
Long-Term Suppression of c-Jun and nNOS Preserves Ultrastructural Features of Lower Motor Neurons and Forelimb Function after Brachial Plexus Roots Avulsion

 ,
,  and
and 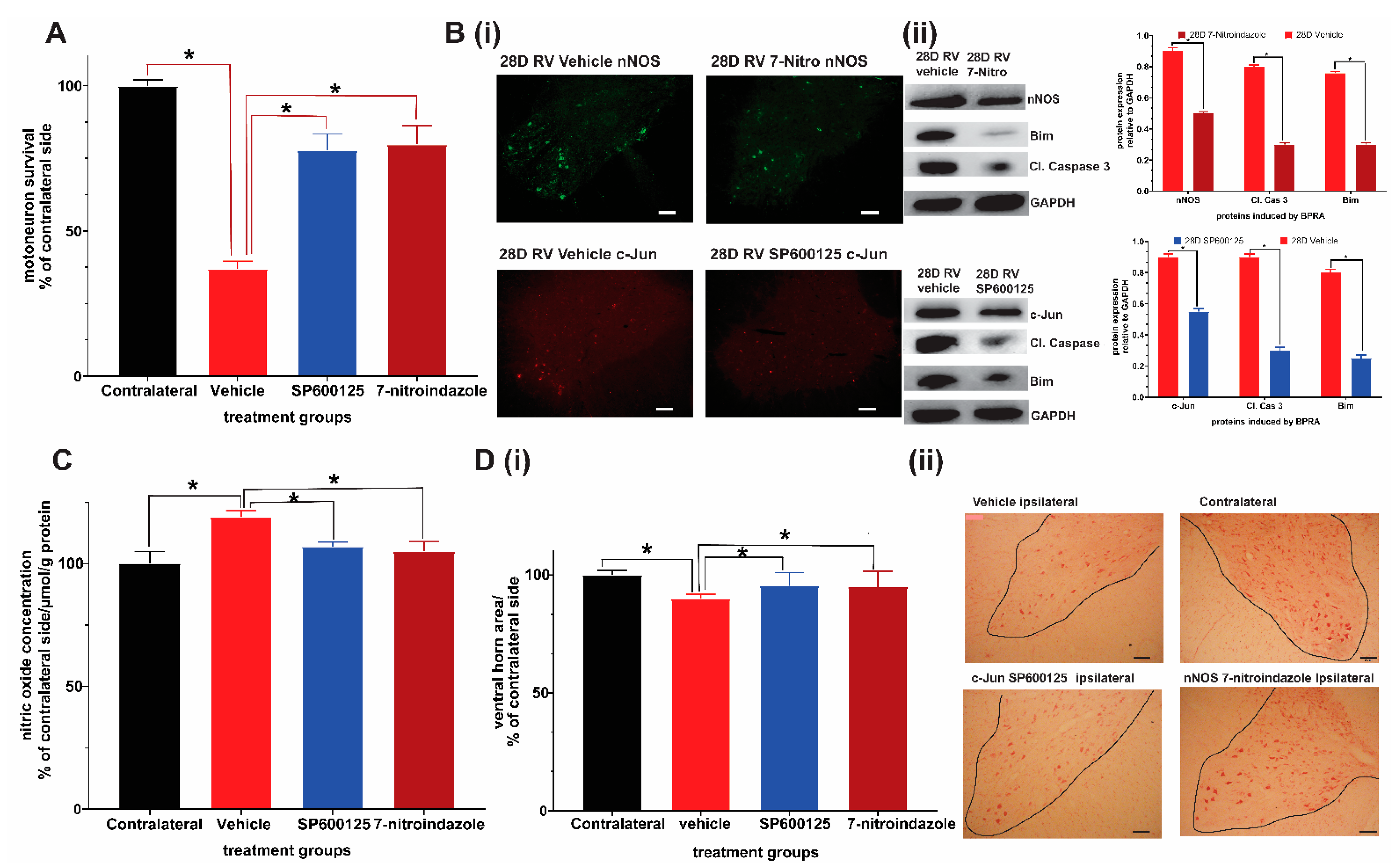{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Ethics Statement
Brachial Plexus Avulsion
2.2. Experimental Grouping and Treatments
2.3. Electron Microscopy
2.4. Analysis of the Ultrathin Sections
2.5. Morphometrical and Statistical Analysis
2.6. Light Microscopy: Motor Neuron Survival
2.7. Immunofluorescence
2.8. Western Blotting
2.9. Oxidative Stress: Nitric Oxide Production Assay
2.10. Functional Evaluations
2.10.1. Terzis Grooming Test
2.10.2. Grid Walk Test
2.11. Statistical Analysis
3. Results
3.1. Chronic Pharmacological Inhibition of c-Jun and nNOS Is Neuroprotective, Relieves Oxidative/Nitrosative Stress, and Promotes Functional Recovery
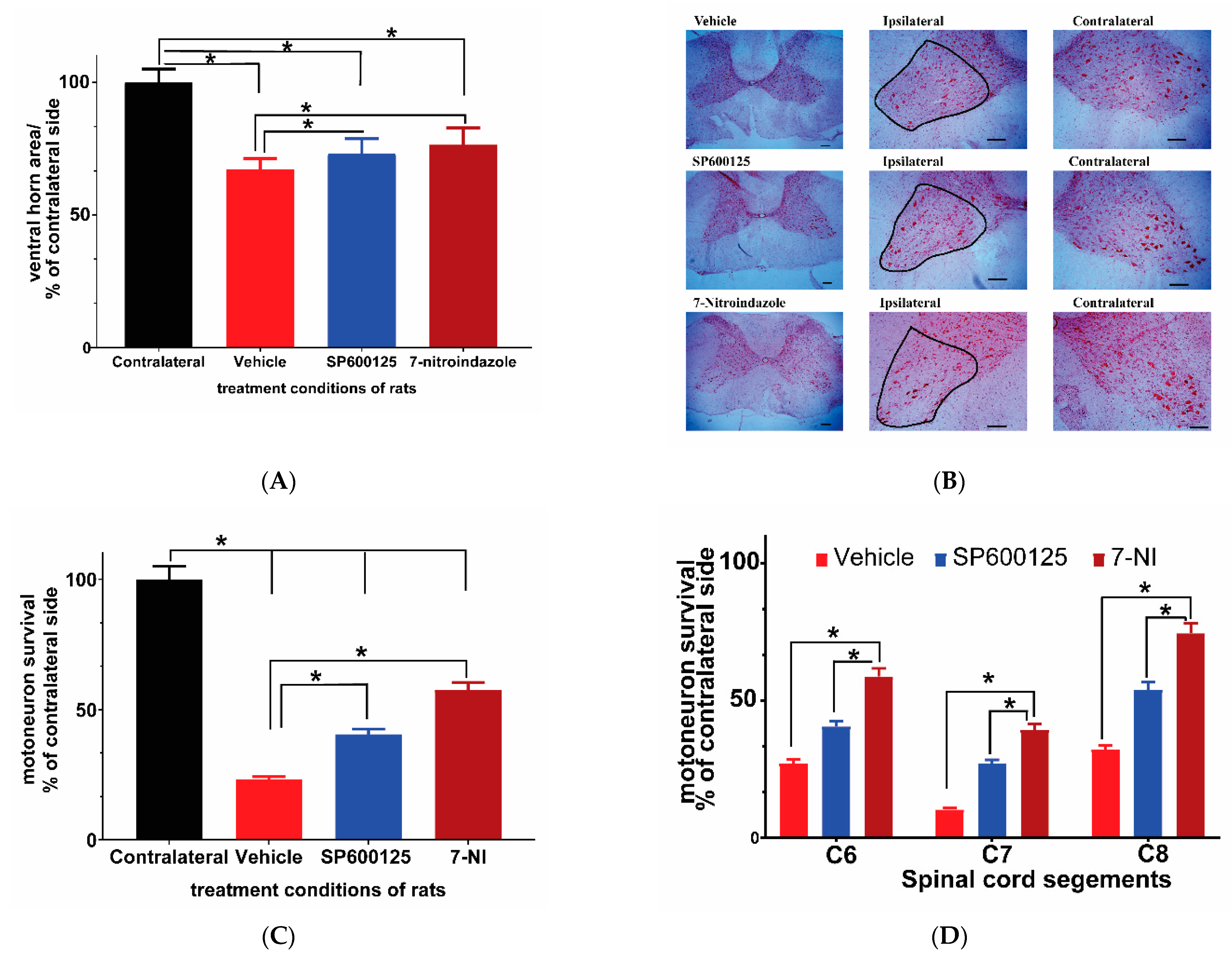
3.2. Avulsion Causes Long-Term Ventral Horn Atrophy and Is Partially Rescued by Neuroprotective Treatments
3.3. Avulsion Causes Motor Neuron Loss at 6 Months Post-Injury and Was Attenuated by Neuroprotective Treatments
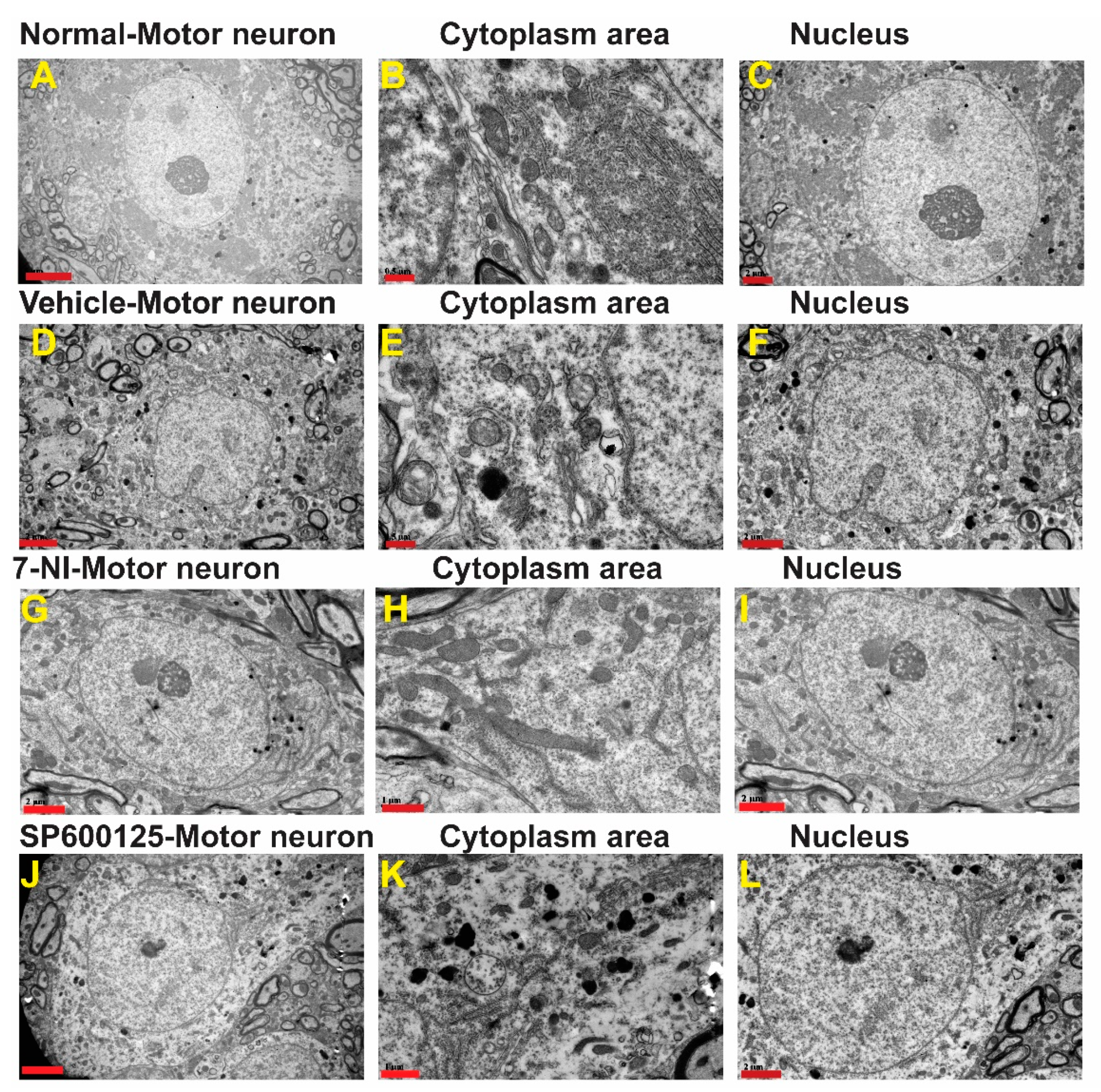
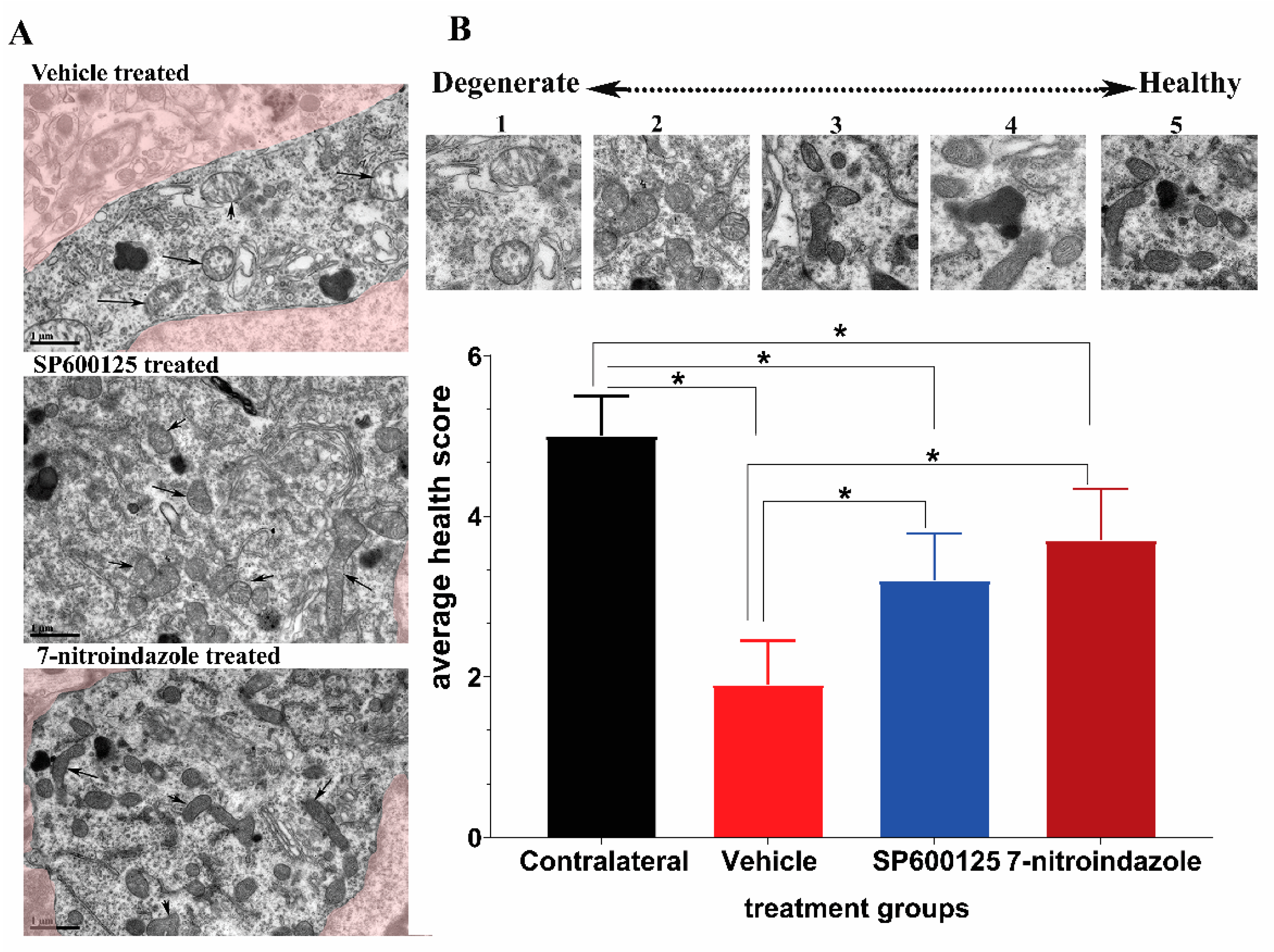
3.4. Avulsion Causes Longstanding Severe Ultrastructural Changes in Motor Neurons
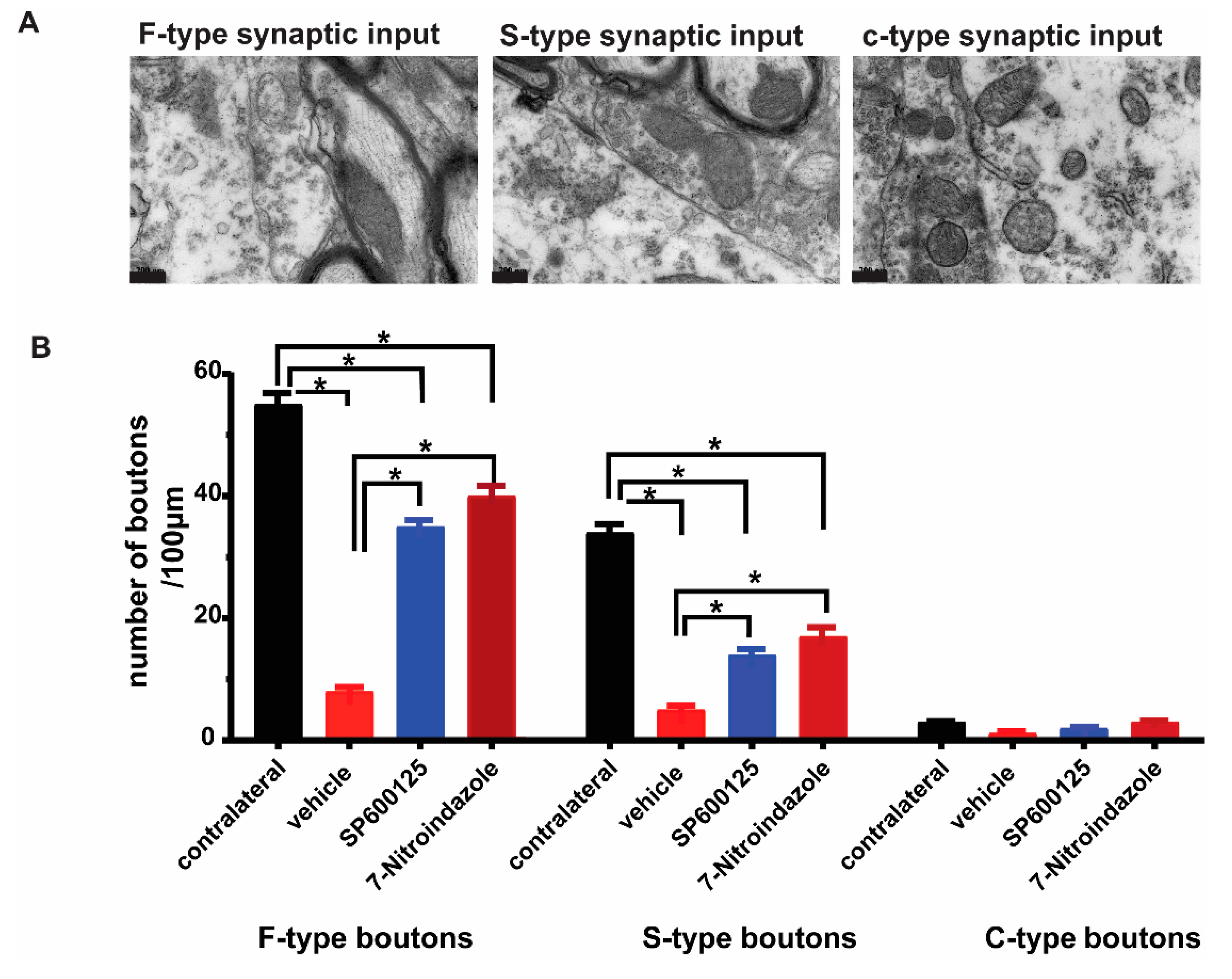
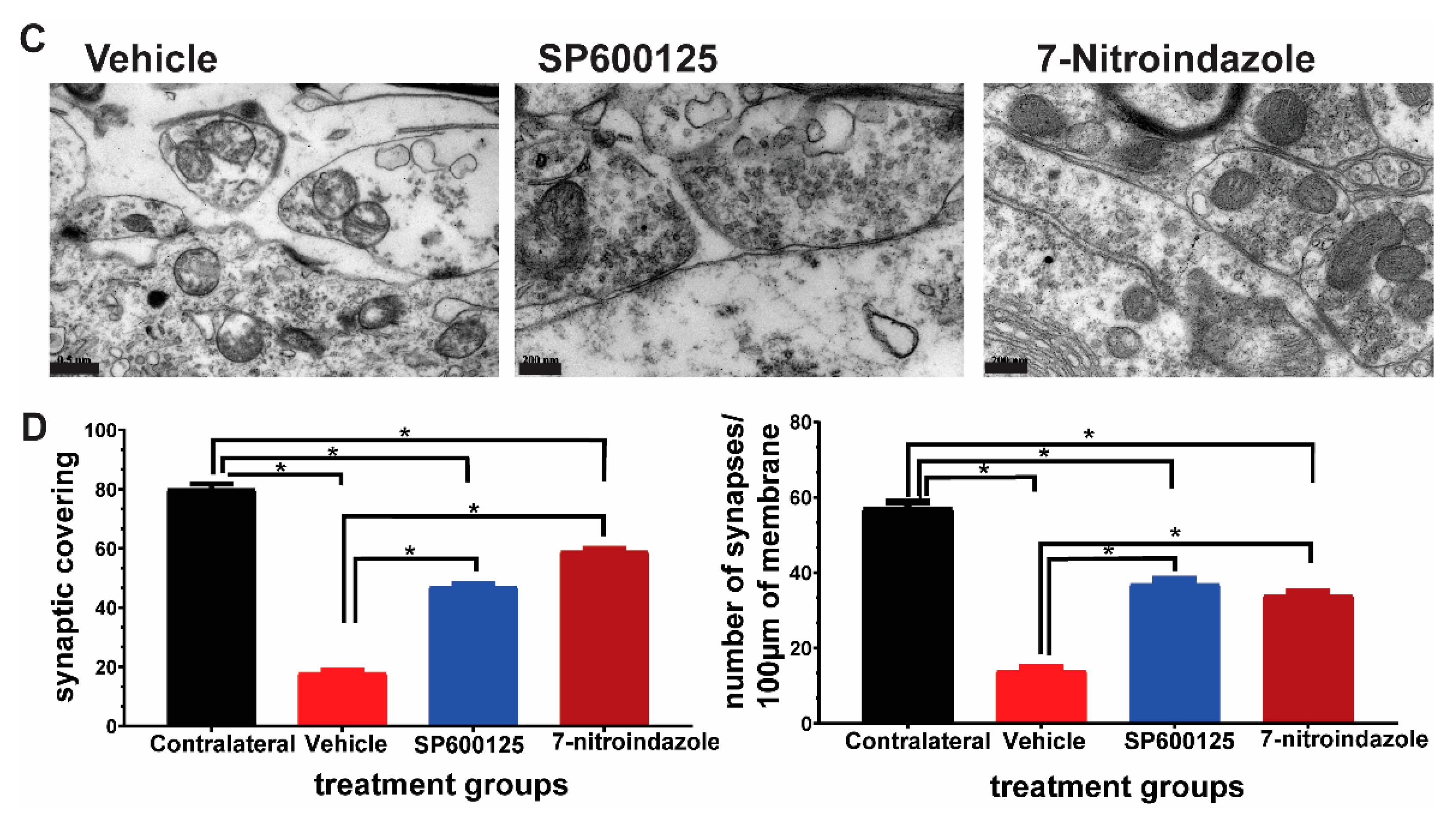
3.5. Downregulation of nNOS by 7-Nitroindazole Preserved Synaptic Inputs on Alpha Motor Neurons
3.6. Ultrastructural Observations of Axons in the Vicinity of Motor Neurons
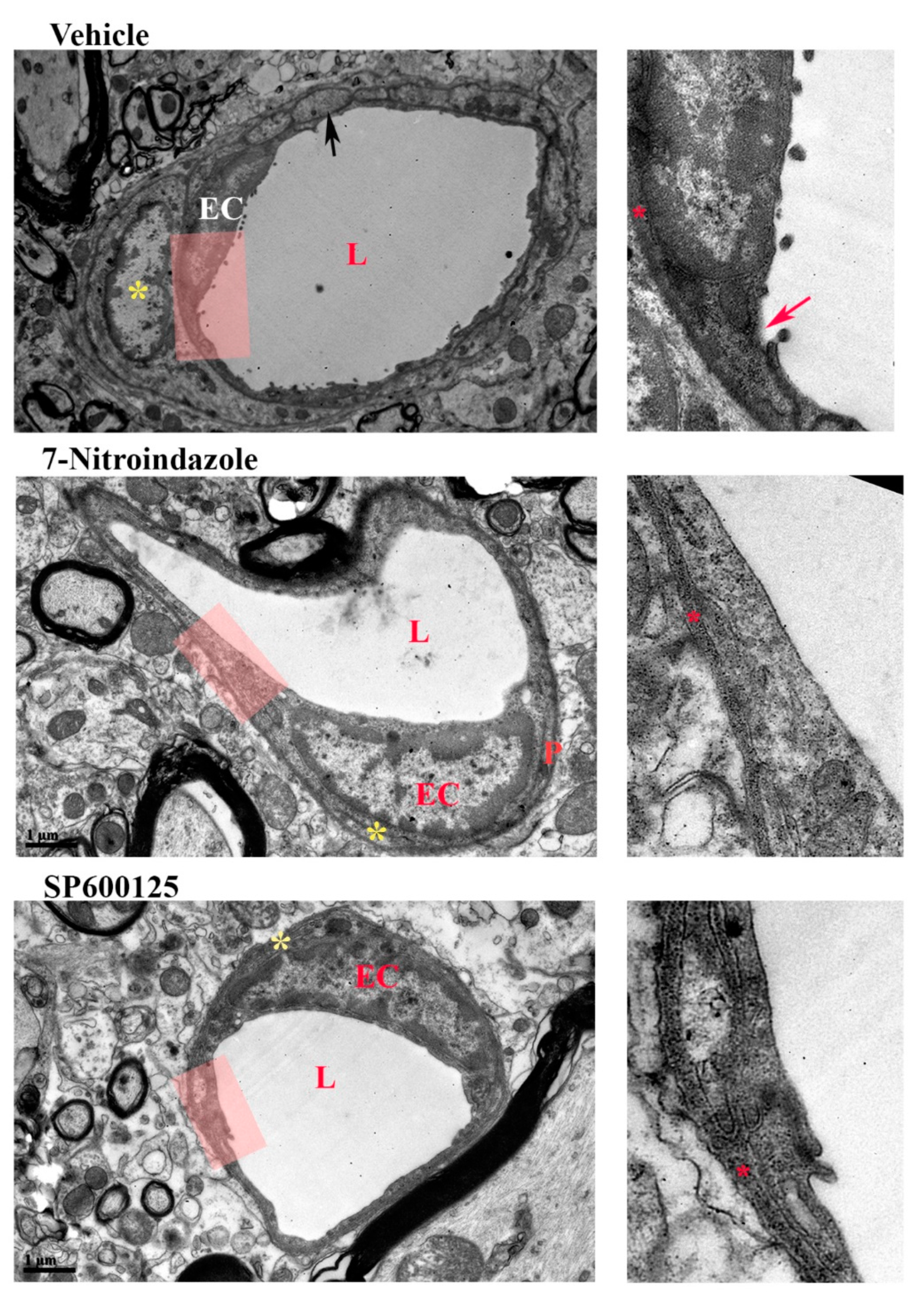
3.7. Neurovascular Unit Preserved by Neuroprotective Treatment
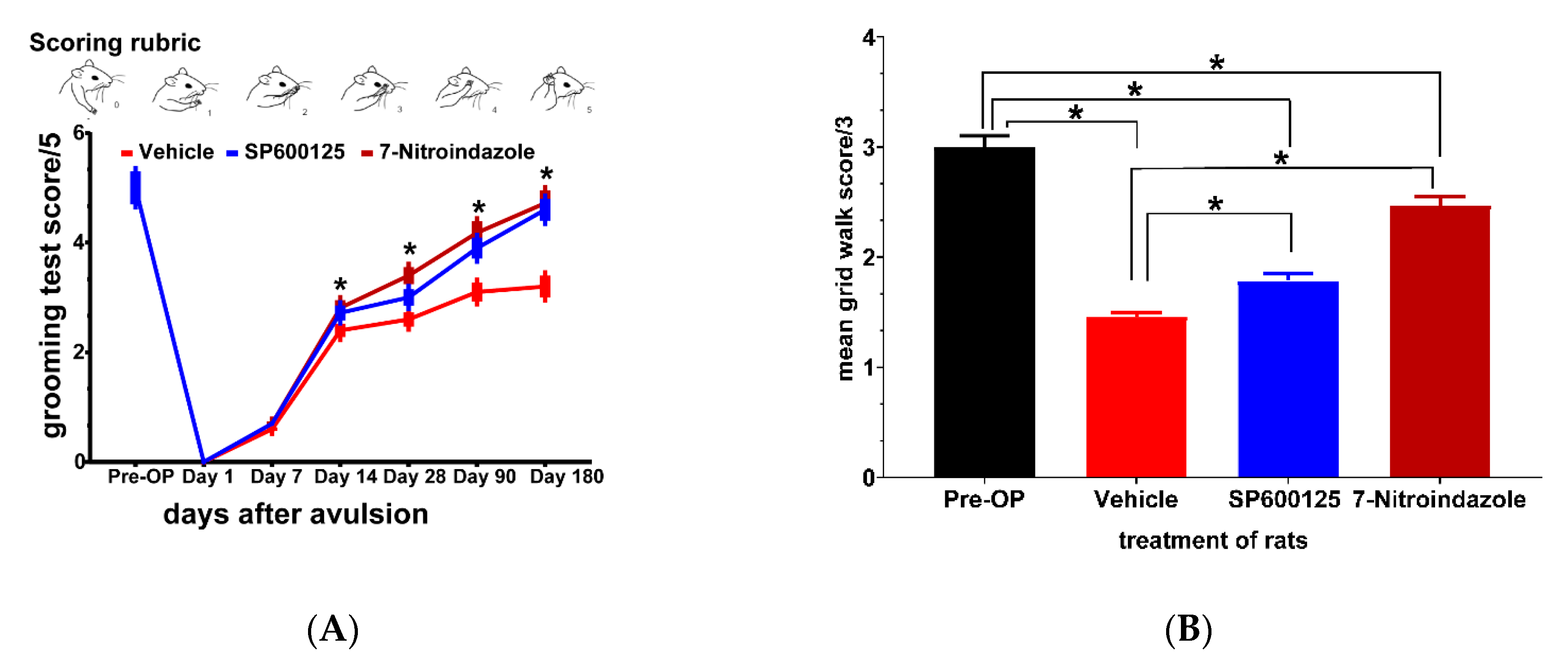
3.8. Functional Recovery at Six Months
3.8.1. Terzis Grooming Test
3.8.2. Grid Walk Test
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bertelli, J.A.; Ghizoni, M.F. Pain after Avulsion Injuries and Complete Palsy of the Brachial Plexus: The Possible Role of Nonavulsed Roots in Pain Generation. Neurosurgery 2008, 62, 1104–1113, discussion 1113–1114. [Google Scholar] [CrossRef]
- Scorisa, J.M.; Zanon, R.G.; Freria, C.M.; de Oliveira, A.L.R. Glatiramer Acetate Positively Influences Spinal Motoneuron Survival and Synaptic Plasticity after Ventral Root Avulsion. Neurosci. Lett. 2009, 451, 34–39. [Google Scholar] [CrossRef]
- Ali, Z.S.; Johnson, V.E.; Stewart, W.; Zager, E.L.; Xiao, R.; Heuer, G.G.; Weber, M.T.; Mallela, A.N.; Smith, D.H. Neuropathological Characteristics of Brachial Plexus Avulsion Injury With and Without Concomitant Spinal Cord Injury. J. Neuropathol. Exp. Neurol. 2016, 75, 69–85. [Google Scholar] [CrossRef] [Green Version]
- Bergerot, A. Co-Treatment with Riluzole and GDNF Is Necessary for Functional Recovery after Ventral Root Avulsion Injury. Exp. Neurol. 2004, 187, 359–366. [Google Scholar] [CrossRef]
