
Siglec Ligands


Abstract
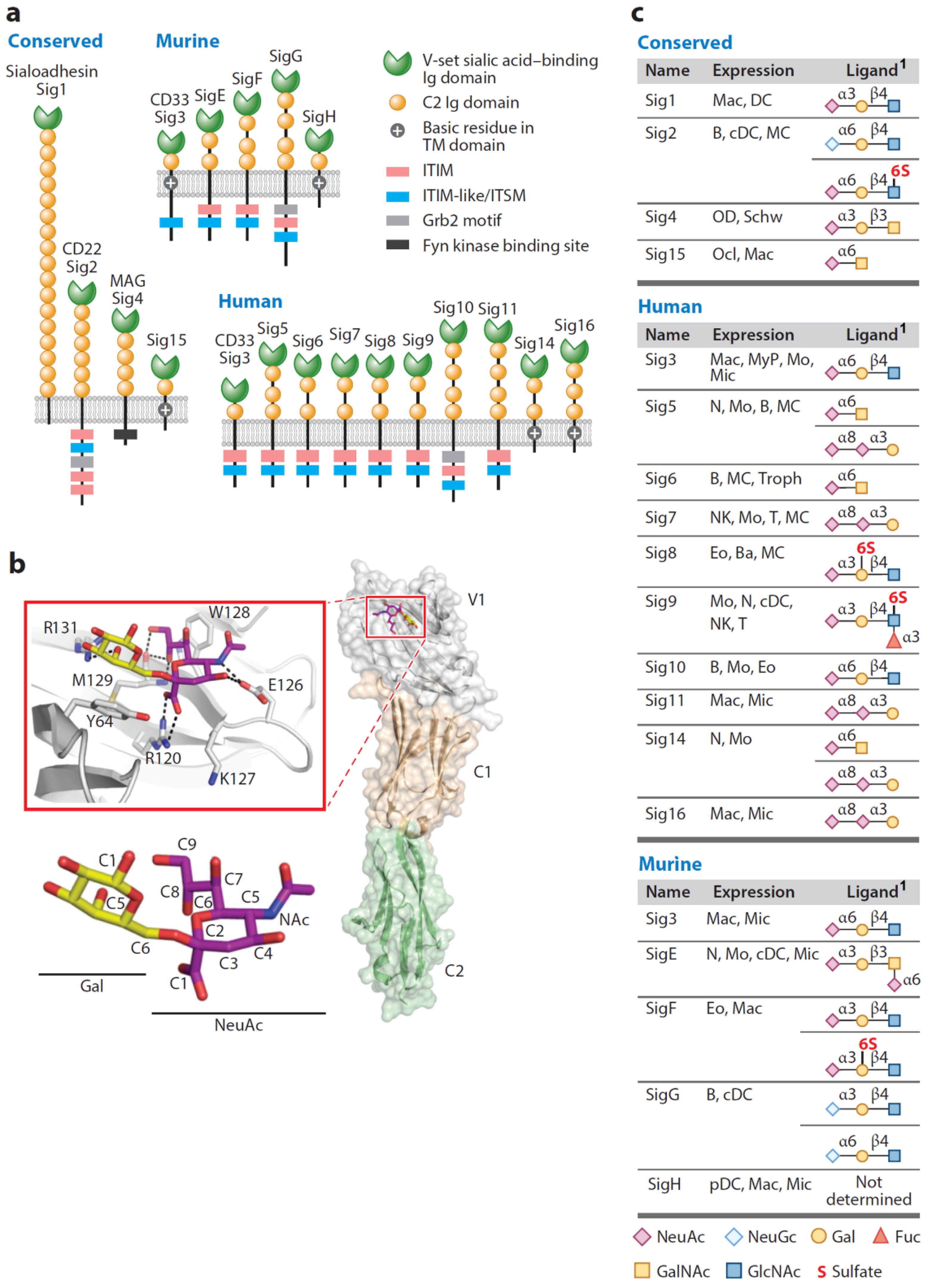
:1. Introduction
2. Experimental Approaches to Identify Siglec Ligands
2.1. Siglec-Based Probes
2.2. Glycan Arrays
2.3. Sialomimetic Design and Screening
2.4. Cell-Based Binding Assays
2.5. Native Siglec Ligand Identification
3. Human Siglec Ligands
3.1. Siglec-4 (Myelin Associated Glycoprotein, MAG)
3.2. Siglec-2 (CD22)
3.3. Siglec-7
3.4. Siglec-8
3.5. Siglec-9
3.6. Siglec-1 (Sialoadhesin, CD169)
3.7. Siglec-3 (CD33)
3.8. Siglec-11
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duan, S.; Paulson, J.C. Siglecs as Immune Cell Checkpoints in Disease. Annu. Rev. Immunol. 2020, 38, 365–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugesan, G.; Weigle, B.; Crocker, P.R. Siglec and anti-Siglec therapies. Curr. Opin. Chem. Biol. 2021, 62, 34–42. [Google Scholar] [CrossRef]
- Läubli, H.; Varki, A. Sialic acid–binding immunoglobulin-like lectins (Siglecs) detect self-associated molecular patterns to regulate immune responses. Cell. Mol. Life Sci. 2020, 77, 593–605. [Google Scholar] [CrossRef]
- Movsisyan, L.D.; Macauley, M.S. Structural advances of Siglecs: Insight into synthetic glycan ligands for immunomodulation. Org. Biomol. Chem. 2020, 18, 5784–5797. [Google Scholar] [CrossRef]
- Lenza, M.P.; Atxabal, U.; Oyenarte, I.; Jiménez-Barbero, J.; Ereño-Orbea, J. Current Status on Therapeutic Molecules Targeting Siglec Receptors. Cells 2020, 9, 2691. [Google Scholar] [CrossRef]
- Manni, M.; Läubli, H. Targeting glyco-immune checkpoints for cancer therapy. Expert Opin. Biol. Ther. 2021, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Crocker, P.R.; Clark, E.A.; Filbin, M.; Gordon, S.; Jones, Y.; Kehrl, J.H.; Kelm, S.; Le Douarin, N.; Powell, L.; Roder, J.; et al. Siglecs: A family of sialic-acid binding lectins. Glycobiology 1998, 8, 5–6. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Schnaar, R.L.; Schauer, R. Sialic Acids and Other Nonulosonic Acids. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2017; pp. 179–195. [Google Scholar]
- O’Reilly, M.K.; Paulson, J.C. Multivalent Ligands for Siglecs. Methods Enzymol. 2010, 478, 343–363. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.Y.; Heimburg-Molinaro, J.; Cummings, R.D.; Goth, C.K. Emerging patterns of tyrosine sulfation and O-glycosylation cross-talk and co-localization. Curr. Opin. Struct. Biol. 2020, 62, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Büll, C.; Heise, T.; Adema, G.J.; Boltje, T.J. Sialic Acid Mimetics to Target the Sialic Acid–Siglec Axis. Trends Biochem. Sci. 2016, 41, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Angata, T.; Margulies, E.H.; Green, E.D.; Varki, A. Large-scale sequencing of the CD33-related Siglec gene cluster in five mammalian species reveals rapid evolution by multiple mechanisms. Proc. Natl. Acad. Sci. USA 2004, 101, 13251–13256. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, S.S.; Vaill, M.; Do, R.; Khan, N.; Verhagen, A.L.; Zhang, W.; Lenz, H.; Johnson-Pais, T.L.; Leach, R.J.; Fraser, G.; et al. Human-specific polymorphic pseudogenization of SIGLEC12 protects against advanced cancer progression. FASEB Bioadv. 2021, 3, 69–82. [Google Scholar] [CrossRef]
- Bhattacherjee, A.; Rodrigues, E.; Jung, J.; Luzentales-Simpson, M.; Enterina, J.R.; Galleguillos, D.; Laurent, C.D.S.; Nakhaei-Nejad, M.; Fuchsberger, F.F.; Streith, L.; et al. Repression of phagocytosis by human CD33 is not conserved with mouse CD33. Commun. Biol. 2019, 2, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angata, T. Possible Influences of Endogenous and Exogenous Ligands on the Evolution of Human Siglecs. Front. Immunol. 2018, 9, 2885. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Crocker, P.R. Evolution of CD33-related siglecs: Regulating host immune functions and escaping pathogen exploitation? Immunology 2010, 132, 18–26. [Google Scholar] [CrossRef]
- Varki, A. Nothing in Glycobiology Makes Sense, except in the Light of Evolution. Cell 2006, 126, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, E.; Jung, J.; Park, H.; Loo, C.; Soukhtehzari, S.; Kitova, E.N.; Mozaneh, F.; Daskhan, G.; Schmidt, E.N.; Aghanya, V.; et al. A versatile soluble siglec scaffold for sensitive and quantitative detection of glycan ligands. Nat. Commun. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, A.; Porell, R.N.; Fernandes, S.M.; Wei, Y.; Yu, H.; Carroll, D.J.; McBride, R.; Paulson, J.C.; Tiemeyer, M.; Aoki, K.; et al. Sialylated keratan sulfate proteoglycans are Siglec-8 ligands in human airways. Glycobiology 2018, 28, 786–801. [Google Scholar] [CrossRef]
- Li, Z.; Feizi, T. The neoglycolipid (NGL) technology-based microarrays and future prospects. FEBS Lett. 2018, 592, 3976–3991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuillan, A.M.; Byrd-Leotis, L.; Heimburg-Molinaro, J.; Cummings, R.D. Natural and Synthetic Sialylated Glycan Microarrays and Their Applications. Front. Mol. Biosci. 2019, 6, 88. [Google Scholar] [CrossRef] [Green Version]
