
Upregulation of Chemoresistance by Mg2+ Deficiency through Elevation of ATP Binding Cassette Subfamily B Member 1 Expression in Human Lung Adenocarcinoma A549 Cells


Abstract
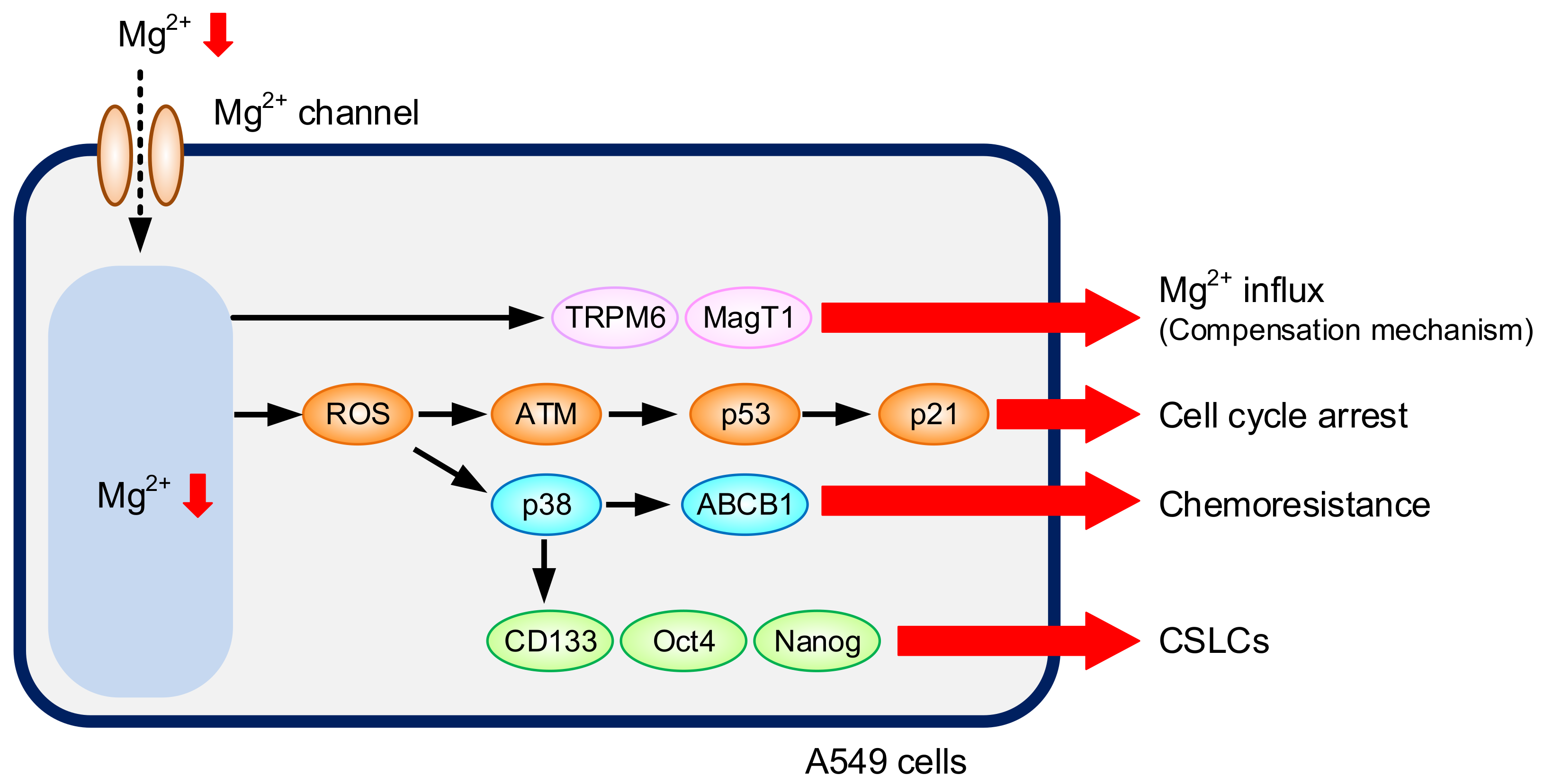
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Measurement of [Mg2+]i
2.3. Reverse Transcription and Quantitative Real-Time Polymerase Chain Reaction (PCR)
2.4. Flow Cytometry
2.5. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blotting
2.6. Oxidative DNA Damage and ROS Production
2.7. Statistical Analysis
3. Results
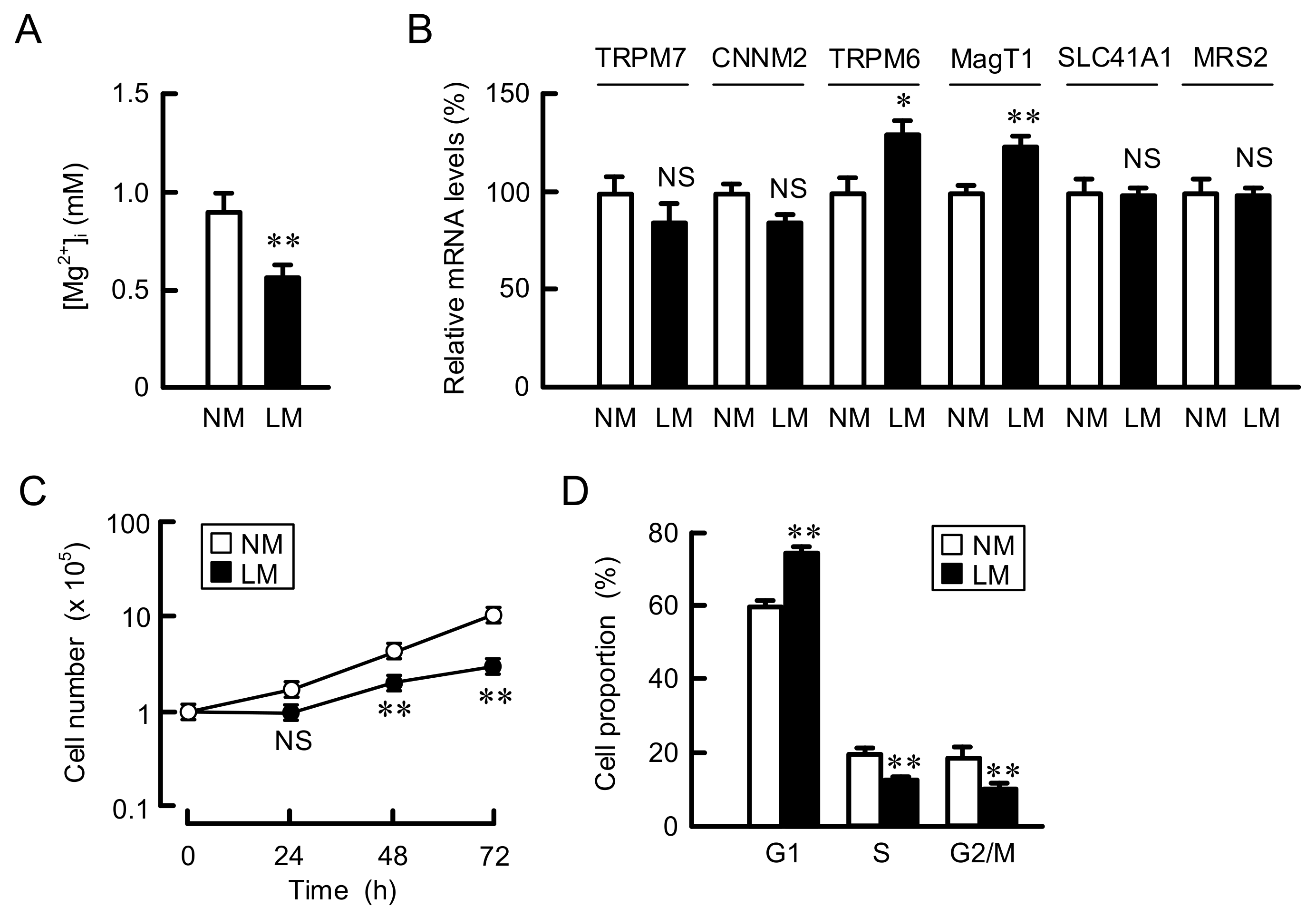
3.1. Inhibition of Cell Proliferation by Culturing in LM Medium
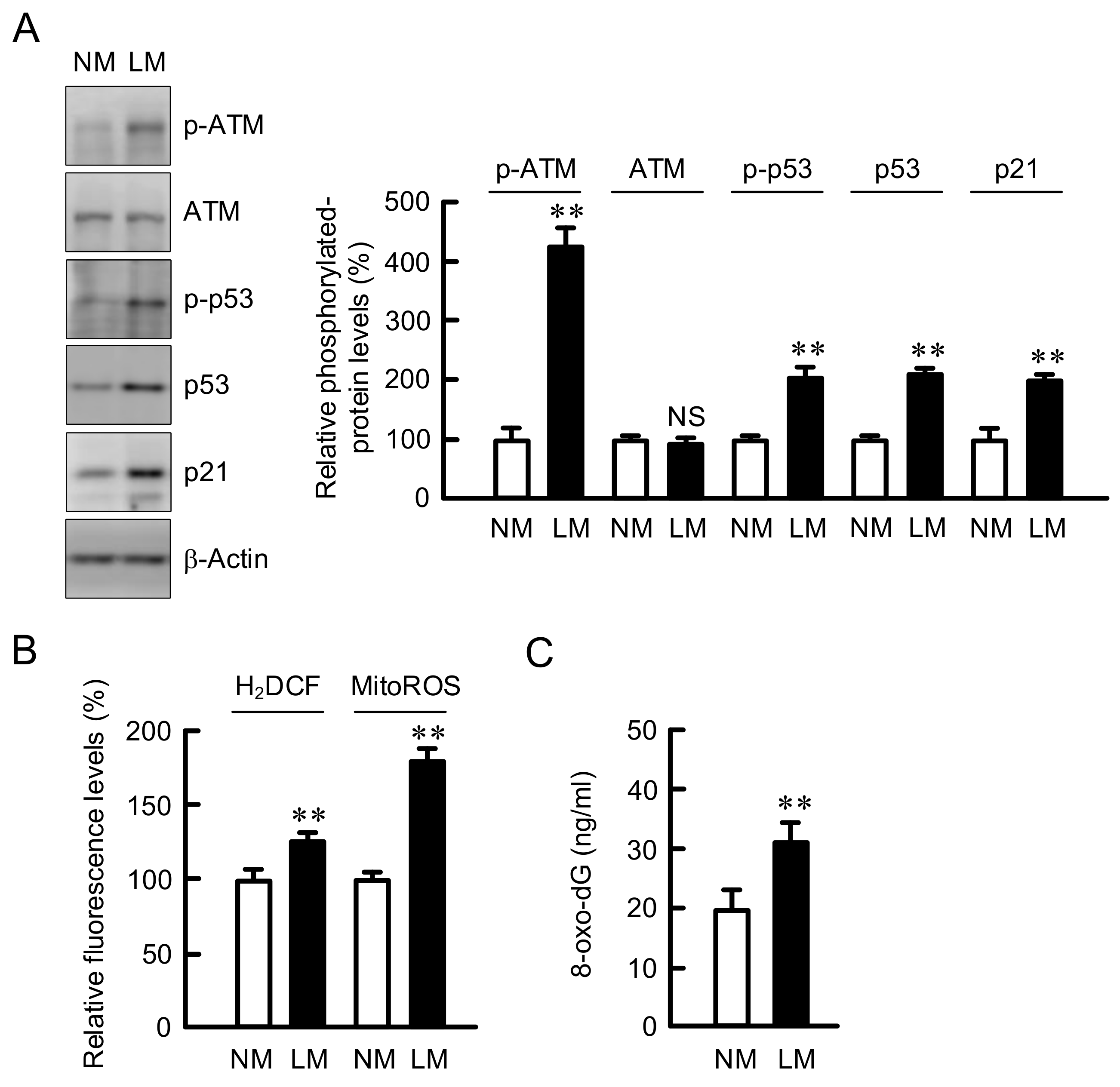
3.2. Increase in the Expression of Negative Cell Cycle Regulators by LM
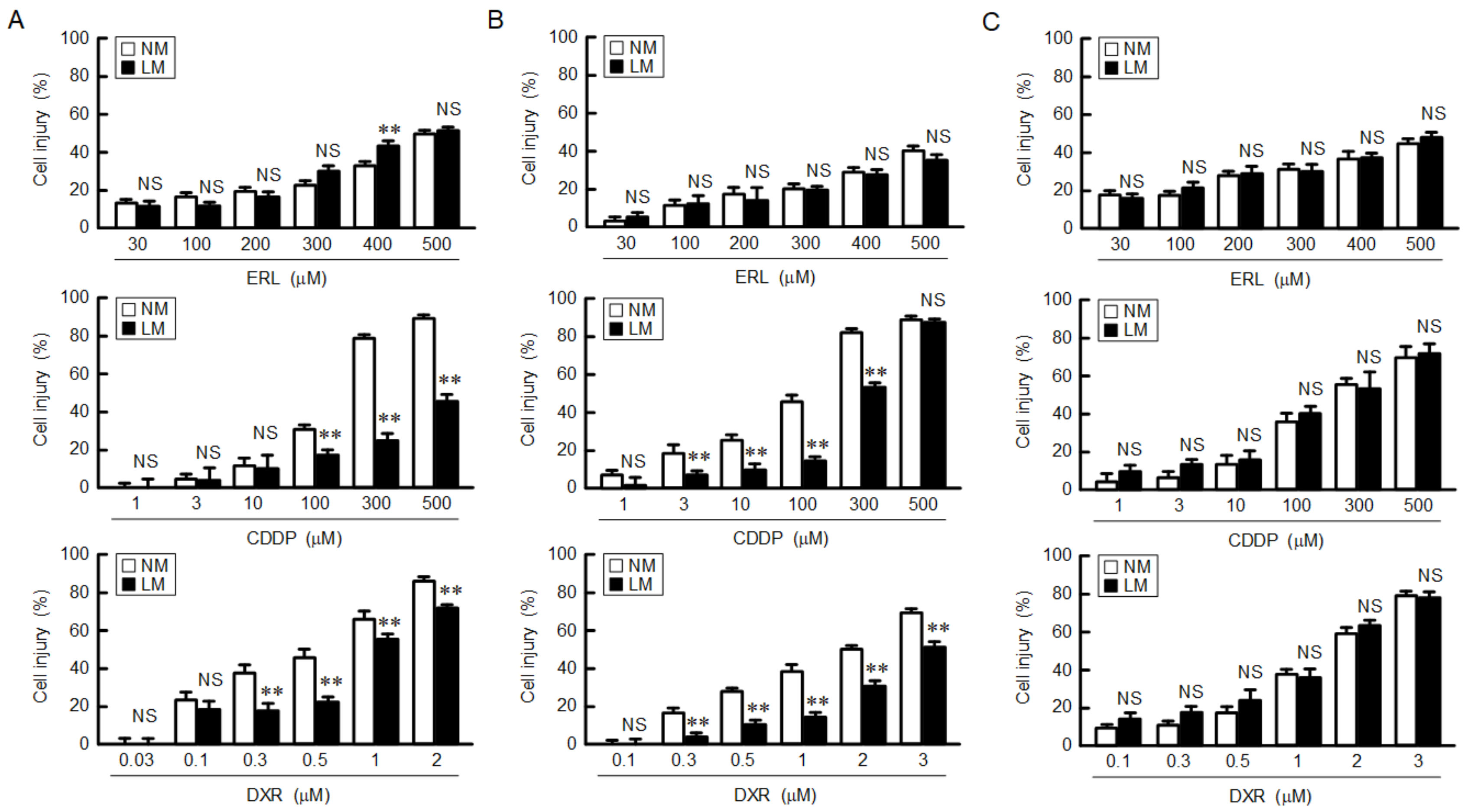
3.3. Decrease in Chemosensitivity of Anticancer Drugs by LM
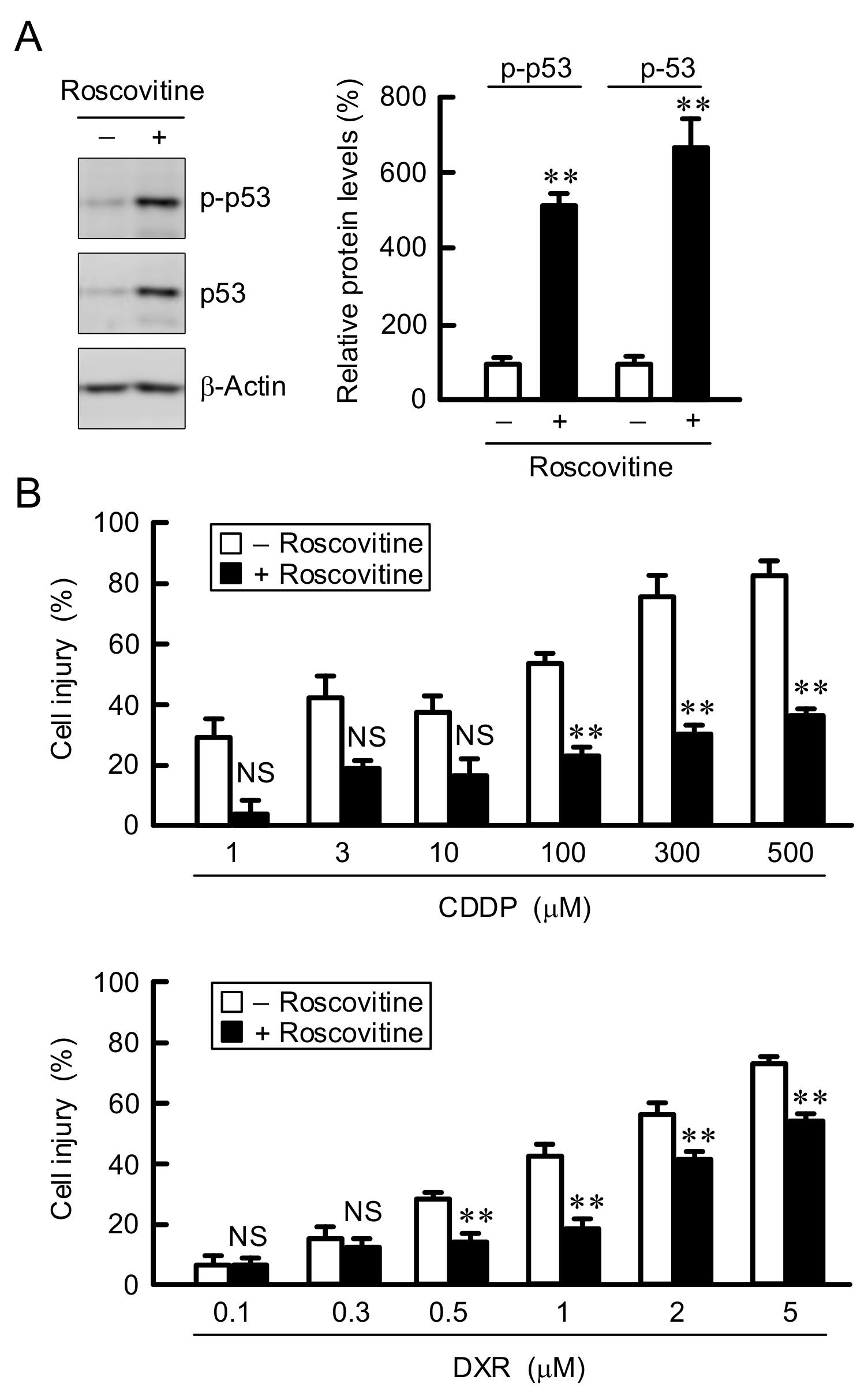
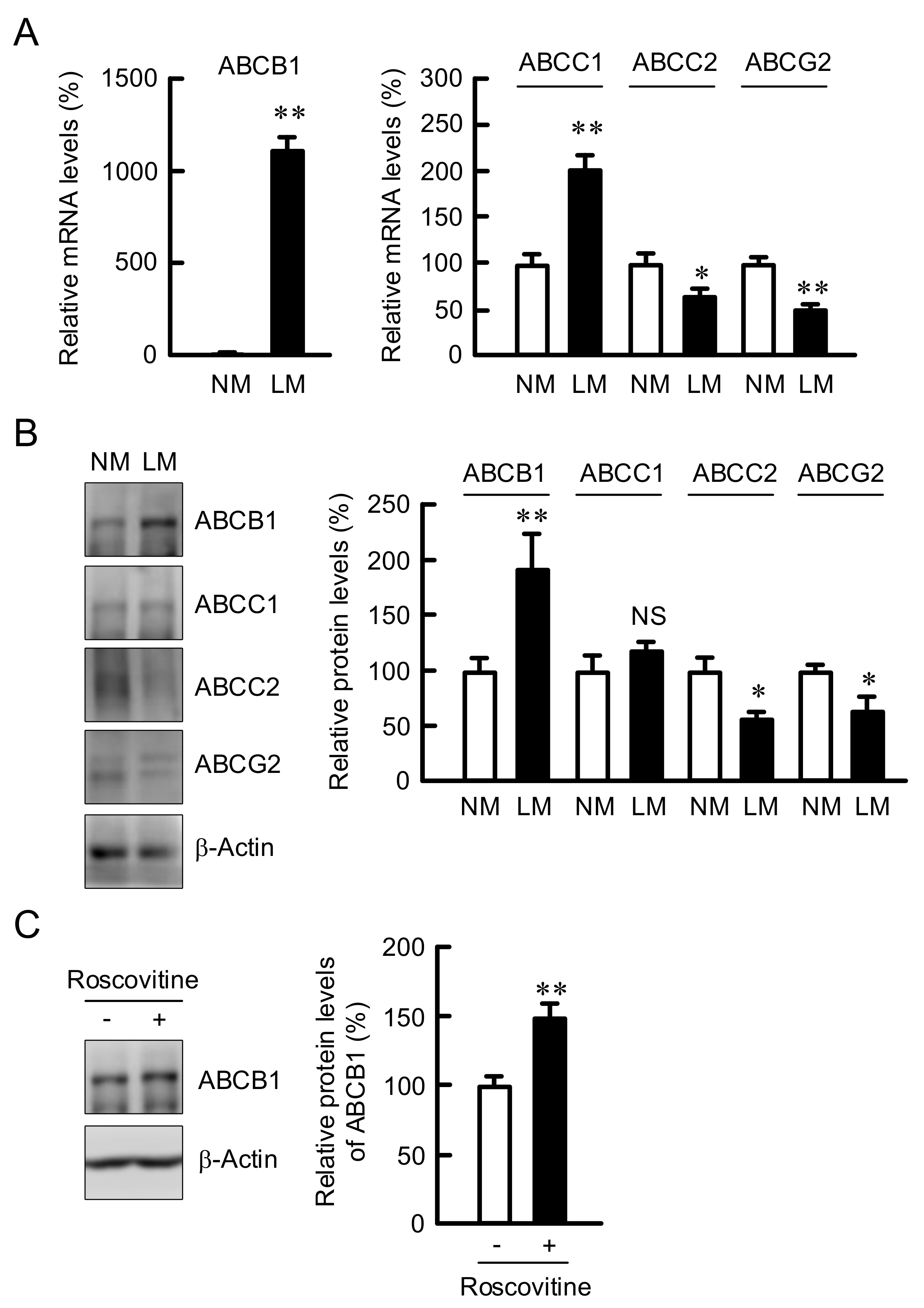
3.4. Elevation of ABCB1 Expression by LM and Roscovitine
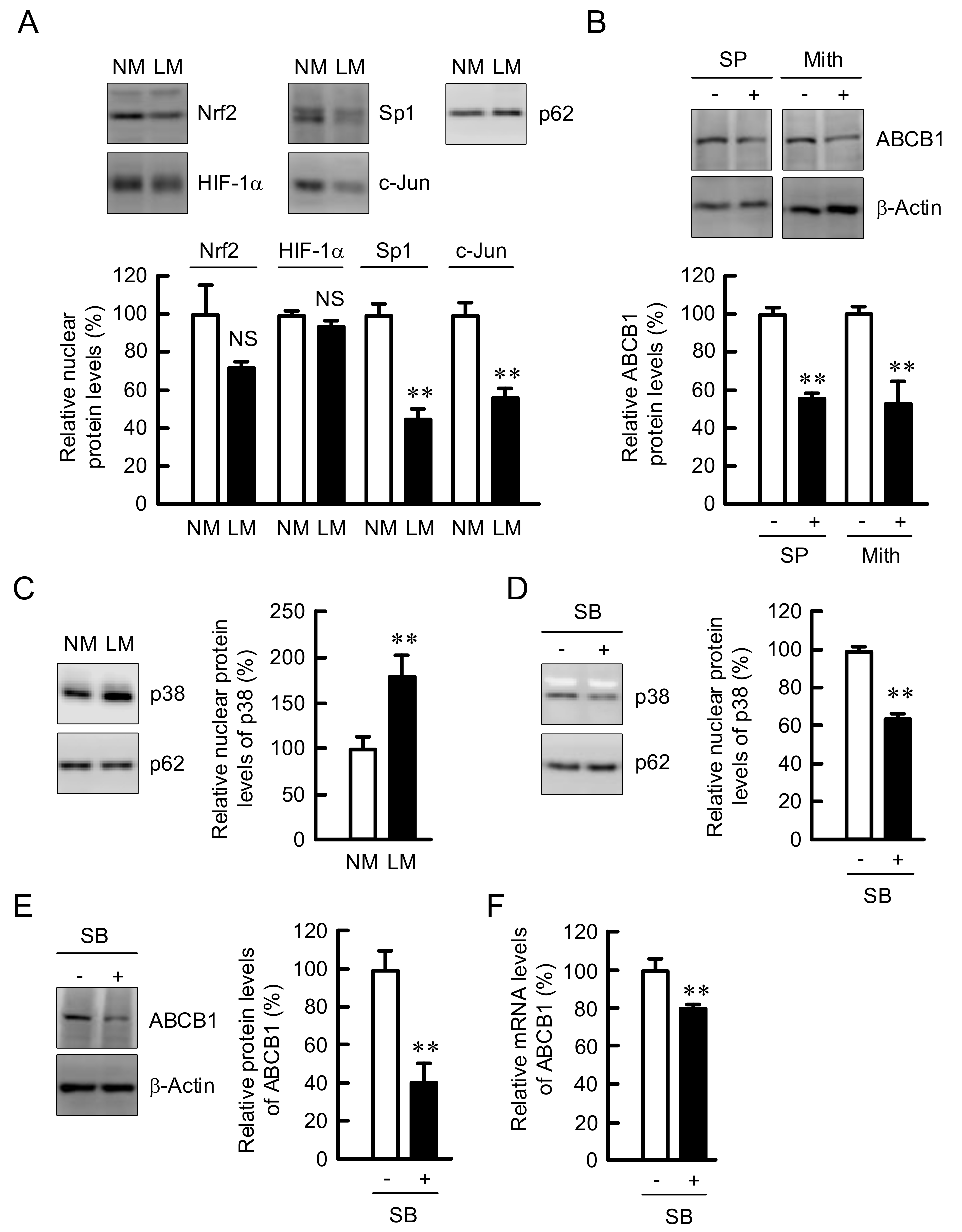
3.5. Involvement of p38 in the Elevation of ABCB1 Expression by LM
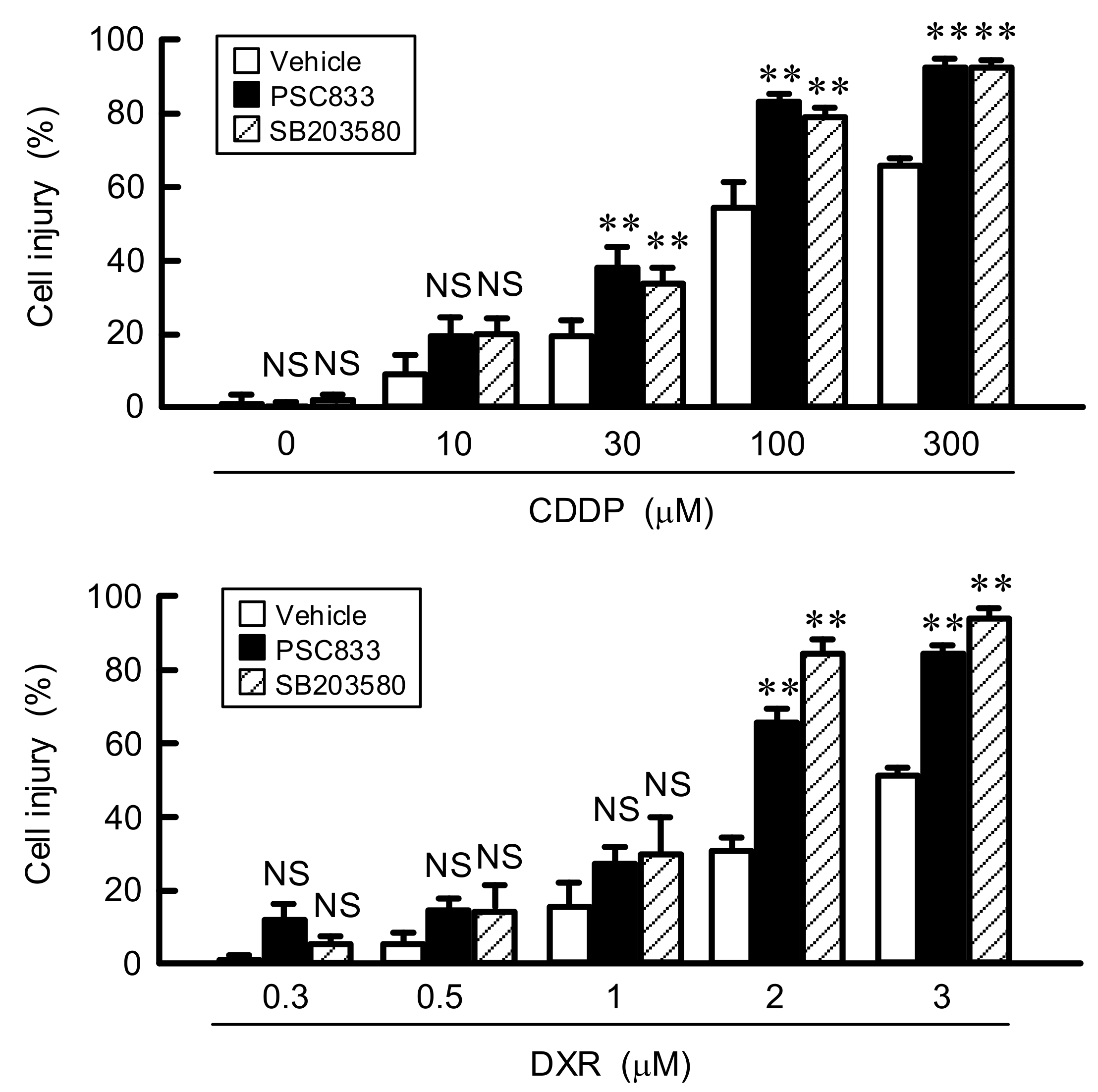
3.6. Effect of LM on Chemoresistance
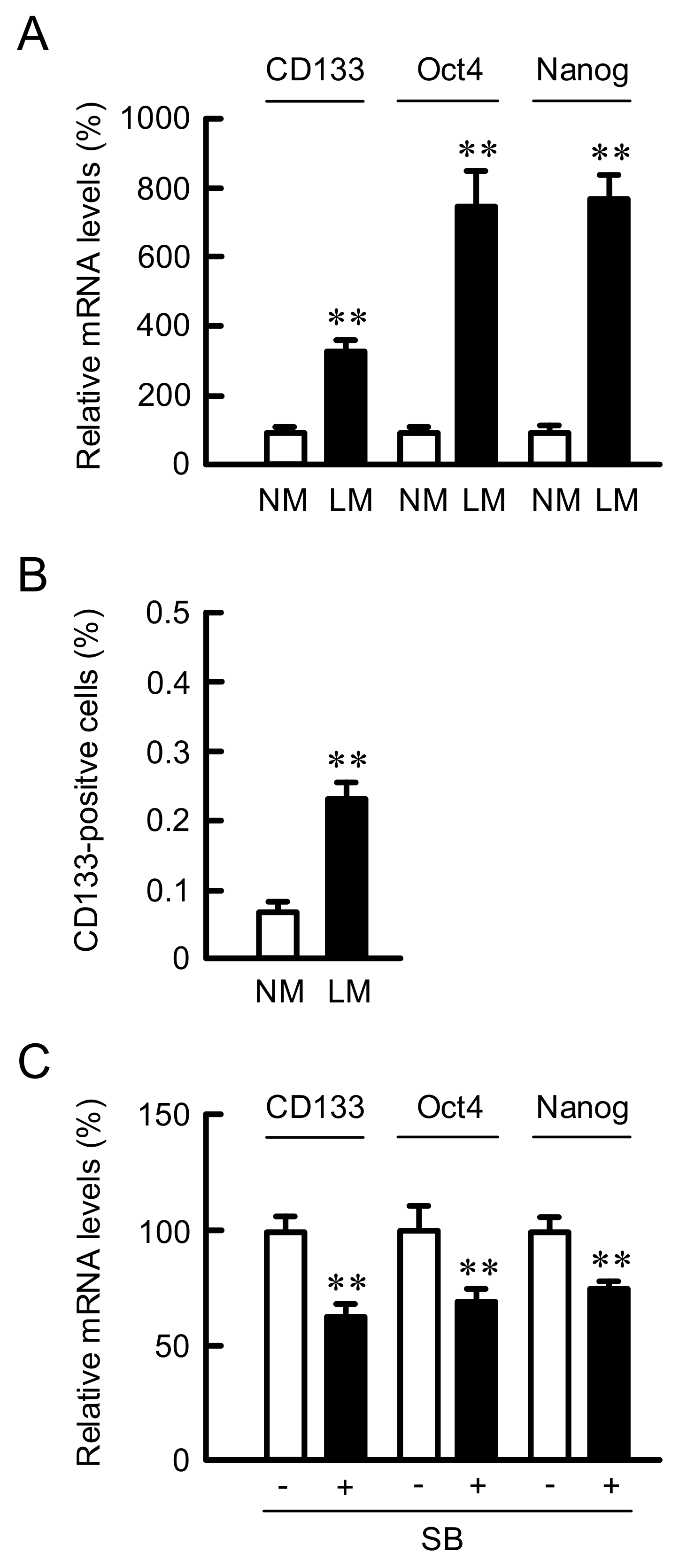
3.7. Effect of LM on the Proportion of Cancer Stem Like Cells (CSLCs)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Baaij, J.H.; Hoenderop, J.G.; Bindels, R.J. Magnesium in man: Implications for health and disease. Physiol. Rev. 2015, 95, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, K.P.; Weber, S.; Peters, M.; Niemann Nejsum, L.; Vitzthum, H.; Klingel, K.; Kratz, M.; Haddad, E.; Ristoff, E.; Dinour, D.; et al. Hypomagnesemia with secondary hypocalcemia is caused by mutations in TRPM6, a new member of the TRPM gene family. Nat. Genet. 