
Mechanical Mechanisms of Chromosome Segregation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
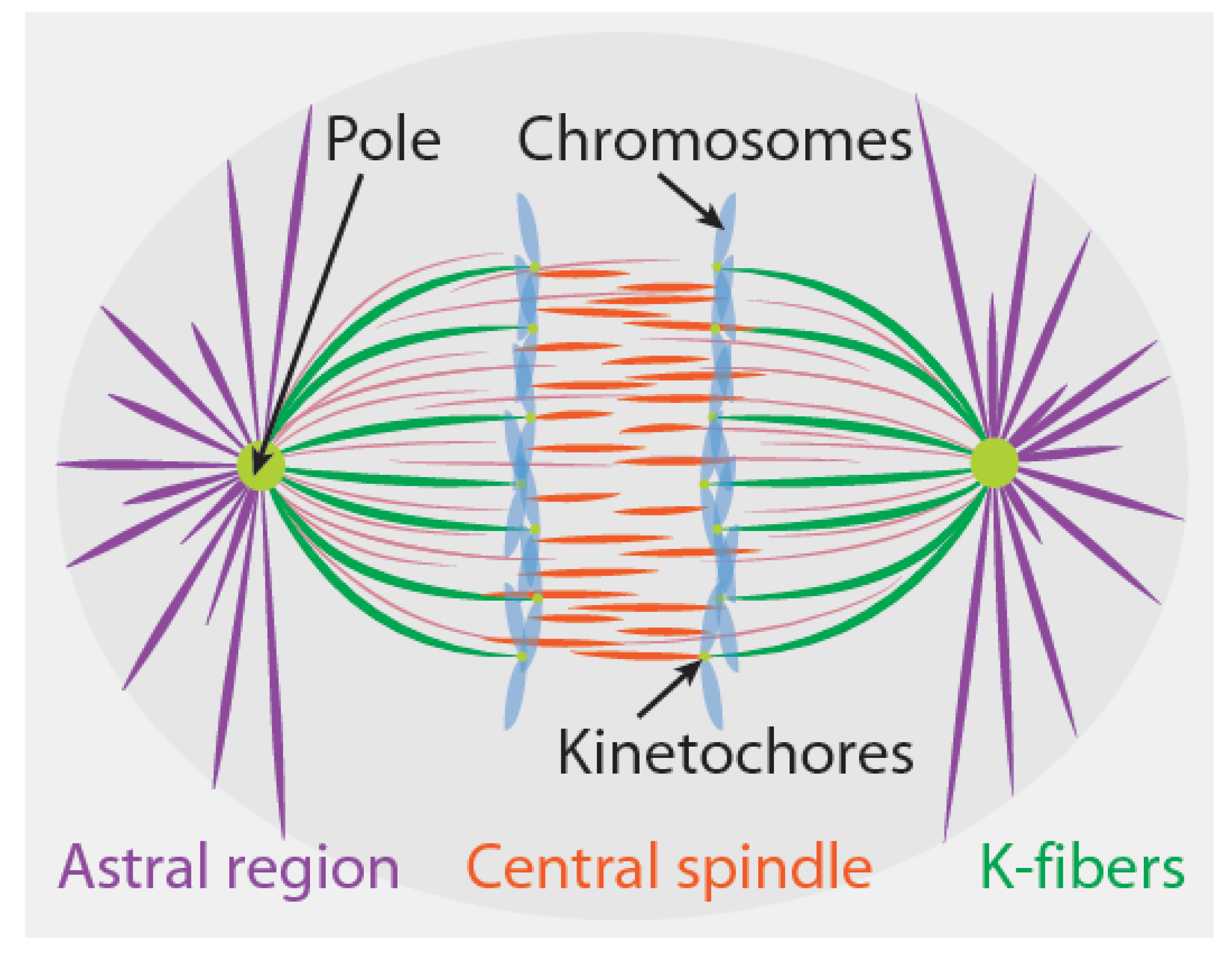
2. Force Generation in Anaphase Spindles
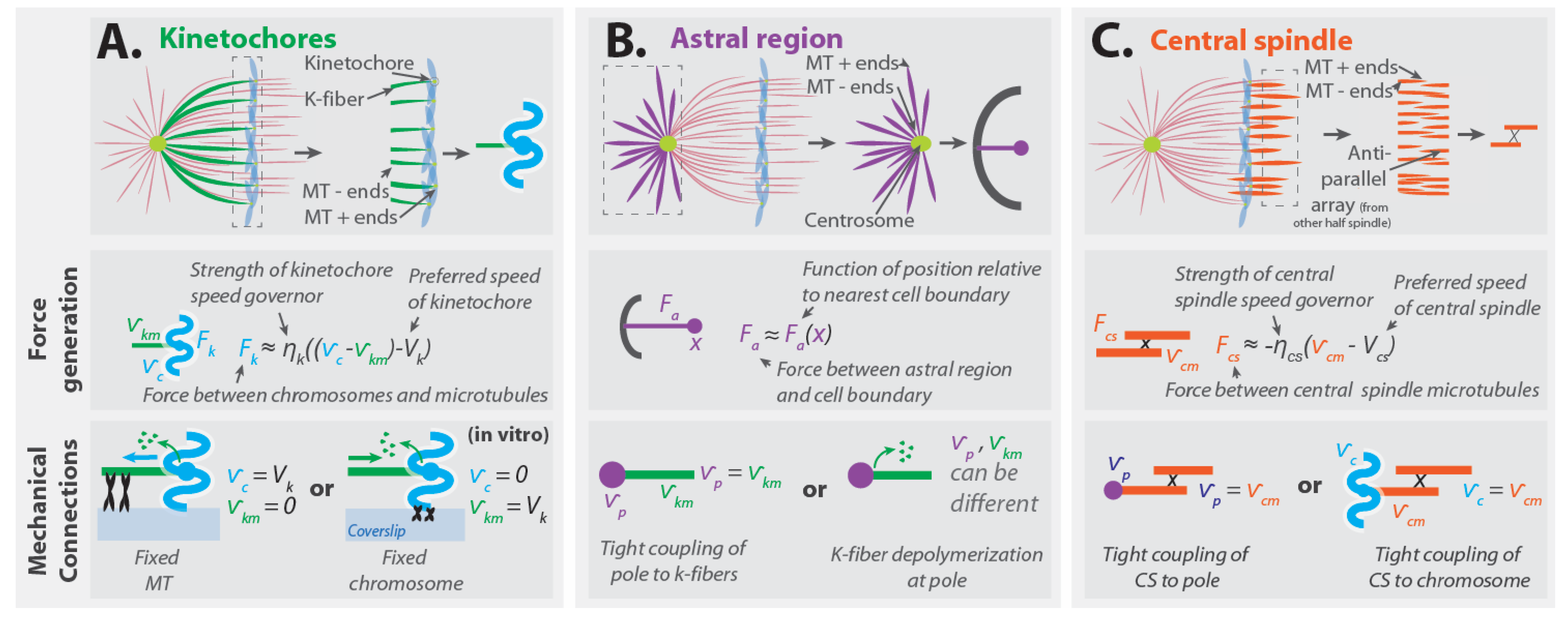
2.1. Kinetochores
2.2. The Astral Region
2.3. The Central Spindle
3. Mechanics and Its Relevance to Anaphase Spindles
3.1. Force Balance
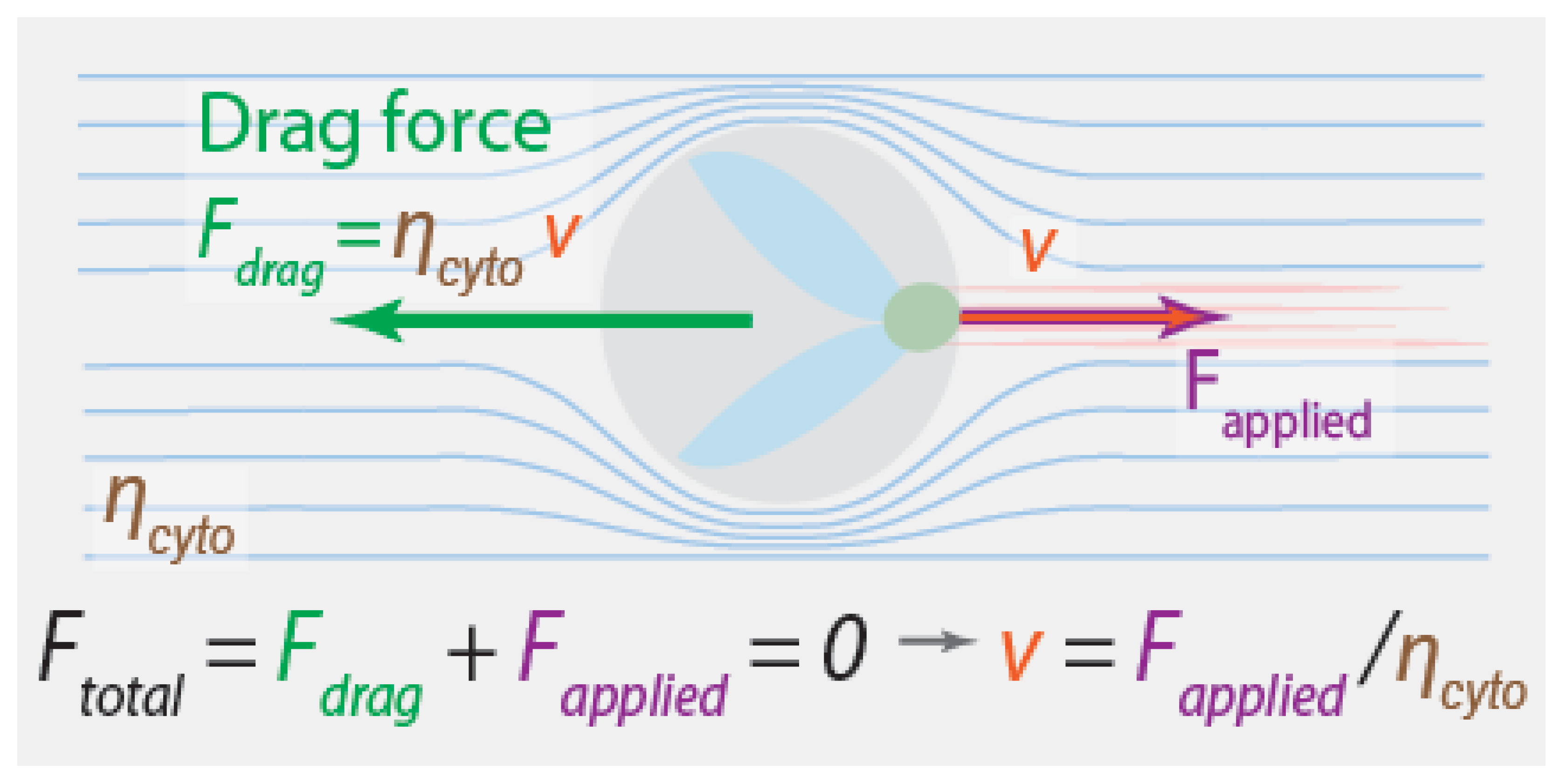
3.2. The Relevance, or Lack Thereof, of Fluid Drag Forces
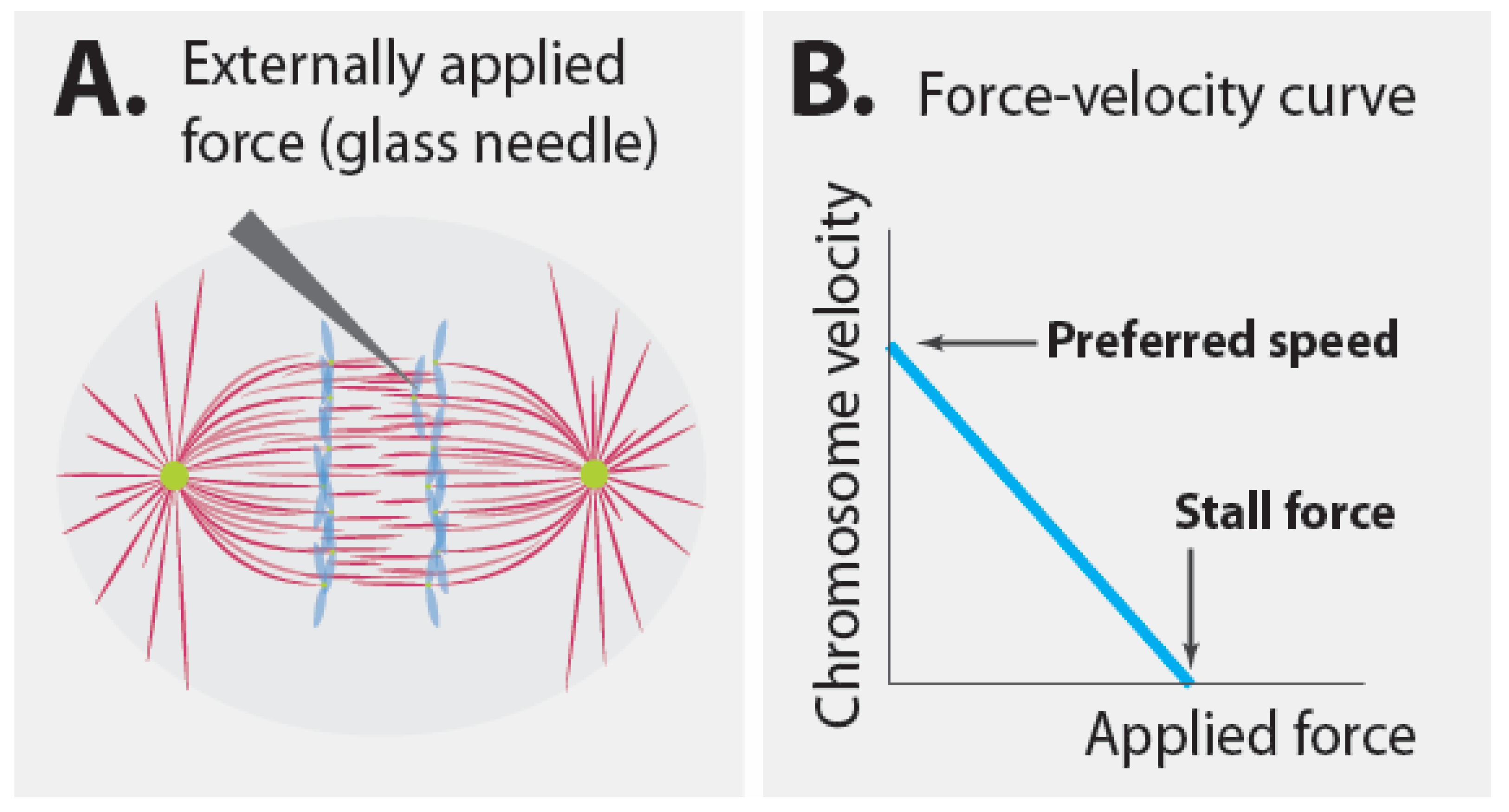
3.3. Mechanics of Speed Governors
3.4. Forces Are Local and Dependent on Relative Positions and Velocities
3.4.1. Implications for Forces from Kinetochores
3.4.2. Implications for Forces from the Astral Region
3.4.3. Implications for Forces from the Central Spindle
4. Mechanics of Chromosome Segregation
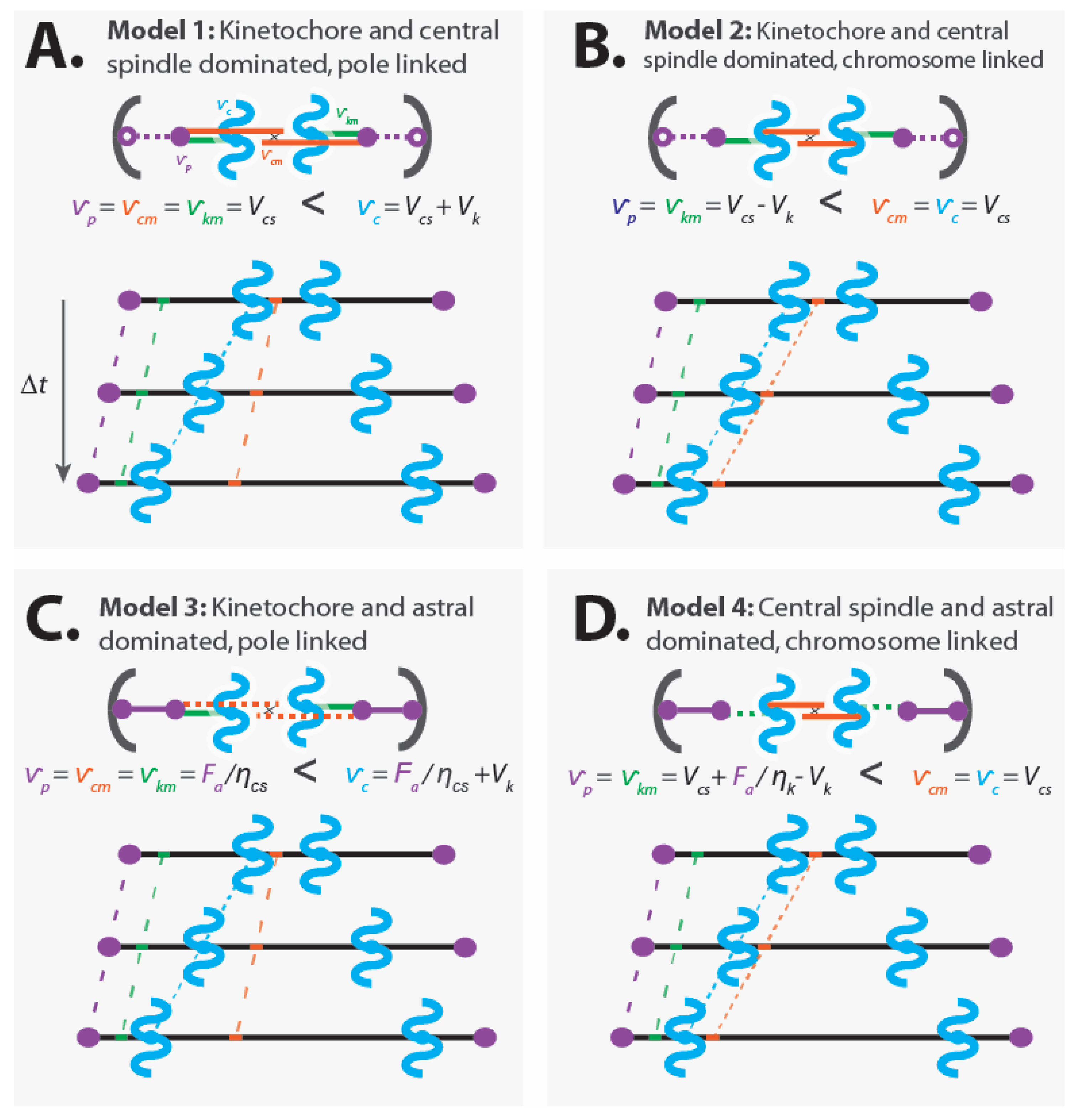
4.1. Four Mechanical Models of Anaphase
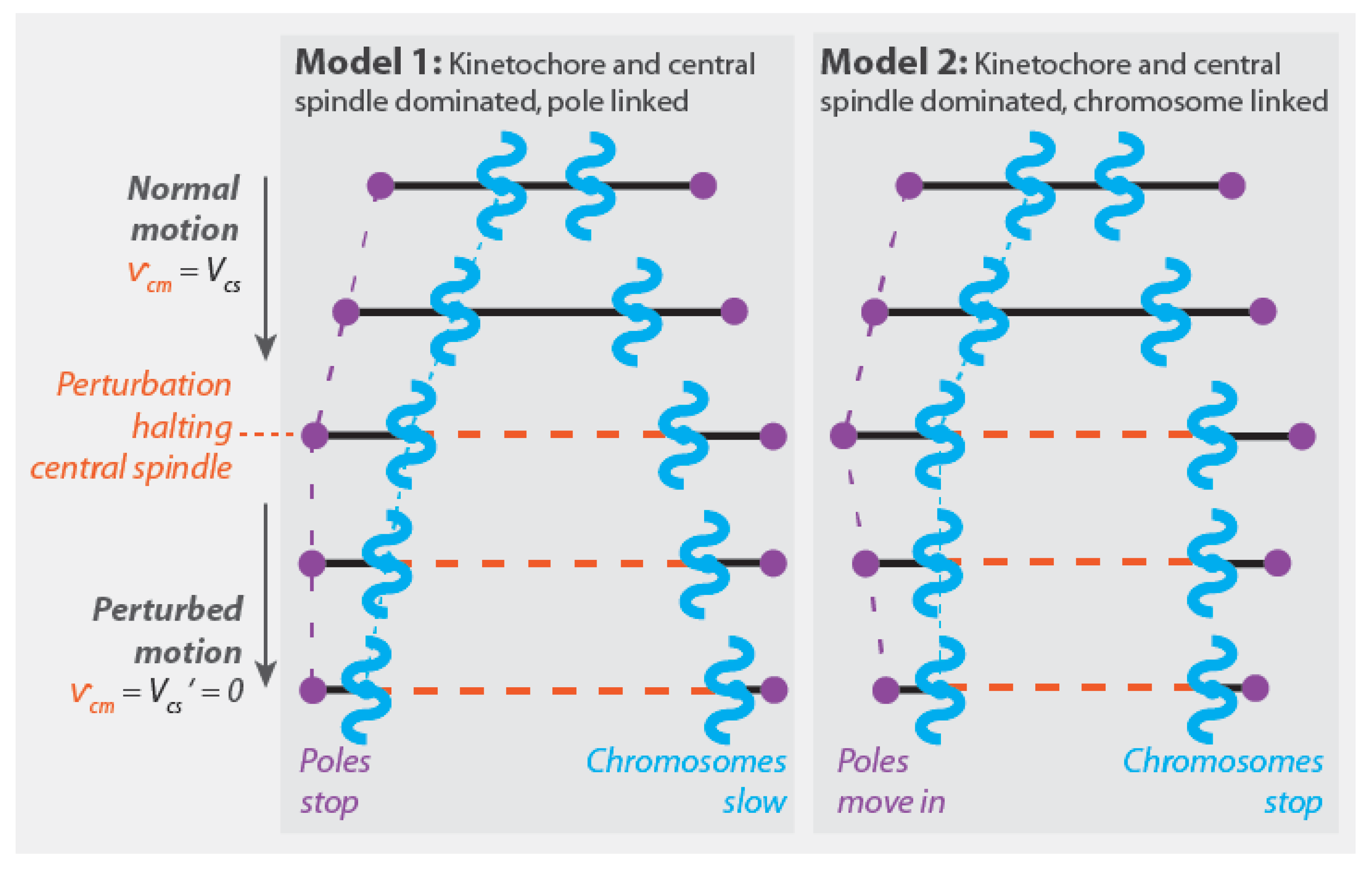
4.1.1. Model 1: Kinetochore- and Central Spindle-Dominated, Pole-Linked
- Dominant kinetochores: (from Equation (2))
- Dominant central spindle: (from Equation (3))
- Central spindle microtubules are linked to the pole:
- Kinetochore microtubules are linked to the pole:
- Astral pulling forces position the spindle, but do not alter its internal dynamics
- Pole movement:
- Chromatid movement:
- Central spindle microtubule movement:
- Kinetochore microtubule movement:
4.1.2. Model 2: Kinetochore- and Central Spindle-Dominated, Chromosome-Linked
- Dominant kinetochores: (from Equation (2))
- Dominant central spindle: (from Equation (3))
- Central spindle microtubules are linked to the chromosomes:
- Kinetochore microtubules are linked to the pole:
- Astral pulling forces position the spindle, but do not alter its internal dynamics
- Pole movement:
- Chromatid movement:
- Central spindle microtubule movement:
- Kinetochore microtubule movement:
4.1.3. Model 3: Kinetochore- and Astral-Dominated, Pole-Linked
- Dominant kinetochores: (from Equation (2))
- Central spindle microtubules are linked to the pole:
- Kinetochore microtubules are linked to the pole:
- Subordinate central spindle mechanics that act passively (), with a force on central spindle microtubules of: (from Equation (3))
- Force on poles is a balance from astral pulling force, , and tension from the central spindle microtubules:
- Pole movement:
- Chromatid movement:
- Central spindle microtubule movement:
- Kinetochore microtubule movement:
4.1.4. Model 4: Central Spindle- and Astral-Dominated, Chromosome-Linked
- Dominant central spindle: (from Equation (3))
- Central spindle microtubules are linked to the chromosomes:
- Kinetochore microtubules are linked to the pole:
- Kinetochore mechanics results in a force on kinetochore microtubules of: (from Equation (2))
- Force on poles is a balance from astral pulling force, , and tension from the kinetochore microtubules:
- Pole movement:
- Chromatid movement:
- Central spindle microtubule movement:
- Kinetochore microtubule movement:
4.1.5. Mechanism of Chromosome Segregation in the Four Models of Anaphase
4.2. Additional Mechanical Models of Anaphase
5. Conclusions and Outlook
Author Contributions
Funding
Conflicts of Interest
References
- Weinberg, R.A. The Biology of Cancer, 2nd illus. ed.; Garland Science: New York, NY, USA, 2013. [Google Scholar]
- McIntosh, J.R.; Hays, T. A Brief History of Research on Mitotic Mechanisms. Biology 2016, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Pavin, N.; Tolić, I.M. Mechanobiology of the Mitotic Spindle. Dev. Cell 2021, 56, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Maiato, H.; Lince-Faria, M. The perpetual movements of anaphase. Cell. Mol. Life Sci. 2010, 67, 2251–2269. [Google Scholar] [CrossRef]
