
Pattern of Tumor-Infiltrating Lymphocytes in Mixed Epithelial and Stromal Tumor of the Kidney: A Review of Five Cases

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Immunohistochemistry
2.3. Flow Cytometric Analysis
3. Results
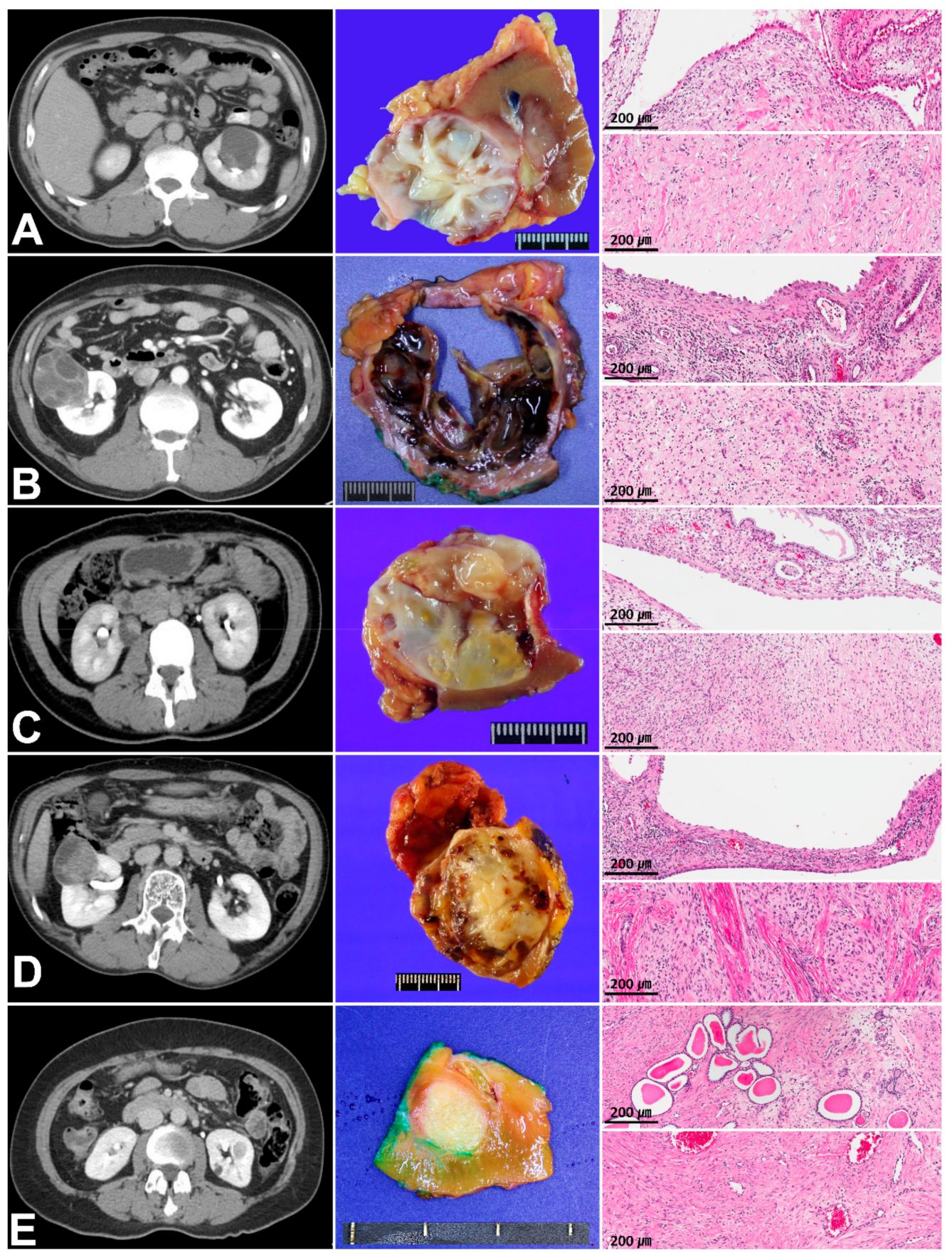
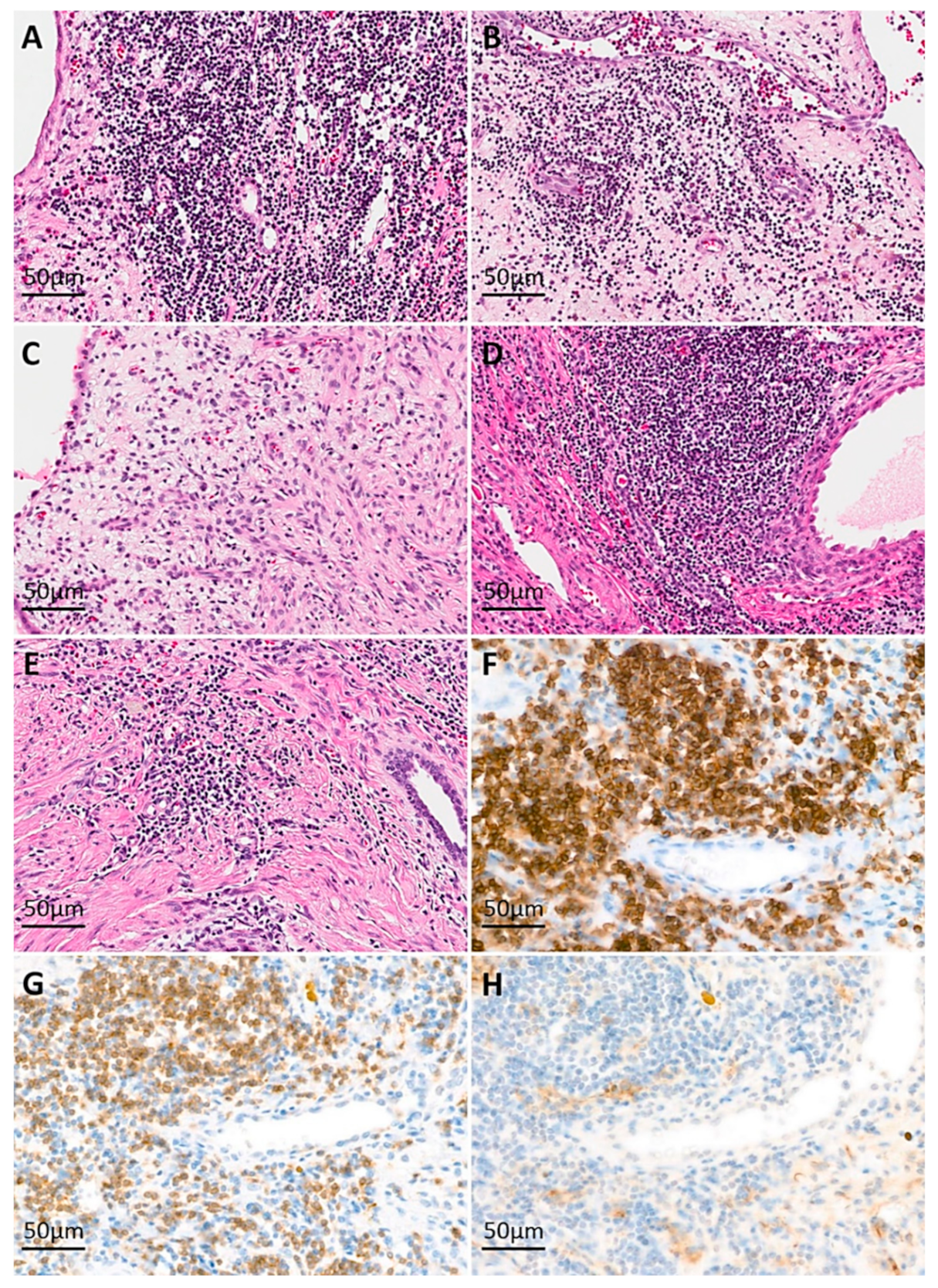
3.1. Clinicopathologic Characteristics and Immunoreactivity of the Five Cases of MESTK
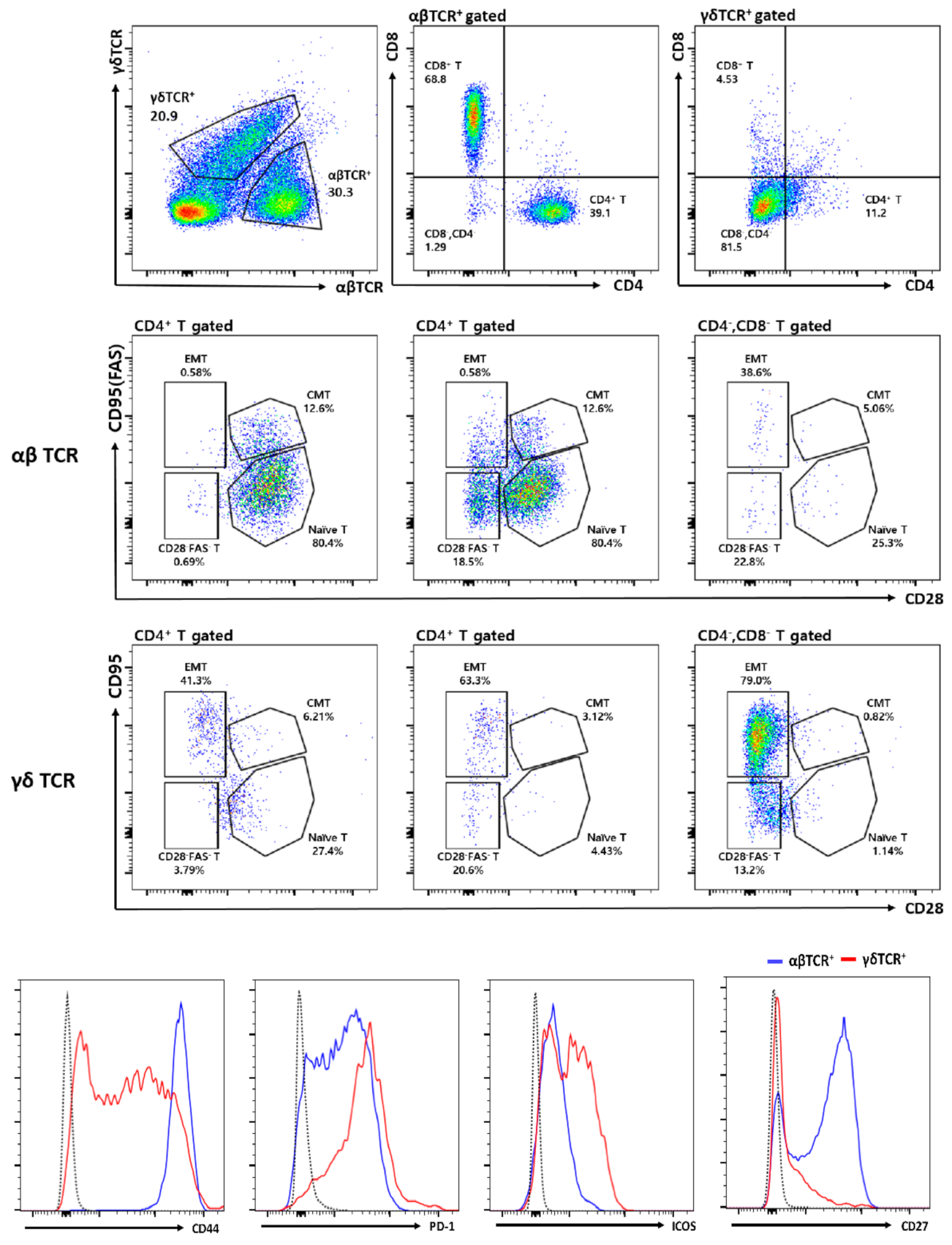
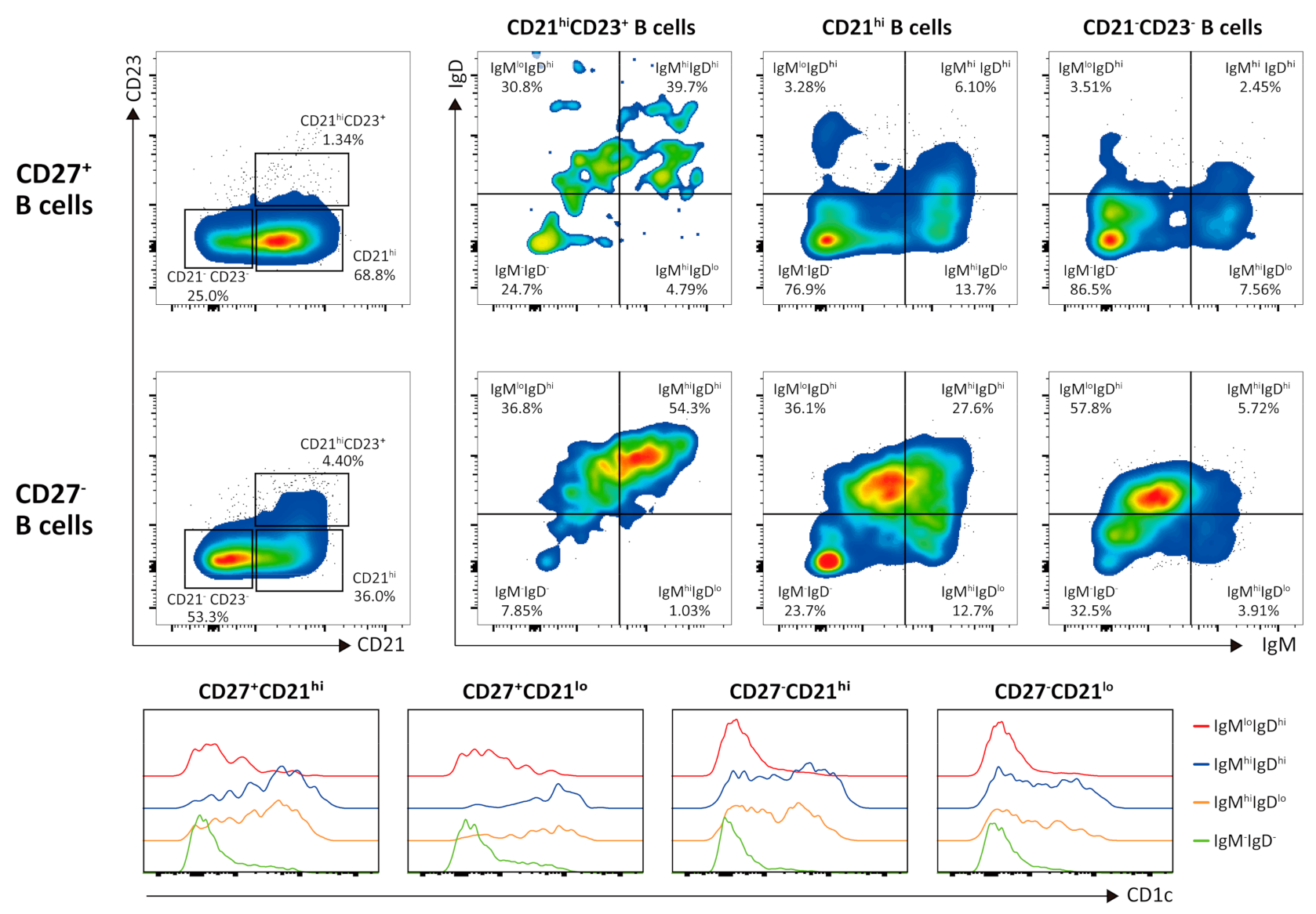
3.2. Flow Cytometric Analysis of Intratumor Lymphocytes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Montironi, R.; Mazzucchelli, R.; Lopez-Beltran, A.; Martignoni, G.; Cheng, L.; Montorsi, F.; Scarpelli, M. Cystic nephroma and mixed epithelial and stromal tumour of the kidney: Opposite ends of the spectrum of the same entity? Eur. Urol. 2008, 54, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Lane, B.R.; Campbell, S.C.; Remer, E.M.; Fergany, A.F.; Williams, S.B.; Novick, A.C.; Weight, C.J.; Magi-Galluzzi, C.; Zhou, M. Adult cystic nephroma and mixed epithelial and stromal tumor of the kidney: Clinical, radiographic, and pathologic characteristics. Urology 2008, 71, 1142–1148. [Google Scholar] [CrossRef] [PubMed]
- Portier, B.P.; Hansel, D.E.; Zhou, M.; Maclennan, G.T. Mixed epithelial and stromal tumor of the kidney. J. Urol. 2009, 181, 1879–1880. [Google Scholar] [CrossRef] [PubMed]
