
Human Umbilical Cord-Derived Mesenchymal Stem Cells Promote Corneal Epithelial Repair In Vitro

 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture of Human Umbilical Cord-Derived Mesenchymal Stem Cells (UC-MSCs)
2.2. Osteogenic, Adipogenic, and Chondrogenic Differentiation of UC-MSCs
2.3. Culture of Human Limbal Epithelial Cells
2.4. Culture of Corneal Epithelial Cells
2.5. Immunofluorescence
2.6. Flow Cytometry
2.7. In Vitro Scratch Assay
2.8. Statistical Analysis
3. Results
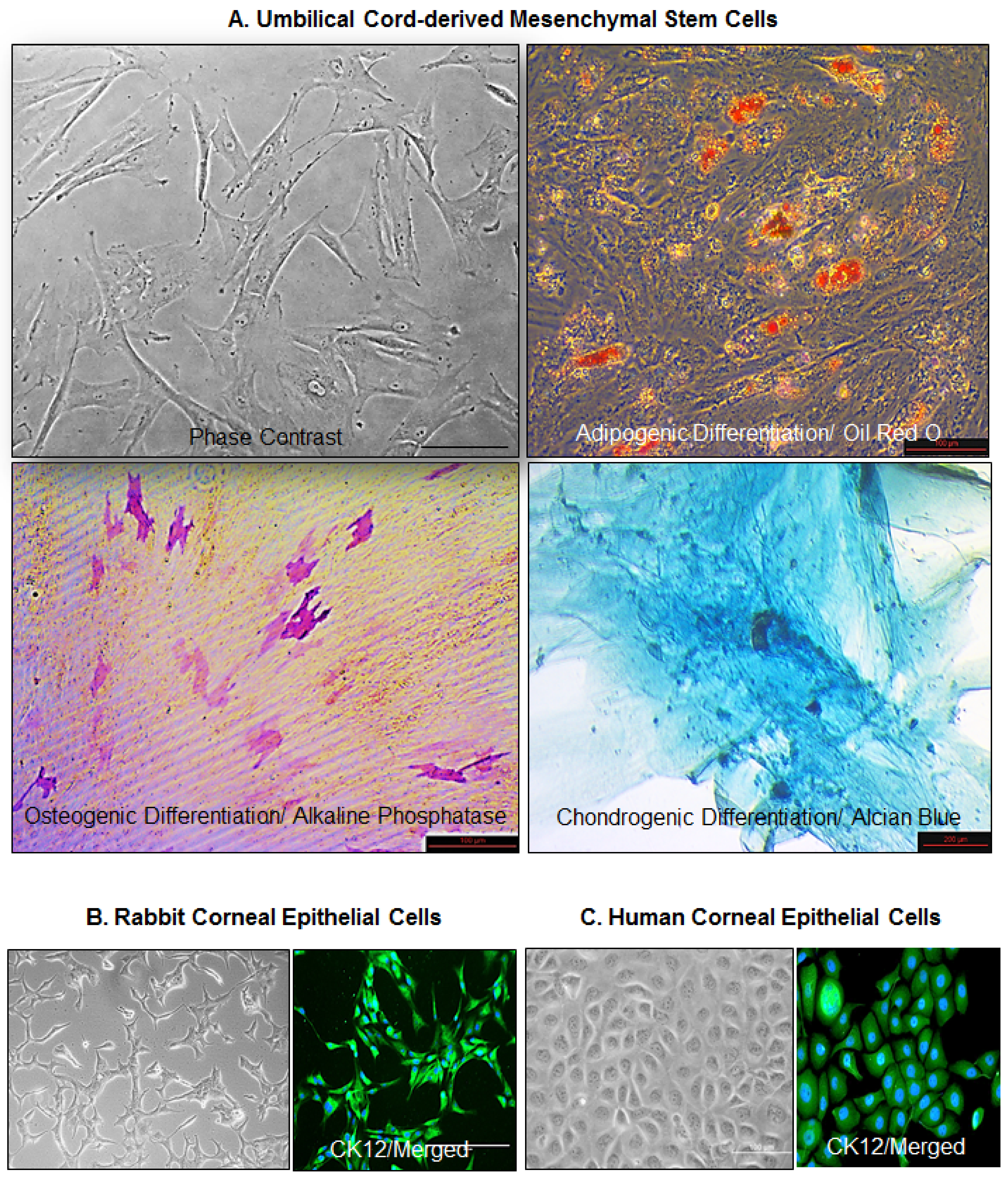
3.1. Culture and Characterization of Cells
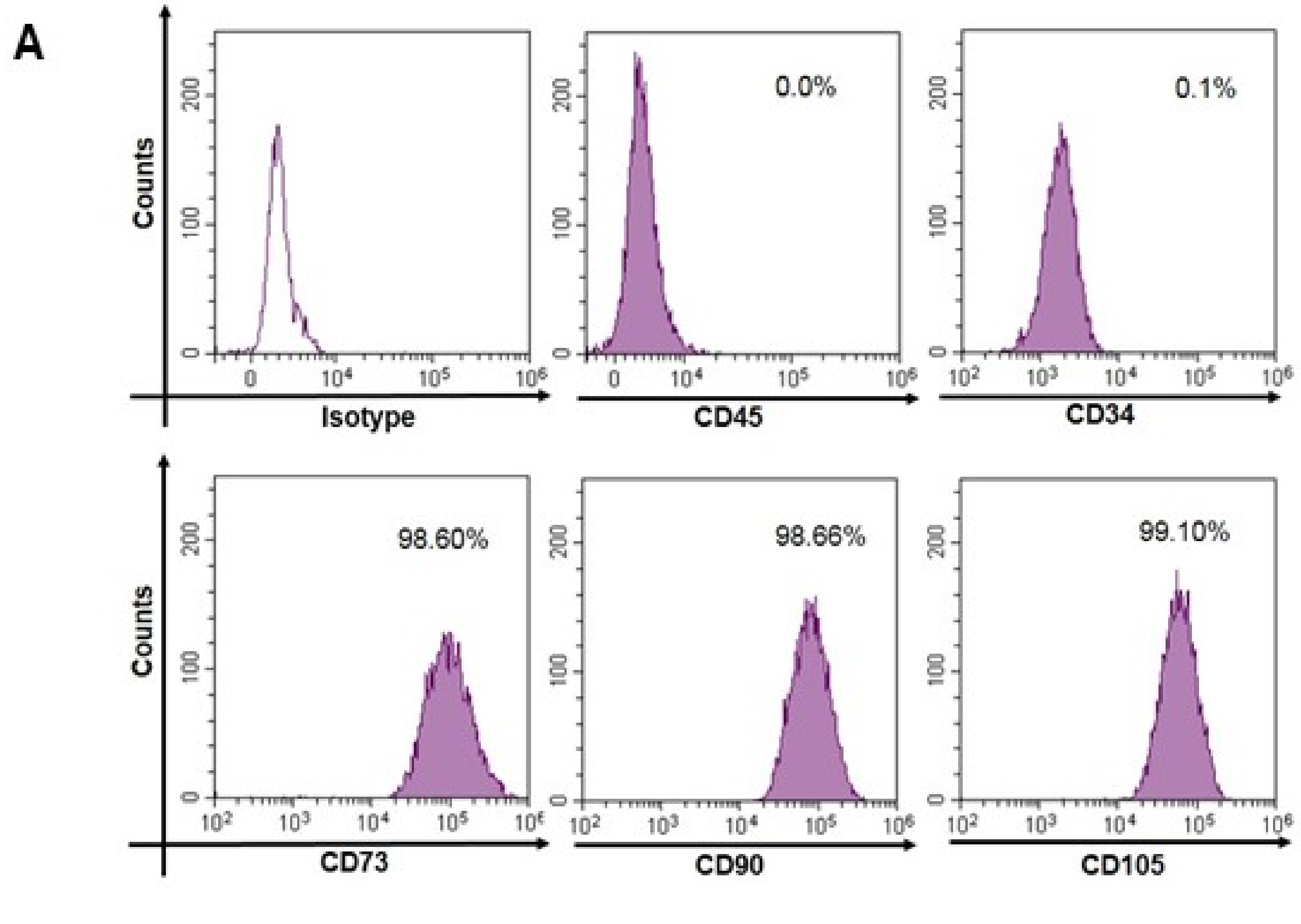
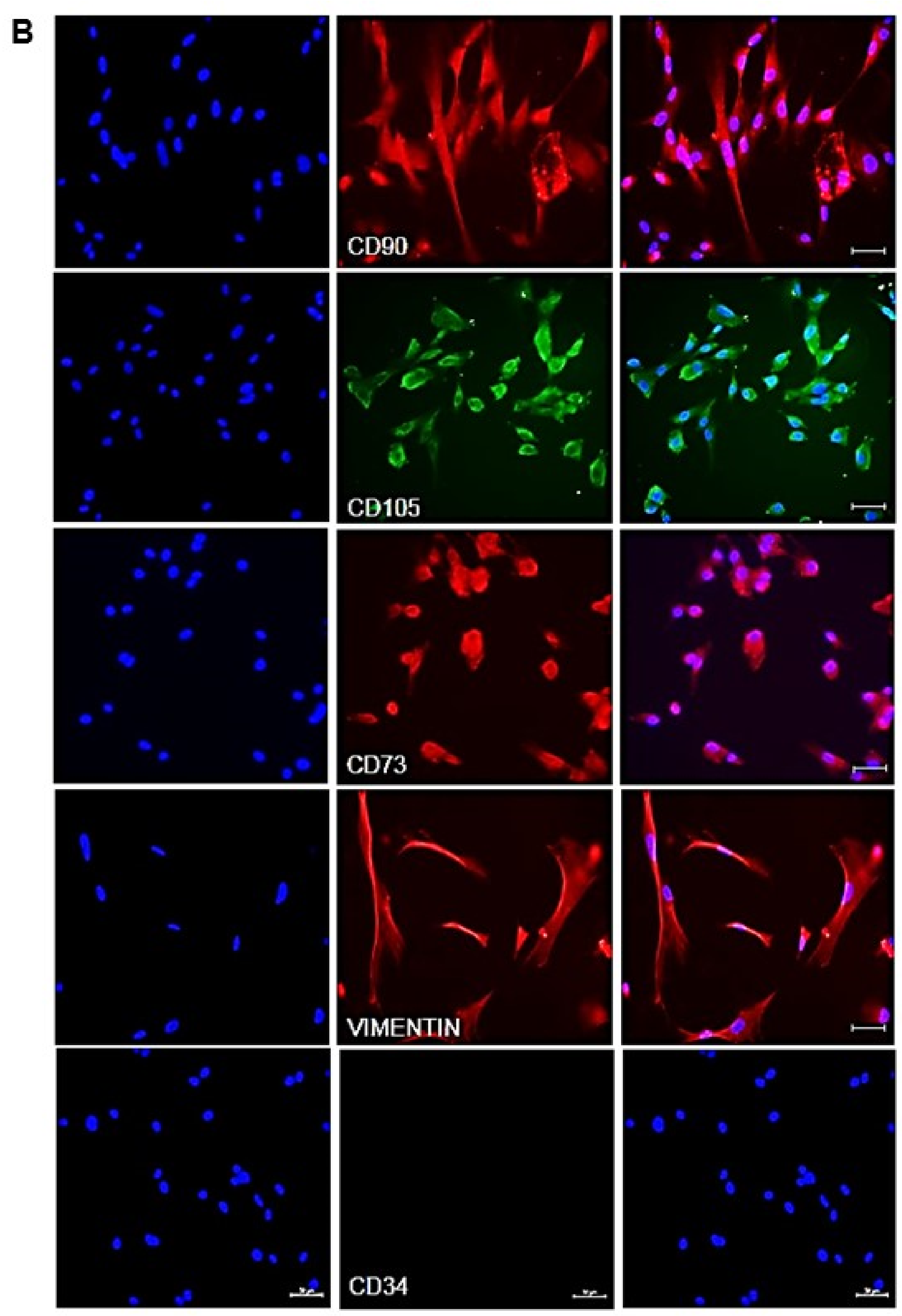
3.2. Expression of Mesenchymal Stem Cell Markers
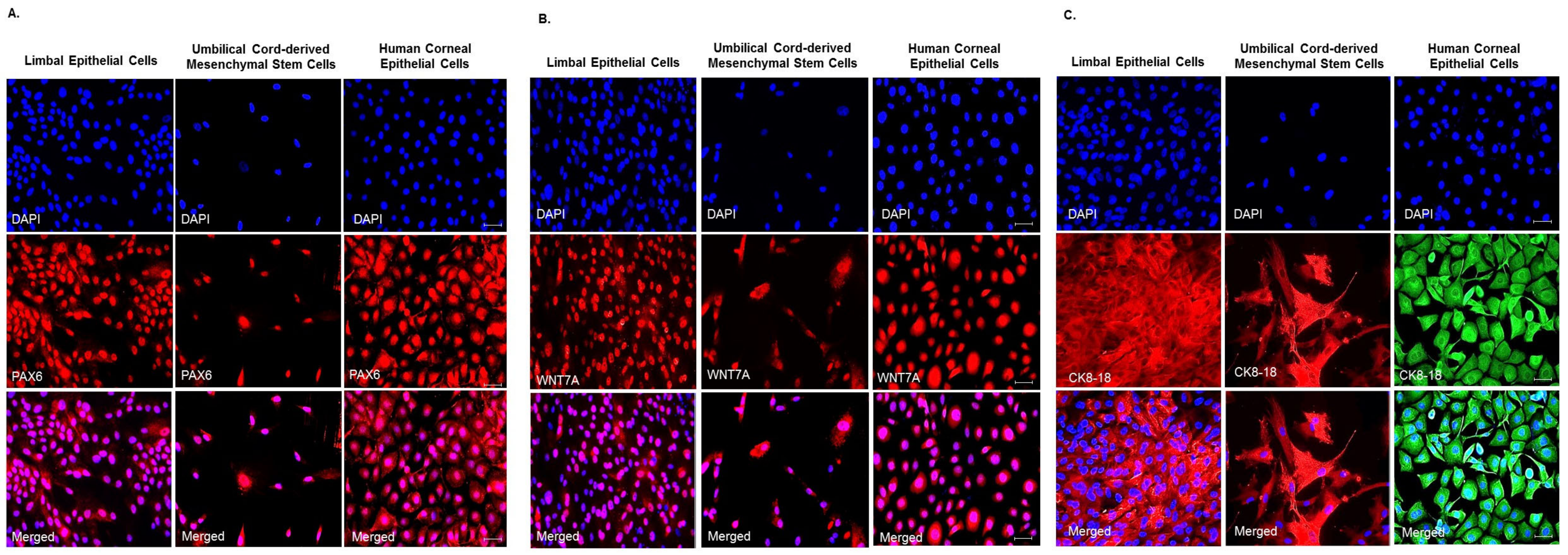
3.3. Expression of Ocular Lineage Markers by UC-MSCs
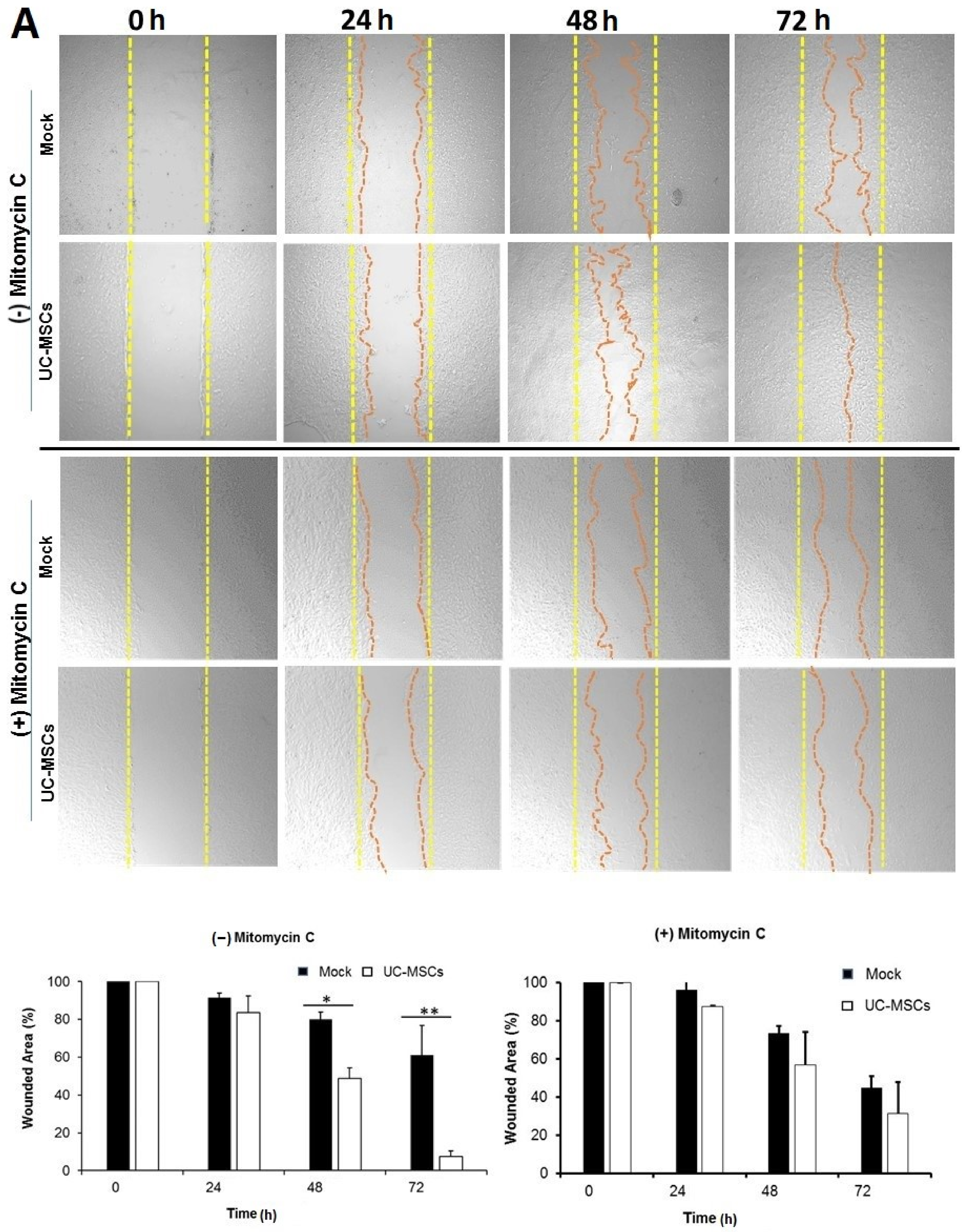
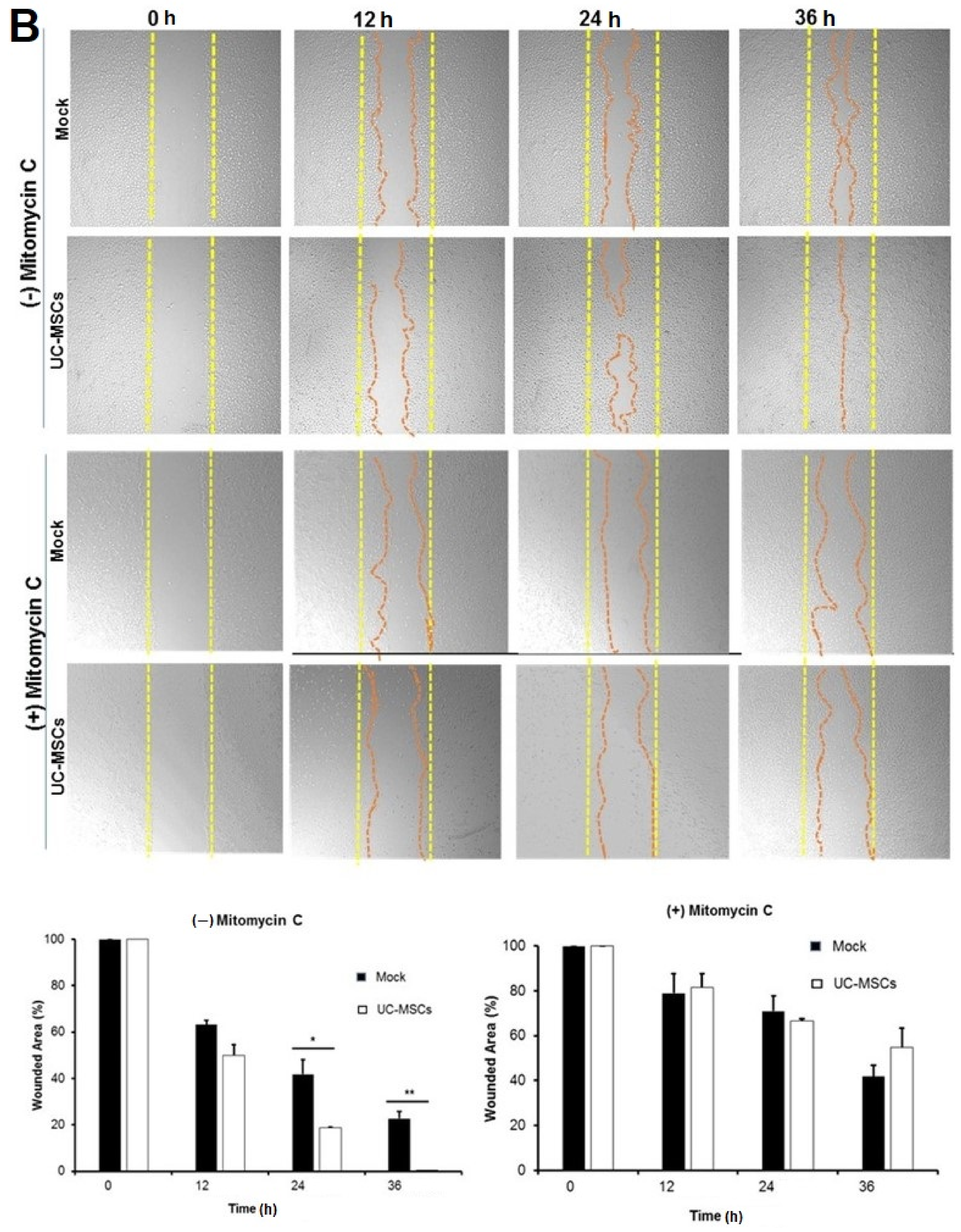
3.4. Role of UC-MSCs in Injury-Induced Corneal Epithelial Repair
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaidyanathan, U.; Hopping, G.C.; Liu, H.Y.; Somani, A.N.; Ronquillo, Y.C.; Hoopes, P.C.; Moshirfar, M. Persistent Corneal Epithelial Defects: A Review Article. Med. Hypothesis Discov. Innov. Ophthalmol. 2019, 8, 163–176. [Google Scholar] [PubMed]
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract. Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Schermer, A.; Galvin, S.; Sun, T.-T. Differentiation-related expression of a major 64K corneal keratin in vivo and in culture suggests limbal location of corneal epithelial stem cells. J. Cell Biol. 1986, 103, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Lavker, R.M.; Sun, T.T. Epithelial stem cells: The eye provides a vision. Eye 2003, 17, 937–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dua, H.S.; Shanmuganathan, A.V.; Powell-Richards, A.; Tighe, P.J.; Joseph, A. Limbal epithelial crypts: A novel anatomical structure and a putative limbal stem cell niche. Br. J. Ophthalmol. 2005, 89, 529–532. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Figueiredo, F.; Lako, M. Corneal epithelial stem cells: Characterization, culture and transplantation. Regen. Med. 2006, 1, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, G.; Traverso, E.C.; Franzi, A.T.; Zingirian, M.; Cancedda, R.; De Luca, M. Long-term restoration of damaged corneal surfaces with autologous cultivated corneal epithelium. Lancet 1997, 349, 990–993. [Google Scholar] [CrossRef]
