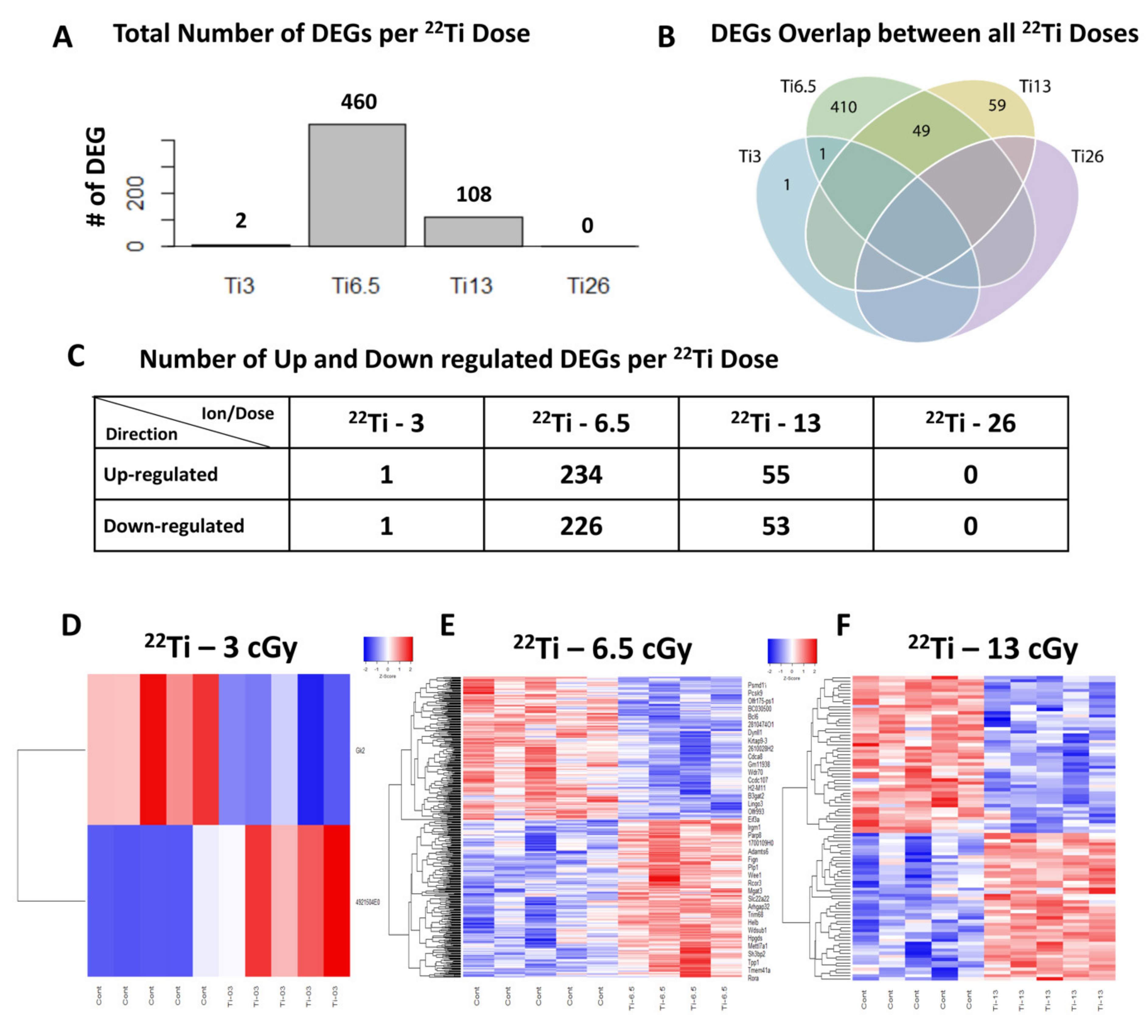
3.4. Different Gene Expression in Mouse Heart Tissue Following 22Ti Irradiation
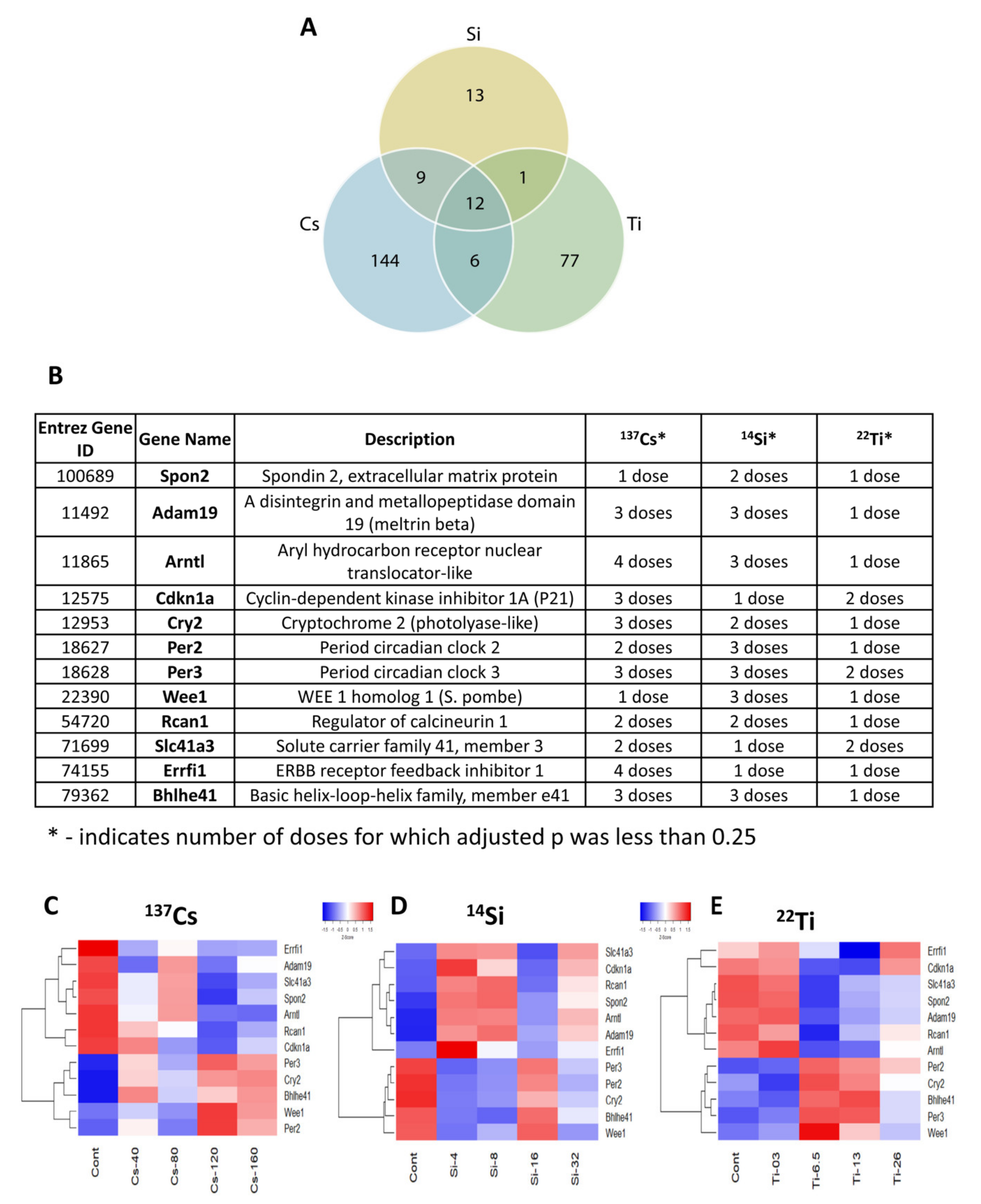
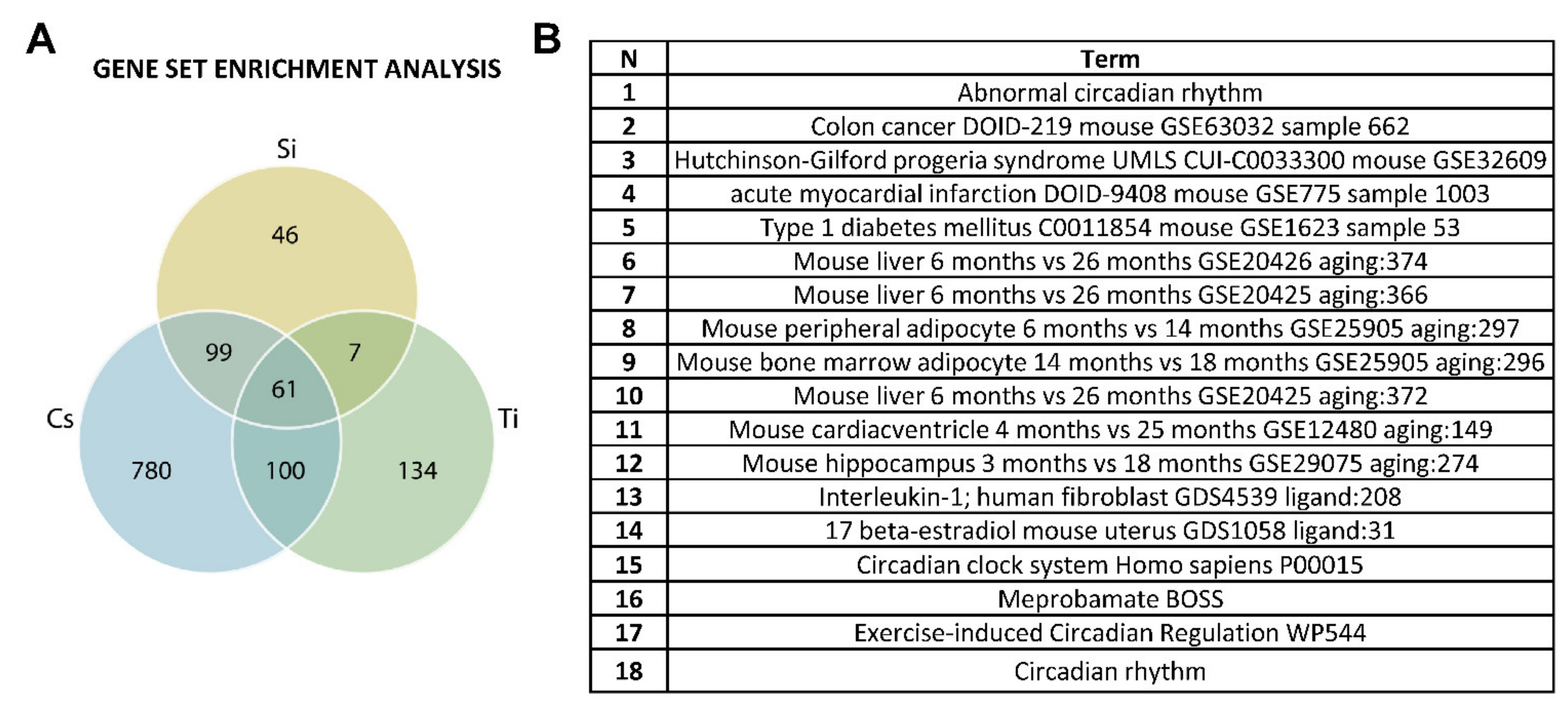
3.5. Common Differentially Expressed Genes across All Radiation Types
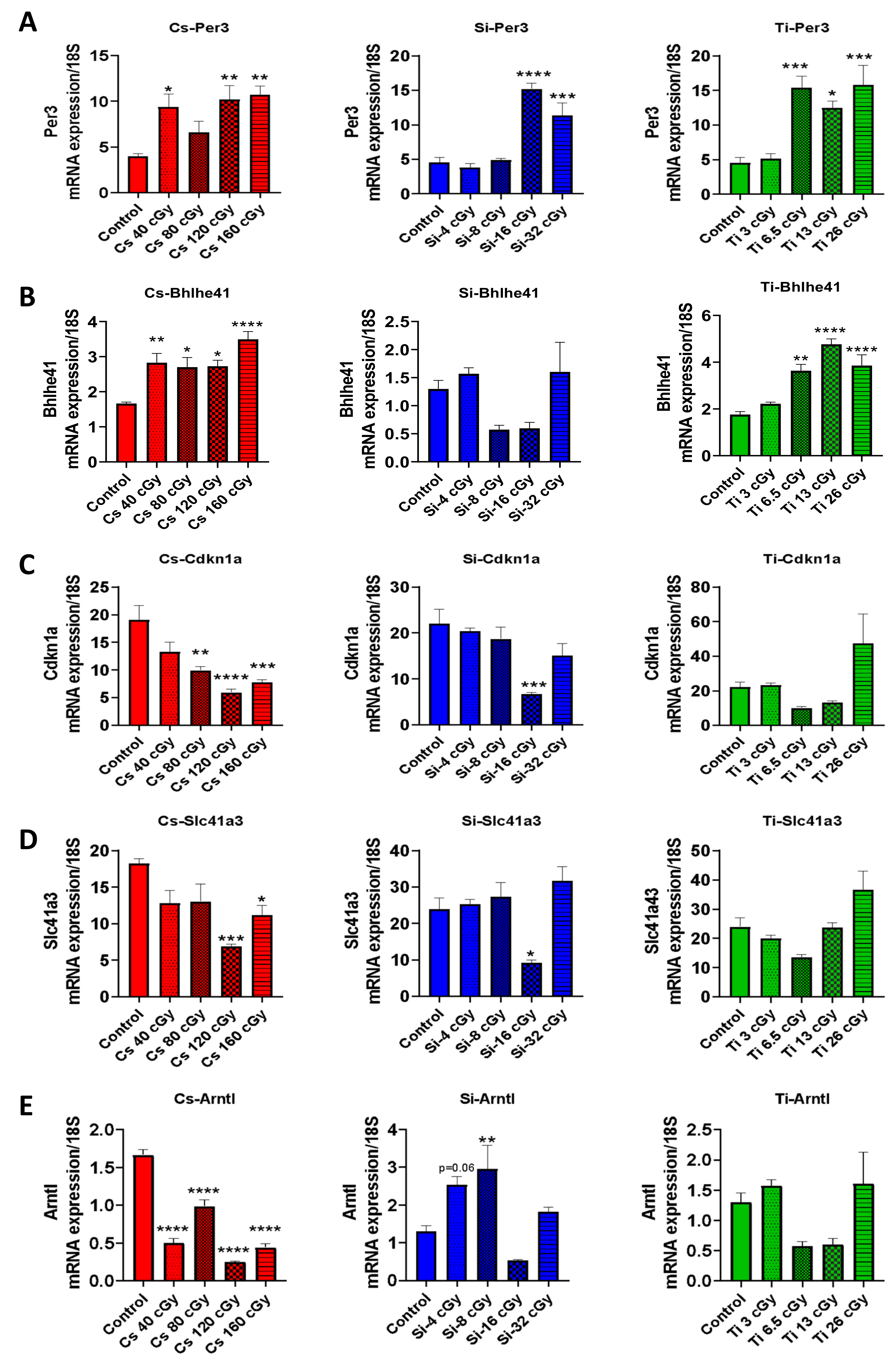
3.6. Microarray Analysis and Validation
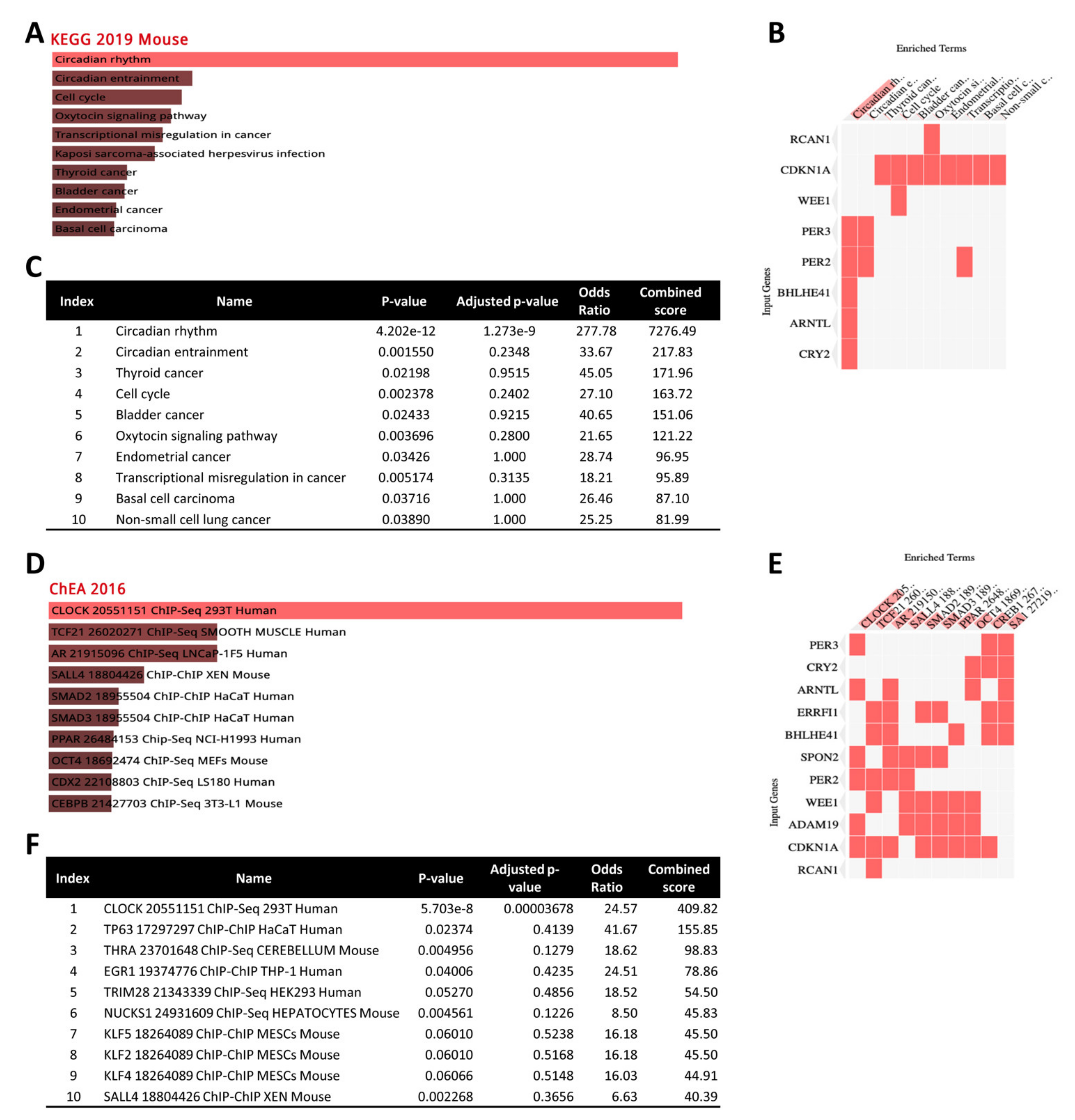
3.7. Gene Enrichment Analysis of 12 Common DEGs
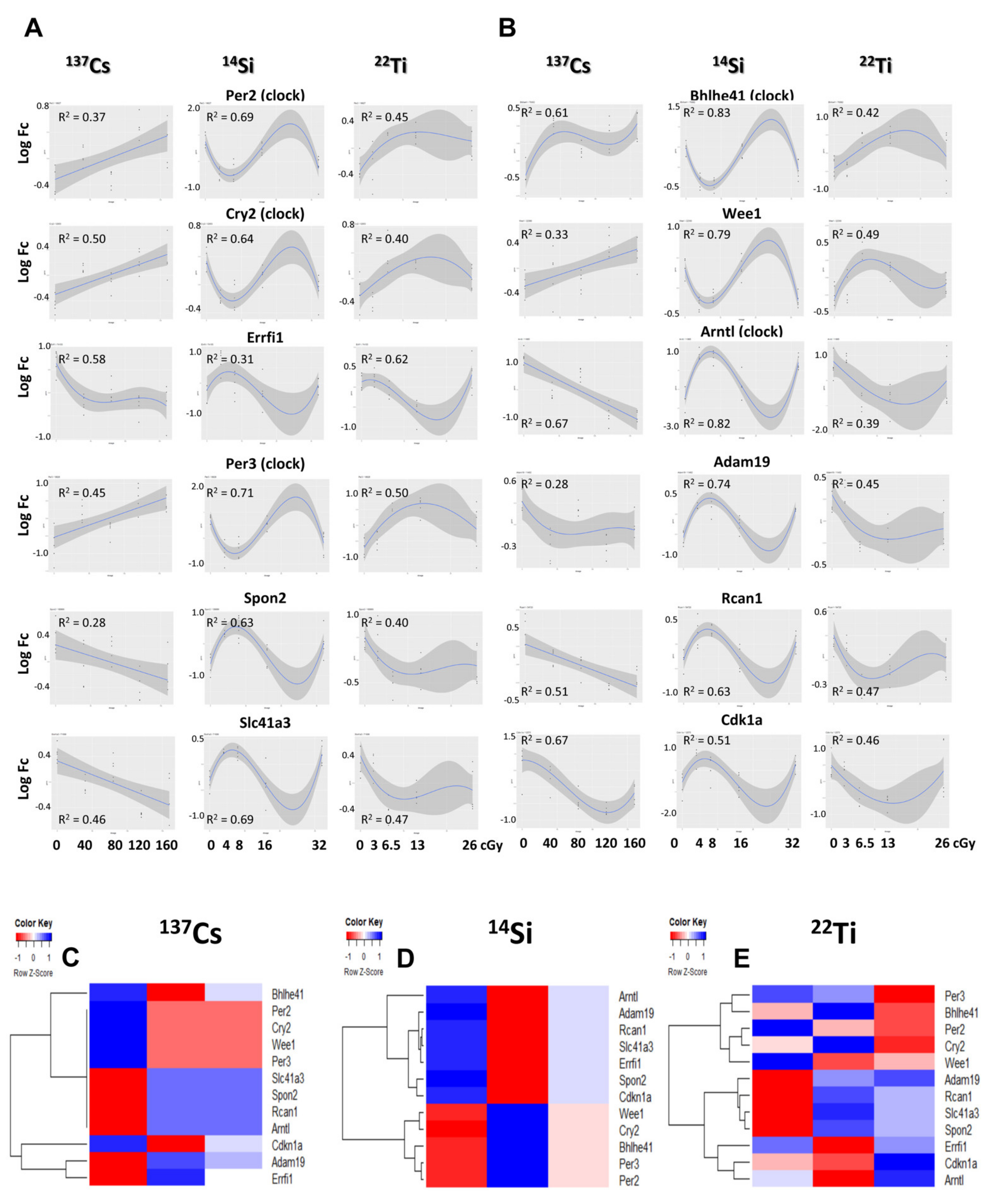
3.8. Regression Analysis of Common Differentially Expressed Genes across Radiation Type
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Czupalla, M.; Horneck, G.; Blome, H.J. The conceptual design of a hybrid life support system based on the evaluation and comparison of terrestrial testbeds. Adv. Space Res. Off. J. Comm. Space Res. (COSPAR) 2005, 35, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human Pathophysiological Adaptations to the Space Environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed]
- Reitz, G. Characteristic of the radiation field in low Earth orbit and in deep space. Z. Med. Phys. 2008, 18, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Norbury, J.W.; Schimmerling, W.; Slaba, T.C.; Azzam, E.I.; Badavi, F.F.; Baiocco, G.; Benton, E.; Bindi, V.; Blakely, E.A.; Blattnig, S.R.; et al. Galactic cosmic ray simulation at the NASA Space Radiation Laboratory. Life Sci. Space Res. 2016, 8, 38–51. [Google Scholar] [CrossRef]
- Council, N.R. Recapturing a Future for Space Exploration: Life and Physical Sciences Research for a New Era; The National Academies Press: Washington, DC, USA, 2011. [Google Scholar] [CrossRef]
- Shtifman, A.; Pezone, M.J.; Sasi, S.P.; Agarwal, A.; Gee, H.; Song, J.; Perepletchikov, A.; Yan, X.; Kishore, R.; Goukassian, D.A. Divergent modification of low-dose ⁵⁶Fe-particle and proton radiation on skeletal muscle. Radiat. Res. 2013, 180, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Sasi, S.P.; Song, J.; Park, D.; Enderling, H.; McDonald, J.T.; Gee, H.; Garrity, B.; Shtifman, A.; Yan, X.; Walsh, K.; et al. TNF-TNFR2/p75 signaling inhibits early and increases delayed nontargeted effects in bone marrow-derived endothelial progenitor cells. J. Biol. Chem. 2015, 290, 27014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delius, W.; Senbening, H.; Wirtzfeld, A.; Lutilsky, L. Atrial stimulation in the therapy of auricular flutter. Mmwmunchener Med. Wochenschr. 1974, 116, 479–484. [Google Scholar]
- Sasi, S.P.; Park, D.; Muralidharan, S.; Wage, J.; Kiladjian, A.; Onufrak, J.; Enderling, H.; Yan, X.; Goukassian, D.A. Particle Radiation-Induced Nontargeted Effects in Bone-Marrow-Derived Endothelial Progenitor Cells. Stem Cells Int. 2015, 2015, 496512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coleman, M.A.; Sasi, S.P.; Onufrak, J.; Natarajan, M.; Manickam, K.; Schwab, J.; Muralidharan, S.; Peterson, L.E.; Alekseyev, Y.O.; Yan, X.; et al. Low-dose radiation affects cardiac physiology: Gene networks and molecular signaling in cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1947–H1963. [Google Scholar] [CrossRef]