- Rillahan, C.D.; Schwartz, E.; McBride, R.; Fokin, V.V.; Paulson, J.C. Click and Pick: Identification of Sialoside Analogues for Siglec-Based Cell Targeting. Angew. Chem. Int. Ed. 2012, 51, 11014–11018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.H.; Hsu, C.H.; Wu, C.Y. Sialoside Arrays: New Synthetic Strategies and Applications. Top. Curr. Chem. 2015, 367, 125–149. [Google Scholar]
- Blixt, O.; Head, S.; Mondala, T.; Scanlan, C.; Huflejt, M.E.; Alvarez, R.; Bryan, M.C.; Fazio, F.; Calarese, D.; Stevens, J.; et al. Printed covalent glycan array for ligand profiling of diverse glycan binding proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 17033–17038. [Google Scholar] [CrossRef] [Green Version]
- Heimburg-Molinaro, J.; Song, X.; Smith, D.F.; Cummings, R.D. Preparation and Analysis of Glycan Microarrays. Curr. Protoc. Protein Sci. 2011, 64, 12.10.1–12.10.29. [Google Scholar] [CrossRef] [Green Version]
- Cummings, R.D. The repertoire of glycan determinants in the human glycome. Mol. BioSyst. 2009, 5, 1087–1104. [Google Scholar] [CrossRef]
- Dam, T.K.; Brewer, C.F. Multivalent lectin-carbohydrate interactions energetics and mechanisms of binding. Adv. Carbohydr. Chem. Biochem. 2010, 63, 139–164. [Google Scholar] [PubMed]
- Nycholat, C.M.; Rademacher, C.; Kawasaki, N.; Paulson, J.C. In Silico-Aided Design of a Glycan Ligand of Sialoadhesin for in Vivo Targeting of Macrophages. J. Am. Chem. Soc. 2012, 134, 15696–15699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwardt, O.; Kelm, S.; Ernst, B. SIGLEC-4 (MAG) Antagonists: From the Natural Carbohydrate Epitope to Glycomimetics. Top. Curr. Chem. 2013, 367, 151–200. [Google Scholar] [CrossRef]
- Rillahan, C.D.; Schwartz, E.; Rademacher, C.; McBride, R.; Rangarajan, J.; Fokin, V.V.; Paulson, J.C. On-Chip Synthesis and Screening of a Sialoside Library Yields a High Affinity Ligand for Siglec-7. ACS Chem. Biol. 2013, 8, 1417–1422. [Google Scholar] [CrossRef] [Green Version]
- Nycholat, C.M.; Duan, S.; Knuplez, E.; Worth, C.; Elich, M.; Yao, A.; O’Sullivan, J.; McBride, R.; Wei, Y.; Fernandes, S.M.; et al. A Sulfonamide Sialoside Analogue for Targeting Siglec-8 and -F on Immune Cells. J. Am. Chem. Soc. 2019, 141, 14032–14037. [Google Scholar] [CrossRef]
- Mesch, S.; Lemme, K.; Wittwer, M.; Koliwer-Brandl, H.; Schwardt, O.; Kelm, S.; Ernst, B. From a Library of MAG Antagonists to Nanomolar CD22 Ligands. ChemMedChem 2011, 7, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Kelm, S.; Pelz, A.; Schauer, R.; Filbin, M.T.; Tang, S.; de Bellard, M.-E.; Schnaar, R.L.; Mahoney, J.A.; Hartnell, A.; Bradfield, P.; et al. Sialoadhesin, myelin-associated glycoprotein and CD22 define a new family of sialic acid-dependent adhesion molecules of the immunoglobulin superfamily. Curr. Biol. 1994, 4, 965–972. [Google Scholar] [CrossRef]
- Freeman, S.; Kelm, S.; Barber, E.; Crocker, P. Characterization of CD33 as a new member of the sialoadhesin family of cellular interaction molecules. Blood 1995, 85, 2005–2012. [Google Scholar] [CrossRef] [Green Version]
- Cornish, A.L.; Freeman, S.; Forbes, G.; Ni, J.; Zhang, M.; Cepeda, M.; Gentz, R.; Augustus, M.; Carter, K.C.; Crocker, P.R. Characterization of siglec-5, a novel glycoprotein expressed on myeloid cells related to CD33. Blood 1998, 92, 2123–2132. [Google Scholar] [CrossRef]
- Nicoll, G.; Ni, J.; Liu, D.; Klenerman, P.; Munday, J.; Dubock, S.; Mattei, M.-G.; Crocker, P.R. Identification and Characterization of a Novel Siglec, Siglec-7, Expressed by Human Natural Killer Cells and Monocytes. J. Biol. Chem. 1999, 274, 34089–34095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floyd, H.; Ni, J.; Cornish, A.L.; Zeng, Z.; Liu, D.; Carter, K.C.; Steel, J.; Crocker, P.R. Siglec-8. A novel eosinophil-specific member of the immunoglobulin superfamily. J. Biol. Chem. 2000, 275, 861–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angata, T.; Varki, A. Cloning, characterization and phylogenetic analysis of Siglec-9, a new member of the CD33-related group of Siglecs. Evidence for co-evolution with sialic acid synthesis pathways. J. Biol. Chem. 2000, 275, 22127–22135. [Google Scholar] [CrossRef] [Green Version]
- Munday, J.; Kerr, S.; Ni, J.; Cornish, A.L.; Zhang, J.Q.; Nicoll, G.; Floyd, H.; Mattei, M.G.; Moore, P.; Liu, D.; et al. Identification, characterization and leucocyte expression of Siglec-10, a novel human sialic acid-binding receptor. Biochem. J. 2001, 355, 489–497. [Google Scholar] [CrossRef]
- Paulson, J.C.; Rogers, G.N. Resialylated eythrocytes for assessment of the specificity of sialyloligosaccharide binding proteins. Methods Enzymol. 1987, 138, 162–168. [Google Scholar]
- Narimatsu, Y.; Büll, C.; Chen, Y.-H.; Wandall, H.H.; Yang, Z.; Clausen, H. Genetic glycoengineering in mammalian cells. J. Biol. Chem. 2021, 296, 100448. [Google Scholar] [CrossRef]
- Narimatsu, Y.; Joshi, H.J.; Nason, R.; Van Coillie, J.; Karlsson, R.; Sun, L.; Ye, Z.; Chen, Y.-H.; Schjoldager, K.T.; Steentoft, C.; et al. An Atlas of Human Glycosylation Pathways Enables Display of the Human Glycome by Gene Engineered Cells. Mol. Cell 2019, 75, 394–407. [Google Scholar] [CrossRef] [PubMed]