- Civelekoglu-Scholey, G.; Scholey, J.M. Mitotic force generators and chromosome segregation. Cell. Mol. Life Sci. 2010, 67, 2231–2250. [Google Scholar] [CrossRef] [Green Version]
- Dumont, S.; Mitchison, T.J. Force and Length in the Mitotic Spindle. Curr. Biol. 2009, 19, R749–R761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntosh, J.R.; Molodtsov, M.I.; Ataullakhanov, F.I. Biophysics of mitosis. Q. Rev. Biophys. 2012, 45, 147–207. [Google Scholar] [CrossRef]
- Wickstead, B.; Gull, K. A “holistic” kinesin phylogeny reveals new kinesin families and predicts protein functions. Mol. Biol. Cell 2006, 17, 1734–1743. [Google Scholar] [CrossRef] [Green Version]
- Wickstead, B.; Gull, K. Dyneins Across Eukaryotes: A Comparative Genomic Analysis. Traffic 2007, 8, 1708–1721. [Google Scholar] [CrossRef] [Green Version]
- Drinnenberg, I.A.; Akiyoshi, B. Evolutionary lessons from species with unique kinetochores. Cent. Kinetochores 2017, 56, 111–138. [Google Scholar]
- Van Hooff, J.J.; Tromer, E.; Van Wijk, L.M.; Snel, B.; Kops, G.J. Evolutionary dynamics of the kinetochore network in eukaryotes as revealed by comparative genomics. EMBO Rep. 2017, 18, 1559–1571. [Google Scholar] [CrossRef]
- Hamilton, G.E.; Davis, T.N. Biochemical evidence for diverse strategies in the inner kinetochore. Open Biol. 2020, 10, 200284. [Google Scholar] [CrossRef]
- Debec, A.; Sullivan, W.; Bettencourt-Dias, M. Centrioles: Active players or passengers during mitosis? Cell. Mol. Life Sci. 2010, 67, 2173–2194. [Google Scholar] [CrossRef] [Green Version]
- Sazer, S.; Lynch, M.; Needleman, D. Deciphering the Evolutionary History of Open and Closed Mitosis. Curr. Biol. 2014, 24, R1099–R1103. [Google Scholar] [CrossRef] [Green Version]
- Asbury, C.L. Anaphase A: Disassembling Microtubules Move Chromosomes toward Spindle Poles. Biology 2017, 6, 15. [Google Scholar] [CrossRef] [Green Version]
- Scholey, J.M.; Civelekoglu-Scholey, G.; Brust-Mascher, I. Anaphase b. Biology 2016, 5, 51. [Google Scholar] [CrossRef] [Green Version]
- Walczak, C.E.; Vernos, I.; Mitchison, T.J.; Karsenti, E.; Heald, R. A model for the proposed roles of different microtubule-based motor proteins in establishing spindle bipolarity. Curr. Biol. 1998, 8, 903–913. [Google Scholar] [CrossRef] [Green Version]
- Goshima, G.; Nédélec, F.; Vale, R.D. Mechanisms for focusing mitotic spindle poles by minus end–directed motor proteins. J. Cell Biol. 2005, 171, 229–240. [Google Scholar] [CrossRef] [Green Version]
- True, J.R.; Haag, E.S. Developmental system drift and flexibility in evolutionary trajectories. Evol. Dev. 2001, 3, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Nicklas, R.B. Measurements of the force produced by the mitotic spindle in anaphase. J. Cell Biol. 1983, 97, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Nicklas, R.B. The forces that move chromosomes in mitosis. Ann. Rev. Biophys. Chem. 1988, 17, 431–449. [Google Scholar] [CrossRef]
- Walczak, C.E.; Cai, S.; Khodjakov, A. Mechanisms of chromosome behaviour during mitosis. Nat. Rev. Mol. Cell Biol. 2010, 11, 91–102. [Google Scholar] [CrossRef]
- Musacchio, A.; Desai, A. A Molecular View of Kinetochore Assembly and Function. Biology 2017, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Rieder, C.L. The Formation, Structure, and Composition of the Mammalian Kinetochore and Kinetochore Fiber. Adv. in Applied Microbiology 1982, 79, 1–58. [Google Scholar] [CrossRef]