- Rama, P.; Matuska, S.; Paganoni, G.; Spinelli, A.; De Luca, M.; Pellegrini, G. Limbal Stem-Cell Therapy and Long-Term Corneal Regeneration. N. Engl. J. Med. 2010, 363, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Marchini, G.; Pedrotti, E.; Pedrotti, M.; Barbaro, V.; Di Iorio, E.; Ferrari, S.; Bertolin, M.; Ferrari, B.; Passilongo, M.; Fasolo, A.; et al. Long-term effectiveness of autologous cultured limbal stem cell grafts in patients with limbal stem cell deficiency due to chemical burns. Clin. Exp. Ophthalmol. 2011, 40, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Espana, E.M.; Grueterich, M.; Romano, A.C.; Touhami, A.; Tseng, S.C.G. Idiopathic limbal stem cell deficiency. Ophthalmol. 2002, 109, 2004–2010. [Google Scholar] [CrossRef]
- Daniels, J.T.; Dart, J.K.; Tuft, S.J.; Kha, P.T. Corneal stem cells in review. Wound Repair Regen. 2001, 9, 483–494. [Google Scholar] [CrossRef]
- Gomes, J.Á.P.; Monteiro, B.G.; Melo, G.B.; Smith, R.L.; da Silva, M.C.P.; Lizier, N.F.; Kerkis, A.; Cerruti, H.; Kerkis, I. Corneal reconstruction with tissue-engineered cell sheets composed of human immature dental pulp stem cells. Inv. Ophthalmol. Vis. Sci. 2010, 51, 1408–1414. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Wang, W.; Zhang, J.; Chen, X.; Li, B.Z.; Li, L.S. Experimental study on repairing damage of corneal surface by mesenchymal stem cells transplantation. Zhonghua Yan Ke Za Zhi 2006, 42, 246–250. [Google Scholar] [CrossRef]
- Zhang, C.; Du, L.; Pang, K.; Wu, X. Differentiation of human embryonic stem cells into corneal epithelial progenitor cells under defined conditions. PLoS ONE 2017, 12, e0183303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cieślar-Pobuda, A.; Rafat, M.; Knoflach, V.; Skonieczna, M.; Hudecki, A.; Małecki, A.; Urasińska, E.; Ghavami, S.; Łos, M.J. Human induced pluripotent stem cell differentiation and direct transdifferentiation into corneal epithelial-like cells. Oncotarget 2016, 7, 42314–42329. [Google Scholar] [CrossRef] [PubMed]