- Wisnovsky, S.; Möckl, L.; Malaker, S.A.; Pedram, K.; Hess, G.T.; Riley, N.M.; Gray, M.A.; Smith, B.A.H.; Bassik, M.C.; Moerner, W.E.; et al. Genome-wide CRISPR screens reveal a specific ligand for the glycan-binding immune checkpoint receptor Siglec-7. Proc. Natl. Acad. Sci. USA 2021, 118, e2015024118. [Google Scholar] [CrossRef] [PubMed]
- Briard, J.G.; Jiang, H.; Moremen, K.W.; Macauley, M.S.; Wu, P. Cell-based glycan arrays for probing glycan–glycan binding protein interactions. Nat. Commun. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Büll, C.; Nason, R.; Sun, L.; Van Coillie, J.; Sørensen, D.M.; Moons, S.J.; Yang, Z.; Arbitman, S.; Fernandes, S.M.; Furukawa, S.; et al. Probing the binding specificities of human Siglecs by cell-based glycan arrays. Proc. Natl. Acad. Sci. USA 2021, 118, e2026102118. [Google Scholar] [CrossRef]
- Hart, G.W.; Varki, A. Future Directions in Glycosciences. In Essentials of Glycobiology, 3rd ed.; Varki, A., Cummings, R.D., Esko, J.D., Stanley, P., Hart, G.W., Aebi, M., Darvill, A.G., Kinoshita, T., Packer, N.H., Prestegard, J.H., et al., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015; pp. 761–768. [Google Scholar]
- Quarles, R.H. Myelin-associated glycoprotein (MAG): Past, present and beyond. J. Neurochem. 2007, 100, 1431–1448. [Google Scholar] [CrossRef] [PubMed]
- Bjartmar, C.; Yin, X.; Trapp, B.D. Axonal pathology in myelin disorders. J. Neurocytol. 1999, 28, 383–395. [Google Scholar] [CrossRef]
- Arquint, M.; Roder, J.; Chia, L.S.; Down, J.; Wilkinson, D.; Bayley, H.; Braun, P.; Dunn, R. Molecular cloning and primary structure of myelin-associated glycoprotein. Proc. Natl. Acad. Sci. USA 1987, 84, 600–604. [Google Scholar] [CrossRef] [Green Version]
- Schnaar, R.L.; Gerardy-Schahn, R.; Hildebrandt, H. Sialic Acids in the Brain: Gangliosides and Polysialic Acid in Nervous System Development, Stability, Disease, and Regeneration. Physiol. Rev. 2014, 94, 461–518. [Google Scholar] [CrossRef] [Green Version]
- Schnaar, R.L. Gangliosides of the Vertebrate Nervous System. J. Mol. Biol. 2016, 428, 3325–3336. [Google Scholar] [CrossRef] [Green Version]
- Chiavegatto, S.; Sun, J.; Nelson, R.J.; Schnaar, R.L. A Functional Role for Complex Gangliosides: Motor Deficits in GM2/GD2 Synthase Knockout Mice. Exp. Neurol. 2000, 166, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Pan, B.; Fromholt, S.E.; Hess, E.J.; Crawford, T.O.; Griffin, J.W.; Sheikh, K.A.; Schnaar, R.L. Myelin-associated glycoprotein and complementary axonal ligands, gangliosides, mediate axon stability in the CNS and PNS: Neuropathology and behavioral deficits in single- and double-null mice. Exp. Neurol. 2005, 195, 208–217. [Google Scholar] [CrossRef] [Green Version]
- Boukhris, A.; Schüle-Freyer, R.; Loureiro, J.L.; Lourenço, C.M.; Mundwiller, E.; Gonzalez, M.A.; Charles, P.; Gauthier, J.; Rekik, I.; Lebrigio, R.F.A.; et al. Alteration of Ganglioside Biosynthesis Responsible for Complex Hereditary Spastic Paraplegia. Am. J. Hum. Genet. 2013, 93, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Roubertie, A.; Charif, M.; Meyer, P.; Manes, G.; Meunier, I.; Taieb, G.; Morales, R.J.; Guichet, A.; Delettre, C.; Sarzi, E.; et al. Hereditary spastic paraplegia and prominent sensorial involvement: Think MAG mutations! Ann. Clin. Transl. Neurol. 2019, 6, 1572–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roda, R.H.; FitzGibbon, E.J.; Boucekkine, H.; Schindler, A.B.; Blackstone, C. Neurologic syndrome associated with homozygous mutation at MAG sialic acid binding site. Ann. Clin. Transl. Neurol. 2016, 3, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.E.; Yang, L.J.-S.; Mukhopadhyay, G.; Filbin, M.T.; Kiso, M.; Hasegawa, A.; Schnaar, R.L. Sialic Acid Specificity of Myelin-associated Glycoprotein Binding. J. Biol. Chem. 1997, 272, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.E.; Ito, H.; Sawada, N.; Ishida, H.; Kiso, M.; Schnaar, R.L. Enhanced Binding of the Neural Siglecs, Myelin-associated Glycoprotein and Schwann Cell Myelin Protein, to Chol-1 (α-Series) Gangliosides and Novel Sulfated Chol-1 Analogs. J. Biol. Chem. 1999, 274, 37637–37643. [Google Scholar] [CrossRef] [Green Version]
- Swanson, B.J.; McDermott, K.M.; Singh, P.K.; Eggers, J.P.; Crocker, P.R.; Hollingsworth, M.A. MUC1 Is a Counter-Receptor for Myelin-Associated Glycoprotein (Siglec-4a) and Their Interaction Contributes to Adhesion in Pancreatic Cancer Perineural Invasion. Cancer Res. 2007, 67, 10222–10229. [Google Scholar] [CrossRef] [Green Version]
- McKerracher, L.; Rosen, K.M. MAG, myelin and overcoming growth inhibition in the CNS. Front. Mol. Neurosci. 2015, 8, 51. [Google Scholar] [CrossRef] [Green Version]
- Vyas, A.A.; Patel, H.V.; Fromholt, S.E.; Heffer-Lauc, M.; Vyas, K.A.; Dang, J.; Schachner, M.; Schnaar, R.L. Gangliosides are functional nerve cell ligands for myelin-associated glycoprotein (MAG), an inhibitor of nerve regeneration. Proc. Natl. Acad. Sci. USA 2002, 99, 8412–8417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelke, S.V.; Gao, G.-P.; Mesch, S.; Gäthje, H.; Kelm, S.; Schwardt, O.; Ernst, B. Synthesis of sialic acid derivatives as ligands for the myelin-associated glycoprotein (MAG). Bioorg. Med. Chem. 2007, 15, 4951–4965. [Google Scholar] [CrossRef]