- Wiberg, R.; Kingham, P.J.; Novikova, L.N. A Morphological and Molecular Characterization of the Spinal Cord after Ventral Root Avulsion or Distal Peripheral Nerve Axotomy Injuries in Adult Rats. J. Neurotrauma 2017, 34, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-H.; Han, S.; Xie, Y.-Y.; Wang, L.-L.; Yao, Z.-B. Differences in C-Jun and nNOS Expression Levels in Motoneurons Following Different Kinds of Axonal Injury in Adult Rats. Brain Cell Biol. 2008, 36, 213–227. [Google Scholar] [CrossRef]
- Eggers, R.; Tannemaat, M.R.; Ehlert, E.M.; Verhaagen, J. A Spatio-Temporal Analysis of Motoneuron Survival, Axonal Regeneration and Neurotrophic Factor Expression after Lumbar Ventral Root Avulsion and Implantation. Exp. Neurol. 2010, 223, 207–220. [Google Scholar] [CrossRef]
- Holtzer, C.A.J.; Feirabend, H.K.P.; Marani, E.; Thomeer, R.T.W.M. Ultrastructural And Quantitative Motoneuronal Changes After Ventral Root Avulsion Favor Early Surgical Repair. Arch. Physiol. Biochem. 2000, 108, 293–309. [Google Scholar] [CrossRef]
- Martin, E.; Senders, J.T.; DiRisio, A.C.; Smith, T.R.; Broekman, M.L.D. Timing of Surgery in Traumatic Brachial Plexus Injury: A Systematic Review. J. Neurosurg. 2019, 130, 1333–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexeeva, N.; Sames, C.; Jacobs, P.L.; Hobday, L.; DiStasio, M.M.; Mitchell, S.A.; Calancie, B. Comparison of Training Methods to Improve Walking in Persons with Chronic Spinal Cord Injury: A Randomized Clinical Trial. J. Spinal Cord Med. 2011, 34, 362–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Azevedo Filho, F.A.S.; Abdouni, Y.A.; Ogawa, G.; de Sá, C.K.C.; da Costa, A.C.; de Moraes Barros Fucs, P.M. Functional Outcome of Oberlin Procedure. Acta Ortopédica Bras. 2019, 27, 294–297. [Google Scholar] [CrossRef] [Green Version]
- Carlstedt, T.; Anand, P.; Hallin, R.; Vigay Misra, P.; Norén, G.; Seferlis, T. Spinal Nerve Root Repair and Reimplantation of Avulsed Ventral Roots into the Spinal Cord after Brachial Plexus Injury. J. Neurosurg. Spine 2000, 93, 237–247. [Google Scholar] [CrossRef]
- Field-Fote, E.C.; Roach, K.E. Influence of a Locomotor Training Approach on Walking Speed and Distance in People With Chronic Spinal Cord Injury: A Randomized Clinical Trial. Phys. Ther. 2011, 91, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Ko, C.-C.; Tu, T.-H.; Wu, J.-C.; Huang, W.-C.; Tsai, Y.-A.; Huang, S.-F.; Huang, H.-C.; Cheng, H. Functional Improvement in Chronic Human Spinal Cord Injury: {Four} Years after Acidic Fibroblast Growth Factor. Sci. Rep. 2018, 8, 12691. [Google Scholar] [CrossRef] [Green Version]
- Sedain, G.; Sharma, M.S.; Sharma, B.S.; Mahapatra, A.K. Outcome After Delayed Oberlin Transfer in Brachial Plexus Injury. Neurosurgery 2011, 69, 822–828. [Google Scholar] [CrossRef]
- Griffin, J.M.; Bradke, F. Therapeutic Repair for Spinal Cord Injury: Combinatory Approaches to Address a Multifaceted Problem. EMBO Mol. Med. 2020, 12. [Google Scholar] [CrossRef]
