
The Role of Trogocytosis in the Modulation of Immune Cell Functions


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
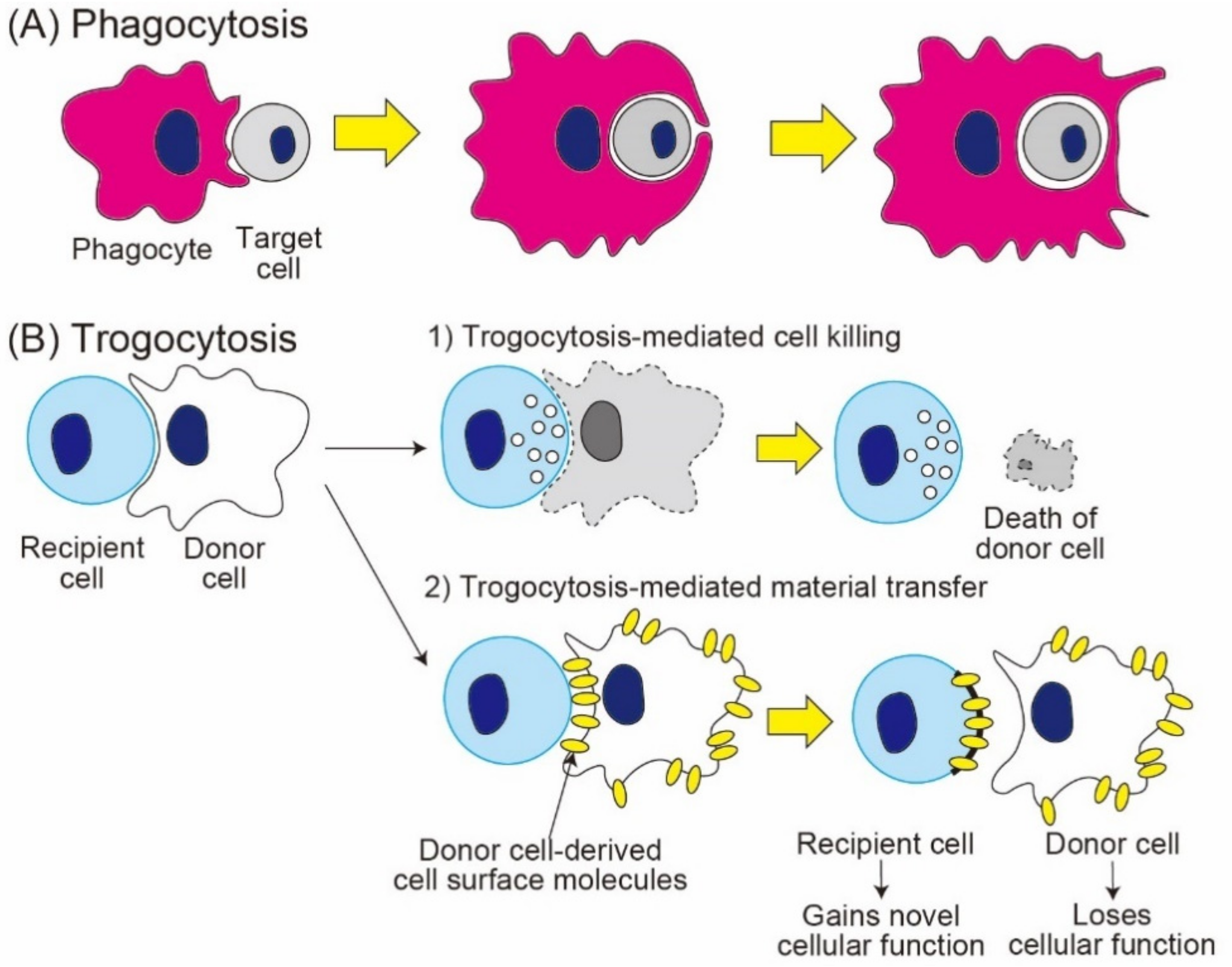
2. Trogocytosis-Mediated Cell Death
3. Trogocytosis Triggered by LIGAND–Receptor Interaction-1: TCR-Mediated Trogocytosis
3.1. Overview of Trogocytosis Triggered by TCR–MHC Interaction
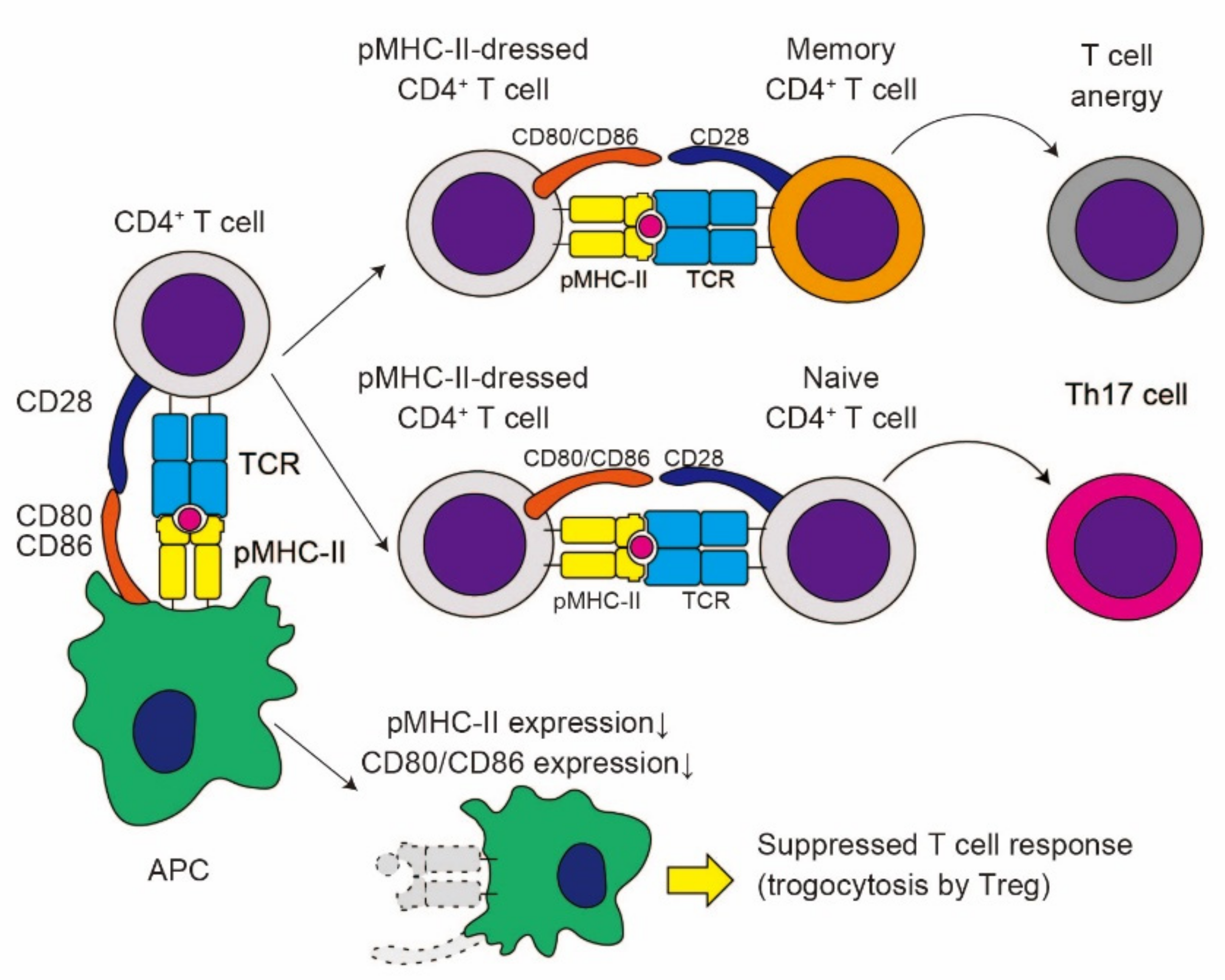
3.2. The Impact of TCR-Mediated Trogocytosis in CD4+ T Cell Functions
3.3. The Impact of TCR-Mediated Trogocytosis on CD8+ T Cell Functions
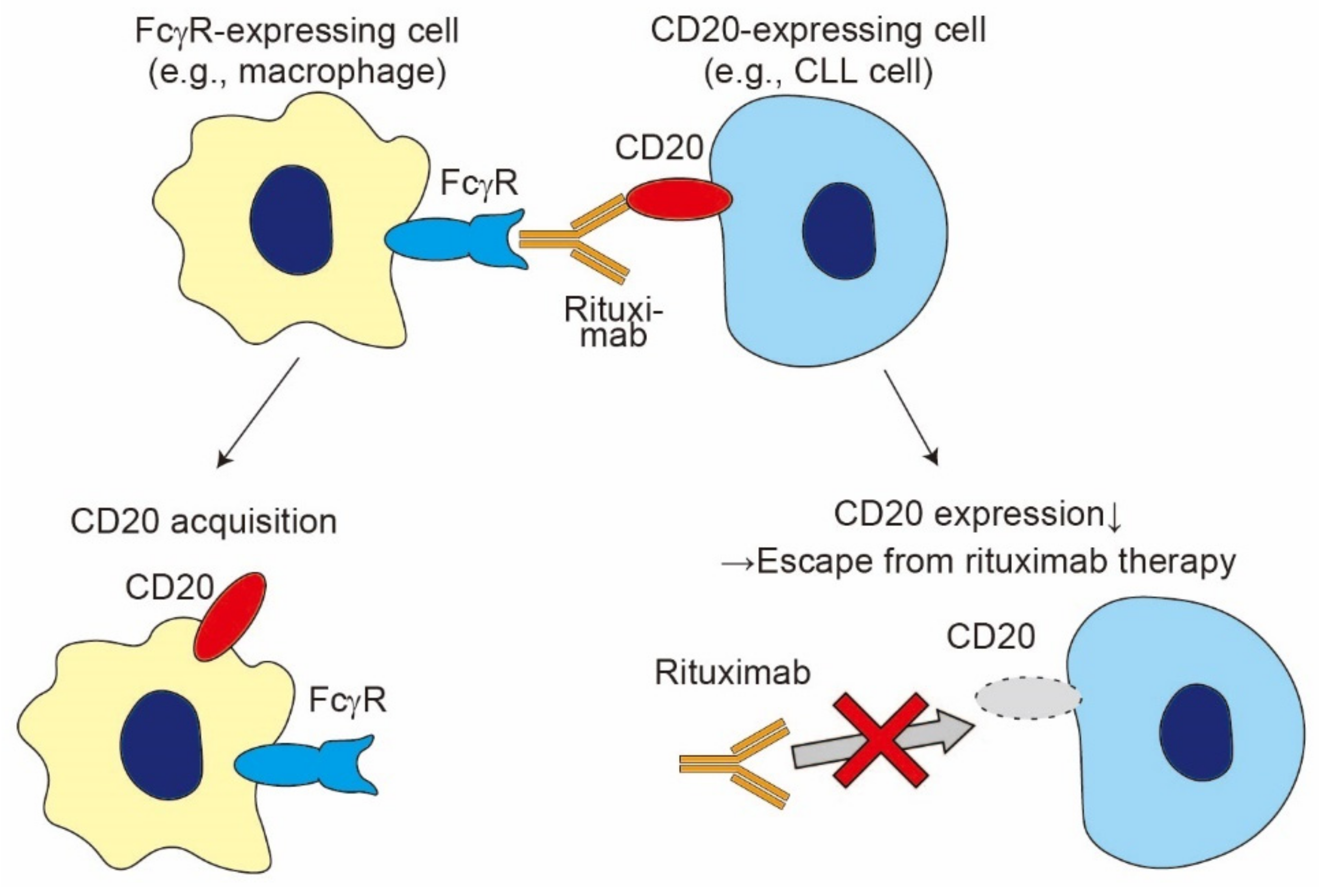
4. Trogocytosis Triggered by Ligand–Receptor Interaction-2: FcγR-Mediated Trogocytosis
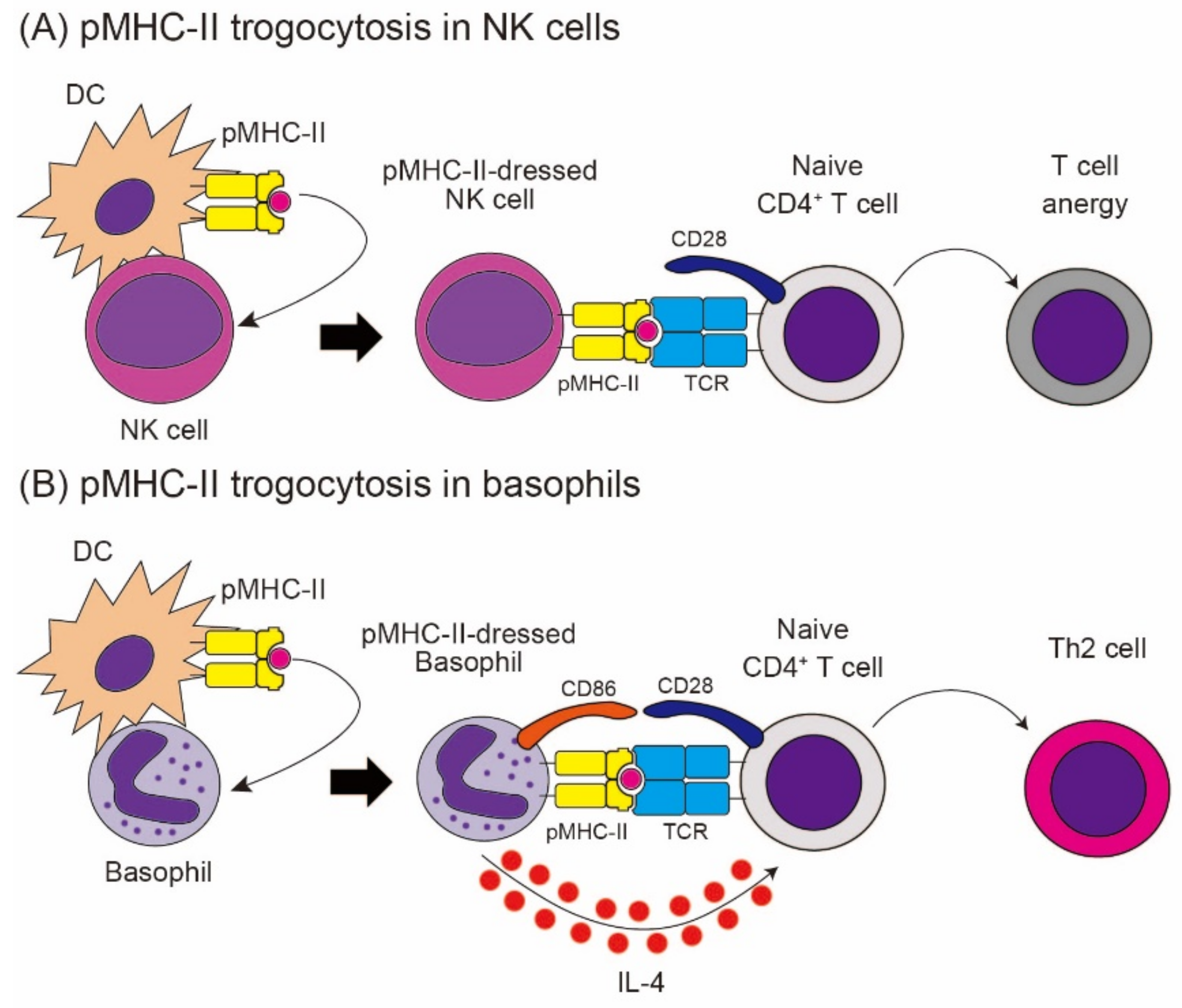
5. Trogocytosis Triggered by Ligand–Receptor Interaction-3: NK Receptor-Mediated Trogocytosis
6. Trogocytosis-Mediated MHC Transfer
6.1. Overview of the Trogocytosis-Mediated Transfer of MHC Molecules
6.2. Transfer of pMHC-I Complexes between DCs (Cross-Dressing)
6.3. pMHC-II Transfer from APCs to Immune Cells Other Than T Cells
7. Immune Cell Trogocytosis Exploited by Pathogen Microbes
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nakayama, M. Antigen presentation by MHC-dressed cells. Front. Immunol. 2014, 5, 672. [Google Scholar] [PubMed] [Green Version]
- Campana, S.; de Pasquale, C.; Carrega, P.; Ferlazzo, G.; Bonaccorsi, I. Cross-dressing: An alternative mechanism for antigen presentation. Immunol. Lett. 2015, 168, 349–354. [Google Scholar]
- Taylor, R.P.; Lindorfer, M.A. Fcgamma-receptor-mediated trogocytosis impacts mAb-based therapies: Historical precedence and recent developments. Blood 2015, 125, 762–766. [Google Scholar] [PubMed] [Green Version]
- Andoh, M.; Ikegaya, Y.; Koyama, R. Synaptic pruning by microglia in epilepsy. J. Clin. Med. 2019, 8, 2170. [Google Scholar]
- Dance, A. Core concept: Cells nibble one another via the under-appreciated process of trogocytosis. Proc. Natl. Acad. Sci. USA 2019, 116, 17608–17610. [Google Scholar] [PubMed] [Green Version]
- Bettadapur, A.; Miller, H.W.; Ralston, K.S. Biting off what can be chewed: Trogocytosis in health, infection, and disease. Infect. Immun. 2020, 88, e00919–e00930. [Google Scholar]
- Li, K.J.; Wu, C.H.; Lu, C.H.; Shen, C.Y.; Kuo, Y.M.; Tsai, C.Y.; Hsieh, S.C.; Yu, C.L. Trogocytosis between non-immune cells for cell clearance, and among immune-related cells for modulating immune responses and autoimmunity. Int. J. Mol. Sci. 2021, 22, 2236. [Google Scholar]
- Joly, E.; Hudrisier, D. What is trogocytosis and what is its purpose? Nat. Immunol. 2003, 4, 815. [Google Scholar]
- Brown, T. Observations by immunofluorescence microscopy and electron microscopy on the cytopathogenicity of Naegleria fowleri in mouse embryo-cell cultures. J. Med. Microbiol. 1979, 12, 363–371. [Google Scholar]
- Ralston, K.S.; Solga, M.D.; Mackey-Lawrence, N.M.; Islamia, J.M.; Bhattacharya, A.; Petri, W.A., Jr. Trogocytosis by Entamoeba histolytica contributes to cell killing and tissue invasion. Nature 2014, 508, 526–530. [Google Scholar]
- Islamia, J.M.; Nakada-Tsukui, K.; Nozaki, T. AGC family kinase 1 participates in trogocytosis but not in phagocytosis in Entamoeba histolytica. Nat. Commun. 2017, 8, 101. [Google Scholar]
- Saito-Nakano, Y.; Wahyuni, R.; Nakada-Tsukui, K.; Tomii, K.; Nozaki, T. Rab7D small GTPase is involved in phago-, trogocytosis and cytoskeletal reorganization in the enteric protozoan Entamoeba histolytica. Cell. Microbiol. 2021, 23, e13267. [Google Scholar] [PubMed]
- Cone, R.E.; Sprent, J.; Marchalonis, J.J. Antigen-binding specificity of isolated cell-surface immunoglobulin from thymus cells activated to histocompatibility antigens. Proc. Natl. Acad. Sci. USA 1972, 69, 2556–2560. [Google Scholar]
- Hudson, L.; Sprent, J.; Miller, J.F.; Playfair, J.H. B cell-derived immunoglobulin on activated mouse T lymphocytes. Nature 1974, 251, 60–62. [Google Scholar]
- Sharrow, S.O.; Ozato, K.; Sachs, D.H. Phenotypic expression of I-A and I-E/C subregion determinants on murine thymocytes. J. Immunol. 1980, 125, 2263–2268. [Google Scholar]
- Sharrow, S.O.; Mathieson, B.J.; Singer, A. Cell surface appearance of unexpected host MHC determinants on thymocytes from radiation bone marrow chimeras. J. Immunol. 1981, 126, 1327–1335. [Google Scholar]
- Pham, T.; Mero, P.; Booth, J.W. Dynamics of macrophage trogocytosis of rituximab-coated B cells. PLoS ONE 2011, 6, e14498. [Google Scholar]