2002, 31, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Bi, C.; Song, Y.; Guo, H.; Liu, L.; Zhou, Z.; Wang, B.; Tang, G.; Liu, C.; Yang, Y.; et al. Plasma Magnesium Concentrations and Risk of Incident Cancer in Adults with Hypertension: A Nested Case-Control Study. Ann. Nutr. Metab. 2020, 76, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Qin, F.; Lao, L.; Huang, M.; Tan, H.; Jin, X.; Ma, X.; Zeng, J. Evaluation of the TRPM protein family as potential biomarkers for various types of human cancer using public database analyses. Exp. Ther. Med. 2020, 20, 770–785. [Google Scholar] [CrossRef]
- Velimirovic, M.; Ziperstein, J.C.; Fenves, A.Z. A case of chronic hypomagnesemia in a cancer survivor. Hosp. Pract. 2017, 45, 58–64. [Google Scholar] [CrossRef]
- Pietropaolo, G.; Pugliese, D.; Armuzzi, A.; Guidi, L.; Gasbarrini, A.; Rapaccini, G.L.; Wolf, F.I.; Trapani, V. Magnesium Absorption in Intestinal Cells: Evidence of Cross-Talk between EGF and TRPM6 and Novel Implications for Cetuximab Therapy. Nutrients 2020, 12, 3277. [Google Scholar] [CrossRef]
- Izzedine, H.; Perazella, M.A. Adverse kidney effects of epidermal growth factor receptor inhibitors. Nephrol. Dial. Transpl. 2017, 32, 1089–1097. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Tejpar, S.; Prenen, H.; Van Cutsem, E. Hypomagnesaemia and targeted anti-epidermal growth factor receptor (EGFR) agents. Target. Oncol. 2011, 6, 227–233. [Google Scholar] [CrossRef]
- Schwalfenberg, G.K.; Genuis, S.J. The Importance of Magnesium in Clinical Healthcare. Scientifica 2017, 2017, 4179326. [Google Scholar] [CrossRef]
- Tyson, J.J.; Csikasz-Nagy, A.; Novak, B. The dynamics of cell cycle regulation. Bioessays 2002, 24, 1095–1109. [Google Scholar] [CrossRef] [Green Version]