- Kachramanoglou, C.; Carlstedt, T.; Koltzenburg, M.; Choi, D. Long-Term Outcome of Brachial Plexus Reimplantation After Complete Brachial Plexus Avulsion Injury. World Neurosurg. 2017, 103, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Bossy-Wetzel, E. Induction of Apoptosis by the Transcription Factor c-Jun. EMBO J. 1997, 16, 1695–1709. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Cao, R.-J.; Zhu, X.-J.; Liu, X.-Y.; Li, L.-Y.; Cui, S.-S. Erythropoietin Attenuates the Apoptosis of Adult Neurons After Brachial Plexus Root Avulsion by Downregulating JNK Phosphorylation and c-Jun Expression and Inhibiting c-PARP Cleavage. J. Mol. Neurosci. 2015, 56, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.; Eilers, A.; Lallemand, D.; Kyriakis, J.; Rubin, L.L.; Ham, J. Phosphorylation of C-Jun Is Necessary for Apoptosis Induced by Survival Signal Withdrawal in Cerebellar Granule Neurons. J. Neurosci. 1998, 18, 751–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.-L.; Zhao, X.-C.; Yan, L.-F.; Wang, Y.-Q.; Cheng, X.; Fu, R.; Zhou, L.-H. C-Jun Phosphorylation Contributes to down Regulation of Neuronal Nitric Oxide Synthase Protein and Motoneurons Death in Injured Spinal Cords Following Root-Avulsion of the Brachial Plexus. Neuroscience 2011, 189, 397–407. [Google Scholar] [CrossRef]
- Kim, B.-J.; Silverman, S.M.; Liu, Y.; Wordinger, R.J.; Pang, I.-H.; Clark, A.F. In Vitro and in Vivo Neuroprotective Effects of CJun N-Terminal Kinase Inhibitors on Retinal Ganglion Cells. Mol. Neurodegener. 2016, 11, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, P.; Cinelli, M.A.; Kang, S.; Silverman, R.B. Development of Nitric Oxide Synthase Inhibitors for Neurodegeneration and Neuropathic Pain. Chem. Soc. Rev. 2014, 43, 6814–6838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Iwasaki, Y.; Kinoshita, M. Neuronal Nitric Oxide Synthase Inhibitor, 7-Nitroindazole, Delays Motor Dysfunction and Spinal Motoneuron Degeneration in the Wobbler Mouse. J. Neurol. Sci. 1998, 160, 9–15. [Google Scholar] [CrossRef]
- Sim, S.K.; Tan, Y.C.; Tee, J.H.; Yusoff, A.A.; Abdullah, J.M. Paclitaxel Inhibits Expression of Neuronal Nitric Oxide Synthase and Prevents Mitochondrial Dysfunction in Spinal Ventral Horn in Rats after C7 Spinal Root Avulsion. Turk. Neurosurg. 2015. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Li, L.; Yick, L.-W.; Chai, H.; Xie, Y.; Yang, Y.; Prevette, D.M.; Oppenheim, R.W. GDNF and BDNF Alter the Expression of Neuronal NOS, c-Jun, and P75 and Prevent Motoneuron Death Following Spinal Root Avulsion in Adult Rats. J. Neurotrauma 2003, 20, 603–612. [Google Scholar] [CrossRef]
- Wu, W.; Li, L. Inhibition of Nitric Oxide Synthase Reduces Motoneuron Death Due to Spinal Root Avulsion. Neurosci. Lett. 1993, 153, 121–124. [Google Scholar] [CrossRef]
- Barbizan, R.; Oliveira, A.L. Impact of Acute Inflammation on Spinal Motoneuron Synaptic Plasticity Following Ventral Root Avulsion. J. Neuroinflamm. 2010, 7, 29. [Google Scholar] [CrossRef] [Green Version]