- Bonaccorsi, I.; Morandi, B.; Antsiferova, O.; Costa, G.; Oliveri, D.; Conte, R.; Pezzino, G.; Vermiglio, G.; Anastasi, G.P.; Navarra, G.; et al. Membrane transfer from tumor cells overcomes deficient phagocytic ability of plasmacytoid dendritic cells for the acquisition and presentation of tumor antigens. J. Immunol. 2014, 192, 824–832. [Google Scholar] [PubMed]
- Li, K.J.; Wu, C.H.; Shen, C.Y.; Kuo, Y.M.; Yu, C.L.; Hsieh, S.C. Membrane transfer from mononuclear cells to polymorphonuclear neutrophils transduces cell survival and activation signals in the recipient cells via anti-extrinsic apoptotic and MAP kinase signaling pathways. PLoS ONE 2016, 11, e0156262. [Google Scholar]
- Miyake, K.; Shiozawa, N.; Nagao, T.; Yoshikawa, S.; Yamanishi, Y.; Karasuyama, H. Trogocytosis of peptide-MHC class II complexes from dendritic cells confers antigen-presenting ability on basophils. Proc. Natl. Acad. Sci. USA 2017, 114, 1111–1116. [Google Scholar]
- Oliphant, C.J.; Hwang, Y.Y.; Walker, J.A.; Salimi, M.; Wong, S.H.; Brewer, J.M.; Englezakis, A.; Barlow, J.L.; Hams, E.; Scanlon, S.T.; et al. MHCII-mediated dialog between group 2 innate lymphoid cells and CD4(+) T cells potentiates type 2 immunity and promotes parasitic helminth expulsion. Immunity 2014, 41, 283–295. [Google Scholar] [PubMed] [Green Version]
- Dubrot, J.; Duraes, F.V.; Potin, L.; Capotosti, F.; Brighouse, D.; Suter, T.; Gut-Landmann, S.L.; Garbi, N.; Reith, W.; Swartz, M.A.; et al. Lymph node stromal cells acquire peptide-MHCII complexes from dendritic cells and induce antigen-specific CD4(+) T cell tolerance. J. Exp. Med. 2014, 211, 1153–1166. [Google Scholar] [PubMed]
- Rafii, A.; Mirshahi, P.; Poupot, M.; Faussat, A.M.; Simon, A.; Ducros, E.; Mery, E.; Couderc, B.; Lis, R.; Capdet, J.; et al. Oncologic trogocytosis of an original stromal cells induces chemoresistance of ovarian tumours. PLoS ONE 2008, 3, e3894. [Google Scholar]
- Weinhard, L.; di Bartolomei, G.; Bolasco, G.; Machado, P.; Schieber, N.L.; Neniskyte, U.; Exiga, M.; Vadisiute, A.; Raggioli, A.; Schertel, A.; et al. Microglia remodel synapses by presynaptic trogocytosis and spine head filopodia induction. Nat. Commun. 2018, 9, 1228. [Google Scholar]
- Andoh, M.; Shibata, K.; Okamoto, K.; Onodera, J.; Morishita, K.; Miura, Y.; Ikegaya, Y.; Koyama, R. Exercise reverses behavioral and synaptic abnormalities after maternal inflammation. Cell Rep. 2019, 27, 2817–2825. [Google Scholar]
- Lim, T.K.; Ruthazer, E.S. Microglial trogocytosis and the complement system regulate axonal pruning in vivo. eLife 2021, 10, e62167. [Google Scholar]
- Vanherberghen, B.; Andersson, K.; Carlin, L.M.; Nolte-’t Hoen, E.N.; Williams, G.S.; Hoglund, P.; Davis, D.M. Human and murine inhibitory natural killer cell receptors transfer from natural killer cells to target cells. Proc. Natl. Acad. Sci. USA 2004, 101, 16873–16878. [Google Scholar]
- Matlung, H.L.; Babes, L.; Zhao, X.W.; van Houdt, M.; Treffers, L.W.; van Rees, D.J.; Franke, K.; Schornagel, K.; Verkuijlen, P.; Janssen, H.; et al. Neutrophils kill antibody-opsonized cancer cells by trogoptosis. Cell Rep. 2018, 23, 3946–3959. [Google Scholar]
- Davis, D.M. Intercellular transfer of cell-surface proteins is common and can affect many stages of an immune response. Nat. Rev. Immunol. 2007, 7, 238–243. [Google Scholar]
- Ahmed, K.A.; Munegowda, M.A.; Xie, Y.; Xiang, J. Intercellular trogocytosis plays an important role in modulation of immune responses. Cell. Mol. Immunol. 2008, 5, 261–269. [Google Scholar]
- Rechavi, O.; Goldstein, I.; Kloog, Y. Intercellular exchange of proteins: The immune cell habit of sharing. FEBS Lett. 2009, 583, 1792–1799. [Google Scholar]
- Dhainaut, M.; Moser, M. Regulation of immune reactivity by intercellular transfer. Front. Immunol. 2014, 5, 112. [Google Scholar]
- Gilmartin, A.A.; Ralston, K.S.; Petri, W.A., Jr. Inhibition of amebic lysosomal acidification blocks amebic trogocytosis and cell killing. mBio 2017, 8, e01117–e01187. [Google Scholar]
- Gilmartin, A.A.; Ralston, K.S.; Petri, W.A. Inhibition of amebic cysteine proteases blocks amebic trogocytosis but not phagocytosis. J. Infect. Dis. 2020, 221, 1734–1739. [Google Scholar]
- Mercer, F.; Ng, S.H.; Brown, T.M.; Boatman, G.; Johnson, P.J. Neutrophils kill the parasite Trichomonas vaginalis using trogocytosis. PLoS Biol. 2018, 16, e2003885. [Google Scholar]