- McDonald, K.L.; O’Toole, E.T.; Mastronarde, D.N.; McIntosh, J.R. Kinetochore microtubules in PTK cells. J. Cell Biol. 1992, 118, 369–383. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.F.; Heagle, A.B.; Cassels, G.O.; Buttle, K.F.; Rieder, C.L. Kinetochore fiber maturation in PtK1 cells and its implications for the mechanisms of chromosome congression and anaphase onset. J. Cell Biol. 1997, 137, 1567–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Toole, E.; Morphew, M.; McIntosh, J.R. Electron tomography reveals aspects of spindle structure important for mechanical stability at metaphase. Mol. Biol. Cell 2020, 31, 184–195. [Google Scholar] [CrossRef]
- Euteneuer, U.; McIntosh, J.R. Structural polarity of kinetochore microtubules in PtK1 cells. J. Cell Biol. 1981, 89, 338–345. [Google Scholar] [CrossRef] [Green Version]
- Redemann, S.; Baumgart, J.; Lindow, N.; Shelley, M.; Nazockdast, E.; Kratz, A.; Prohaska, S.; Brugués, J.; Fürthauer, S.; Müller-Reichert, T. C. elegans chromosomes connect to centrosomes by anchoring into the spindle network. Nat. Commun. 2017, 8, 15288. [Google Scholar] [CrossRef] [Green Version]
- LaFountain, J.R., Jr.; Cohan, C.S.; Siegel, A.S.; LaFountain, D.J. Direct visualization of microtubule flux during metaphase and anaphase in crane-fly spermatocytes. Mol. Biol. Cell 2004, 15, 5724–5732. [Google Scholar] [CrossRef]
- McIntosh, J.R. Assessing the Contributions of Motor Enzymes and Microtubule Dynamics to Mitotic Chromosome Motions. Annu. Rev. Cell Dev. Biol. 2017, 33, 1–22. [Google Scholar] [CrossRef]
- Akiyoshi, B.; Sarangapani, K.K.; Powers, A.F.; Nelson, C.R.; Reichow, S.L.; Arellano-Santoyo, H.; Gonen, T.; Ranish, J.A.; Asbury, C.L.; Biggins, S. Tension directly stabilizes reconstituted kinetochore-microtubule attachments. Nat. Cell Biol. 2010, 468, 576–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asbury, C.L.; Tien, J.F.; Davis, T.N. Kinetochores’ gripping feat: Conformational wave or biased diffusion? Trends Cell Biol. 2011, 21, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Armond, J.W.; Turner, M.S. Force Transduction by the Microtubule-Bound Dam1 Ring. Biophys. J. 2010, 98, 1598–1607. [Google Scholar] [CrossRef] [Green Version]
- Grishchuk, E.L. Biophysics of microtubule end coupling at the kinetochore. Cent. Kinetochores 2017, 56, 397–428. [Google Scholar]
- Hill, T.L. Theoretical problems related to the attachment of microtubules to kinetochores. Proc. Natl. Acad. Sci. USA 1985, 82, 4404–4408. [Google Scholar] [CrossRef] [Green Version]
- Rieder, C.L. Formation of the astral mitotic spindle: Ultrastructural basis for the centrosome-kinetochore interaction. Electron Microsc. Rev. 1990, 3, 269–300. [Google Scholar] [CrossRef]
- Doxsey, S. Re-evaluating centrosome function. Nat. Rev. Mol. Cell Biol. 2001, 2, 688–698. [Google Scholar] [CrossRef]
- McNally, F.J. Mechanisms of spindle positioning. J. Cell Biol. 2013, 200, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Howard, J.; Garzon-Coral, C. Physical limits on the precision of mitotic spindle positioning by microtubule pushing forces: Mechanics of mitotic spindle positioning. BioEssays 2017, 39, 1700122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Minc, N. Cytoskeleton Force Exertion in Bulk Cytoplasm. Front. Cell Dev. Biol. 2020, 8, 69. [Google Scholar] [CrossRef]
- Wu, H.-Y.; Nazockdast, E.; Shelley, M.J.; Needleman, D.J. Forces positioning the mitotic spindle: Theories, and now experiments. BioEssays 2016, 39, 1600212. [Google Scholar] [CrossRef]