- Chang, P.Y.; Cucinotta, F.A.; Bjornstad, K.A.; Bakke, J.; Rosen, C.J.; Du, N.; Fairchild, D.G.; Cacao, E.; Blakely, E.A. Harderian Gland Tumorigenesis: Low-Dose and LET Response. Radiat. Res. 2016, 185, 449–460. [Google Scholar] [CrossRef]
- Leek, J.T.; Johnson, W.E.; Parker, H.S.; Jaffe, A.E.; Storey, J.D. The sva package for removing batch effects and other unwanted variation in high-throughput experiments. Bioinformatics 2012, 28, 882–883. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Oytam, Y.; Sobhanmanesh, F.; Duesing, K.; Bowden, J.C.; Osmond-McLeod, M.; Ross, J. Risk-conscious correction of batch effects: Maximising information extraction from high-throughput genomic datasets. BMC Bioinform. 2016, 17, 332. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Bushel, P.; Chu, T.-M.; Wolfinger, R. Principal Variance Components Analysis: Estimating Batch Effects in Microarray Gene Expression Data. In Batch Effects and Noise in Microarray Experiments: Sources and Solutions; Wiley: Hoboken, NJ, USA, 2009; pp. 141–154. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loffler-Wirth, H.; Kalcher, M.; Binder, H. oposSOM: R-package for high-dimensional portraying of genome-wide expression landscapes on bioconductor. Bioinformatics 2015, 31, 3225–3227. [Google Scholar] [CrossRef]
- Wirth, H.; Loffler, M.; von Bergen, M.; Binder, H. Expression cartography of human tissues using self organizing maps. BMC Bioinform. 2011, 12, 306. [Google Scholar] [CrossRef] [Green Version]
- Kusano, K.F.; Pola, R.; Murayama, T.; Curry, C.; Kawamoto, A.; Iwakura, A.; Shintani, S.; Ii, M.; Asai, J.; Tkebuchava, T.; et al. Sonic hedgehog myocardial gene therapy: Tissue repair through transient reconstitution of embryonic signaling. Nat. Med. 2005, 11, 1197–1204. [Google Scholar] [CrossRef]
- Preitner, N.; Damiola, F.; Lopez-Molina, L.; Zakany, J.; Duboule, D.; Albrecht, U.; Schibler, U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell 2002, 110, 251–260. [Google Scholar] [CrossRef]
- Lopez-Molina, L.; Conquet, F.; Dubois-Dauphin, M.; Schibler, U. The DBP gene is expressed according to a circadian rhythm in the suprachiasmatic nucleus and influences circadian behavior. EMBO J. 1997, 16, 6762–6771. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.M.; Shay, J.; Zeng, T.C.; Chou, J.L.; Huang, T.H.; Lai, H.C.; Chan, M.W. Epigenetic silencing of ARNTL, a circadian gene and potential tumor suppressor in ovarian cancer. Int. J. Oncol. 2014, 45, 2101–2107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Zhou, X.; Fang, Z.; Pan, Z. Effect of STC2 gene silencing on colorectal cancer cells. Mol. Med. Rep. 2019, 20, 977–984. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Arimura, T.; Itoh-Satoh, M.; Ueda, K.; Hohda, S.; Inagaki, N.; Takahashi, M.; Hori, H.; Yasunami, M.; Nishi, H.; et al. Tcap gene mutations in hypertrophic cardiomyopathy and dilated cardiomyopathy. J. Am. Coll. Cardiol. 2004, 44, 2192–2201. [Google Scholar] [CrossRef] [Green Version]