- Zeng, Y.; Rademacher, C.; Nycholat, C.M.; Futakawa, S.; Lemme, K.; Ernst, B.; Paulson, J.C. High affinity sialoside ligands of myelin associated glycoprotein. Bioorg. Med. Chem. Lett. 2011, 21, 5045–5049. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.E.; Blixt, O.; Han, S.; Duong, B.; Li, H.; Nathan, J.K.; Bovin, N.; Paulson, J.C. High-affinity ligand probes of CD22 overcome the threshold set by cis ligands to allow for binding, endocytosis, and killing of B cells. J. Immunol. 2006, 177, 2994–3003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rillahan, C.D.; Macauley, M.; Schwartz, E.; He, Y.; McBride, R.; Arlian, B.M.; Rangarajan, J.; Fokin, V.V.; Paulson, J.C. Disubstituted sialic acid ligands targeting siglecs CD33 and CD22 associated with myeloid leukaemias and B cell lymphomas. Chem. Sci. 2014, 5, 2398–2406. [Google Scholar] [CrossRef] [Green Version]
- Prescher, H.; Schweizer, A.; Kuhfeldt, E.; Nitschke, L.; Brossmer, R. Discovery of Multifold Modified Sialosides as Human CD22/Siglec-2 Ligands with Nanomolar Activity on B-Cells. ACS Chem. Biol. 2014, 9, 1444–1450. [Google Scholar] [CrossRef] [PubMed]
- Prescher, H.; Frank, M.; Gütgemann, S.; Kuhfeldt, E.; Schweizer, A.; Nitschke, L.; Watzl, C.; Brossmer, R. Design, Synthesis, and Biological Evaluation of Small, High-Affinity Siglec-7 Ligands: Toward Novel Inhibitors of Cancer Immune Evasion. J. Med. Chem. 2017, 60, 941–956. [Google Scholar] [CrossRef] [PubMed]
- Kroezen, B.S.; Conti, G.; Girardi, B.; Cramer, J.; Jiang, X.H.; Rabbani, S.; Muller, J.; Kokot, M.; Luisoni, E.; Ricklin, D.; et al. A Potent Mimetic of the Siglec-8 Ligand 6′-Sulfo-Sialyl Lewis(x). Chemmedchem 2020, 15, 1706–1719. [Google Scholar] [CrossRef]
- Bednar, K.J.; Shanina, E.; Ballet, R.; Connors, E.P.; Duan, S.; Juan, J.; Arlian, B.M.; Kulis, M.D.; Butcher, E.C.; Fung-Leung, W.-P.; et al. Human CD22 Inhibits Murine B Cell Receptor Activation in a Human CD22 Transgenic Mouse Model. J. Immunol. 2017, 199, 3116–3128. [Google Scholar] [CrossRef]
- Nitschke, L.; Tsubata, T. Molecular interactions regulate BCR signal inhibition by CD22 and CD72. Trends Immunol. 2004, 25, 543–550. [Google Scholar] [CrossRef]
- Walker, J.A.; Smith, K.G.C. CD22: An inhibitory enigma. Immunology 2008, 123, 314–325. [Google Scholar] [CrossRef]
- Müller, J.; Obermeier, I.; Wöhner, M.; Brandl, C.; Mrotzek, S.; Angermüller, S.; Maity, P.C.; Reth, M.; Nitschke, L. CD22 ligand-binding and signaling domains reciprocally regulate B-cell Ca2+ signaling. Proc. Natl. Acad. Sci. USA 2013, 110, 12402–12407. [Google Scholar] [CrossRef] [Green Version]
- Macauley, M.; Kawasaki, N.; Peng, W.; Wang, S.-H.; He, Y.; Arlian, B.M.; McBride, R.; Kannagi, R.; Khoo, K.-H.; Paulson, J.C. Unmasking of CD22 Co-receptor on Germinal Center B-cells Occurs by Alternative Mechanisms in Mouse and Man. J. Biol. Chem. 2015, 290, 30066–30077. [Google Scholar] [CrossRef] [Green Version]
- Ereño-Orbea, J.; Sicard, T.; Cui, H.; Mazhab-Jafari, M.T.; Benlekbir, S.; Guarné, A.; Rubinstein, J.L.; Julien, J.-P. Molecular basis of human CD22 function and therapeutic targeting. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Hennet, T.; Chui, D.; Paulson, J.C.; Marth, J.D. Immune regulation by the ST6Gal sialyltransferase. Proc. Natl. Acad. Sci. USA 1998, 95, 4504–4509. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.E.; Smith, B.A.; Bengtson, P.; Paulson, J.C. Ablation of CD22 in ligand-deficient mice restores B cell receptor signaling. Nat. Immunol. 2005, 7, 199–206. [Google Scholar] [CrossRef]
- Han, S.; Collins, B.E.; Bengtson, P.; Paulson, J.C. Homomultimeric complexes of CD22 in B cells revealed by protein-glycan cross-linking. Nat. Chem. Biol. 2005, 1, 93–97. [Google Scholar] [CrossRef]
- Sheikh, A.A.D.; Akatsu, C.; Imamura, A.; Abdu-Allah, H.H.; Takematsu, H.; Ando, H.; Ishida, H.; Tsubata, T. Proximity labeling of cis-ligands of CD22/Siglec-2 reveals stepwise α2,6 sialic acid-dependent and -independent interactions. Biochem. Biophys. Res. Commun. 2018, 495, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Enterina, J.R.; Jung, J.; Macauley, M.S. Coordinated roles for glycans in regulating the inhibitory function of CD22 on B cells. Biomed. J. 2019, 42, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, F.; Feest, C.; Bruckbauer, A.; Mattila, P.K.; Müller, J.; Nitschke, L.; Bray, D.; Batista, F.D. Nanoscale organization and dynamics of the siglec CD 22 cooperate with the cytoskeleton in restraining BCR signalling. EMBO J. 2015, 35, 258–280. [Google Scholar] [CrossRef] [Green Version]
- Ramya, T.N.; Weerapana, E.; Liao, L.; Zeng, Y.; Tateno, H.; Liao, L.; Yates, J.R.; Cravatt, B.F.; Paulson, J.C. In situ trans ligands of CD22 identified by glycan-protein photocross-linking-enabled proteomics. Mol. Cell. Proteom. 2010, 9, 1339–1351. [Google Scholar] [CrossRef] [Green Version]