- Farhadifar, R.; Yu, C.-H.; Fabig, G.; Wu, H.-Y.; Stein, D.B.; Rockman, M.; Müller-Reichert, T.; Shelley, M.J.; Needleman, D.J. Stoichiometric interactions explain spindle dynamics and scaling across 100 million years of nematode evolution. eLife 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Garzon-Coral, C.; Fantana, H.A.; Howard, J. A force-generating machinery maintains the spindle at the cell center during mitosis. Science. 2016, 352, 1124–1127. [Google Scholar] [CrossRef]
- Grill, S.W.; Gönczy, P.; Stelzer, E.H.K.; Hyman, A.A. Polarity controls forces governing asymmetric spindle positioning in the Caenorhabditis elegans embryo. Nat. Cell Biol. 2001, 409, 630–633. [Google Scholar] [CrossRef]
- Grill, S.W.; Howard, J.; Schäffer, E.; Stelzer, E.H.K.; Hyman, A.A. The Distribution of Active Force Generators Controls Mitotic Spindle Position. Science 2003, 301, 518–521. [Google Scholar] [CrossRef] [Green Version]
- Winey, M.; Mamay, C.L.; O’Toole, E.T.; Mastronarde, D.N.; Giddings, T.H.; McDonald, K.L.; McIntosh, J.R. Three-dimensional ultrastructural analysis of the Saccharomyces cerevisiae mitotic spindle. J. Cell Biol. 1995, 129, 1601–1615. [Google Scholar] [CrossRef] [Green Version]
- Ding, R.; McDonald, K.L.; McIntosh, J.R. Three-dimensional reconstruction and analysis of mitotic spindles from the yeast, Schizosaccharomyces pombe. J. Cell Biol. 1993, 120, 141–151. [Google Scholar] [CrossRef]
- Mastronarde, D.N.; McDonald, K.L.; Ding, R.; McIntosh, J.R. Interpolar spindle microtubules in PTK cells. J. Cell Biol. 1993, 123, 1475–1489. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.-H.; Redemann, S.; Wu, H.-Y.; Kiewisz, R.; Yoo, T.Y.; Conway, W.; Farhadifar, R.; Müller-Reichert, T.; Needleman, D. Central-spindle microtubules are strongly coupled to chromosomes during both anaphase A and anaphase B. Mol. Biol. Cell 2019, 30, 2503–2514. [Google Scholar] [CrossRef]
- Lim, H.W.G.; Huber, G.; Torii, Y.; Hirata, A.; Miller, J.; Sazer, S. Vesicle-like biomechanics governs important aspects of nuclear geometry in fission yeast. PLoS ONE 2007, 2, e948. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.J.; Roque, H.; Antony, C.; Nedelec, F.J. Mechanical design principles of a mitotic spindle. eLife 2014, 3, 03398. [Google Scholar] [CrossRef] [PubMed]
- Lanksy, Z.; Braun, M.; Lüdecke, A.; Schlierf, M.; ten Wolde, P.R.; Janson, M.E.; Diez, S. Diffusible crosslinkers generate directed forces in microtubule networks. Cell 2015, 6, 1159–1168. [Google Scholar]
- Gaska, I.; Armstrong, M.E.; Alfieri, A.; Forth, S. The mitotic crosslinking protein PRC1 acts like a mechanical dashpot to resist microtubule sliding. Dev. Cell 2020, 3, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Lera-Ramirez, M.; Nédélec, F.J. Theory of antiparallel microtubule overlap stabilization by motors and diffusible cross-linkers. Cytoskeleton 2019, 76, 600–610. [Google Scholar] [CrossRef]
- Hannabuss, J.; Lera-Ramirez, M.; Cade, N.I.; Fourniol, F.J.; Nédélec, F.; Surrey, T. Self-Organization of Minimal Anaphase Spindle Midzone Bundles. Curr. Biol. 2019, 29, 2120–2130.e7. [Google Scholar] [CrossRef] [Green Version]
- Wijeratne, S.; Subramanian, R. Geometry of antiparallel microtubule bundles regulates relative sliding and stalling by PRC1 and Kif4A. eLife 2018, 7. [Google Scholar] [CrossRef]
- Purcell, E.M. Life at low Reynolds number. Am. J. Phys. 1977, 45, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Needleman, D.; Shelley, M. The stormy fluid dynamics of the living cell. Phys. Today 2019, 72, 32–38. [Google Scholar] [CrossRef]
- Berg, H.C.; Dyson, F. Random Walks in Biology. Phys. Today 1987, 40, 73–74. [Google Scholar] [CrossRef]