- Velmurugan, R.; Challa, D.K.; Ram, S.; Ober, R.J.; Ward, E.S. Macrophage-mediated trogocytosis leads to death of antibody-opsonized tumor cells. Mol. Cancer Ther. 2016, 15, 1879–1889. [Google Scholar]
- Bouti, P.; Zhao, X.W.; Verkuijlen, P.; Tool, A.T.J.; van Houdt, M.; Koker, N.; Koker, M.Y.; Keskin, O.; Akbayram, S.; van Bruggen, R.; et al. Kindlin3-dependent CD11b/CD18-integrin activation is required for potentiation of neutrophil cytotoxicity by CD47-SIRPα checkpoint disruption. Cancer Immunol. Res. 2021, 9, 147–155. [Google Scholar]
- Olivera-Valle, I.; Latorre, M.C.; Calvo, M.; Gaspar, B.; Gomez-Oro, C.; Collazos, A.; Breton, A.; Caballero-Campo, P.; Ardoy, M.; Asensio, F.; et al. Vaginal neutrophils eliminate sperm by trogocytosis. Hum. Reprod. 2020, 35, 2567–2578. [Google Scholar]
- Dustin, M.L.; Choudhuri, K. Signaling and polarized communication across the T cell immunological synapse. Annu. Rev. Cell Dev. Biol. 2016, 32, 303–325. [Google Scholar]
- Huang, J.F.; Yang, Y.; Sepulveda, H.; Shi, W.; Hwang, I.; Peterson, P.A.; Jackson, M.R.; Sprent, J.; Cai, Z. TCR-mediated internalization of peptide-MHC complexes acquired by T cells. Science 1999, 286, 952–954. [Google Scholar]
- Hudrisier, D.; Riond, J.; Mazarguil, H.; Gairin, J.E.; Joly, E. Cutting edge: CTLs rapidly capture membrane fragments from target cells in a TCR signaling-dependent manner. J. Immunol. 2001, 166, 3645–3649. [Google Scholar] [PubMed] [Green Version]
- Wetzel, S.A.; McKeithan, T.W.; Parker, D.C. Peptide-specific intercellular transfer of MHC class II to CD4+ T cells directly from the immunological synapse upon cellular dissociation. J. Immunol. 2005, 174, 80–89. [Google Scholar] [PubMed] [Green Version]
- Aucher, A.; Magdeleine, E.; Joly, E.; Hudrisier, D. Capture of plasma membrane fragments from target cells by trogocytosis requires signaling in T cells but not in B cells. Blood 2008, 111, 5621–5628. [Google Scholar] [PubMed] [Green Version]
- Martinez-Martin, N.; Fernandez-Arenas, E.; Cemerski, S.; Delgado, P.; Turner, M.; Heuser, J.; Irvine, D.J.; Huang, B.; Bustelo, X.R.; Shaw, A.; et al. T cell receptor internalization from the immunological synapse is mediated by TC21 and RhoG GTPase-dependent phagocytosis. Immunity 2011, 35, 208–222. [Google Scholar]
- Hwang, I.; Huang, J.F.; Kishimoto, H.; Brunmark, A.; Peterson, P.A.; Jackson, M.R.; Surh, C.D.; Cai, Z.; Sprent, J. T cells can use either T cell receptor or CD28 receptors to absorb and internalize cell surface molecules derived from antigen-presenting cells. J. Exp. Med. 2000, 191, 1137–1148. [Google Scholar]
- Sabzevari, H.; Kantor, J.; Jaigirdar, A.; Tagaya, Y.; Naramura, M.; Hodge, J.; Bernon, J.; Schlom, J. Acquisition of CD80 (B7-1) by T cells. J. Immunol. 2001, 166, 2505–2513. [Google Scholar]
- Baba, E.; Takahashi, Y.; Lichtenfeld, J.; Tanaka, R.; Yoshida, A.; Sugamura, K.; Yamamoto, N.; Tanaka, Y. Functional CD4 T cells after intercellular molecular transfer of 0X40 ligand. J. Immunol. 2001, 167, 875–883. [Google Scholar]
- Hudrisier, D.; Aucher, A.; Puaux, A.L.; Bordier, C.; Joly, E. Capture of target cell membrane components via trogocytosis is triggered by a selected set of surface molecules on T or B cells. J. Immunol. 2007, 178, 3637–3647. [Google Scholar]
- Daubeuf, S.; Aucher, A.; Bordier, C.; Salles, A.; Serre, L.; Gaibelet, G.; Faye, J.C.; Favre, G.; Joly, E.; Hudrisier, D. Preferential transfer of certain plasma membrane proteins onto T and B cells by trogocytosis. PLoS ONE 2010, 5, e8716. [Google Scholar]
- Xiang, J.; Huang, H.; Liu, Y. A new dynamic model of CD8+ T effector cell responses via CD4+ T helper-antigen-presenting cells. J. Immunol. 2005, 174, 7497–7505. [Google Scholar]
- He, T.; Zong, S.; Wu, X.; Wei, Y.; Xiang, J. CD4+ T cell acquisition of the bystander pMHC I colocalizing in the same immunological synapse comprising pMHC II and costimulatory CD40, CD54, CD80, OX40L, and 41BBL. Biochem. Biophys. Res. Commun. 2007, 362, 822–828. [Google Scholar]
- Ahmed, K.A.; Xie, Y.; Zhang, X.; Xiang, J. Acquired pMHC I complexes greatly enhance CD4(+) Th cell’s stimulatory effect on CD8(+) T cell-mediated diabetes in transgenic RIP-mOVA mice. Cell. Mol. Immunol. 2008, 5, 407–415. [Google Scholar]