- Macauley, M.; Pfrengle, F.; Rademacher, C.; Nycholat, C.M.; Gale, A.J.; von Drygalski, A.; Paulson, J.C. Antigenic liposomes displaying CD22 ligands induce antigen-specific B cell apoptosis. J. Clin. Investig. 2013, 123, 3074–3083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bednar, K.J.; Nycholat, C.M.; Rao, T.S.; Paulson, J.C.; Fung-Leung, W.-P.; Macauley, M.S.; Rao, T.S. Exploiting CD22 To Selectively Tolerize Autoantibody Producing B-Cells in Rheumatoid Arthritis. ACS Chem. Biol. 2019, 14, 644–654. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Paulson, J.C. CD22 Ligands on a NaturalN-Glycan Scaffold Efficiently Deliver Toxins to B-Lymphoma Cells. J. Am. Chem. Soc. 2017, 139, 12450–12458. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.G.M.; Keating, N.; Nicholson, S.E. Natural Killer Cells: Tumor Surveillance and Signaling. Cancers 2020, 12, 952. [Google Scholar] [CrossRef] [Green Version]
- Daly, J.; Carlsten, M.; O’Dwyer, M. Sugar Free: Novel Immunotherapeutic Approaches Targeting Siglecs and Sialic Acids to Enhance Natural Killer Cell Cytotoxicity Against Cancer. Front. Immunol. 2019, 10, 1047. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Handa, K.; Withers, D.A.; Satoh, M.; Hakomori, S. Binding specificity of siglec7 to disialogangliosides of renal cell carcinoma: Possible role of disialogangliosides in tumor progression. FEBS Lett. 2001, 504, 82–86. [Google Scholar] [CrossRef]
- Blixt, O.; Collins, B.E.; van den Nieuwenhof, I.; Crocker, P.R.; Paulson, J.C. Sialoside specificity of the siglec family assessed using novel multivalent probes: Identification of potent inhibitors of myelin-associated glycoprotein. J. Biol. Chem. 2003, 278, 31007–31019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, T.; Teranishi, T.; Alphey, M.S.; Crocker, P.R.; Hashimoto, Y. A Small Region of the Natural Killer Cell Receptor, Siglec-7, Is Responsible for Its Preferred Binding to α2,8-Disialyl and Branched α2,6-Sialyl Residues. J. Biol. Chem. 2002, 277, 6324–6332. [Google Scholar] [CrossRef] [Green Version]
- Nicoll, G.; Avril, T.; Lock, K.; Furukawa, K.; Bovin, N.; Crocker, P.R. Ganglioside GD3 expression on target cells can modulate NK cell cytotoxicity via siglec-7-dependent and -independent mechanisms. Eur. J. Immunol. 2003, 33, 1642–1648. [Google Scholar] [CrossRef]
- Attrill, H.; Imamura, A.; Sharma, R.S.; Kiso, M.; Crocker, P.R.; van Aalten, D.M.F. Siglec-7 Undergoes a Major Conformational Change When Complexed with the α(2,8)-Disialylganglioside GT1b. J. Biol. Chem. 2006, 281, 32774–32783. [Google Scholar] [CrossRef] [Green Version]
- Campanero-Rhodes, M.A.; Childs, R.A.; Kiso, M.; Komba, S.; Le Narvor, C.; Warren, J.; Otto, D.; Crocker, P.R.; Feizi, T. Carbohydrate microarrays reveal sulphation as a modulator of siglec binding. Biochem. Biophys. Res. Commun. 2006, 344, 1141–1146. [Google Scholar] [CrossRef]
- Yoshimura, A.; Asahina, Y.; Chang, L.-Y.; Angata, T.; Tanaka, H.; Kitajima, K.; Sato, C. Identification and functional characterization of a Siglec-7 counter-receptor on K562 cells. J. Biol. Chem. 2021, 296, 100477. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Yoshimura, A.; Yasuda, Y.; Mori, A.; Tanaka, H.; Takahashi, T.; Kitajima, K.; Sato, C. Chemical Synthesis and Evaluation of a Disialic Acid-Containing Dextran Polymer as an Inhibitor for the Interaction between Siglec 7 and Its Ligand. ChemBioChem 2017, 18, 1194–1203. [Google Scholar] [CrossRef] [PubMed]
- Kikly, K.K.; Bochner, B.S.; Freeman, S.D.; Tan, K.; Gallagher, K.T.; D’Alessio, K.J.; Holmes, S.D.; Abrahamson, J.A.; Erickson-Miller, C.L.; Murdock, P.R.; et al. Identification of SAF-2, a novel siglec expressed on eosinophils, mast cells, and basophils. J. Allergy Clin. Immunol. 2000, 105, 1093–1100. [Google Scholar] [CrossRef]
- Youngblood, B.A.; Leung, J.; Falahati, R.; Williams, J.; Schanin, J.; Brock, E.C.; Singh, B.; Chang, A.T.; O’Sullivan, J.A.; Schleimer, R.P.; et al. Discovery, Function, and Therapeutic Targeting of Siglec-8. Cells 2020, 10, 19. [Google Scholar] [CrossRef]
- Nutku-Bilir, E.; Hudson, S.A.; Bochner, B.S. Interleukin-5 Priming of Human Eosinophils Alters Siglec-8–Mediated Apoptosis Pathways. Am. J. Respir. Cell Mol. Biol. 2008, 38, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Yokoi, H.; Choi, O.H.; Hubbard, W.; Lee, H.S.; Canning, B.J.; Lee, H.H.; Ryu, S.D.; von Gunten, S.; Bickel, C.A.; Hudson, S.A.; et al. Inhibition of FcepsilonRI-dependent mediator release and calcium flux from human mast cells by sialic acid-binding immunoglobulin-like lectin 8 engagement. J. Allergy Clin. Immunol. 2008, 121, 499–505. [Google Scholar] [CrossRef]