- Nicklas, R.B. Chromosome velocity during mitosis as a function of chromosome size and position. J. Cell Biol. 1965, 25, 119–135. [Google Scholar] [CrossRef] [Green Version]
- Fürthauer, S.; Lemma, B.; Foster, P.J.; Ems-McClung, S.C.; Yu, C.-H.; Walczak, C.E.; Dogic, Z.; Needleman, D.J.; Shelley, M.J. Self-straining of actively crosslinked microtubule networks. Nat. Phys. 2019, 15, 1295–1300. [Google Scholar] [CrossRef]
- Striebel, M.; Graf, I.R.; Frey, E. A Mechanistic View of Collective Filament Motion in Active Nematic Networks. Biophys. J. 2020, 118, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Ye, A.A.; Cane, S.; Maresca, T.J. Chromosome biorientation produces hundreds of piconewtons at a metazoan kinetochore. Nat. Commun. 2016, 7, 13221. [Google Scholar] [CrossRef] [Green Version]
- Salmon, E.D.; Bloom, K. Tension sensors reveal how the kinetochore shares its load. BioEssays 2017, 39, 1600216. [Google Scholar] [CrossRef]
- Suzuki, A.; Badger, B.L.; Haase, J.; Ohashi, T.; Erickson, H.P.; Salmon, E.D.; Bloom, K. How the kinetochore couples microtubule force and centromere stretch to move chromosomes. Nat. Cell Biol. 2016, 18, 382–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, J. Mechanics of Motor Proteins and the Cytoskeleton; Sinauer Associates: Sunderland, MA, USA, 2001; pp. 123–125. [Google Scholar]
- Israelachvili, J.N. Intermolecular and Surface Forces, 3rd ed.; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Koshland, D.E.; Mitchison, T.J.; Kirschner, M.W. Polewards chromosome movement driven by microtubule depolymerization in vitro. Nature 1988, 331, 499–504. [Google Scholar] [CrossRef]
- Coue, M.; Lombillo, V.A.; McIntosh, J.R. Microtubule depolymerization promotes particle and chromosome movement in vitro. J. Cell Biol. 1991, 112, 1165–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanimoto, H.; Sallé, J.; Dodin, L.; Minc, N. Physical forces determining the persistency and centring precision of microtubule asters. Nat. Phys. 2018, 14, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Elting, M.W.; Hueschen, C.L.; Udy, D.B.; Dumont, S. Force on spindle microtubule minus ends moves chromosomes. J. Cell Biol. 2014, 206, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Sikirzhytski, V.; Magidson, V.; Steinman, J.B.; He, J.; Le Berre, M.; Tikhonenko, I.; Ault, J.G.; McEwen, B.F.; Chen, J.K.; Sui, H.; et al. Direct kinetochore–spindle pole connections are not required for chromosome segregation. J. Cell Biol. 2014, 206, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Wollman, R.; Civelekoglu-Scholey, G.; Scholey, J.M.; Mogilner, A. Reverse engineering of force integration during mitosis in the Drosophila embryo. Mol. Syst. Biol. 2008, 4, 195. [Google Scholar] [CrossRef]
- Vukušić, K.; Buđa, R.; Tolić, I.M. Force-generating mechanisms of anaphase in human cells. J. Cell Sci. 2019, 132, jcs231985. [Google Scholar] [CrossRef] [Green Version]
- Suresh, P.; Long, A.F.; Dumont, S. Microneedle manipulation of the mammalian spindle reveals specialized, short-lived reinforcement near chromosomes. eLife 2020, 9, e53807. [Google Scholar] [CrossRef]