- Abumaree, M.; Al Jumah, M.; Pace, R.A.; Kalionis, B. Immunosuppressive Properties of Mesenchymal Stem Cells. Stem Cell Rev. Rep. 2011, 8, 375–392. [Google Scholar] [CrossRef]
- Martínez-Conesa, E.M.; Espel, E.; Reina, M.; Casaroli-Marano, R.P. Characterization of ocular surface epithelial and progenitor cell markers in human adipose stromal cells derived from lipoaspirates. Investig. Ophthalmol. Vis. Sci. 2012, 53, 513–520. [Google Scholar] [CrossRef]
- Hou, G.H.; Ye, N.; Wu, J.; Xu, J.T.; Shi, W.J.; Chen, Y.; Liu, X.H.; Zhang, Z.H. Preliminary study on human bone marrow mesenchymal stem cells differentiation into epithelial-like cells. Zhonghua Yan Ke Za Zhi 2010, 46, 719–724. [Google Scholar]
- Reinshagen, H.; Sorg, R.V.; Boehringer, D.; Eberwein, P.; Sundmacher, R.; Reinhard, T.; Auw-Haedrich, C.; Schwartzkopff, J. Corneal surface reconstruction using adult mesenchymal stem cells in experimental limbal stem cell deficiency in rabbits. Acta Ophthalmol. 2009, 89, 741–748. [Google Scholar] [CrossRef]
- Monteiro, B.G.; Serafim, R.C.; Melo, G.B.; Silva, M.C.P.; Lizier, N.F.; Maranduba, C.M.C.; Smith, R.L.; Kerkis, A.; Cerruti, H.; Gomes, J.A.P.; et al. Human immature dental pulp stem cells share key characteristic features with limbal stem cells. Cell Prolif. 2009, 42, 587–594. [Google Scholar] [CrossRef]
- Saleh, R.; Reza, H.M. Short review on human umbilical cord lining epithelial cells and their potential clinical applications. Stem. Cell. Res. Ther. 2017, 8, 31–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conconi, M.T.; Burra, P.; Di Liddo, R.; Calore, C.; Turetta, M.; Bellini, S.; Bo, P.; Nussdorfer, G.G.; Parnigotto, P.P. CD105(+) cells from Wharton’s jelly show in vitro and in vivo myogenic differentiative potential. Int. J. Mol. Med. 2006, 18, 1089–1096. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.E.; Weiss, M.L.; Mitchell, B.M.; Martin, P.; Davis, D.; Morales, L.; Helwig, B.; Beerenstrauch, M.; Abou-Easa, K.; Hildreth, T.; et al. Matrix Cells from Wharton’s Jelly Form Neurons and Glia. Stem. Cells 2003, 21, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.T.; Fan, J.; Cai, Y.Q.; Zhao, S.J.; Xue, S.; Lin, J.H.; Jiang, X.D.; Xu, R.X. Human Wharton’s jelly cells can be induced to differentiate into growth factor-secreting oligodendrocyte progenitor-like cells. Differentiation 2010, 79, 15–20. [Google Scholar] [CrossRef]