- Kang, J.; Ferguson, D.; Song, H.; Bassing, C.; Eckersdorff, M.; Alt, F.W.; Xu, Y. Functional interaction of H2AX, NBS1, and p53 in ATM-dependent DNA damage responses and tumor suppression. Mol. Cell Biol. 2005, 25, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Luan, Y.; Yu, R.; Zhang, Z.; Zhang, J.; Wang, W. Transient receptor potential (TRP) channels, promising potential diagnostic and therapeutic tools for cancer. Biosci. Trends 2014, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Guilbert, A.; Gautier, M.; Dhennin-Duthille, I.; Haren, N.; Sevestre, H.; Ouadid-Ahidouch, H. Evidence that TRPM7 is required for breast cancer cell proliferation. Am. J. Physiol. Cell Physiol. 2009, 297, C493–C502. [Google Scholar] [CrossRef]
- Kim, B.J.; Park, E.J.; Lee, J.H.; Jeon, J.H.; Kim, S.J.; So, I. Suppression of transient receptor potential melastatin 7 channel induces cell death in gastric cancer. Cancer Sci. 2008, 99, 2502–2509. [Google Scholar] [CrossRef]
- Min, H.Y.; Lee, H.Y. Mechanisms of resistance to chemotherapy in non-small cell lung cancer. Arch. Pharm. Res. 2021, 44, 146–164. [Google Scholar] [CrossRef]
- Evan, G.I.; Vousden, K.H. Proliferation, cell cycle and apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Klein, I.; Sarkadi, B.; Varadi, A. An inventory of the human ABC proteins. Biochim. Biophys Acta 1999, 1461, 237–262. [Google Scholar] [CrossRef] [Green Version]
- Zou, F.; Seike, M.; Noro, R.; Kunugi, S.; Kubota, K.; Gemma, A. Prognostic significance of ABCB1 in stage I lung adenocarcinoma. Oncol. Lett. 2017, 14, 313–321. [Google Scholar] [CrossRef]
- Kong, W.; Ling, X.; Chen, Y.; Wu, X.; Zhao, Z.; Wang, W.; Wang, S.; Lai, G.; Yu, Z. Hesperetin reverses Pglycoproteinmediated cisplatin resistance in DDPresistant human lung cancer cells via modulation of the nuclear factorkappaB signaling pathway. Int. J. Mol. Med. 2020, 45, 1213–1224. [Google Scholar]