- Amoroso, F.; Falzoni, S.; Adinolfi, E.; Ferrari, D.; Di Virgilio, F. The P2X7 receptor is a key modulator of aerobic glycolysis. Cell Death Dis. 2012, 3, e370. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, X.; Gao, J.; Xu, C.; Xu, P.; Li, Y.; Zhu, Y.; Yu, C. Long noncoding RNA FLRL2 alleviated nonalcoholic fatty liver disease through Arntl-Sirt1 pathway. FASEB J. 2019, 33, 11411–11419. [Google Scholar] [CrossRef]
- Tan, K.L.; Board, P.G. Purification and characterization of a recombinant human Theta-class glutathione transferase (GSTT2-2). Biochem. J. 1996, 315 Pt 3, 727–732. [Google Scholar] [CrossRef] [Green Version]
- Marnett, L.J. Oxy radicals, lipid peroxidation and DNA damage. Toxicology 2002, 181–182, 219–222. [Google Scholar] [CrossRef]
- Landi, S.; Gemignani, F.; Moreno, V.; Gioia-Patricola, L.; Chabrier, A.; Guino, E.; Navarro, M.; de Oca, J.; Capella, G.; Canzian, F.; et al. A comprehensive analysis of phase I and phase II metabolism gene polymorphisms and risk of colorectal cancer. Pharm. Genom. 2005, 15, 535–546. [Google Scholar] [CrossRef] [PubMed]
- Rissiek, B.; Haag, F.; Boyer, O.; Koch-Nolte, F.; Adriouch, S. P2X7 on Mouse T Cells: One Channel, Many Functions. Front. Immunol. 2015, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Wang, J.; Wang, X.; Li, X.; Song, J.; Fang, J.; Liu, X.; Liu, T.; Wang, D.; Li, Q.; et al. Pik3ip1 Is a Negative Immune Regulator that Inhibits Antitumor T-Cell Immunity. Clin. Cancer Res. 2019, 25, 6180–6194. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.N.; Zhu, G.J. Progress in the research of stanniocalcin. Sheng Li Ke Xue Jin Zhan 2008, 39, 225–228. [Google Scholar]
- Douglas, R.; Kellaway, L.; Mintz, M.; van Wageningen, G. The crossed nigrostriatal projection decussates in the ventral tegmental decussation. Brain Res. 1987, 418, 111–121. [Google Scholar] [CrossRef]
- He, X.; Zhu, Z.; Johnson, C.; Stoops, J.; Eaker, A.E.; Bowen, W.; DeFrances, M.C. PIK3IP1, a negative regulator of PI3K, suppresses the development of hepatocellular carcinoma. Cancer Res. 2008, 68, 5591–5598. [Google Scholar] [CrossRef] [Green Version]
- Yu, M.; Li, W.; Wang, Q.; Wang, Y.; Lu, F. Circadian regulator NR1D2 regulates glioblastoma cell proliferation and motility. Oncogene 2018, 37, 4838–4853. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Y.; He, H.T.; Yang, Q. MiR-589-5p is a potential prognostic marker of hepatocellular carcinoma and regulates tumor cell growth by targeting MIG-6. Neoplasma 2018, 65, 753–761. [Google Scholar] [CrossRef]
- Jia, W.; Li, H.; He, Y.W. The extracellular matrix protein mindin serves as an integrin ligand and is critical for inflammatory cell recruitment. Blood 2005, 106, 3854–3859. [Google Scholar] [CrossRef] [PubMed]
- Feinstein, Y.; Klar, A. The neuronal class 2 TSR proteins F-spondin and Mindin: A small family with divergent biological activities. Int. J. Biochem. Cell Biol. 2004, 36, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Wei, X.; Tang, Q.Z.; Feng, J.; Zhang, Y.; Liu, C.; Bian, Z.Y.; Zhang, L.F.; Chen, M.; Bai, X.; et al. Cardiac-specific mindin overexpression attenuates cardiac hypertrophy via blocking AKT/GSK3beta and TGF-beta1-Smad signalling. Cardiovasc. Res. 2011, 92, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badie, C.; Dziwura, S.; Raffy, C.; Tsigani, T.; Alsbeih, G.; Moody, J.; Finnon, P.; Levine, E.; Scott, D.; Bouffler, S. Aberrant CDKN1A transcriptional response associates with abnormal sensitivity to radiation treatment. Br. J. Cancer 2008, 98, 1845–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartel, A.L.; Serfas, M.S.; Tyner, A.L. p21—Negative regulator of the cell cycle. Proc. Soc. Exp. Biol. Med. 1996, 213, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.W.; Adami, G.R.; Wei, N.; Keyomarsi, K.; Elledge, S.J. The p21 Cdk-interacting protein Cip1 is a potent inhibitor of G1 cyclin-dependent kinases. Cell 1993, 75, 805–816. [Google Scholar] [CrossRef]
- Scatizzi, J.C.; Mavers, M.; Hutcheson, J.; Young, B.; Shi, B.; Pope, R.M.; Ruderman, E.M.; Samways, D.S.K.; Corbett, J.A.; Egan, T.M.; et al. The CDK domain of p21 is a suppressor of IL-1beta-mediated inflammation in activated macrophages. Eur. J. Immunol. 2009, 39, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Caballero, J.; Flores, J.M.; Garcia-Palencia, P.; Serrano, M. Tumor susceptibility of p21(Waf1/Cip1)-deficient mice. Cancer Res. 2001, 61, 6234–6238. [Google Scholar]
- Khanna, A.K. Enhanced susceptibility of cyclin kinase inhibitor p21 knockout mice to high fat diet induced atherosclerosis. J. Biomed. Sci. 2009, 16, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.A. Circadian clock-mediated control of stem cell division and differentiation: Beyond night and day. Development 2014, 141, 3105–3111. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.A.; Kowalska, E.; Dallmann, R. (Re)inventing the circadian feedback loop. Dev. Cell 2012, 22, 477–487. [Google Scholar] [CrossRef] [Green Version]
- Mallis, M.M.; DeRoshia, C.W. Circadian rhythms, sleep, and performance in space. Aviat. Space Environ. Med. 2005, 76, B94–B107. [Google Scholar]
- Chellappa, S.L.; Vujovic, N.; Williams, J.S.; Scheer, F. Impact of Circadian Disruption on Cardiovascular Function and Disease. Trends Endocrinol. Metab. 2019, 30, 767–779. [Google Scholar] [CrossRef]
- Koczor, C.A.; Ludlow, I.; Hight, R.S., 2nd; Jiao, Z.; Fields, E.; Ludaway, T.; Russ, R.; Torres, R.A.; Lewis, W. Ecstasy (MDMA) Alters Cardiac Gene Expression and DNA Methylation: Implications for Circadian Rhythm Dysfunction in the Heart. Toxicol. Sci. Off. J. Soc. Toxicol. 2015, 148, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Bonney, S.; Hughes, K.; Harter, P.N.; Mittelbronn, M.; Walker, L.; Eckle, T. Cardiac period 2 in myocardial ischemia: Clinical implications of a light dependent protein. Int. J. Biochem. Cell Biol. 2013, 45, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Eckle, T.; Hartmann, K.; Bonney, S.; Reithel, S.; Mittelbronn, M.; Walker, L.A.; Lowes, B.D.; Han, J.; Borchers, C.H.; Buttrick, P.M.; et al. Adora2b-elicited Per2 stabilization promotes a HIF-dependent metabolic switch crucial for myocardial adaptation to ischemia. Nat. Med. 2012, 18, 774–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.Y.; Wen, M.S.; Wang, H.W.; Hsieh, I.C.; Li, Y.; Liu, P.Y.; Lin, F.C.; Liao, J.K. Increased vascular senescence and impaired endothelial progenitor cell function mediated by mutation of circadian gene Per2. Circulation 2008, 118, 2166–2173. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 controls lipid metabolism by direct regulation of PPARgamma. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [Green Version]