- Duan, S.; Arlian, B.M.; Nycholat, C.M.; Wei, Y.; Tateno, H.; Smith, S.A.; Macauley, M.S.; Zhu, Z.; Bochner, B.S.; Paulson, J.C. Nanoparticles Displaying Allergen and Siglec-8 Ligands Suppress IgE-FcepsilonRI-Mediated Anaphylaxis and Desensitize Mast Cells to Subsequent Antigen Challenge. J. Immunol. 2021, 206, 2290–2300. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, J.A.; Chang, A.T.; Youngblood, B.A.; Bochner, B.S. Eosinophil and mast cell Siglecs: From biology to drug target. J. Leukoc. Biol. 2020, 108, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Bochner, B.S.; Alvarez, R.A.; Mehta, P.; Bovin, N.V.; Blixt, O.; White, J.R.; Schnaar, R.L. Glycan Array Screening Reveals a Candidate Ligand for Siglec-8. J. Biol. Chem. 2005, 280, 4307–4312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.; Gonzalez-Gil, A.; Wei, Y.; Fernandes, S.M.; Porell, R.N.; Vajn, K.; Paulson, J.C.; Nycholat, C.M.; Schnaar, R.L. Siglec-8 and Siglec-9 binding specificities and endogenous airway ligand distributions and properties. Glycobiology 2017, 27, 657–668. [Google Scholar] [CrossRef]
- Tateno, H.; Crocker, P.R.; Paulson, J.C. Mouse Siglec-F and human Siglec-8 are functionally convergent paralogs that are selectively expressed on eosinophils and recognize 6′-sulfo-sialyl Lewis X as a preferred glycan ligand. Glycobiology 2005, 15, 1125–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pröpster, J.M.; Yang, F.; Rabbani, S.; Ernst, B.; Allain, F.H.-T.; Schubert, M. Structural basis for sulfation-dependent self-glycan recognition by the human immune-inhibitory receptor Siglec-8. Proc. Natl. Acad. Sci. USA 2016, 113, E4170–E4179. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Angata, T.; Cho, J.Y.; Miller, M.; Broide, D.H.; Varki, A. Defining the in vivo function of Siglec-F, a CD33-related Siglec expressed on mouse eosinophils. Blood 2007, 109, 4280–4287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Yu, H.; Fernandes, S.M.; Wei, Y.; Gonzalez-Gil, A.; Motari, M.G.; Vajn, K.; Stevens, W.W.; Peters, A.T.; Bochner, B.S.; et al. Expression of ligands for Siglec-8 and Siglec-9 in human airways and airway cells. J. Allergy Clin. Immunol. 2015, 135, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Funderburgh, J.L. MINI REVIEW Keratan sulfate: Structure, biosynthesis, and function. Glycobiology 2000, 10, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, A.; Li, T.A.; Porell, R.N.; Fernandes, S.M.; Tarbox, H.E.; Lee, H.S.; Aoki, K.; Tiemeyer, M.; Kim, J.; Schnaar, R.L. Isolation, identification, and characterization of the human airway ligand for the eosinophil and mast cell immunoinhibitory receptor Siglec-8. J. Allergy Clin. Immunol. 2021, 147, 1442–1452. [Google Scholar] [CrossRef] [PubMed]
- Reichhardt, M.; Holmskov, U.; Meri, S. SALSA—A dance on a slippery floor with changing partners. Mol. Immunol. 2017, 89, 100–110. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Gonzalez-Gil, A.; Drake, V.; Li, T.A.; Schnaar, R.L.; Kim, J. Induction of the endogenous sialoglycan ligand for eosinophil death receptor Siglec-8 in chronic rhinosinusitis with hyperplastic nasal polyposis. Glycobiology 2021. [Google Scholar] [CrossRef]
- Zhang, J.Q.; Nicoll, G.; Jones, C.; Crocker, P.R. Siglec-9, a Novel Sialic Acid Binding Member of the Immunoglobulin Superfamily Expressed Broadly on Human Blood Leukocytes. J. Biol. Chem. 2000, 275, 22121–22126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaveris, C.S.; Chiu, S.H.; Riley, N.M.; Bertozzi, C.R. Modulation of immune cell reactivity with cis-binding Siglec agonists. Proc. Natl. Acad. Sci. USA 2021, 118, e2012408118. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, E.; Boelaars, K.; Brown, K.; Li, R.J.E.; Kruijssen, L.; Bruijns, S.C.M.; van Ee, T.; Schetters, S.T.T.; Crommentuijn, M.H.W.; van der Horst, J.C.; et al. Sialic acids in pancreatic cancer cells drive tumour-associated macrophage differentiation via the Siglec receptors Siglec-7 and Siglec-9. Nat. Commun. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Von Gunten, S.; Yousefi, S.; Seitz, M.; Jakob, S.M.; Schaffner, T.; Seger, R.; Takala, J.; Villiger, P.M.; Simon, H.-U. Siglec-9 transduces apoptotic and nonapoptotic death signals into neutrophils depending on the proinflammatory cytokine environment. Blood 2005, 106, 1423–1431. [Google Scholar] [CrossRef]
- Delaveris, C.S.; Wilk, A.J.; Riley, N.M.; Stark, J.C.; Yang, S.S.; Rogers, A.J.; Ranganath, T.; Nadeau, K.C.; Blish, C.A.; Bertozzi, C.R. Synthetic Siglec-9 Agonists Inhibit Neutrophil Activation Associated with COVID-19. ACS Cent. Sci. 2021, 7, 650–657. [Google Scholar] [CrossRef]
- Secundino, I.; Lizcano, A.; Roupé, K.M.; Wang, X.; Cole, J.N.; Olson, J.; Ali, S.R.; Dahesh, S.; Amayreh, L.K.; Henningham, A.; et al. Host and pathogen hyaluronan signal through human siglec-9 to suppress neutrophil activation. J. Mol. Med. 2015, 94, 219–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lizcano, A.; Secundino, I.; Döhrmann, S.; Corriden, R.; Rohena, C.; Diaz, S.; Ghosh, P.; Deng, L.; Nizet, V.; Varki, A. Erythrocyte sialoglycoproteins engage Siglec-9 on neutrophils to suppress activation. Blood 2017, 129, 3100–3110. [Google Scholar] [CrossRef] [Green Version]
- Kiser, Z.M.; Lizcano, A.; Nguyen, J.; Becker, G.L.; Belcher, J.D.; Varki, A.P.; Vercellotti, G.M. Decreased erythrocyte binding of Siglec-9 increases neutrophil activation in sickle cell disease. Blood Cells Mol. Dis. 2020, 81, 102399. [Google Scholar] [CrossRef]