- Kebsa, W.; Lahouel, M.; Rouibah, H.; Zihlif, M.; Ahram, M.; Abu-Irmaileh, B.; Mustafa, E.; Al-Ameer, H.J.; Al Shhab, M. Reversing Multidrug Resistance in Chemo-resistant Human Lung Adenocarcinoma (A549/DOX) Cells by Algerian Propolis Through Direct Inhibiting the P-gp Efflux-pump, G0/G1 Cell Cycle Arrest and Apoptosis Induction. Anticancer. Agents Med. Chem. 2018, 18, 1330–1337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Song, Y.; Cheng, X.; Xu, Z.; Matthew, O.A.; Wang, J.; Sun, Z.; Zhang, X. Apatinib Reverses Paclitaxel-resistant Lung Cancer Cells (A549) Through Blocking the Function of ABCB1 Transporter. Anticancer. Res. 2019, 39, 5461–5471. [Google Scholar] [CrossRef] [PubMed]
- Yeldag, G.; Rice, A.; Del Rio Hernandez, A. Chemoresistance and the Self-Maintaining Tumor Microenvironment. Cancers 2018, 10, 471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broekgaarden, M.; Weijer, R.; van Gulik, T.M.; Hamblin, M.R.; Heger, M. Tumor cell survival pathways activated by photodynamic therapy: A molecular basis for pharmacological inhibition strategies. Cancer Metastasis Rev. 2015, 34, 643–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Vitos-Faleato, J.; Real, S.M.; Gutierrez-Prat, N.; Villanueva, A.; Llonch, E.; Drosten, M.; Barbacid, M.; Nebreda, A.R. Requirement for epithelial p38alpha in KRAS-driven lung tumor progression. Proc. Natl. Acad. Sci. USA 2020, 117, 2588–2596. [Google Scholar] [CrossRef]
- Akizuki, R.; Maruhashi, R.; Eguchi, H.; Kitabatake, K.; Tsukimoto, M.; Furuta, T.; Matsunaga, T.; Endo, S.; Ikari, A. Decrease in paracellular permeability and chemosensitivity to doxorubicin by claudin-1 in spheroid culture models of human lung adenocarcinoma A549 cells. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 769–780. [Google Scholar] [CrossRef]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef]