- Romagnoli, P.A.; Premenko-Lanier, M.F.; Loria, G.D.; Altman, J.D. CD8 T cell memory recall is enhanced by novel direct interactions with CD4 T cells enabled by MHC class II transferred from APCs. PLoS ONE 2013, 8, e56999. [Google Scholar]
- Delgado, P.; Cubelos, B.; Calleja, E.; Martinez-Martin, N.; Cipres, A.; Merida, I.; Bellas, C.; Bustelo, X.R.; Alarcon, B. Essential function for the GTPase TC21 in homeostatic antigen receptor signaling. Nat. Immunol. 2009, 10, 880–888. [Google Scholar]
- Zhou, J.; Tagaya, Y.; Tolouei-Semnani, R.; Schlom, J.; Sabzevari, H. Physiological relevance of antigen presentasome (APS), an acquired MHC/costimulatory complex, in the sustained activation of CD4+ T cells in the absence of APCs. Blood 2005, 105, 3238–3246. [Google Scholar]
- Osborne, D.G.; Wetzel, S.A. Trogocytosis results in sustained intracellular signaling in CD4(+) T cells. J. Immunol. 2012, 189, 4728–4739. [Google Scholar]
- Reed, J.; Wetzel, S.A. Trogocytosis-mediated intracellular signaling in CD4(+) T cells drives TH2-associated effector cytokine production and differentiation. J. Immunol. 2019, 202, 2873–2887. [Google Scholar]
- Yamane, H.; Paul, W.E. Early signaling events that underlie fate decisions of naive CD4(+) T cells toward distinct T-helper cell subsets. Immunol. Rev. 2013, 252, 12–23. [Google Scholar]
- Boccasavia, V.L.; Bovolenta, E.R.; Villanueva, A.; Borroto, A.; Oeste, C.L.; van Santen, H.M.; Prieto, C.; Alonso-Lopez, D.; Diaz-Munoz, M.D.; Batista, F.D.; et al. Antigen presentation between T cells drives Th17 polarization under conditions of limiting antigen. Cell Rep. 2021, 34, 108861. [Google Scholar]
- Tsang, J.Y.; Chai, J.G.; Lechler, R. Antigen presentation by mouse CD4+ T cells involving acquired MHC class II:peptide complexes: Another mechanism to limit clonal expansion? Blood 2003, 101, 2704–2710. [Google Scholar]
- Game, D.S.; Rogers, N.J.; Lechler, R.I. Acquisition of HLA-DR and costimulatory molecules by T cells from allogeneic antigen presenting cells. Am. J. Transplant. 2005, 5, 1614–1625. [Google Scholar] [PubMed]
- Helft, J.; Jacquet, A.; Joncker, N.T.; Grandjean, I.; Dorothee, G.; Kissenpfennig, A.; Malissen, B.; Matzinger, P.; Lantz, O. Antigen-specific T-T interactions regulate CD4 T-cell expansion. Blood 2008, 112, 1249–1258. [Google Scholar] [PubMed]
- Zhou, G.; Ding, Z.C.; Fu, J.; Levitsky, H.I. Presentation of acquired peptide-MHC class II ligands by CD4+ regulatory T cells or helper cells differentially regulates antigen-specific CD4+ T cell response. J. Immunol. 2011, 186, 2148–2155. [Google Scholar]
- Akkaya, B.; Oya, Y.; Akkaya, M.; Souz, J.A.; Holstein, A.H.; Kamenyeva, O.; Kabat, J.; Matsumura, R.; Dorward, D.W.; Glass, D.D.; et al. Regulatory T cells mediate specific suppression by depleting peptide-MHC class II from dendritic cells. Nat. Immunol. 2019, 20, 218–231. [Google Scholar]
- Gu, P.; Gao, J.F.; D’Souza, C.A.; Kowalczyk, A.; Chou, K.Y.; Zhang, L. Trogocytosis of CD80 and CD86 by induced regulatory T cells. Cell. Mol. Immunol. 2012, 9, 136–146. [Google Scholar] [PubMed]
- Qureshi, O.S.; Zheng, Y.; Nakamura, K.; Attridge, K.; Manzotti, C.; Schmidt, E.M.; Baker, J.; Jeffery, L.E.; Kaur, S.; Briggs, Z.; et al. Trans-endocytosis of CD80 and CD86: A molecular basis for the cell-extrinsic function of CTLA-4. Science 2011, 332, 600–603. [Google Scholar] [PubMed] [Green Version]
- Ovcinnikovs, V.; Ross, E.M.; Petersone, L.; Edner, N.M.; Heuts, F.; Ntavli, E.; Kogimtzis, A.; Kennedy, A.; Wang, C.J.; Bennett, C.L.; et al. CTLA-4-mediated transendocytosis of costimulatory molecules primarily targets migratory dendritic cells. Sci. Immunol. 2019, 4, eaaw0902. [Google Scholar]
- Tekguc, M.; Wing, J.B.; Osaki, M.; Long, J.; Sakaguchi, S. CTLA-4-dependent trogocytosis promotes the interactions between Tregs and antigen-presenting cells. J. Immunol. 2020, 204, 228.8. [Google Scholar]
- Dhainaut, M.; Coquerelle, C.; Uzureau, S.; Denoeud, J.; Acolty, V.; Oldenhove, G.; Galuppo, A.; Sparwasser, T.; Thielemans, K.; Pays, E.; et al. Thymus-derived regulatory T cells restrain pro-inflammatory Th1 responses by downregulating CD70 on dendritic cells. EMBO J. 2015, 34, 1336–1348. [Google Scholar]
- Luu, K.; Patwardhan, M.V.; Zeng, Q.; Wickstrom, S.L.; Lundqvist, A.; Schwarz, H. Regulatory T cells inhibit T cell activity by downregulating CD137 ligand via CD137 trogocytosis. Cells 2021, 10, 353. [Google Scholar]
