
Application of Stem Cell Therapy for Infertility



Abstract
:
1. Introduction
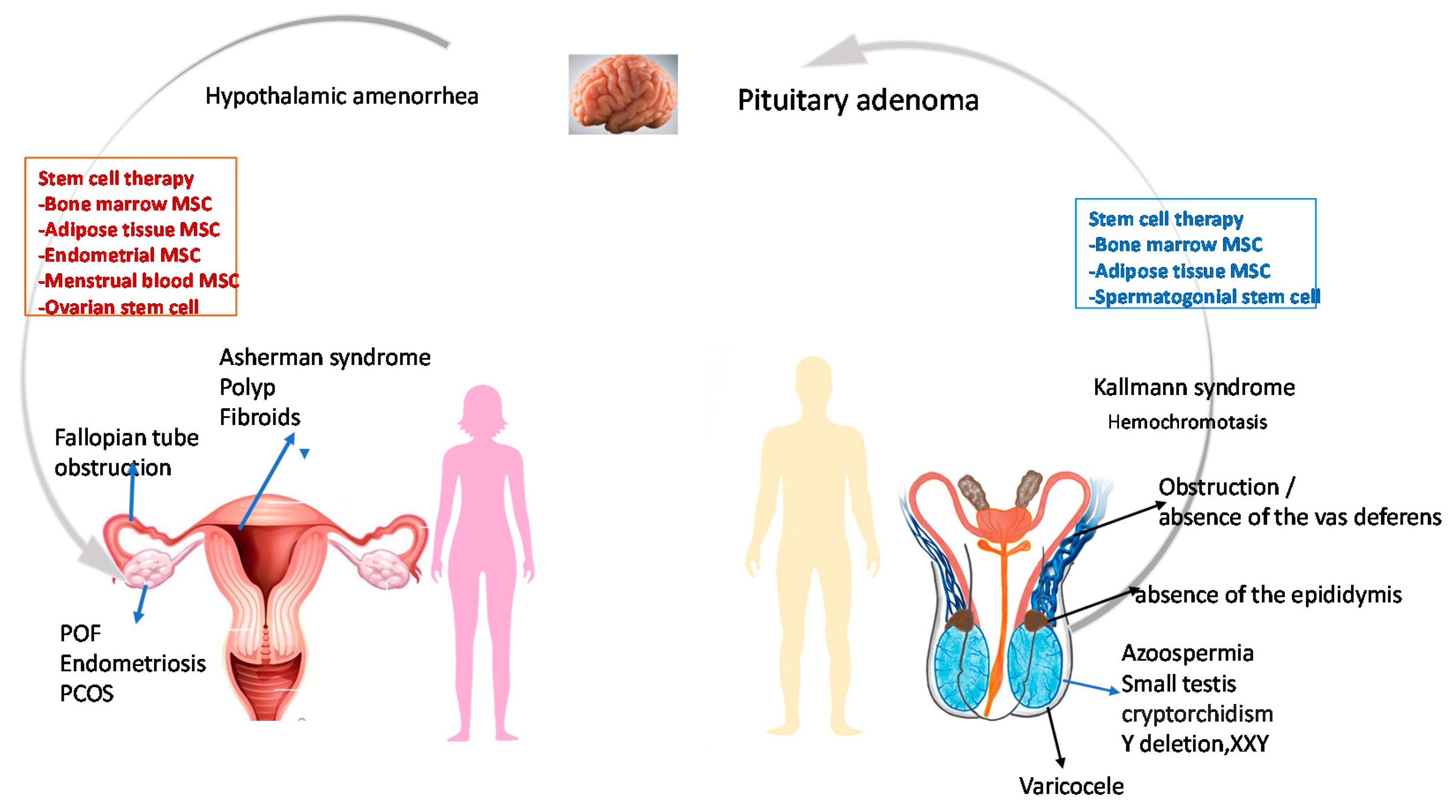
2. Infertility Risk Factors
3. Conventional Treatment
4. Assisted Reproduction Technology (ART)
4.1. Ovarian Stimulation