- Jeyaraj, D.; Haldar, S.M.; Wan, X.; McCauley, M.D.; Ripperger, J.A.; Hu, K.; Lu, Y.; Eapen, B.L.; Sharma, N.; Ficker, E.; et al. Circadian rhythms govern cardiac repolarization and arrhythmogenesis. Nature 2012, 483, 96–99. [Google Scholar] [CrossRef]
- Arai, S.; Meagher, R.; Swearingen, M.; Myint, H.; Rich, E.; Martinson, J.; Klingemann, H. Infusion of the allogeneic cell line NK-92 in patients with advanced renal cell cancer or melanoma: A phase I trial. Cytotherapy 2008, 10, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Angelousi, A.; Kassi, E.; Nasiri-Ansari, N.; Weickert, M.O.; Randeva, H.; Kaltsas, G. Clock genes alterations and endocrine disorders. Eur. J. Clin. Investig. 2018, 48, e12927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samten, B. Regulation of B-1a cells: Another novel function of the basic helix-loop-helix transcriptional regulator BHLHE41. Cell. Mol. Immunol. 2017, 14, 802–804. [Google Scholar] [CrossRef] [Green Version]
- Asanoma, K.; Liu, G.; Yamane, T.; Miyanari, Y.; Takao, T.; Yagi, H.; Ohgami, T.; Ichinoe, A.; Sonoda, K.; Wake, N.; et al. Regulation of the Mechanism of TWIST1 Transcription by BHLHE40 and BHLHE41 in Cancer Cells. Mol. Cell. Biol. 2015, 35, 4096–4109. [Google Scholar] [CrossRef] [Green Version]
- Flynn-Evans, E.E.; Barger, L.K.; Kubey, A.A.; Sullivan, J.P.; Czeisler, C.A. Circadian misalignment affects sleep and medication use before and during spaceflight. NPJ Microgravity 2016, 2, 15019. [Google Scholar] [CrossRef]
- Tyagi, S.; Gupta, P.; Saini, A.S.; Kaushal, C.; Sharma, S. The peroxisome proliferator-activated receptor: A family of nuclear receptors role in various diseases. J. Adv. Pharm. Technol. Res. 2011, 2, 236–240. [Google Scholar] [CrossRef]
- De Filippis, D.; D’Amico, A.; Cinelli, M.P.; Esposito, G.; Di Marzo, V.; Iuvone, T. Adelmidrol, a palmitoylethanolamide analogue, reduces chronic inflammation in a carrageenin-granuloma model in rats. J. Cell. Mol. Med. 2009, 13, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Zahid, M.D.K.; Rogowski, M.; Ponce, C.; Choudhury, M.; Moustaid-Moussa, N.; Rahman, S.M. CCAAT/enhancer-binding protein beta (C/EBPbeta) knockdown reduces inflammation, ER stress, and apoptosis, and promotes autophagy in oxLDL-treated RAW264.7 macrophage cells. Mol. Cell. Biochem. 2020, 463, 211–223. [Google Scholar] [CrossRef]
- Tang, Q.Q.; Otto, T.C.; Lane, M.D. CCAAT/enhancer-binding protein beta is required for mitotic clonal expansion during adipogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 850–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giltiay, N.V.; Karakashian, A.A.; Alimov, A.P.; Ligthle, S.; Nikolova-Karakashian, M.N. Ceramide- and ERK-dependent pathway for the activation of CCAAT/enhancer binding protein by interleukin-1beta in hepatocytes. J. Lipid Res. 2005, 46, 2497–2505. [Google Scholar] [CrossRef] [Green Version]