- Hudrisier, D.; Riond, J.; Garidou, L.; Duthoit, C.; Joly, E. T cell activation correlates with an increased proportion of antigen among the materials acquired from target cells. Eur. J. Immunol. 2005, 35, 2284–2294. [Google Scholar]
- Hamieh, M.; Dobrin, A.; Cabriolu, A.; van der Stegen, S.J.C.; Giavridis, T.; Mansilla-Soto, J.; Eyquem, J.; Zhao, Z.; Whitlock, B.M.; Miele, M.M.; et al. CAR T cell trogocytosis and cooperative killing regulate tumour antigen escape. Nature 2019, 568, 112–116. [Google Scholar]
- Son, J.M.; Ha, S.J. Extrinsic acquisition of CD80 by antigen-specific CD8(+) T cells regulates their recall immune responses to acute viral infection. Immune Netw. 2019, 19, e25. [Google Scholar]
- Riond, J.; Elhmouzi, J.; Hudrisier, D.; Gairin, J.E. Capture of membrane components via trogocytosis occurs in vivo during both dendritic cells and target cells encounter by CD8(+) T cells. Scand. J. Immunol. 2007, 66, 441–450. [Google Scholar]
- Machlenkin, A.; Uzana, R.; Frankenburg, S.; Eisenberg, G.; Eisenbach, L.; Pitcovski, J.; Gorodetsky, R.; Nissan, A.; Peretz, T.; Lotem, M. Capture of tumor cell membranes by trogocytosis facilitates detection and isolation of tumor-specific functional CTLs. Cancer Res. 2008, 68, 2006–2013. [Google Scholar] [PubMed] [Green Version]
- Kedl, R.M.; Schaefer, B.C.; Kappler, J.W.; Marrack, P. T cells down-modulate peptide-MHC complexes on APCs in vivo. Nat. Immunol. 2002, 3, 27–32. [Google Scholar] [PubMed]
- Chung, B.; Stuge, T.B.; Murad, J.P.; Beilhack, G.; Andersen, E.; Armstrong, B.D.; Weber, J.S.; Lee, P.P. Antigen-specific inhibition of high-avidity T cell target lysis by low-avidity T cells via trogocytosis. Cell Rep. 2014, 8, 871–882. [Google Scholar] [PubMed] [Green Version]
- LeMaoult, J.; Caumartin, J.; Daouya, M.; Favier, B.; Le Rond, S.; Gonzalez, A.; Carosella, E.D. Immune regulation by pretenders: Cell-to-cell transfers of HLA-G make effector T cells act as regulatory cells. Blood 2007, 109, 2040–2048. [Google Scholar]
- Brown, R.; Kabani, K.; Favaloro, J.; Yang, S.; Ho, P.J.; Gibson, J.; Fromm, P.; Suen, H.; Woodland, N.; Nassif, N.; et al. CD86+ or HLA-G+ can be transferred via trogocytosis from myeloma cells to T cells and are associated with poor prognosis. Blood 2012, 120, 2055–2063. [Google Scholar]
- Brown, R.; Suen, H.; Favaloro, J.; Yang, S.; Ho, P.J.; Gibson, J.; Joshua, D. Trogocytosis generates acquired regulatory T cells adding further complexity to the dysfunctional immune response in multiple myeloma. Oncoimmunology 2012, 1, 1658–1660. [Google Scholar]
- June, C.H.; O’Connor, R.S.; Kawalekar, O.U.; Ghassemi, S.; Milone, M.C. CAR T cell immunotherapy for human cancer. Science 2018, 359, 1361–1365. [Google Scholar]
- Brown, C.E.; Mackall, C.L. CAR T cell therapy: Inroads to response and resistance. Nat. Rev. Immunol. 2019, 19, 73–74. [Google Scholar]
- Hafeez, U.; Gan, H.K.; Scott, A.M. Monoclonal antibodies as immunomodulatory therapy against cancer and autoimmune diseases. Curr. Opin. Pharmacol. 2018, 41, 114–121. [Google Scholar]
- Xu, H.; Heyman, B. IgG-mediated suppression of antibody responses: Hiding or snatching epitopes? Scand. J. Immunol. 2020, 92, e12921. [Google Scholar]
- Beum, P.V.; Kennedy, A.D.; Williams, M.E.; Lindorfer, M.A.; Taylor, R.P. The shaving reaction: Rituximab/CD20 complexes are removed from mantle cell lymphoma and chronic lymphocytic leukemia cells by THP-1 monocytes. J. Immunol. 2006, 176, 2600–2609. [Google Scholar]
- Beum, P.V.; Peek, E.M.; Lindorfer, M.A.; Beurskens, F.J.; Engelberts, P.J.; Parren, P.W.; van de Winkel, J.G.; Taylor, R.P. Loss of CD20 and bound CD20 antibody from opsonized B cells occurs more rapidly because of trogocytosis mediated by Fc receptor-expressing effector cells than direct internalization by the B cells. J. Immunol. 2011, 187, 3438–3447. [Google Scholar]
- Iwasaki, S.; Masuda, S.; Baba, T.; Tomaru, U.; Katsumata, K.; Kasahara, M.; Ishizu, A. Plasma-dependent, antibody- and Fcgamma receptor-mediated translocation of CD8 molecules from T cells to monocytes. Cytom. Part A 2011, 79, 46–56. [Google Scholar]
- Masuda, S.; Iwasaki, S.; Tomaru, U.; Sato, J.; Kawakami, A.; Ichijo, K.; Sogo, S.; Baba, T.; Katsumata, K.; Kasahara, M.; et al. Mechanism of Fcgamma receptor-mediated trogocytosis-based false-positive results in flow cytometry. PLoS ONE 2012, 7, e52918. [Google Scholar]