- Elting, M.W.; Prakash, M.; Udy, D.B.; Dumont, S. Mapping load-bearing in the mammalian spindle reveals local kinetochore-fiber anchorage that provides mechanical isolation and redundancy. Curr. Biol. 2017, 27, 2112–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vukušić, K.; Buđa, R.; Bosilj, A.; Milas, A.; Pavin, N.; Tolić, I.M. Microtubule sliding within the bridging fiber pushes kinetochore fibers apart to segregate chromosomes. Dev. Cell 2017, 43, 11–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kajtez, J.; Solomatina, A.; Novak, M.; Polak, B.; Vukušić, K.; Rüdiger, J.; Cojoc, G.; Milas, A.; Šestak, I.S.; Risteki, P.; et al. Overlap microtubules link sister k-fibres and balance the forces on bi-oriented kinetochores. Nat. Comm. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Brust-Mascher, I.; Civelekoglu-Scholey, G.; Kwon, M.; Mogilner, A.; Scholey, J.M. Model for anaphase B: Role of three mitotic motors in a switch from poleward flux to spindle elongation. Proc. Natl. Acad. Sci. USA 2004, 101, 15938–15943. [Google Scholar] [CrossRef] [Green Version]
- Civelekoglu-Scholey, G.; Sharp, D.J.; Mogilner, A.; Scholey, J.M. Model of Chromosome Motility in Drosophila Embryos: Adaptation of a General Mechanism for Rapid Mitosis. Biophys. J. 2006, 90, 3966–3982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khodjakov, A.; La Terra, S.; Chang, F. Laser microsurgery in fission yeast; role of the mitotic spindle midzone in anaphase B. Curr. Biol. 2004, 14, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Gay, G.; Courtheoux, T.; Reyes, C.; Tournier, S.; Gachet, Y. A stochastic model of kinetochore-microtubule attachment accurately describes fission yeast chromosome segregation. J. Cell Biol. 2012, 196, 757–774. [Google Scholar] [CrossRef] [Green Version]
- Ris, H. The anaphase movement of chromosomes in the spermatocytes of the grasshopper. Biol. Bull. 1949, 96, 90–106. [Google Scholar] [CrossRef]
- Nicklas, R.B. The motor for poleward chromosome movement in anaphase is in or near the kinetochore. J. Cell Biol. 1989, 109, 2245–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nahaboo, W.; Zouak, M.; Askjaer, P.; Delattre, M. Chromatids segregate without centrosomes during Caenorhabditis elegans mitosis in a Ran and CLASP-dependent manner. Mol. Biol. Cell 2015, 26, 2020–2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spurck, T.P.; Stonington, O.G.; Snyder, J.A.; Pickett-Heaps, J.D.; Bajer, A.; Mole-Bajer, J. UV microbeam irradiations of the mitotic spindle. II. Spindle fiber dynamics and force production. J. Cell Biol. 1990, 111, 1505–1518. [Google Scholar] [CrossRef] [PubMed]
- Spurck, T.; Forer, A.; Pickett-Heaps, J. Ultraviolet microbeam irradiations of epithelial and spermatocyte spindles suggest that forces act on the kinetochore fibre and are not generated by its disassembly. Cell Motil. Cytoskelet. 1997, 36, 136–148. [Google Scholar] [CrossRef]
- Aist, J.R.; Berns, M.W. Mechanics of chromosome separation during mitosis in Fusarium (Fungi imperfecti): New evidence from ultrastructural and laser microbeam experiments. J. Cell Biol. 1981, 91, 446–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aist, J.R.; Liang, H.; Berns, M.W. Astral and spindle forces in PtK2 cells during anaphase B: A laser microbeam study. J. Cell Sci. 1993, 104, 1207–1216. [Google Scholar] [PubMed]
- Bayles, C.J.; Aist, J.R.; Berns, M.W. The mechanics of anaphase-B in a basidiomycete as revealed by laser microbeam micro-surgery. Exp. Mycol. 1993, 17, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Dumont, J.; Oegema, K.; Desai, A. A kinetochore-independent mechanism drives anaphase chromosome separation during acentrosomal meiosis. Nat. Cell Biol. 2010, 12, 894–901. [Google Scholar] [CrossRef] [Green Version]
- Danlasky, B.M.; Panzica, M.T.; McNally, K.P.; Vargas, E.; Bailey, C.; Li, W.; Gong, T.; Fishman, E.S.; Jiang, X.; McNally, F.J. Evidence for anaphase pulling forces during C. elegans meiosis. J. Cell Biol. 2020, 219, 202005179. [Google Scholar] [CrossRef]