- Menet, J.S.; Pescatore, S.; Rosbash, M. CLOCK:BMAL1 is a pioneer-like transcription factor. Genes Dev. 2014, 28, 8–13. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Martin, K.A. TCF21: Flipping the Phenotypic Switch in SMC. Circ. Res. 2020, 126, 530–532. [Google Scholar] [CrossRef]
- Zhang, J.; Tam, W.L.; Tong, G.Q.; Wu, Q.; Chan, H.Y.; Soh, B.S.; Lou, Y.; Yang, J.; Ma, Y.; Chai, L.; et al. Sall4 modulates embryonic stem cell pluripotency and early embryonic development by the transcriptional regulation of Pou5f1. Nat. Cell Biol. 2006, 8, 1114–1123. [Google Scholar] [CrossRef]
- Zeineddine, D.; Hammoud, A.A.; Mortada, M.; Boeuf, H. The Oct4 protein: More than a magic stemness marker. Am. J. Stem Cells 2014, 3, 74–82. [Google Scholar]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zi, Z.; Chapnick, D.A.; Liu, X. Dynamics of TGF-beta/Smad signaling. FEBS Lett. 2012, 586, 1921–1928. [Google Scholar] [CrossRef] [Green Version]
- Hata, A.; Chen, Y.G. TGF-beta Signaling from Receptors to Smads. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef]
- Massague, J. TGFbeta signalling in context. Nat. Rev. Mol. Cell Biol. 2012, 13, 616–630. [Google Scholar] [CrossRef]
- Cohen, P. Molecular mechanisms involved in the control of glycogenolysis in skeletal muscle by calcium ions and cyclic AMP. Biochem. Soc. Trans. 1987, 15, 999–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.H.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E.L. Androgen receptor: Structure, role in prostate cancer and drug discovery. Acta Pharmacol. Sin. 2015, 36, 3–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takakura, Y.; Hinoi, T.; Oue, N.; Sasada, T.; Kawaguchi, Y.; Okajima, M.; Akyol, A.; Fearon, E.R.; Yasui, W.; Ohdan, H. CDX2 regulates multidrug resistance 1 gene expression in malignant intestinal epithelium. Cancer Res. 2010, 70, 6767–6778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Liu, D.; Sun, X.; Yang, K.; Yao, J.; Cheng, C.; Wang, C.; Zheng, J. CDX2 inhibits the proliferation and tumor formation of colon cancer cells by suppressing Wnt/beta-catenin signaling via transactivation of GSK-3beta and Axin2 expression. Cell Death Dis. 2019, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, E.; Matthias, P.; Muller, M.M.; Schaffner, W. Rapid detection of octamer binding proteins with ’mini-extracts’, prepared from a small number of cells. Nucleic Acids Res. 1989, 17, 6419. [Google Scholar] [CrossRef] [Green Version]
- Koo, B.S.; Lee, S.H.; Kim, J.M.; Huang, S.; Kim, S.H.; Rho, Y.S.; Bae, W.J.; Kang, H.J.; Kim, Y.S.; Moon, J.H.; et al. Oct4 is a critical regulator of stemness in head and neck squamous carcinoma cells. Oncogene 2015, 34, 2317–2324. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, P.; Shao, M.; Zang, X.; Zhang, J.; Mao, F.; Qian, H.; Xu, W. SALL4 activates TGF-beta/SMAD signaling pathway to induce EMT and promote gastric cancer metastasis. Cancer Manag. Res. 2018, 10, 4459–4470. [Google Scholar] [CrossRef] [Green Version]