- Calio, A.; Eble, J.N.; Grignon, D.J.; Delahunt, B. Mixed Epithelial and Stromal Tumor of the Kidney: A Clinicopathologic Study of 53 Cases. Am. J. Surg. Pathol. 2016, 40, 1538–1549. [Google Scholar] [CrossRef]
- Jung, S.J.; Shen, S.S.; Tran, T.; Jun, S.Y.; Truong, L.; Ayala, A.G.; Ro, J.Y. Mixed epithelial and stromal tumor of kidney with malignant transformation: Report of two cases and review of literature. Hum. Pathol. 2008, 39, 463–468. [Google Scholar] [CrossRef]
- Kuroda, N.; Sakaida, N.; Kinoshita, H.; Matsuda, T.; Hes, O.; Michal, M.; Okamoto, S.; Nagashima, Y.; Tanaka, Y. Carcinosarcoma arising in mixed epithelial and stromal tumor of the kidney. APMIS 2008, 116, 1013–1015. [Google Scholar] [CrossRef]
- Yap, Y.S.; Coleman, M.; Olver, I. Aggressive mixed epithelial-stromal tumour of the kidney treated with chemotherapy and radiotherapy. Lancet Oncol. 2004, 5, 747–749. [Google Scholar] [CrossRef]
- Kum, J.B.; Grignon, D.J.; Wang, M.; Zhou, M.; Montironi, R.; Shen, S.S.; Zhang, S.; Lopez-Beltran, A.; Eble, J.N.; Cheng, L. Mixed epithelial and stromal tumors of the kidney: Evidence for a single cell of origin with capacity for epithelial and stromal differentiation. Am. J. Surg. Pathol. 2011, 35, 1114–1122. [Google Scholar] [CrossRef]