- Joyce, N.C.; Harris, D.L.; Markov, V.; Zhang, Z.; Saitta, B. Potential of human umbilical cord blood mesenchymal stem cells to heal damaged corneal endothelium. Mol. Vis. 2012, 18, 64–547. [Google Scholar]
- Campard, D.; Lysy, P.A.; Najimi, M.; Sokal, E.M. Native umbilical cord matrix stem cells express hepatic markers and differ-entiate into hepatocyte-like cells. Gastroenterology 2008, 134, 833–848. [Google Scholar] [CrossRef]
- El Omar, R.; Beroud, J.; Stoltz, J.F.; Menu, P.; Velot, E.; Decot, V. Umbilical cord mesenchymal stem cells: The new gold standard for mesenchymal stem cell-based therapies? Tissue Eng. Part B Rev. 2014, 20, 523–544. [Google Scholar] [CrossRef]
- Reza, H.M.; Ng, B.-Y.; Gimeno, F.L.; Phan, T.T.; Ang, L.P.-K. Umbilical Cord Lining Stem Cells as a Novel and Promising Source for Ocular Surface Regeneration. Stem Cell Rev. Rep. 2011, 7, 935–947. [Google Scholar] [CrossRef]
- Reza, H.M.; Ng, B.-Y.; Phan, T.T.; Tan, D.T.H.; Beuerman, R.W.; Ang, L.P.-K. Characterization of a Novel Umbilical Cord Lining Cell with CD227 Positivity and Unique Pattern of P63 Expression and Function. Stem Cell Rev. Rep. 2011, 7, 624–638. [Google Scholar] [CrossRef] [PubMed]
- Garzón, I.; Martín-Piedra, M.A.; Alfonso-Rodríguez, C.; González-Andrades, M.; Carriel, V.; Martínez-Gómez, C.; Campos, A.; Alaminos, M. Generation of a Biomimetic Human Artificial Cornea Model Using Wharton’s Jelly Mesenchymal Stem Cells. Investig. Opthalmology Vis. Sci. 2014, 55, 4073–4083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Ballen, J.; Liu, C.Y.; Wang, I.J.; Sieber, M.; Chang, J.; Jester, J.V.; Kao, W.W. Cell therapy of congenital corneal diseases with um-bilical mesenchymal stem cells: Lumican null mice. PLoS ONE 2010, 19, e10707. [Google Scholar] [CrossRef]