- Pawitan, Y.; Michiels, S.; Koscielny, S.; Gusnanto, A.; Ploner, A. False discovery rate, sensitivity and sample size for microarray studies. Bioinformatics 2005, 21, 3017–3024. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Hwang, J.T. Quick calculation for sample size while controlling false discovery rate with application to microarray analysis. Bioinformatics 2007, 23, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Tod, P.; Roka, B.; Kaucsar, T.; Szatmari, K.; Vizovisek, M.; Vidmar, R.; Fonovic, M.; Szenasi, G.; Hamar, P. Time-Dependent miRNA Profile during Septic Acute Kidney Injury in Mice. Int. J. Mol. Sci. 2020, 21, 5316. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Xie, X.; Lu, Z.; Liu, L.; Qu, Y.; Wu, S.; Li, Y.; Li, G.; Wang, H.; Xu, G. Establishment of a Colorectal Cancer-Related MicroRNA-mRNA Regulatory Network by Microarray and Bioinformatics. Front. Genet. 2020, 11, 560186. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ion | Dose (cGy) | Energy (MeV/n) | Entrance LET (keV/µ) |
|---|---|---|---|
| 137Cs | 0 40 | 0.662 | 0.8 |
| 80 | |||
| 120 | |||
| 160 | |||
| 14Si | 0 4 | 260 | 70 |
| 8 | |||
| 16 | |||
| 32 | |||
| 22Ti | 0 3 | 1000 | 100 |
| 6.5 | |||
| 13 | |||
| 26 |
| Gene | Primer |
|---|---|
| ARNTL | Probe-/56-FAM/ACC TGC TCC/ZEN/AGT GTT TCC TCA TCA/3IABkFQ/ |
| R-ATC CAC AGC TAG CCC AAA C | |
| F-CCA CCT CAG AGC CAT TGA TAC | |
| Slc41a3 | Probe-/56-FAM/CTG GTT TCT/ZEN/GTG CCT CCC TGA CTG/3IABkFQ/ |
| R-CAC TGA GGA CAT GAG GGA AAG | |
| F-CTT CTT CCT GGA CTG GTT ACT G | |
| Cdkn1a | Probe-/56-FAM/CAG CCT AGA/ZEN/ACA GGG ATG GCA GTT/3IABkFQ/ |
| R-GAG TCG GGA TAT TAC GGT TGA G | |
| F-CCA GCT AGG ATG ACA GTG AAG | |
| PER3 | Probe-/56-FAM/AGC CGG AAG/ZEN/GTC TCC TTC ATC ATT/3IABkFQ/ |
| R-TGG ACT CGT TCG GAC TTT ATG | |
| F-GCA CTC AGA ACG GAG AGT ATG | |
| Bhlhe41 | Probe-/56-FAM/AAC CGG AAG /ZEN/CCA CAG CTC ATA CAT/3IABkFQ/ |
| R-TGC CTG ACT TTC TTC CCT TAC | |
| F-TGA CTG TGA CAA GCT GAC TG |
| Gene Name | Extracellular | Plasma Membrane | Nucleus | Cytosol | Human Protein Atlas |
|---|---|---|---|---|---|
| Spon2 | Y | Plasma proteins, predicted membrane proteins, predicted secreted proteins | |||
| Adam19 | Y | Y | Plasma proteins, predicted membrane proteins | ||
| Arntl | Y | predicted intracellular proteins, transcription factors | |||
| Cdkn1a | Y | Y | Cancer-related genes, predicted intracellular proteins | ||
| Cry2 | Y | Y | Y | Predicted intracellular proteins | |
| Per2 | Y | Disease-related genes, predicted intracellular proteins | |||
| Per3 | Y | Y | Disease-related genes, predicted intracellular proteins | ||
| Wee1 | Y | Cancer-related genes, enzymes, plasma proteins, Predicted intracellular proteins | |||
| Rcan1 | Y | Y | Predicted intracellular proteins | ||
| Slc41a3 | Y | Predicted membrane proteins, transporters | |||
| Errfi1 | Y | Y | Y | Predicted intracellular proteins | |
| Bhlhe41 | Y | Predicted intracellular proteins, transcription factors |
| Term Description | Genes |
|---|---|
| RNA polymerase II-specific DNA-binding transcription factor binding | Arntl, Bhlhe41, Cry2, Per2 |