- Sonoki, H.; Tanimae, A.; Endo, S.; Matsunaga, T.; Furuta, T.; Ichihara, K.; Ikari, A. Kaempherol and luteolin decrease claudin-2 expression mediated by inhibition of STAT3 in lung adenocarcinoma A549 cells. Nutrients 2017, 9, 597. [Google Scholar] [CrossRef]
- Smith, H.L.; Southgate, H.; Tweddle, D.A.; Curtin, N.J. DNA damage checkpoint kinases in cancer. Expert. Rev. Mol. Med. 2020, 22, e2. [Google Scholar] [CrossRef]
- Wu, X.J.; Kassie, F.; Mersch-Sundermann, V. The role of reactive oxygen species (ROS) production on diallyl disulfide (DADS) induced apoptosis and cell cycle arrest in human A549 lung carcinoma cells. Mutat. Res. 2005, 579, 115–124. [Google Scholar] [CrossRef]
- Yang, Y.M.; Noh, K.; Han, C.Y.; Kim, S.G. Transactivation of genes encoding for phase II enzymes and phase III transporters by phytochemical antioxidants. Molecules 2010, 15, 6332–6348. [Google Scholar] [CrossRef] [Green Version]
- Franzellitti, S.; Fabbri, E. Cyclic-AMP mediated regulation of ABCB mRNA expression in mussel haemocytes. PLoS ONE 2013, 8, e61634. [Google Scholar]
- Del Re, M.; Arrigoni, E.; Restante, G.; Passaro, A.; Rofi, E.; Crucitta, S.; De Marinis, F.; Di Paolo, A.; Danesi, R. Concise Review: Resistance to tyrosine kinase inhibitors in non-small cell lung cancer: The Role of Cancer Stem Cells. Stem. Cells 2018, 36, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Safa, A.R. Resistance to Cell Death and Its Modulation in Cancer Stem Cells. Crit. Rev. Oncog. 2016, 21, 203–219. [Google Scholar] [CrossRef]
- Schaffers, O.J.M.; Hoenderop, J.G.J.; Bindels, R.J.M.; de Baaij, J.H.F. The rise and fall of novel renal magnesium transporters. Am. J. Physiol. Renal. Physiol. 2018, 314, F1027–F1033. [Google Scholar] [CrossRef] [Green Version]