- Rossi, E.A.; Goldenberg, D.M.; Michel, R.; Rossi, D.L.; Wallace, D.J.; Chang, C.H. Trogocytosis of multiple B-cell surface markers by CD22 targeting with epratuzumab. Blood 2013, 122, 3020–3029. [Google Scholar]
- Jones, J.D.; Hamilton, B.J.; Rigby, W.F. Rituximab mediates loss of CD19 on B cells in the absence of cell death. Arthritis Rheum. 2012, 64, 3111–3118. [Google Scholar]
- Skopelja-Gardner, S.; Jones, J.D.; Hamilton, B.J.; Danilov, A.V.; Rigby, W.F.C. Role for ZAP-70 signaling in the differential effector functions of rituximab and obinutuzumab (GA101) in chronic lymphocytic leukemia B cells. J. Immunol. 2017, 199, 1275–1282. [Google Scholar] [PubMed] [Green Version]
- Beum, P.V.; Mack, D.A.; Pawluczkowycz, A.W.; Lindorfer, M.A.; Taylor, R.P. Binding of rituximab, trastuzumab, cetuximab, or mAb T101 to cancer cells promotes trogocytosis mediated by THP-1 cells and monocytes. J. Immunol. 2008, 181, 8120–8132. [Google Scholar] [PubMed] [Green Version]
- Valgardsdottir, R.; Cattaneo, I.; Klein, C.; Introna, M.; Figliuzzi, M.; Golay, J. Human neutrophils mediate trogocytosis rather than phagocytosis of CLL B cells opsonized with anti-CD20 antibodies. Blood 2017, 129, 2636–2644. [Google Scholar] [PubMed] [Green Version]
- Zhang, Y.; McClellan, M.; Efros, L.; Shi, D.; Bielekova, B.; Tang, M.T.; Vexler, V.; Sheridan, J.P. Daclizumab reduces CD25 levels on T cells through monocyte-mediated trogocytosis. Mult. Scler. J. 2014, 20, 156–164. [Google Scholar]
- Suzuki, E.; Kataoka, T.R.; Hirata, M.; Kawaguchi, K.; Nishie, M.; Haga, H.; Toi, M. Trogocytosis-mediated expression of HER2 on immune cells may be associated with a pathological complete response to trastuzumab-based primary systemic therapy in HER2-overexpressing breast cancer patients. BMC Cancer 2015, 15, 39. [Google Scholar]
- Krejcik, J.; Frerichs, K.A.; Nijhof, I.S.; van Kessel, B.; van Velzen, J.F.; Bloem, A.C.; Broekmans, M.E.C.; Zweegman, S.; van Meerloo, J.; Musters, R.J.P.; et al. Monocytes and granulocytes reduce CD38 expression levels on myeloma cells in patients treated with daratumumab. Clin. Cancer Res. 2017, 23, 7498–7511. [Google Scholar]
- Beurskens, F.J.; Lindorfer, M.A.; Farooqui, M.; Beum, P.V.; Engelberts, P.; Mackus, W.J.; Parren, P.W.; Wiestner, A.; Taylor, R.P. Exhaustion of cytotoxic effector systems may limit monoclonal antibody-based immunotherapy in cancer patients. J. Immunol. 2012, 188, 3532–3541. [Google Scholar]
- Baig, N.A.; Taylor, R.P.; Lindorfer, M.A.; Church, A.K.; LaPlant, B.R.; Pettinger, A.M.; Shanafelt, T.D.; Nowakowski, G.S.; Zent, C.S. Induced resistance to ofatumumab-mediated cell clearance mechanisms, including complement-dependent cytotoxicity, in chronic lymphocytic leukemia. J. Immunol. 2014, 192, 1620–1629. [Google Scholar]
- Williams, M.E.; Densmore, J.J.; Pawluczkowycz, A.W.; Beum, P.V.; Kennedy, A.D.; Lindorfer, M.A.; Hamil, S.H.; Eggleton, J.C.; Taylor, R.P. Thrice-weekly low-dose rituximab decreases CD20 loss via shaving and promotes enhanced targeting in chronic lymphocytic leukemia. J. Immunol. 2006, 177, 7435–7443. [Google Scholar]
- Pinney, J.J.; Rivera-Escalera, F.; Chu, C.C.; Whitehead, H.E.; VanDerMeid, K.R.; Nelson, A.M.; Barbeau, M.C.; Zent, C.S.; Elliott, M.R. Macrophage hypophagia as a mechanism of innate immune exhaustion in mAb-induced cell clearance. Blood 2020, 136, 2065–2079. [Google Scholar]
- Carlin, L.M.; Eleme, K.; McCann, F.E.; Davis, D.M. Intercellular transfer and supramolecular organization of human leukocyte antigen C at inhibitory natural killer cell immune synapses. J. Exp. Med. 2001, 194, 1507–1517. [Google Scholar] [PubMed] [Green Version]
- Sjöström, A.; Eriksson, M.; Cerboni, C.; Johansson, M.H.; Sentman, C.L.; Kärre, K.; Höglund, P. Acquisition of external major histocompatibility complex class I molecules by natural killer cells expressing inhibitory Ly49 receptors. J. Exp. Med. 2001, 194, 1519–1530. [Google Scholar]
- Zimmer, J.; Ioannidis, V.; Held, W. H-2D ligand expression by Ly49A+ natural killer (NK) cells precludes ligand uptake from environmental cells: Implications for NK cell function. J. Exp. Med. 2001, 194, 1531–1539. [Google Scholar] [PubMed]
- Tabiasco, J.; Espinosa, E.; Hudrisier, D.; Joly, E.; Fournié, J.J.; Vercellone, A. Active trans-synaptic capture of membrane fragments by natural killer cells. Eur. J. Immunol. 2002, 32, 1502–1508. [Google Scholar] [PubMed]