4.2. Oocyte Retrieval
4.3. In Vitro Fertilization and Embryo Culture
4.4. Embryo Transfer
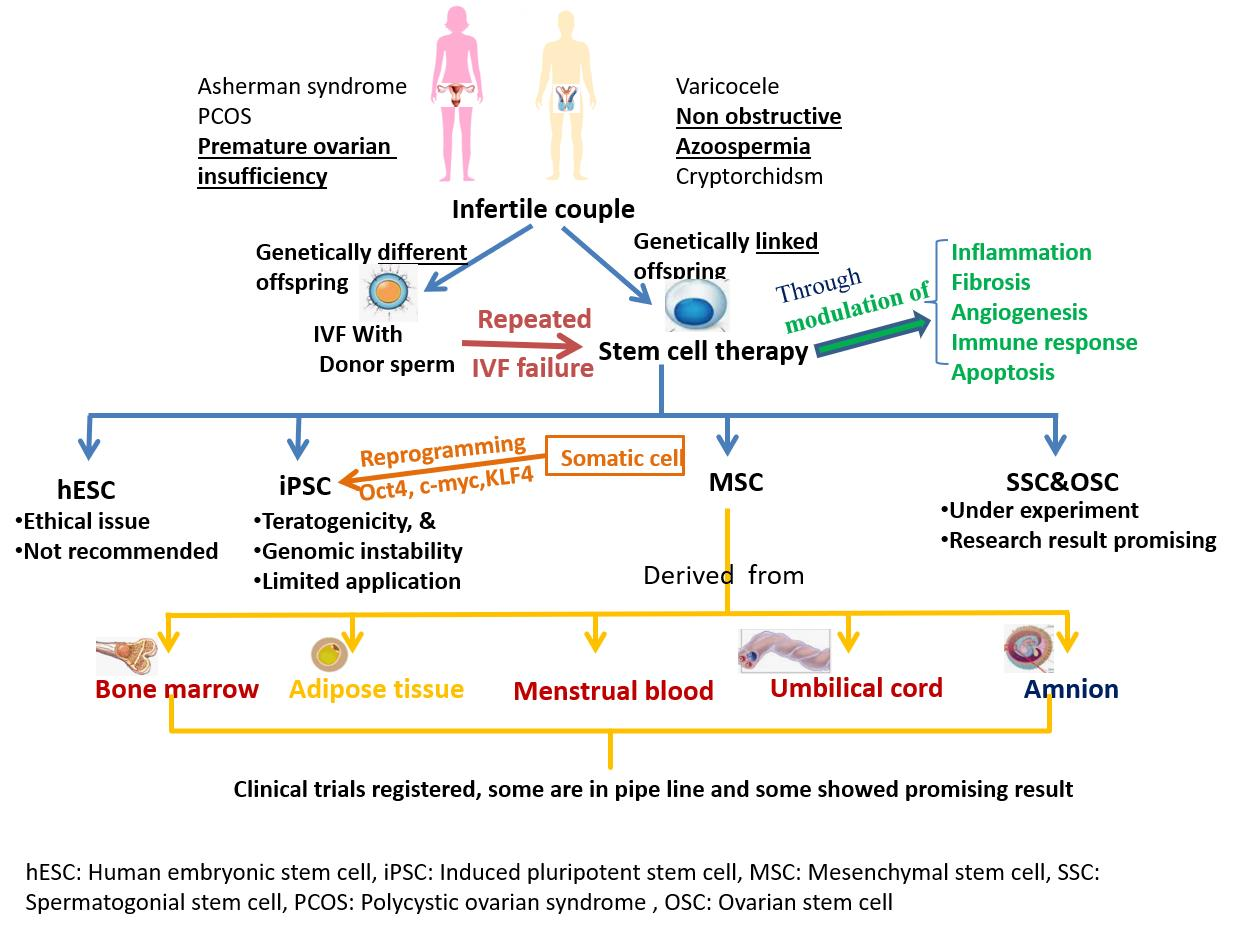
5. Implications for Stem Cells in Infertility
5.1. Embryonic Stem Cells (ESC)
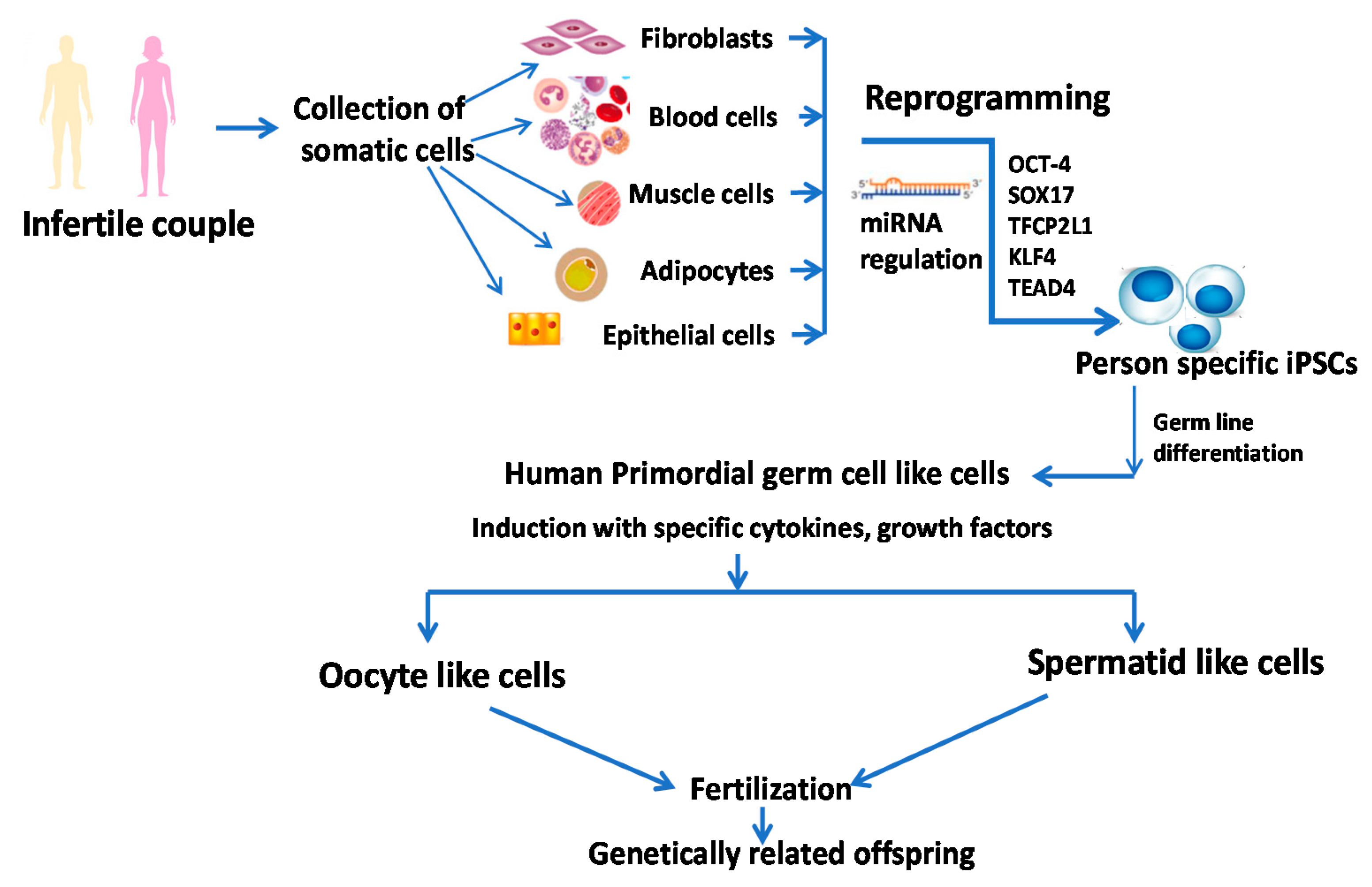