- Bartek, J.; Lukas, J. Mammalian G1- and S-phase checkpoints in response to DNA damage. Curr. Opin. Cell Biol. 2001, 13, 738–747. [Google Scholar] [CrossRef]
- Mahabir, S.; Wei, Q.; Barrera, S.L.; Dong, Y.Q.; Etzel, C.J.; Spitz, M.R.; Forman, M.R. Dietary magnesium and DNA repair capacity as risk factors for lung cancer. Carcinogenesis 2008, 29, 949–956. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, W. Classical resistance mechanisms. Int. J. Clin. Pharmacol. Ther. 2009, 47, 46–48. [Google Scholar] [CrossRef]
- Herbst, R.S.; Bunn, P.A., Jr. Targeting the epidermal growth factor receptor in non-small cell lung cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2003, 9, 5813–5824. [Google Scholar]
- Durmus, S.; Hendrikx, J.J.; Schinkel, A.H. Apical ABC transporters and cancer chemotherapeutic drug disposition. Adv. Cancer Res. 2015, 125, 1–41. [Google Scholar] [PubMed]
- Kim, S.J.; Kim, H.S.; Seo, Y.R. Understanding of ROS-Inducing Strategy in Anticancer Therapy. Oxid. Med. Cell Longev. 2019, 2019, 5381692. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.C.; Chiou, P.C.; Chen, P.C.; Liu, P.Y.; Huang, W.C.; Chao, C.C.; Tang, C.H. Melatonin reduces lung cancer stemness through inhibiting of PLC, ERK, p38, beta-catenin, and Twist pathways. Environ. Toxicol. 2019, 34, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Wang, J.; Wang, G.; Zhou, C.; Wang, P.; Zhao, S.; Zhao, S.; Huang, S.; Su, W.; Jiang, P.; et al. Inactivation of p38 MAPK contributes to stem cell-like properties of non-small cell lung cancer. Oncotarget 2017, 8, 26702–26717. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Direction | Sequence (5′→3′) |
|---|---|---|
| TRPM6 | Sense | AAGGACTCCAGGTGCCAAT |