- Laband, K.; Le Borgne, R.; Edwards, F.; Stefanutti, M.; Canman, J.C.; Verbavatz, J.-M.; Dumont, J. Chromosome segregation occurs by microtubule pushing in oocytes. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Gao, J.; Suraneni, P.; Li, R. Kinetochore-independent chromosome poleward movement during anaphase of mei-osis II in mouse eggs. PLoS ONE 2009, 4, e5249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warecki, B.; Sullivan, W. Mechanisms driving acentric chromosome transmission. Chromosom. Res. 2020, 28, 229–246. [Google Scholar] [CrossRef] [PubMed]
- Needleman, D.; Dogic, Z. Active matter at the interface between materials science and cell biology. Nat. Rev. Mater. 2017, 2, 17048. [Google Scholar] [CrossRef]
- Marchetti, M.C.; Joanny, J.F.; Ramaswamy, S.; Liverpool, T.B.; Prost, J.; Rao, M.; Simha, R.A. Hydrodynamics of soft active matter. Rev. Mod. Phys. 2013, 85, 1143–1189. [Google Scholar] [CrossRef] [Green Version]
- Brugués, J.; Needleman, D. Physical basis of spindle self-organization. Proc. Natl. Acad. Sci. USA 2014, 111, 18496–18500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oriola, D.; Needleman, D.J.; Brugués, J. The Physics of the Metaphase Spindle. Annu. Rev. Biophys. 2018, 47, 655–673. [Google Scholar] [CrossRef]
- Oriola, D.; Jülicher, F.; Brugués, J. Active forces shape the metaphase spindle through a mechanical instability. Proc. Natl. Acad. Sci. USA 2020, 117, 16154–16159. [Google Scholar] [CrossRef]
- Takagi, J.; Sakamoto, R.; Shiratsuchi, G.; Maeda, Y.T.; Shimamoto, Y. Mechanically Distinct Microtubule Arrays Determine the Length and Force Response of the Meiotic Spindle. Dev. Cell 2019, 49, 267–278.e5. [Google Scholar] [CrossRef] [PubMed]
- Shimamoto, Y.; Maeda, Y.T.; Ishiwata, S.; Libchaber, A.J.; Kapoor, T.M. Insights into the Micromechanical Properties of the Metaphase Spindle. Cell 2011, 145, 1062–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, W.F.; Marko, J.F.; Agard, D.A.; Sedat, J.W. Chromosome elasticity and mitotic polar ejection force measured in living Drosophila embryos by four-dimensional microscopy-based motion analysis. Curr. Biol. 2001, 11, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.; Peskin, C.S. The influence of chromosome flexibility on chromosome transport during anaphase A. Proc. Natl. Acad. Sci. USA 2006, 103, 5349–5354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houchmandzadeh, B.; Marko, J.F.; Chatenay, D.; Libchaber, A. Elasticity and Structure of Eukaryote Chromosomes Studied by Micromanipulation and Micropipette Aspiration. J. Cell Biol. 1997, 139, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Marko, J.F. Micromechanical studies of mitotic chromosomes. Chromosom. Res. 2008, 16, 469–497. [Google Scholar] [CrossRef] [Green Version]
- Stephens, A.D.; Haggerty, R.A.; Vasquez, P.A.; Vicci, L.; Snider, C.E.; Shi, F.; Quammen, C.; Mullins, C.; Haase, J.; Taylor, R.M., III; et al. Pericentric chromatin loops function as a nonlinear spring in mitotic force balance. J. Cell Biol. 2013, 200, 757–772. [Google Scholar] [CrossRef] [Green Version]
- Foster, P.J.; Fürthauer, S.; Shelley, M.J.; Needleman, D.J. From cytoskeletal assemblies to living materials. Curr. Opin. Cell Biol. 2019, 56, 109–114. [Google Scholar] [CrossRef] [PubMed]
- Fürthauer, S.; Needleman, D.J.; Shelley, M.J. A design framework for actively crosslinked filament networks. New J. Phys. 2020, 23, 013012. [Google Scholar] [CrossRef]
- Marshall, W.F. Centriole evolution. Curr. Opin. Cell Biol. 2009, 21, 14–19. [Google Scholar] [CrossRef]