- Beiko, D.T.; Nickel, J.C.; Boag, A.H.; Srigley, J.R. Benign mixed epithelial stromal tumor of the kidney of possible mullerian origin. J. Urol. 2001, 166, 1381–1382. [Google Scholar] [CrossRef]
- Binnewies, M.; Roberts, E.W.; Kersten, K.; Chan, V.; Fearon, D.F.; Merad, M.; Coussens, L.M.; Gabrilovich, D.I.; Ostrand-Rosenberg, S.; Hedrick, C.C.; et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat. Med. 2018, 24, 541–550. [Google Scholar] [CrossRef]
- Turbiner, J.; Amin, M.B.; Humphrey, P.A.; Srigley, J.R.; De Leval, L.; Radhakrishnan, A.; Oliva, E. Cystic nephroma and mixed epithelial and stromal tumor of kidney: A detailed clinicopathologic analysis of 34 cases and proposal for renal epithelial and stromal tumor (REST) as a unifying term. Am. J. Surg. Pathol. 2007, 31, 489–500. [Google Scholar] [CrossRef]
- Dieu-Nosjean, M.C.; Goc, J.; Giraldo, N.A.; Sautes-Fridman, C.; Fridman, W.H. Tertiary lymphoid structures in cancer and beyond. Trends. Immunol. 2014, 35, 571–580. [Google Scholar] [CrossRef]
- Mahnke, Y.D.; Brodie, T.M.; Sallusto, F.; Roederer, M.; Lugli, E. The who’s who of T-cell differentiation: Human memory T-cell subsets. Eur. J. Immunol. 2013, 43, 2797–2809. [Google Scholar] [CrossRef]