- Thomas, V.J.C.-; Caterson, B.; Kao, W.W.-Y. Transplantation of human umbilical mesenchymal stem cells cures the corneal defects of mucopolysaccharidosis VII mice. Stem Cells 2013, 31, 2116–2126. [Google Scholar] [CrossRef] [Green Version]
- Mennan, C.; Wright, K.; Bhattacharjee, A.; Balain, B.; Richardson, J.; Roberts, S. Isolation and Characterisation of Mesenchymal Stem Cells from Different Regions of the Human Umbilical Cord. Biomed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Eswaramoorthy, S.D.; Dhiman, N.; Korra, G.; Oranges, C.M.; Schaefer, D.J.; Rath, S.N.; Madduri, S. Isogenic-induced endothelial cells enhance osteogenic differentiation of mesenchymal stem cells on silk fibroin scaffold. Regen. Med. 2019, 14, 647–661. [Google Scholar] [CrossRef] [Green Version]
- Mariappan, I.; Maddileti, S.; Savy, S.; Tiwari, S.; Gaddipati, S.; Fatima, A.; Sangwan, V.S.; Balasubramanian, D.; Vemuganti, G.K. In vitro culture and expansion of human limbal epithelial cells. Nat. Protoc. 2010, 5, 1470–1479. [Google Scholar] [CrossRef]
- Araki-Sasaki, K.; Ohashi, Y.; Sasabe, T.; Hayashi, K.; Watanabe, H.; Tano, Y.; Handa, H. An SV40-immortalized human corneal ep-ithelial cell line and its characterization. Investig. Ophthalmol. Vis. Sci. 1995, 36, 614–621. [Google Scholar]
- Knorr, M.; Steuhl, K.P.; Tatje, D.; Hoppe, J. A rabbit corneal epithelial cell line expresses functional platelet-derived growth factor beta-type receptors. Investig. Ophthalmol. Vis. Sci. 1992, 33, 2207–2211. [Google Scholar]
- Majumdar, S.; Tirucherai, G.S.; Pal, D.; Mitra, A.K. Functional differences in nucleoside and nucleobase transporters expressed on the rabbit corneal epithelial cell line (SIRC) and isolated rabbit cornea. Aaps Pharm.Sci. 2003, 5, 72–85. [Google Scholar] [CrossRef] [Green Version]
- Olivieri, M.; Cristaldi, M.; Pezzino, S.; Rusciano, D.; Tomasello, B.; Anfuso, C.D.; Lupo, G. Phenotypic characterization of the SIRC (Statens Seruminstitut Rabbit Cornea) cell line reveals a mixed epithelial and fibroblastic nature. Exp. Eye Res. 2018, 172, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Swamynathan, S.K. Ocular Surface Development and Gene Expression. J. Ophthalmol. 2013, 2013, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simpson, T.I.; Price, D.J. Pax6; A pleiotropic player in development. BioEssays 2002, 24, 1041–1051. [Google Scholar] [CrossRef]