- Conradi, S. Ultrastructure and Distribution of Neuronal and Glial Elements on the Motoneuron Surface in the Lumbosacral Spinal Cord of the Adult Cat. Acta Physiol. Scand Suppl. 1969, 332, 5–48. [Google Scholar] [PubMed]
- Coughlin, L.; Morrison, R.S.; Horner, P.J.; Inman, D.M. Mitochondrial Morphology Differences and Mitophagy Deficit in Murine Glaucomatous Optic Nerve. Invest. Ophthalmol. Vis. Sci. 2015, 56, 1437–1446. [Google Scholar] [CrossRef] [Green Version]
- Berling, K.; Mannström, P.; Ulfendahl, M.; Danckwardt Lillieström, N.; von Unge, M. The Chorda Tympani Degenerates during Chronic Otitis Media: An Electron Microscopy Study. Acta Otolaryngol. (Stockh.) 2015, 135, 542–548. [Google Scholar] [CrossRef]
- Yu, G.; Zilundu, P.L.M.; Liu, L.; Zhong, K.; Tang, Y.; Ling, Z.; Zhou, L.-H. ERRγ Is Downregulated in Injured Motor Neuron Subpopulations Following Brachial Plexus Root Avulsion. Exp. Ther. Med. 2020, 19, 205–213. [Google Scholar] [CrossRef] [Green Version]
- Ruven, C.; Badea, S.-R.; Wong, W.-M.; Wu, W. Combination Treatment With Exogenous GDNF and Fetal Spinal Cord Cells Results in Better Motoneuron Survival and Functional Recovery After Avulsion Injury With Delayed Root Reimplantation. J. Neuropathol. Exp. Neurol. 2018, 77, 325–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelli, J.A.; Mira, J.-C. Behavioral Evaluating Methods in the Objective Clinical Assessment of Motor Function after Experimental Brachial Plexus Reconstruction in the Rat. J. Neurosci. Methods 1993, 46, 203–208. [Google Scholar] [CrossRef]
- Korak, K.J.; Tam, S.L.; Gordon, T.; Frey, M.; Aszmann, O.C. Changes in Spinal Cord Architecture after Brachial Plexus Injury in the Newborn. Brain 2004, 127, 1488–1495. [Google Scholar] [CrossRef] [Green Version]
- Metz, G.A.S.; Merkler, D.; Dietz, V.; Schwab, M.E.; Fouad, K. Efficient Testing of Motor Function in Spinal Cord Injured Rats. Brain Res. 2000, 883, 165–177. [Google Scholar] [CrossRef]
- Pintér, S.; Gloviczki, B.; Szabó, A.; Márton, G.; Nógrádi, A. Increased Survival and Reinnervation of Cervical Motoneurons by Riluzole after Avulsion of the C7 Ventral Root. J. Neurotrauma 2010, 27, 2273–2282. [Google Scholar] [CrossRef] [Green Version]
- Drozdz, M.M.; Jiang, H.; Pytowski, L.; Grovenor, C.; Vaux, D.J. Formation of a Nucleoplasmic Reticulum Requires de Novo Assembly of Nascent Phospholipids and Shows Preferential Incorporation of Nascent Lamins. Sci. Rep. 2017, 7, 7454. [Google Scholar] [CrossRef] [Green Version]
- Malhas, A.; Goulbourne, C.; Vaux, D.J. The Nucleoplasmic Reticulum: Form and Function. Trends Cell Biol. 2011, 21, 362–373. [Google Scholar] [CrossRef]
- Lindenboim, L.; Zohar, H.; Worman, H.J.; Stein, R. The Nuclear Envelope: Target and Mediator of the Apoptotic Process. Cell Death Discov. 2020, 6, 29. [Google Scholar] [CrossRef]