| Antisense | TCCTCTTCAGAGATGCTGTTTTC | |
| TRPM7 | Sense | GCCACTTGGAAACTGGAACC |
| Antisense | CGGTAGATGGCCTTCTACTG | |
| CNNM2 | Sense | GTTCTGGGAATCGTCACCTTAG |
| Antisense | TTTCAGTTCCTGGATGACATTG | |
| MagT1 | Sense | GCAAACTCCTGGCGATACTCC |
| Antisense | ACTGGGCTTGACTGCTTCC | |
| SLC41A1 | Sense | GGTCTTCATCCTAGTGCCTG |
| Antisense | CAAGGTGATGAGGTCGCC | |
| MRS2 | Sense | GACTAATGGGAGTTGCTTTTGG |
| Antisense | AATGGAGCTTCTAGCTGTCGTC | |
| ABCB1 | Sense | CCCATCATTGCAATAGCAGG |
| Antisense | TGTTCAAACTTCTGCTCCTGA | |
| ABCC1 | Sense | ATGTCACGTGGAATACCAGC |
| Antisense | GAAGACTGAACTCCCTTCCT | |
| ABCC2 | Sense | ACAGAGGCTGGTGGCAACC |
| Antisense | ACCATTACCTTGTCACTGTCCATGA | |
| ABCG2 | Sense | AGATGGGTTTCCAAGCGTTCAT |
| Antisense | CCAGTCCCAGTACGACTGTGACA | |
| CD133 | Sense | CGACAATGTAACTCAGCGTCTT |
| Antisense | CACACAGTAAGCCCAGGTAGTA | |
| Oct4 | Sense | GGATCACCCTGGGATATACACA |
| Antisense | TTCATTGTTGTCAGCTTCCTCC | |
| Nanog | Sense | AAATGTCTTCTGCTGAGATGCC |
| Antisense | CTTTGGGACTGGTGGAAGAATC | |
| β-Actin | Sense | CCTGAGGCACTCTTCCAGCCTT |
| Antisense | TGCGGATGTCCACGTCACACTTC |
| Name | Catalog No. | Lot No. | Supplier | Address |
|---|---|---|---|---|
| p-ATM (D6H9) | 5883T | 6 | Cell Signaling Technology | Danvers, MA, USA |
| ATM | 27156-1-AP | 00055078 | ProteinTech | Rosemont, IL, USA |
| p-p53 | 65415 | 1091 | Full Moon Biosystems | Sunnyvale, CA, USA |
| p53 (PAb122) | MS-182-P0 | 182P810D | Thermo Fisher Scientific | San Jose, CA, USA |
| p21 | 14-6715 | 81 | Thermo Fisher Scientific | San Jose, CA, USA |
| ABCB1 | GTX108354 | 39834 | GeneTex | Irvine, CA, USA |
| ABCC1 | GTX116046 | 40135 | GeneTex | Irvine, CA, USA |
| ABCC2 | 4446S | 1 | Cell Signaling Technology | Danvers, MA, USA |
| ABCG2 | GTX100437 | 39471 | GeneTex | Irvine, CA, USA |