- Shaham, O.; Menuchin, Y.; Farhy, C.; Ashery-Padan, R. Pax6: A multi-level regulator of ocular development. Prog. Retin. Eye Res. 2012, 31, 351–376. [Google Scholar] [CrossRef]
- Nakatsu, M.N.; Ding, Z.; Ng, M.Y.; Truong, T.T.; Yu, F.; Deng, S.X. Wnt/β-Catenin Signaling Regulates Proliferation of Human Cornea Epithelial Stem/Progenitor Cells. Investig. Opthalmology Vis. Sci. 2011, 52, 4734–4741. [Google Scholar] [CrossRef] [PubMed]
- Merjava, S.; Neuwirth, A.; Tanzerova, M.; Jirsova, K. The spectrum of cytokeratins expressed in the adult human cornea, limbus and perilimbal conjunctiva. Histol. Histopathol. 2011, 26, 323–331. [Google Scholar] [CrossRef]
- Ballen, K.K.; Gluckman, E.; Broxmeyer, H.E. Umbilical cord blood transplantation: The first 25 years and beyond. Blood 2013, 122, 491–498. [Google Scholar] [CrossRef] [Green Version]
- Sivalingam, J.; Krishnan, S.; Ng, W.H.; Lee, S.S.; Phan, T.T.; Kon, O.L. Biosafety Assessment of Site-directed Transgene Integration in Human Umbilical Cord–lining Cells. Mol. Ther. 2010, 18, 1346–1356. [Google Scholar] [CrossRef] [PubMed]
- Eswaramoorthy, S.D.; Ramakrishna, S.; Rath, S.N. Recent advances in three-dimensional bioprinting of stem cells. J. Tissue Eng. Regen. Med. 2019, 13, 908–924. [Google Scholar] [CrossRef] [PubMed]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Gehring, W.J.; Ikeo, K. Pax 6: Mastering eye morphogenesis and eye evolution. Trends Genet. 1999, 15, 371–377. [Google Scholar] [CrossRef]
- Shukla, S.; Mishra, R. Functional analysis of missense mutations G36A and G51A in PAX6, and PAX6(5a) causing ocular anomalies. Exp. Eye Res. 2011, 93, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Xue, Y.; Lin, Y.; Zhang, X.; Xi, L.; Patel, S.; Cai, H.; Luo, J.; Zhang, M.; Yang, Y. WNT7A and PAX6 define corneal epithelium homeostasis and pathogenesis. Nature 2014, 511, 358–361. [Google Scholar] [CrossRef]
- Sun, J.; Liu, W.; Deng, F.; Luo, Y.; Wen, K.; Zhang, H.; Liu, H.; Wu, J.; Su, B.; Liu, Y. Differentiation of rat adipose-derived mesenchymal stem cells into corneal-like epithelial cells driven by PAX6. Exp. Ther. Med. 2017, 15, 1424–1432. [Google Scholar] [CrossRef]