- De freria, C.M.; Barbizan, R.; De Oliveira, A.L.R. Granulocyte Colony Stimulating Factor Neuroprotective Effects on Spinal Motoneurons after Ventral Root Avulsion. Synapse 2012, 66, 128–141. [Google Scholar] [CrossRef]
- Barbizan, R.; Castro, M.V.; Ferreira, R.S.; Barraviera, B.; Oliveira, A.L.R. Long-Term Spinal Ventral Root Reimplantation, but Not Bone Marrow Mononuclear Cell Treatment, Positively Influences Ultrastructural Synapse Recovery and Motor Axonal Regrowth. Int. J. Mol. Sci. 2014, 15, 19535–19551. [Google Scholar] [CrossRef]
- Ohlsson, M.; Nieto, J.H.; Christe, K.L.; Havton, L.A. Long-Term Effects of a Lumbosacral Ventral Root Avulsion Injury on Axotomized Motor Neurons and Avulsed Ventral Roots in a Non-Human Primate Model of Cauda Equina Injury. Neuroscience 2013, 250, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Koliatsos, V.E.; Price, W.L.; Pardo, C.A.; Price, D.L. Ventral Root Avulsion: An Experimental Model of Death of Adult Motor Neurons. J. Comp. Neurol. 1994, 342, 35–44. [Google Scholar] [CrossRef]
- Noguchi, T.; Ohta, S.; Kakinoki, R.; Kaizawa, Y.; Matsuda, S. A New Cervical Nerve Root Avulsion Model Using a Posterior Extra-Vertebral Approach in Rats. J. Brachial Plex. Peripher. Nerve Inj. 2014, 08, e19–e27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbizan, R.; Castro, M.V.; Rodrigues, A.C.; Barraviera, B.; Ferreira, R.S.; Oliveira, A.L.R. Motor Recovery and Synaptic Preservation after Ventral Root Avulsion and Repair with a Fibrin Sealant Derived from Snake Venom. PLoS ONE 2013, 8, e63260. [Google Scholar] [CrossRef] [Green Version]
- Fox, I.K.; Brenner, M.J.; Johnson, P.J.; Hunter, D.A.; Mackinnon, S.E. Axonal Regeneration and Motor Neuron Survival after Microsurgical Nerve Reconstruction: Nerve Regeneration Over Time. Microsurgery 2012, 32, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.X. Functional Reinnervation of the Rat Lower Urinary Tract after Cauda Equina Injury and Repair. J. Neurosci. 2006, 26, 8672–8679. [Google Scholar] [CrossRef] [Green Version]
- Hoang, T.X.; Nieto, J.H.; Tillakaratne, N.J.K.; Havton, L.A. Autonomic and Motor Neuron Death Is Progressive and Parallel in a Lumbosacral Ventral Root Avulsion Model of Cauda Equina Injury. J. Comp. Neurol. 2003, 467, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.-Y.; Chai, H.; Zhang, J.-Y.; Yao, Z.-B.; Zhou, L.-H.; Wong, W.-M.; Bruce, I.C.; Wu, W.-T. Survival, Regeneration and Functional Recovery of Motoneurons after Delayed Reimplantation of Avulsed Spinal Root in Adult Rat. Exp. Neurol. 2005, 192, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Lang, E.M.; Schlegel, N.; Sendtner, M.; Asan, E. Effects of Root Replantation and Neurotrophic Factor Treatment on Long-Term Motoneuron Survival and Axonal Regeneration after C7 Spinal Root Avulsion. Exp. Neurol. 2005, 194, 341–354. [Google Scholar] [CrossRef]