- Rubtsova, K.; Rubtsov, A.V.; Cancro, M.P.; Marrack, P. Age-Associated B Cells: A T-bet-Dependent Effector with Roles in Protective and Pathogenic Immunity. J. Immunol. 2015, 195, 1933–1937. [Google Scholar] [CrossRef]
- Kurts, C.; Panzer, U.; Anders, H.J.; Rees, A.J. The immune system and kidney disease: Basic concepts and clinical implications. Nat. Rev. Immunol. 2013, 13, 738–753. [Google Scholar] [CrossRef]
- Chevrier, S.; Levine, J.H.; Zanotelli, V.R.T.; Silina, K.; Schulz, D.; Bacac, M.; Ries, C.H.; Ailles, L.; Jewett, M.A.S.; Moch, H.; et al. An Immune Atlas of Clear Cell Renal Cell Carcinoma. Cell 2017, 169, 736–749.e18. [Google Scholar] [CrossRef] [Green Version]
- Boor, P.; Floege, J. The renal (myo-)fibroblast: A heterogeneous group of cells. Nephrol. Dial. Transplant. 2012, 27, 3027–3036. [Google Scholar] [CrossRef] [Green Version]
- Harper, J.; Sainson, R.C. Regulation of the anti-tumour immune response by cancer-associated fibroblasts. Semin. Cancer Biol. 2014, 25, 69–77. [Google Scholar] [CrossRef]
- Cunha, G.R.; Robboy, S.J.; Kurita, T.; Isaacson, D.; Shen, J.; Cao, M.; Baskin, L.S. Development of the human female reproductive tract. Differentiation 2018, 103, 46–65. [Google Scholar] [CrossRef] [Green Version]