- Zhang, Y.; Zheng, Y.; Li, J.; Nie, L.; Hu, Y.; Wang, F.; Liu, H.; Fernandes, S.M.; Zhong, Q.; Li, X.; et al. Immunoregulatory Siglec ligands are abundant in human and mouse aorta and are up-regulated by high glucose. Life Sci. 2019, 216, 189–199. [Google Scholar] [CrossRef]
- Pinho, S.S.; Reis, C.A. Glycosylation in cancer: Mechanisms and clinical implications. Nat. Rev. Cancer 2015, 15, 540–555. [Google Scholar] [CrossRef]
- Tanida, S.; Akita, K.; Ishida, A.; Mori, Y.; Toda, M.; Inoue, M.; Ohta, M.; Yashiro, M.; Sawada, T.; Hirakawa, K.; et al. Binding of the sialic acid-binding lectin, Siglec-9, to the membrane mucin, MUC1, induces recruitment of beta-catenin and subsequent cell growth. J. Biol. Chem. 2013, 288, 31842–31852. [Google Scholar] [CrossRef] [Green Version]
- Beatson, R.; Tajadura-Ortega, V.; Achkova, D.; Picco, G.; Tsourouktsoglou, T.-D.; Klausing, S.; Hillier, M.; Maher, D.A.J.; Noll, S.K.T.; Crocker, P.R.; et al. The mucin MUC1 modulates the tumor immunological microenvironment through engagement of the lectin Siglec-9. Nat. Immunol. 2016, 17, 1273–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belisle, J.A.; Horibata, S.; Gubbels, J.A.; Petrie, S.; Kapur, A.; Andre, S.; Gabius, H.-J.; Rancourt, C.; Connor, J.; Paulson, J.C.; et al. Identification of Siglec-9 as the receptor for MUC16 on human NK cells, B cells, and monocytes. Mol. Cancer 2010, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, A.S.G.; Berg, T.K.V.D.; Mullen, G.E.D. Sialoadhesin—A macrophage-restricted marker of immunoregulation and inflammation. Immunology 2013, 138, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Klaas, M.; Crocker, P.R. Sialoadhesin in recognition of self and non-self. Semin. Immunopathol. 2012, 34, 353–364. [Google Scholar] [CrossRef] [PubMed]
- der Linden, E.C.M.B.-V.; Varki, A. New aspects of siglec binding specificities, including the significance of fucosylation and of the sialyl-Tn epitope. Sialic acid-binding immunoglobulin superfamily lectins. J. Biol. Chem. 2000, 275, 8625–8632. [Google Scholar] [CrossRef] [Green Version]
- Heikema, A.P.; Bergman, M.P.; Richards, H.; Crocker, P.R.; Gilbert, M.; Samsom, J.N.; Van Wamel, W.J.B.; Endtz, H.P.; Van Belkum, A. Characterization of the Specific Interaction between Sialoadhesin and Sialylated Campylobacter jejuni Lipooligosaccharides. Infect. Immun. 2010, 78, 3237–3246. [Google Scholar] [CrossRef] [Green Version]
- Kidder, D.; Richards, H.E.; Ziltener, H.J.; Garden, O.A.; Crocker, P.R. Sialoadhesin Ligand Expression Identifies a Subset of CD4+Foxp3—T Cells with a Distinct Activation and Glycosylation Profile. J. Immunol. 2013, 190, 2593–2602. [Google Scholar] [CrossRef] [Green Version]
- May, A.; Robinson, R.; Vinson, M.; Crocker, P.; Jones, E. Crystal Structure of the N-Terminal Domain of Sialoadhesin in Complex with 3′ Sialyllactose at 1.85 Å Resolution. Mol. Cell 1998, 1, 719–728. [Google Scholar] [CrossRef]
- Berg, T.K.V.D.; Nath, D.; Ziltener, H.J.; Vestweber, D.; Fukuda, M.; Van Die, I.; Crocker, P.R. Cutting Edge: CD43 Functions as a T Cell Counterreceptor for the Macrophage Adhesion Receptor Sialoadhesin (Siglec-1). J. Immunol. 2001, 166, 3637–3640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nath, D.; Hartnell, A.; Happerfield, L.; Miles, D.W.; Burchell, J.; Taylor-Papadimitriou, J.; Crocker, P.R. Macrophage-tumour cell interactions: Identification of MUC1 on breast cancer cells as a potential counter-receptor for the macrophage-restricted receptor, sialoadhesin. Immunology 1999, 98, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Twilhaar, M.K.N.; Czentner, L.; Grabowska, J.; Affandi, A.J.; Lau, C.Y.J.; Olesek, K.; Kalay, H.; Van Nostrum, C.F.; Van Kooyk, Y.; Storm, G.; et al. Optimization of Liposomes for Antigen Targeting to Splenic CD169+ Macrophages. Pharmaceutics 2020, 12, 1138. [Google Scholar] [CrossRef] [PubMed]
- Blixt, O.; Han, S.; Liao, L.; Zeng, Y.; Hoffmann, J.; Futakawa, S.; Paulson, J.C. Sialoside Analogue Arrays for Rapid Identification of High Affinity Siglec Ligands. J. Am. Chem. Soc. 2008, 130, 6680–6681. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, N.; Vela, J.L.; Nycholat, C.M.; Rademacher, C.; Khurana, A.; van Rooijen, N.; Crocker, P.R.; Kronenberg, M.; Paulson, J.C. Targeted delivery of lipid antigen to macrophages via the CD169/sialoadhesin endocytic pathway induces robust invariant natural killer T cell activation. Proc. Natl. Acad. Sci. USA 2013, 110, 7826–7831. [Google Scholar] [CrossRef] [Green Version]
- Edgar, L.J.; Kawasaki, N.; Nycholat, C.M.; Paulson, J.C. Targeted Delivery of Antigen to Activated CD169+ Macrophages Induces Bias for Expansion of CD8+ T Cells. Cell Chem. Biol. 2019, 26, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Laszlo, G.S.; Estey, E.H.; Walter, R.B. The past and future of CD33 as therapeutic target in acute myeloid leukemia. Blood Rev. 2014, 28, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Jen, E.Y.; Ko, C.-W.; Lee, J.E.; Del Valle, P.L.; Aydanian, A.; Jewell, C.; Norsworthy, K.J.; Przepiorka, D.; Nie, L.; Liu, J.; et al. FDA Approval: Gemtuzumab Ozogamicin for the Treatment of Adults with Newly Diagnosed CD33-Positive Acute Myeloid Leukemia. Clin. Cancer Res. 2018, 24, 3242–3246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crocker, P.R.; Paulson, J.C.; Varki, A. Siglecs and their roles in the immune system. Nat. Rev. Immunol. 2007, 7, 255–266. [Google Scholar] [CrossRef] [PubMed]