- Lindå, H.; Shupliakov, O.; Ornung, G.; Ottersen, O.P.; Storm-Mathisen, J.; Risling, M.; Cullheim, S. Ultrastructural Evidence for a Preferential Elimination of Glutamate-Immunoreactive Synaptic Terminals from Spinal Motoneurons after Intramedullary Axotomy. J. Comp. Neurol. 2000, 425, 10–23. [Google Scholar] [CrossRef]
- Carlstedt, T. Nerve Root Replantation. Neurosurg. Clin. N. Am. 2009, 20, 39–50. [Google Scholar] [CrossRef]
- Eggers, R.; Tannemaat, M.R.; De Winter, F.; Malessy, M.J.A.; Verhaagen, J. Clinical and Neurobiological Advances in Promoting Regeneration of the Ventral Root Avulsion Lesion. Eur. J. Neurosci. 2016, 43, 318–335. [Google Scholar] [CrossRef]
- Vanaclocha-Vanaclocha, V.; Saiz-Sapena, N.; María Ortiz-Criado, J.; Vanaclocha, L. Nerve Root Reimplantation in Brachial Plexus Injuries. In Treatment of Brachial Plexus Injuries; Vanaclocha, V., Sáiz-Sapena, N., Eds.; IntechOpen: London, UK, 2019; ISBN 978-1-83880-403-9. [Google Scholar]
- Ibrahim, A.G.; Kirkwood, P.A.; Raisman, G.; Li, Y. Restoration of Hand Function in a Rat Model of Repair of Brachial Plexus Injury. Brain 2009, 132, 1268–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jivan, S.; Novikova, L.N.; Wiberg, M.; Novikov, L.N. The Effects of Delayed Nerve Repair on Neuronal Survival and Axonal Regeneration after Seventh Cervical Spinal Nerve Axotomy in Adult Rats. Exp. Brain Res. 2006, 170, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Chan, T.-K.; Ruven, C. Spinal Root Avulsion: An Excellent Model for Studying Motoneuron Degeneration and Regeneration after Severe Axonal Injury. Neural Regen. Res. 2014, 9, 117. [Google Scholar] [CrossRef]
- Huber, E.; David, G.; Thompson, A.J.; Weiskopf, N.; Mohammadi, S.; Freund, P. Dorsal and Ventral Horn Atrophy Is Associated with Clinical Outcome after Spinal Cord Injury. Neurology 2018, 90, e1510–e1522. [Google Scholar] [CrossRef] [Green Version]
- Konishi, H.; Okamoto, T.; Hara, Y.; Komine, O.; Tamada, H.; Maeda, M.; Osako, F.; Kobayashi, M.; Nishiyama, A.; Kataoka, Y.; et al. Astrocytic Phagocytosis Is a Compensatory Mechanism for Microglial Dysfunction. EMBO J. 2020, 39. [Google Scholar] [CrossRef]
- Lööv, C.; Hillered, L.; Ebendal, T.; Erlandsson, A. Engulfing Astrocytes Protect Neurons from Contact-Induced Apoptosis Following Injury. PLoS ONE 2012, 7, e33090. [Google Scholar] [CrossRef] [Green Version]
- Trias, E.; Barbeito, L.; Yamanaka, K. Phenotypic Heterogeneity of Astrocytes in Motor Neuron Disease. Clin. Exp. Neuroimmunol. 2018, 9, 225–234. [Google Scholar] [CrossRef]
- Alvarez, F.J.; Rotterman, T.M.; Akhter, E.T.; Lane, A.R.; English, A.W.; Cope, T.C. Synaptic Plasticity on Motoneurons After Axotomy: A Necessary Change in Paradigm. Front. Mol. Neurosci. 2020, 13, 68. [Google Scholar] [CrossRef]
- Aldskogius, H.; Liu, L.; Svensson, M. Glial Responses to Synaptic Damage and Plasticity. J Neurosci. Res 1999, 58, 33–41. [Google Scholar] [CrossRef]