- Caumartin, J.; Favier, B.; Daouya, M.; Guillard, C.; Moreau, P.; Carosella, E.D.; LeMaoult, J. Trogocytosis-based generation of suppressive NK cells. EMBO J. 2007, 26, 1423–1433. [Google Scholar] [PubMed] [Green Version]
- Nakamura, K.; Nakayama, M.; Kawano, M.; Amagai, R.; Ishii, T.; Harigae, H.; Ogasawara, K. Fratricide of natural killer cells dressed with tumor-derived NKG2D ligand. Proc. Natl. Acad. Sci. USA 2013, 110, 9421–9426. [Google Scholar]
- Miner, C.A.; Giri, T.K.; Meyer, C.E.; Shabsovich, M.; Tripathy, S.K. Acquisition of activation receptor ligand by trogocytosis renders NK cells hyporesponsive. J. Immunol. 2015, 194, 1945–1953. [Google Scholar] [PubMed] [Green Version]
- Dolan, B.P.; Gibbs, K.D., Jr.; Ostrand-Rosenberg, S. Dendritic cells cross-dressed with peptide MHC class I complexes prime CD8+ T cells. J. Immunol. 2006, 177, 6018–6024. [Google Scholar]
- Wakim, L.M.; Bevan, M.J. Cross-dressed dendritic cells drive memory CD8+ T-cell activation after viral infection. Nature 2011, 471, 629–632. [Google Scholar]
- Hughes, A.D.; Zhao, D.; Dai, H.; Abou-Daya, K.I.; Tieu, R.; Rammal, R.; Williams, A.L.; Landsittel, D.P.; Shlomchik, W.D.; Morelli, A.E.; et al. Cross-dressed dendritic cells sustain effector T cell responses in islet and kidney allografts. J. Clin. Investig. 2020, 130, 287–294. [Google Scholar]
- Nakayama, M.; Takeda, K.; Kawano, M.; Takai, T.; Ishii, N.; Ogasawara, K. Natural killer (NK)-dendritic cell interactions generate MHC class II-dressed NK cells that regulate CD4+ T cells. Proc. Natl. Acad. Sci. USA 2011, 108, 18360–18365. [Google Scholar]
- Embgenbroich, M.; Burgdorf, S. Current concepts of antigen cross-presentation. Front. Immunol. 2018, 9, 1643. [Google Scholar]
- Blander, J.M. Regulation of the cell biology of antigen cross-presentation. Annu. Rev. Immunol. 2018, 36, 717–753. [Google Scholar]
- Li, B.; Lu, C.; Oveissi, S.; Song, J.; Xiao, K.; Zanker, D.; Duan, M.; Chen, J.; Xu, H.; Zou, Q.; et al. Host CD8α(+) and CD103(+) dendritic cells prime transplant antigen-specific CD8(+) T cells via cross-dressing. Immunol. Cell Biol. 2020, 98, 563–576. [Google Scholar]
- Kambayashi, T.; Laufer, T.M. Atypical MHC class II-expressing antigen-presenting cells: Can anything replace a dendritic cell? Nat. Rev. Immunol. 2014, 14, 719–730. [Google Scholar]
- Lin, A.; Loré, K. Granulocytes: New members of the antigen-presenting cell family. Front. Immunol. 2017, 8, 1781. [Google Scholar]
- Schuijs, M.J.; Hammad, H.; Lambrecht, B.N. Professional and ‘amateur’ antigen-presenting cells in type 2 immunity. Trends Immunol. 2019, 40, 22–34. [Google Scholar]
- Voehringer, D. Recent advances in understanding basophil functions in vivo. F1000Research 2017, 6, 1464. [Google Scholar]
- Varricchi, G.; Raap, U.; Rivellese, F.; Marone, G.; Gibbs, B.F. Human mast cells and basophils—How are they similar how are they different? Immunol. Rev. 2018, 282, 8–34. [Google Scholar] [PubMed]
- Nakashima, C.; Otsuka, A.; Kabashima, K. Recent advancement in the mechanism of basophil activation. J. Dermatol. Sci. 2018, 91, 3–8. [Google Scholar]
- Miyake, K.; Shibata, S.; Yoshikawa, S.; Karasuyama, H. Basophils and their effector molecules in allergic disorders. Allergy 2020. [Google Scholar] [CrossRef]
- Yamanishi, Y.; Karasuyama, H. Basophil-derived IL-4 plays versatile roles in immunity. Semin. Immunopathol. 2016, 38, 615–622. [Google Scholar] [PubMed]
- Sokol, C.L.; Barton, G.M.; Farr, A.G.; Medzhitov, R. A mechanism for the initiation of allergen-induced T helper type 2 responses. Nat. Immunol. 2008, 9, 310–318. [Google Scholar]
- Sokol, C.L.; Chu, N.Q.; Yu, S.; Nish, S.A.; Laufer, T.M.; Medzhitov, R. Basophils function as antigen-presenting cells for an allergen-induced T helper type 2 response. Nat. Immunol. 2009, 10, 713–720. [Google Scholar]
- Perrigoue, J.G.; Saenz, S.A.; Siracusa, M.C.; Allenspach, E.J.; Taylor, B.C.; Giacomin, P.R.; Nair, M.G.; Du, Y.; Zaph, C.; van Rooijen, N. MHC class II-dependent basophil-CD4+ T cell interactions promote TH2 cytokine-dependent immunity. Nat. Immunol. 2009, 10, 697–705. [Google Scholar]
- Yoshimoto, T.; Yasuda, K.; Tanaka, H.; Nakahira, M.; Imai, Y.; Fujimori, Y.; Nakanishi, K. Basophils contribute to TH2-IgE responses in vivo via IL-4 production and presentation of peptide-MHC class II complexes to CD4+ T cells. Nat. Immunol. 2009, 10, 706–712. [Google Scholar]