| Kinase binding | Cdkn1a, Cry2, Errfi1, Per2, Per3 |
| Transcription regulatory region sequence-specific DNA binding | Arntl, Bhlhe41, Cry2, Per2 |
| Histone deacetylase binding | Bhlhe41, Per2 |
| SH3 domain binding | Adam19, Errfi1 |
| Nuclear hormone receptor binding | Cry2, Per2 |
| Transcription regulator activity | Arntl, Bhlhe41, Cry2, Per2 |
| Signaling receptor binding | Arntl, Cry2, Per2, Spon2 |
| Protein kinase activity | Wee1 |
| Nucleic acid binding | Rcan1 |
| Cation transmembrane transporter activity | Slc41a3 |
| Spon2 | Adam19 | Arntl | Cdkn1a | Cry2 | Per2 | Per3 | Wee1 | Rcan1 | Slc41a3 | Errfi1 | Bhlhe41 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Circadian rhythm | + | + | + | + | + | |||||||
| ErbB signaling pathway | + | |||||||||||
| HIF-1 signaling pathway | + | |||||||||||
| FoxO signaling pathway | + | |||||||||||
| Cell cycle, circadian regulated | + | + | ||||||||||
| p53 signaling pathway | + | |||||||||||
| PI3K-Akt signaling pathway | + | |||||||||||
| Cellular senescence | + | |||||||||||
| JAK-STAT signaling pathway | + | |||||||||||
| Oxytocin signaling pathway | + | + | ||||||||||
| Transcriptional misregulation in cancer | + | + | ||||||||||
| Circadian entrainment | + | + | ||||||||||
| Acute myeloid leukemia | + | |||||||||||
| Human Immuno-deficiency virus 1 infection | + | |||||||||||
| Thyroid hormone signaling pathway | + |
| Icons | 137Cs | 14Si | 22Ti | |
|---|---|---|---|---|
| Type of Regression | Linear Regression | 423 | 0 | 1 |
| Non-Linear Regression | 215 | 53 | 123 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garikipati, V.N.S.; Arakelyan, A.; Blakely, E.A.; Chang, P.Y.; Truongcao, M.M.; Cimini, M.; Malaredy, V.; Bajpai, A.; Addya, S.; Bisserier, M.; et al. Long-Term Effects of Very Low Dose Particle Radiation on Gene Expression in the Heart: Degenerative Disease Risks. Cells 2021, 10, 387. https://doi.org/10.3390/cells10020387
Garikipati VNS, Arakelyan A, Blakely EA, Chang PY, Truongcao MM, Cimini M, Malaredy V, Bajpai A, Addya S, Bisserier M, et al. Long-Term Effects of Very Low Dose Particle Radiation on Gene Expression in the Heart: Degenerative Disease Risks. Cells. 2021; 10(2):387. https://doi.org/10.3390/cells10020387
Chicago/Turabian StyleGarikipati, Venkata Naga Srikanth, Arsen Arakelyan, Eleanor A. Blakely, Polly Y. Chang, May M. Truongcao, Maria Cimini, Vandana Malaredy, Anamika Bajpai, Sankar Addya, Malik Bisserier, and et al. 2021. "Long-Term Effects of Very Low Dose Particle Radiation on Gene Expression in the Heart: Degenerative Disease Risks" Cells 10, no. 2: 387. https://doi.org/10.3390/cells10020387
APA StyleGarikipati, V. N. S., Arakelyan, A., Blakely, E. A., Chang, P. Y., Truongcao, M. M., Cimini, M., Malaredy, V., Bajpai, A., Addya, S., Bisserier, M., Brojakowska, A., Eskandari, A., Khlgatian, M. K., Hadri, L., Fish, K. M., Kishore, R., & Goukassian, D. A. (2021). Long-Term Effects of Very Low Dose Particle Radiation on Gene Expression in the Heart: Degenerative Disease Risks. Cells, 10(2), 387. https://doi.org/10.3390/cells10020387