- Cheng, X.; Liu, S.; Wang, Y.-Q.; Li, Y.-Q.; Fu, R.; Tang, Y.; Zheng, W.-H.; Zhou, L.-H. Suppression of C-Jun Influences NNOS Expression in Differentiated PC12 Cells. Mol. Med. Rep. 2012, 6, 750–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Luo, H.; Hou, Z.; Huang, Y.; Sun, J.; Zhou, L. Neuronal Nitric Oxide Synthase, as a Downstream Signaling Molecule of C-jun, Regulates the Survival of Differentiated PC12 Cells. Mol. Med. Rep. 2014, 10, 1881–1886. [Google Scholar] [CrossRef]
- Montero, F.; Sunico, C.R.; Liu, B.; Paton, J.F.R.; Kasparov, S.; Moreno-López, B. Transgenic Neuronal Nitric Oxide Synthase Expression Induces Axotomy-like Changes in Adult Motoneurons. J. Physiol. 2010, 588, 3425–3443. [Google Scholar] [CrossRef] [PubMed]
- Sunico, C.R.; Portillo, F.; González-Forero, D.; Moreno-López, B. Nitric-Oxide-Directed Synaptic Remodeling in the Adult Mammal {CNS}. J. Neurosci. 2005, 25, 1448–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunico, C.R.; González-Forero, D.; Domínguez, G.; García-Verdugo, J.M.; Moreno-López, B. Nitric Oxide Induces Pathological Synapse Loss by a Protein Kinase G-, Rho Kinase-Dependent Mechanism Preceded by Myosin Light Chain Phosphorylation. J. Neurosci. 2010, 30, 973–984. [Google Scholar] [CrossRef]
- Moreno-López, B.; Sunico, C.R.; González-Forero, D. NO Orchestrates the Loss of Synaptic Boutons from Adult “Sick” Motoneurons: Modeling a Molecular Mechanism. Mol. Neurobiol. 2011, 43, 41–66. [Google Scholar] [CrossRef]
- Pytowski, L.; Drozdz, M.M.; Jiang, H.; Hernandez, Z.; Kumar, K.; Knott, E.; Vaux, D.J. Vaux Nucleoplasmic Reticulum Formation in Human Endometrial Cells Is Steroid Hormone Responsive and Recruits Nascent Components. Int. J. Mol. Sci. 2019, 20, 5839. [Google Scholar] [CrossRef] [Green Version]
- Raefsky, S.M.; Mattson, M.P. Adaptive Responses of Neuronal Mitochondria to Bioenergetic Challenges: Roles in Neuroplasticity and Disease Resistance. Free Radic. Biol. Med. 2017, 102, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.L.-H.; Chiang, S.; Kalinowski, D.S.; Bae, D.-H.; Sahni, S.; Richardson, D.R. The Role of the Antioxidant Response in Mitochondrial Dysfunction in Degenerative Diseases: Cross-Talk between Antioxidant Defense, Autophagy, and Apoptosis. Oxid. Med. Cell. Longev. 2019, 2019, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Kodavati, M.; Wang, H.; Hegde, M.L. Altered Mitochondrial Dynamics in Motor Neuron Disease: An Emerging Perspective. Cells 2020, 9, 1065. [Google Scholar] [CrossRef]
- Jiang, J.; Duan, Z.; Nie, X.; Xi, H.; Li, A.; Guo, A.; Wu, Q.; Jiang, S.; Zhao, J.; Chen, G. Activation of Neuronal Nitric Oxide Synthase (NNOS) Signaling Pathway in 2,3,7,8-Tetrachlorodibenzo-p-Dioxin (TCDD)-Induced Neurotoxicity. Environ. Toxicol. Pharmacol. 2014, 38, 119–130. [Google Scholar] [CrossRef]