- Shukla, S.; Mittal, S.K.; Foulsham, W.; Elbasiony, E.; Singhania, D.; Sahu, S.K.; Chauhan, S.K. Therapeutic efficacy of different routes of mesenchymal stem cell administration in corneal injury. Ocul. Surf. 2019, 17, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Garzón, I.; Alfonso-Rodríguez, C.; Martínez-Gómez, C.; Carriel, V.; Martin-Piedra, M.; Fernández-Valadés, R.; Sanchez-Quevedo, M.; Alaminos, M. Expression of epithelial markers by human umbilical cord stem cells. A topographical analysis. Placenta 2014, 35, 994–1000. [Google Scholar] [CrossRef]
- Ye, J.; Yao, K.; Kim, J.C. Mesenchymal stem cell transplantation in a rabbit corneal alkali burn model: Engraftment and involvement in wound healing. Eye 2005, 20, 482–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabrowski, F.A.; Burdzinska, A.; Kulesza, A.; Sladowska, A.; Zolocinska, A.; Gala, K.; Paczek, L.; Wielgos, M. Comparison of the paracrine activity of mesenchymal stem cells derived from human umbilical cord, amniotic membrane and adipose tissue. J. Obstet. Gynaecol. Res. 2017, 43, 1758–1768. [Google Scholar] [CrossRef]
- Morita, S.-I.; Shirakata, Y.; Shiraishi, A.; Kadota, Y.; Hashimoto, K.; Higashiyama, S.; Ohashi, Y. Human corneal epithelial cell proliferation by epiregulin and its cross-induction by other EGF family members. Mol. Vis. 2007, 13, 2119–2128. [Google Scholar]
- Basu, S.; Hertsenberg, A.J.; Funderburgh, M.L.; Burrow, M.K.; Mann, M.M.; Du, Y.; Lathrop, K.L.; Syed-Picard, F.N.; Adams, S.M.; Birk, D.E.; et al. Human limbal biopsy-derived stromal stem cells prevent corneal scarring. Sci. Transl. Med. 2014, 6, 266ra172. [Google Scholar] [CrossRef] [Green Version]
- Ziaei, M.; Zhang, J.; Patel, D.V.; McGhee, C.N. Umbilical cord stem cells in the treatment of corneal disease. Surv. Ophthalmol. 2017, 62, 803–815. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No. | Antibody | Supplier | Catalogue/Clone | Concentration * | Dilution |
|---|---|---|---|---|---|
| 1 | CK-8/18 Mouse monoclonal | Santacruz | sc-52325/NCL-5D3 | 200 µg/mL | 1:100 |