- Carvalho-Santos, Z.; Azimzadeh, J.; Pereira-Leal, J.B.; Bettencourt-Dias, M. Tracing the origins of centrioles, cilia, and flagella. J. Cell Biol. 2011, 194, 165–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, M. The frailty of adaptive hypotheses for the origins of organismal complexity. Proc. Natl. Acad. Sci. USA 2007, 104, 8597–8604. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.W.; Lukes, J.; Archibald, J.M.; Keeling, P.J.; Doolittle, W.F. Irremediable Complexity? Science 2010, 330, 920–921. [Google Scholar] [CrossRef]
- Lukeš, J.; Archibald, J.M.; Keeling, P.J.; Doolittle, W.F.; Gray, M.W. How a neutral evolutionary ratchet can build cellular complexity. IUBMB Life 2011, 63, 528–537. [Google Scholar] [CrossRef]
- Lynch, M.; Walsh, B. The Origins of Genome Architecture; Sinauer Associates: Sunderland, MA, USA, 2007. [Google Scholar]
- Valfort, A.C.; Launay, C.; Sémon, M.; Delattre, M. Evolution of mitotic spindle behavior during the first asymmetric embryonic division of nematodes. PLoS Biol. 2018, 16, e2005099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, W.H. Elements of Evolutionary Genetics, 1st ed.; Roberts Publishers: Greenwood Village, CO, USA, 2010. [Google Scholar]
- Walsh, B.; Lynch, M. Evolution and Selection of Quantitative Traits; Oxford University Press (OUP): Oxford, UK, 2018. [Google Scholar]
- Lynch, M.; Field, M.C.; Goodson, H.V.; Malik, H.S.; Pereira-Leal, J.B.; Roos, D.S.; Turkewitz, A.P.; Sazer, S. Evolutionary cell biology: Two origins, one objective: Figure 1. Proc. Natl. Acad. Sci. USA 2014, 111, 16990–16994. [Google Scholar] [CrossRef] [Green Version]
- Lynch, M.; Trickovic, B. A Theoretical Framework for Evolutionary Cell Biology. J. Mol. Biol. 2020, 432, 1861–1879. [Google Scholar] [CrossRef] [PubMed]
- Drinnenberg, I.A.; Berger, F.; Elsässer, S.J.; Andersen, P.R.; Ausió, J.; Bickmore, W.A.; Blackwell, A.R.; Erwin, U.H.; Gahan, J.M.; Gaut, B.S.; et al. EvoChromo: Towards a synthesis of chromatin biology and evolution. Development 2019, 146, dev178962. [Google Scholar] [CrossRef] [Green Version]
- Malik, H.S. The Centromere-Drive Hypothesis: A Simple Basis for Centromere Complexity. Centromere 2009, 48, 33–52. [Google Scholar] [CrossRef]
- Kursel, L.E.; Malik, H.S. The cellular mechanisms and consequences of centromere drive. Curr. Opin. Cell Biol. 2018, 52, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Farhadifar, R.; Ponciano, J.M.; Andersen, E.C.; Needleman, D.J.; Baer, C.F. Mutation Is a Sufficient and Robust Predictor of Genetic Variation for Mitotic Spindle Traits in Caenorhabditis elegans. Genetics 2016, 203, 1859–1870. [Google Scholar] [CrossRef] [Green Version]
- Farhadifar, R.; Baer, C.F.; Valfort, A.-C.; Andersen, E.C.; Müller-Reichert, T.; Delattre, M.; Needleman, D.J. Scaling, Selection, and Evolutionary Dynamics of the Mitotic Spindle. Curr. Biol. 2015, 25, 732–740. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anjur-Dietrich, M.I.; Kelleher, C.P.; Needleman, D.J. Mechanical Mechanisms of Chromosome Segregation. Cells 2021, 10, 465. https://doi.org/10.3390/cells10020465
Anjur-Dietrich MI, Kelleher CP, Needleman DJ. Mechanical Mechanisms of Chromosome Segregation. Cells. 2021; 10(2):465. https://doi.org/10.3390/cells10020465
Chicago/Turabian StyleAnjur-Dietrich, Maya I., Colm P. Kelleher, and Daniel J. Needleman. 2021. "Mechanical Mechanisms of Chromosome Segregation" Cells 10, no. 2: 465. https://doi.org/10.3390/cells10020465
APA StyleAnjur-Dietrich, M. I., Kelleher, C. P., & Needleman, D. J. (2021). Mechanical Mechanisms of Chromosome Segregation. Cells, 10(2), 465. https://doi.org/10.3390/cells10020465