- Prakash Mishra, O.; Zubrow, A.B.; Ashraf, Q.M.; Delivoria-Papadopoulos, M. Effect of Nitric Oxide Synthase Inhibition during Post-Hypoxic Reoxygenation on Bax and Bcl-2 Protein Expression and DNA Fragmentation in Neuronal Nuclei of Newborn Piglets. Brain Res. 2006, 1101, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Natsume, A.; Mata, M.; Wolfe, D.; Oligino, T.; Goss, J.; Huang, S.; Glorioso, J.; Fink, D.J. Bcl-2 and GDNF Delivered by HSV-Mediated Gene Transfer after Spinal Root Avulsion Provide a Synergistic Effect. J. Neurotrauma 2002, 19, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.R.; Voeltz, G.K. The ER in 3D: A Multifunctional Dynamic Membrane Network. Trends Cell Biol. 2011, 21, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Wang, W.; Hwang, J.; Namgung, U.; Min, K.-T. Increased ER-Mitochondria Tethering Promotes Axon Regeneration. Proc. Natl. Acad. Sci. USA 2019, 116, 16074–16079. [Google Scholar] [CrossRef] [Green Version]
- Shibata, Y.; Voeltz, G.K.; Rapoport, T.A. Rough Sheets and Smooth Tubules. Cell 2006, 126, 435–439. [Google Scholar] [CrossRef] [Green Version]
- Dziewulska, D.; Gogol, A.; Gogol, P.; Rafalowska, J. Enlargement of the Nissl Substance as a Manifestation of Early Damage to Spinal Cord Motoneurons in Amyotrophic Lateral Sclerosis. Clin. Neuropathol. 2013, 32, 480–485. [Google Scholar] [CrossRef]
- Terasaki, M. Axonal Endoplasmic Reticulum Is Very Narrow. J. Cell Sci. 2018, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, D.L.; Porter, K.R. The Response of Ventral Horn Neurons to Axonal Transection. J. Cell Biol. 1972, 53, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Fricker, M.; Tolkovsky, A.M.; Borutaite, V.; Coleman, M.; Brown, G.C. Neuronal Cell Death. Physiol. Rev. 2018, 98, 813–880. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zilundu, P.L.M.; Xu, X.; Liaquat, Z.; Wang, Y.; Zhong, K.; Fu, R.; Zhou, L. Long-Term Suppression of c-Jun and nNOS Preserves Ultrastructural Features of Lower Motor Neurons and Forelimb Function after Brachial Plexus Roots Avulsion. Cells 2021, 10, 1614. https://doi.org/10.3390/cells10071614
Zilundu PLM, Xu X, Liaquat Z, Wang Y, Zhong K, Fu R, Zhou L. Long-Term Suppression of c-Jun and nNOS Preserves Ultrastructural Features of Lower Motor Neurons and Forelimb Function after Brachial Plexus Roots Avulsion. Cells. 2021; 10(7):1614. https://doi.org/10.3390/cells10071614
Chicago/Turabian StyleZilundu, Prince Last Mudenda, Xiaoying Xu, Zaara Liaquat, Yaqiong Wang, Ke Zhong, Rao Fu, and Lihua Zhou. 2021. "Long-Term Suppression of c-Jun and nNOS Preserves Ultrastructural Features of Lower Motor Neurons and Forelimb Function after Brachial Plexus Roots Avulsion" Cells 10, no. 7: 1614. https://doi.org/10.3390/cells10071614
APA StyleZilundu, P. L. M., Xu, X., Liaquat, Z., Wang, Y., Zhong, K., Fu, R., & Zhou, L. (2021). Long-Term Suppression of c-Jun and nNOS Preserves Ultrastructural Features of Lower Motor Neurons and Forelimb Function after Brachial Plexus Roots Avulsion. Cells, 10(7), 1614. https://doi.org/10.3390/cells10071614