- Tickoo, S.K.; Gopalan, A.; Tu, J.J.; Harik, L.R.; Al-Ahmadie, H.A.; Fine, S.W.; Olgac, S.; Reuter, V.E. Estrogen and progesterone-receptor-positive stroma as a non-tumorous proliferation in kidneys: A possible metaplastic response to obstruction. Mod. Pathol. 2008, 21, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Makuch, M.; Wilson, R.; Al-Diwani, A.; Varley, J.; Kienzler, A.K.; Taylor, J.; Berretta, A.; Fowler, D.; Lennox, B.; Leite, M.I.; et al. N-methyl-D-aspartate receptor antibody production from germinal center reactions: Therapeutic implications. Ann. Neurol. 2018, 83, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Lawand, M.; Dechanet-Merville, J.; Dieu-Nosjean, M.C. Key Features of Gamma-Delta T-Cell Subsets in Human Diseases and Their Immunotherapeutic Implications. Front. Immunol. 2017, 8, 761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauza, C.D.; Liou, M.L.; Lahusen, T.; Xiao, L.; Lapidus, R.G.; Cairo, C.; Li, H. Gamma Delta T Cell Therapy for Cancer: It Is Good to be Local. Front. Immunol. 2018, 9, 1305. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Niu, C.; Cui, J. Gamma-delta (gammadelta) T cells: Friend or foe in cancer development? J. Transl. Med. 2018, 16, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeres, T.; Smetak, M.; Pretscher, D.; Wilhelm, M. Improving the Efficiency of Vgamma9Vdelta2 T-Cell Immunotherapy in Cancer. Front. Immunol. 2018, 9, 800. [Google Scholar] [CrossRef] [PubMed]