| 2 | CD-34 Mouse monoclonal | Santacruz | sc-7324/ICO115 | 200 µg/mL | 1:100 |
| 3 | CD-73 Rabbit monoclonal | Cell Signaling | #13160/D7F9A | 155.2 µg/mL | 1:100 |
| 4 | CD-90 Mouse monoclonal | Santacruz | sc-59396/AF-9 | 100 µg/mL | 1:100 |
| 5 | CD-105 Mouse monoclonal | Santacruz | sc-376381/A-8 | 200 µg/mL | 1:100 |
| 6 | PAX-6 Rabbit polyclonal | Santacruz | sc-11357/H-295 | 200 µg/mL | 1:100 |
| 7 | WNT-7A Mouse monoclonal | Santacruz | sc-365665/E-9 | 200 µg/mL | 1:100 |
| 8 | VIMENTIN Mouse monoclonal | Santacruz | sc-6260/V9 | 200 µg/mL | 1:100 |
| S. No. | Antibody | Supplier/Catalogue | Concentration * | Dilution |
|---|---|---|---|---|
| 1 | Alexa flour® 488 Goat Anti-Mouse IgG | Abcam/ab150113 | 2 mg/mL | 1:400 |
| 2 | Alexa Fluor® 594 Goat Anti-Mouse IgG | Abcam/ab150120 | 2 mg/mL | 1:400 |
| 3 | Alexa Fluor® 488 Goat Anti-Rabbit IgG | Abcam/ab150077 | 2 mg/mL | 1:400 |
| 4 | Alexa Fluor® 594 Goat Anti-Rabbit IgG | Abcam/ab150080 | 2 mg/mL | 1:400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kacham, S.; Bhure, T.S.; Eswaramoorthy, S.D.; Naik, G.; Rath, S.N.; Parcha, S.R.; Basu, S.; Sangwan, V.S.; Shukla, S. Human Umbilical Cord-Derived Mesenchymal Stem Cells Promote Corneal Epithelial Repair In Vitro. Cells 2021, 10, 1254. https://doi.org/10.3390/cells10051254
Kacham S, Bhure TS, Eswaramoorthy SD, Naik G, Rath SN, Parcha SR, Basu S, Sangwan VS, Shukla S. Human Umbilical Cord-Derived Mesenchymal Stem Cells Promote Corneal Epithelial Repair In Vitro. Cells. 2021; 10(5):1254. https://doi.org/10.3390/cells10051254
Chicago/Turabian StyleKacham, Santhosh, Tejal Sunil Bhure, Sindhuja D. Eswaramoorthy, Gaurav Naik, Subha Narayan Rath, Sreenivasa Rao Parcha, Sayan Basu, Virender Singh Sangwan, and Sachin Shukla. 2021. "Human Umbilical Cord-Derived Mesenchymal Stem Cells Promote Corneal Epithelial Repair In Vitro" Cells 10, no. 5: 1254. https://doi.org/10.3390/cells10051254
APA StyleKacham, S., Bhure, T. S., Eswaramoorthy, S. D., Naik, G., Rath, S. N., Parcha, S. R., Basu, S., Sangwan, V. S., & Shukla, S. (2021). Human Umbilical Cord-Derived Mesenchymal Stem Cells Promote Corneal Epithelial Repair In Vitro. Cells, 10(5), 1254. https://doi.org/10.3390/cells10051254