| Nrf2 | 16396-1-AP | 10898000 | ProteinTech | Rosemont, IL, USA |
| HIF-1α | GTX113850 | W | GeneTex | Irvine, CA, USA |
| Sp1 (1C6) | sc-420 | K1907 | Santa Cruz Biotechnology | Santa Cruz, CA, USA |
| c-Jun (60A8) | 21021-1 | 7 | Signal Antibody Technology | College Park, MD, USA |
| Nucleoporin p62 | 610497 | 5352647 | BD Biosciences | Franklin Lakes, NJ, USA |
| p38 | 612168 | 15187 | BD Biosciences | Franklin Lakes, NJ, USA |
| β-Actin | sc-1615 | H3016 | Santa Cruz Biotechnology | Santa Cruz, CA, USA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onuma, S.; Manabe, A.; Yoshino, Y.; Matsunaga, T.; Asai, T.; Ikari, A. Upregulation of Chemoresistance by Mg2+ Deficiency through Elevation of ATP Binding Cassette Subfamily B Member 1 Expression in Human Lung Adenocarcinoma A549 Cells. Cells 2021, 10, 1179. https://doi.org/10.3390/cells10051179
Onuma S, Manabe A, Yoshino Y, Matsunaga T, Asai T, Ikari A. Upregulation of Chemoresistance by Mg2+ Deficiency through Elevation of ATP Binding Cassette Subfamily B Member 1 Expression in Human Lung Adenocarcinoma A549 Cells. Cells. 2021; 10(5):1179. https://doi.org/10.3390/cells10051179
Chicago/Turabian StyleOnuma, Saki, Aya Manabe, Yuta Yoshino, Toshiyuki Matsunaga, Tomohiro Asai, and Akira Ikari. 2021. "Upregulation of Chemoresistance by Mg2+ Deficiency through Elevation of ATP Binding Cassette Subfamily B Member 1 Expression in Human Lung Adenocarcinoma A549 Cells" Cells 10, no. 5: 1179. https://doi.org/10.3390/cells10051179
APA StyleOnuma, S., Manabe, A., Yoshino, Y., Matsunaga, T., Asai, T., & Ikari, A. (2021). Upregulation of Chemoresistance by Mg2+ Deficiency through Elevation of ATP Binding Cassette Subfamily B Member 1 Expression in Human Lung Adenocarcinoma A549 Cells. Cells, 10(5), 1179. https://doi.org/10.3390/cells10051179