- Gocze, I.; Ehehalt, K.; Zeman, F.; Riquelme, P.; Pfister, K.; Graf, B.M.; Bein, T.; Geissler, E.K.; Kasprzak, P.; Schlitt, H.J.; et al. Postoperative cellular stress in the kidney is associated with an early systemic gammadelta T-cell immune cell response. Crit. Care 2018, 22, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsou, P.; Katayama, H.; Ostrin, E.J.; Hanash, S.M. The Emerging Role of B Cells in Tumor Immunity. Cancer Res. 2016, 76, 5597–5601. [Google Scholar] [CrossRef] [Green Version]
- Montfort, A.; Pearce, O.; Maniati, E.; Vincent, B.G.; Bixby, L.; Bohm, S.; Dowe, T.; Wilkes, E.H.; Chakravarty, P.; Thompson, R.; et al. A Strong B-cell Response Is Part of the Immune Landscape in Human High-Grade Serous Ovarian Metastases. Clin. Cancer Res. 2017, 23, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Weill, J.C.; Weller, S.; Reynaud, C.A. Human marginal zone B cells. Annu. Rev. Immunol. 2009, 27, 267–285. [Google Scholar] [CrossRef]
- Hao, Y.; O’Neill, P.; Naradikian, M.S.; Scholz, J.L.; Cancro, M.P. A B-cell subset uniquely responsive to innate stimuli accumulates in aged mice. Blood 2011, 118, 1294–1304. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Number |
|---|---|
| Sex | |
| Male | 3 |
| Female | 2 |
| Age (years) | 51.4 (34–60) |