- der Linden, E.C.M.B.-V.; Angata, T.; Reynolds, S.A.; Powell, L.D.; Hedrick, S.M.; Varki, A. CD33/Siglec-3 Binding Specificity, Expression Pattern, and Consequences of Gene Deletion in Mice. Mol. Cell. Biol. 2003, 23, 4199–4206. [Google Scholar] [CrossRef] [Green Version]
- Varki, A.; Angata, T. Siglecs—The major subfamily of I-type lectins. Glycobiology 2005, 16, 1R–27R. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, S.P.; Taylor, L.S.; Stansbury, E.K.; McVicar, D.W. Myeloid specific human CD33 is an inhibitory receptor with differential ITIM function in recruiting the phosphatases SHP-1 and SHP-2. Blood 2000, 96, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Miles, L.A.; Hermans, S.J.; Crespi, G.A.N.; Gooi, J.H.; Doughty, L.; Nero, T.L.; Markulic, J.; Ebneth, A.; Wroblowski, B.; Oehlrich, D.; et al. Small Molecule Binding to Alzheimer Risk Factor CD33 Promotes Abeta Phagocytosis. iScience 2019, 19, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.; Koziol-White, C.J.; Jester, W.F., Jr.; Smith, S.A.; Nycholat, C.M.; Macauley, M.S.; Panettieri, R.A., Jr.; Paulson, J.C. CD33 recruitment inhibits IgE-mediated anaphylaxis and desensitizes mast cells to allergen. J. Clin. Investig. 2019, 129, 1387–1401. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, J.A.; Wei, Y.; Carroll, D.J.; Moreno-Vinasco, L.; Cao, Y.; Zhang, F.; Lee, J.J.; Zhu, Z.; Bochner, B.S. Frontline Science: Characterization of a novel mouse strain expressing human Siglec-8 only on eosinophils. J. Leukoc. Biol. 2018, 104, 11–19. [Google Scholar] [CrossRef]
- Wei, Y.; Chhiba, K.D.; Zhang, F.; Ye, X.; Wang, L.; Zhang, L.; Robida, P.A.; Moreno-Vinasco, L.; Schnaar, R.L.; Roers, A.; et al. Mast Cell-Specific Expression of Human Siglec-8 in Conditional Knock-in Mice. Int. J. Mol. Sci. 2018, 20, 19. [Google Scholar] [CrossRef] [Green Version]
- Angata, T.; Kerr, S.C.; Greaves, D.R.; Varki, N.M.; Crocker, P.R.; Varki, A. Cloning and characterization of human Siglec-11: A recently evolved signaling molecule that can interact with SHP-1 and SHP-2 and is expressed by tissue macrophages, including brain microglia. J. Biol. Chem 2002, 277, 24466–24474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayakawa, T.; Angata, T.; Lewis, A.L.; Mikkelsen, T.S.; Varki, N.M.; Varki, A. A human-specific gene in microglia. Science 2005, 309, 1693. [Google Scholar]
- Wang, Y.; Neumann, H. Alleviation of Neurotoxicity by Microglial Human Siglec-11. J. Neurosci. 2010, 30, 3482–3488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahraz, A.; Kopatz, J.; Mathy, R.; Kappler, J.; Winter, M.; Kapoor, S.; Schütza, V.; Scheper, T.; Gieselmann, V.; Neumann, H. Anti-inflammatory activity of low molecular weight polysialic acid on human macrophages. Sci. Rep. 2015, 5, 16800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hane, M.; Chen, D.Y.; Varki, A. Human-specific microglial Siglec-11 transcript variant has the potential to affect polysialic acid-mediated brain functions at a distance. Glycobiology 2020, 31, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Angata, T.; Nycholat, C.M.; Macauley, M.S. Therapeutic Targeting of Siglecs using Antibody- and Glycan-Based Approaches. Trends Pharmacol. Sci. 2015, 36, 645–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Siglecs | Expression | Diseases |
|---|---|---|
| Siglec-1/ sialoadhesin/CD169 |
|
|
| Siglec-2/CD22 |
|
|
| Siglec-3/CD33 |
|
|
| Siglec-4/MAG |
|
|
| Siglec-5 |
|
|
| Siglec-6 |
|
|
| Siglec-7 |
|
|
| Siglec-8 |
|
|
| Siglec-9 |
|
|
| Siglec-10 |
|
|
| Siglec-11 |
|
|
| Siglec-14 |
|
|
| Siglec-15 |
|
|
| Siglec-16 |
|
| Siglec | Endogenous Ligands 1 | Sialomimetics | [References] |
|---|---|---|---|
| Siglec-1 | CD43S1L, PSGL-1S1L, MUC1S1L |  | [28] |
| Siglec-2 (CD22) | CD22, IgMS2L, CD45S2L |  | [64] |
 | [65] | ||
 | [66] | ||
| Siglec-3 (CD33) |  | [65] | |
| Siglec-4 (MAG) | GD1a, GT1b, GQ1bα |  | [32] |
 | [63] | ||
| Siglec-7 | CD43S7L, GD3 |  | [30] |
 | [67] | ||
| Siglec-8 | aggrecanS8L, DMBT1S8L |  | [31] |
 | [68] | ||
| Siglec-9 | glycophorinS9L, hyaluronic acid, MUC5BS9L, MUC1S9L, MUC16S9L |  | [22] |
| Siglec-11 | polysialic acid (NCAM) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez-Gil, A.; Schnaar, R.L. Siglec Ligands. Cells 2021, 10, 1260. https://doi.org/10.3390/cells10051260
Gonzalez-Gil A, Schnaar RL. Siglec Ligands. Cells. 2021; 10(5):1260. https://doi.org/10.3390/cells10051260
Chicago/Turabian StyleGonzalez-Gil, Anabel, and Ronald L. Schnaar. 2021. "Siglec Ligands" Cells 10, no. 5: 1260. https://doi.org/10.3390/cells10051260
APA StyleGonzalez-Gil, A., & Schnaar, R. L. (2021). Siglec Ligands. Cells, 10(5), 1260. https://doi.org/10.3390/cells10051260