- Hammad, H.; Plantinga, M.; Deswarte, K.; Pouliot, P.; Willart, M.A.; Kool, M.; Muskens, F.; Lambrecht, B.N. Inflammatory dendritic cells—Not basophils—Are necessary and sufficient for induction of Th2 immunity to inhaled house dust mite allergen. J. Exp. Med. 2010, 207, 2097–2111. [Google Scholar]
- Tang, H.; Cao, W.; Kasturi, S.P.; Ravindran, R.; Nakaya, H.I.; Kundu, K.; Murthy, N.; Kepler, T.B.; Malissen, B.; Pulendran, B. The T helper type 2 response to cysteine proteases requires dendritic cell-basophil cooperation via ROS-mediated signaling. Nat. Immunol. 2010, 11, 608–617. [Google Scholar]
- Phythian-Adams, A.T.; Cook, P.C.; Lundie, R.J.; Jones, L.H.; Smith, K.A.; Barr, T.A.; Hochweller, K.; Anderton, S.M.; Hammerling, G.J.; Maizels, R.M.; et al. CD11c depletion severely disrupts Th2 induction and development in vivo. J. Exp. Med. 2010, 207, 2089–2096. [Google Scholar]
- Dudeck, J.; Medyukhina, A.; Frobel, J.; Svensson, C.M.; Kotrba, J.; Gerlach, M.; Gradtke, A.C.; Schroder, B.; Speier, S.; Figge, M.T.; et al. Mast cells acquire MHCII from dendritic cells during skin inflammation. J. Exp. Med. 2017, 214, 3791–3811. [Google Scholar]
- Carroll-Portillo, A.; Cannon, J.L.; Riet, J.T.; Holmes, A.; Kawakami, Y.; Kawakami, T.; Cambi, A.; Lidke, D.S. Mast cells and dendritic cells form synapses that facilitate antigen transfer for T cell activation. J. Cell Biol. 2015, 210, 851–864. [Google Scholar]
- Sumpter, T.L.; Balmert, S.C.; Kaplan, D.H. Cutaneous immune responses mediated by dendritic cells and mast cells. JCI Insight 2019, 4, e123947. [Google Scholar]
- Steele, S.; Radlinski, L.; Taft-Benz, S.; Brunton, J.; Kawula, T.H. Trogocytosis-associated cell to cell spread of intracellular bacterial pathogens. eLife 2016, 5, e10625. [Google Scholar]
- Steele, S.P.; Chamberlain, Z.; Park, J.; Kawula, T.H. Francisella tularensis enters a double membraned compartment following cell-cell transfer. eLife 2019, 8, e45252. [Google Scholar] [PubMed]
- Cambier, C.J.; O’Leary, S.M.; O’Sullivan, M.P.; Keane, J.; Ramakrishnan, L. Phenolic glycolipid facilitates mycobacterial escape from microbicidal tissue-resident macrophages. Immunity 2017, 47, 552–565.e4. [Google Scholar] [PubMed] [Green Version]
- Perez, O.A.; Yeung, S.T.; Vera-Licona, P.; Romagnoli, P.A.; Samji, T.; Ural, B.B.; Maher, L.; Tanaka, M.; Khanna, K.M. CD169(+) macrophages orchestrate innate immune responses by regulating bacterial localization in the spleen. Sci. Immunol. 2017, 2, eaah5520. [Google Scholar] [PubMed] [Green Version]
- Dragotakes, Q.; Fu, M.S.; Casadevall, A. Dragotcytosis: Elucidation of the mechanism for Cryptococcus neoformans macrophage-to-macrophage transfer. J. Immunol. 2019, 202, 2661–2670. [Google Scholar] [PubMed]
- Lersritwimanmaen, P.; Na-Ek, P.; Thanunchai, M.; Thewsoongnoen, J.; Sa-Ard-Iam, N.; Wiboon-ut, S.; Mahanonda, R.; Thitithanyanont, A. The presence of monocytes enhances the susceptibility of B cells to highly pathogenic avian influenza (HPAI) H5N1 virus possibly through the increased expression of α2,3 SA receptor. Biochem. Biophys. Res. Commun. 2015, 464, 888–893. [Google Scholar]
- Kongsomros, S.; Thanunchai, M.; Manopwisedjaroen, S.; Na-Ek, P.; Wang, S.F.; Taechalertpaisarn, T.; Thitithanyanont, A. Trogocytosis with monocytes associated with increased α2,3 sialic acid expression on B cells during H5N1 influenza virus infection. PLoS ONE 2020, 15, e0239488. [Google Scholar]
- Tabiasco, J.; Vercellone, A.; Meggetto, F.; Hudrisier, D.; Brousset, P.; Fournié, J.J. Acquisition of viral receptor by NK cells through immunological synapse. J. Immunol. 2003, 170, 5993–5998. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyake, K.; Karasuyama, H. The Role of Trogocytosis in the Modulation of Immune Cell Functions. Cells 2021, 10, 1255. https://doi.org/10.3390/cells10051255
Miyake K, Karasuyama H. The Role of Trogocytosis in the Modulation of Immune Cell Functions. Cells. 2021; 10(5):1255. https://doi.org/10.3390/cells10051255
Chicago/Turabian StyleMiyake, Kensuke, and Hajime Karasuyama. 2021. "The Role of Trogocytosis in the Modulation of Immune Cell Functions" Cells 10, no. 5: 1255. https://doi.org/10.3390/cells10051255
APA StyleMiyake, K., & Karasuyama, H. (2021). The Role of Trogocytosis in the Modulation of Immune Cell Functions. Cells, 10(5), 1255. https://doi.org/10.3390/cells10051255