| History of Hormone therapy | |
| Absent | 4 |
| Present | 1 |
| Tumor size (mm) | 37.6 (18–55) |
| Recurrence | |
| Absent | 5 |
| Survival status | |
| Alive | 5 |
| Follow-up period (months) | 9.3 (1.1–27.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.W.; Lee, H.; Park, C.; Oh, W.J.; Kim, T.J.; Kwon, G.Y.; Seo, S.I. Pattern of Tumor-Infiltrating Lymphocytes in Mixed Epithelial and Stromal Tumor of the Kidney: A Review of Five Cases. Cells 2021, 10, 917. https://doi.org/10.3390/cells10040917
Lee HW, Lee H, Park C, Oh WJ, Kim TJ, Kwon GY, Seo SI. Pattern of Tumor-Infiltrating Lymphocytes in Mixed Epithelial and Stromal Tumor of the Kidney: A Review of Five Cases. Cells. 2021; 10(4):917. https://doi.org/10.3390/cells10040917
Chicago/Turabian StyleLee, Hye Won, Hyunwoo Lee, Chanho Park, Won Joon Oh, Tae Jin Kim, Ghee Young Kwon, and Seong Il Seo. 2021. "Pattern of Tumor-Infiltrating Lymphocytes in Mixed Epithelial and Stromal Tumor of the Kidney: A Review of Five Cases" Cells 10, no. 4: 917. https://doi.org/10.3390/cells10040917
APA StyleLee, H. W., Lee, H., Park, C., Oh, W. J., Kim, T. J., Kwon, G. Y., & Seo, S. I. (2021). Pattern of Tumor-Infiltrating Lymphocytes in Mixed Epithelial and Stromal Tumor of the Kidney: A Review of Five Cases. Cells, 10(4), 917. https://doi.org/10.3390/cells10040917