5.2. Induced Pluripotent Stem Cells (iPSC)
5.3. Mesenchymal Stem Cells (MSCs)
5.3.1. Bone Marrow Mesenchymal Stem Cells (BMSC)
5.3.2. Menstrual Blood Mesenchymal Stem Cell (MB-MSC)
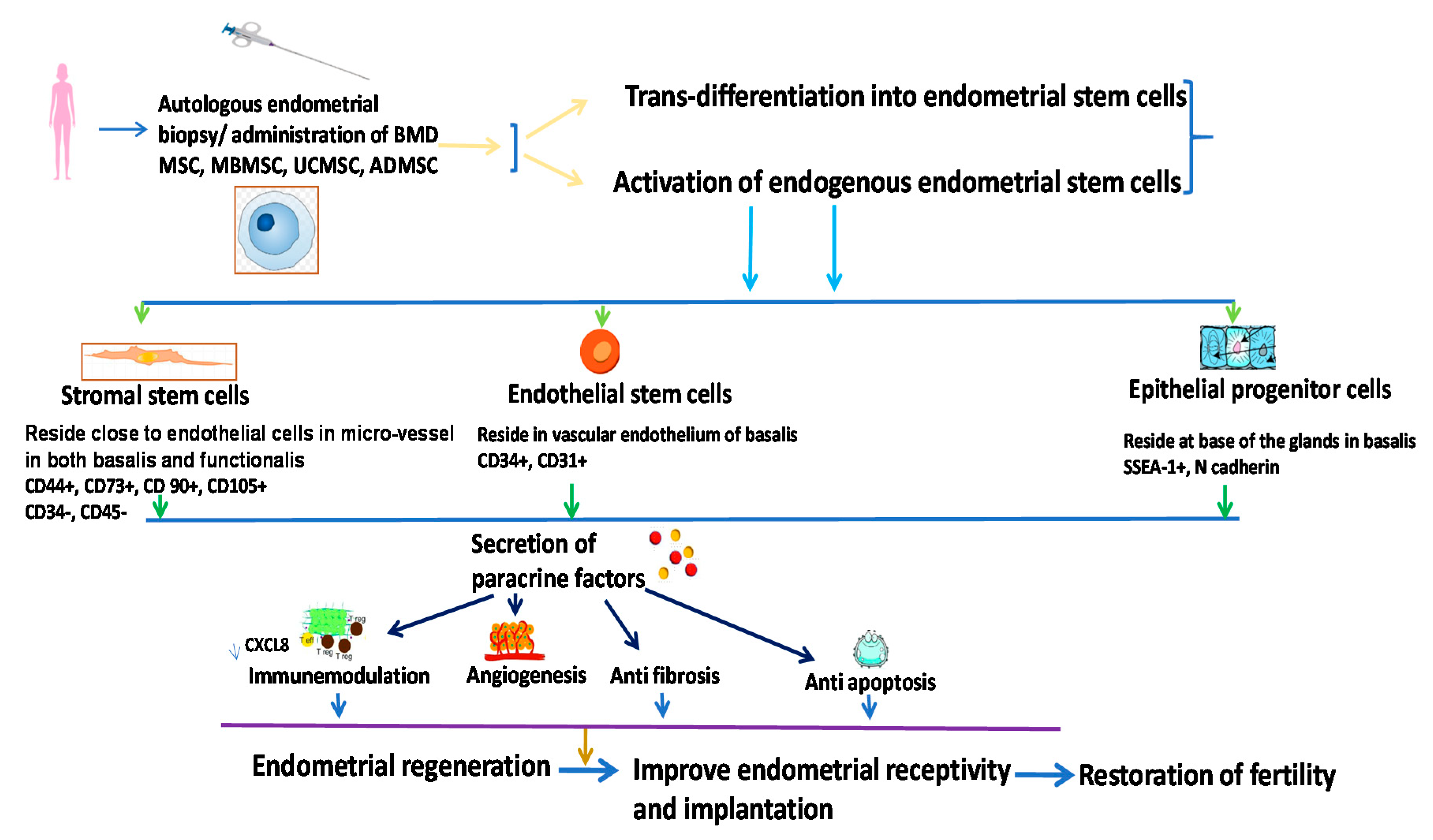
5.3.3. Endometrial Stem Cells (EndSCs)
5.3.4. Umbilical Cord Mesenchymal Stem Cells (UC-MSCs)
5.3.5. Amniotic Fluid Stem Cells (AFSCs)
5.3.6. Amnion-Derived Mesenchymal Stem Cells (AmDMSCs)
5.3.7. Placenta-Derived Mesenchymal Stem Cells (PDMSCs)
5.3.8. Adipose-Tissue-Derived Stem Cells (AD MSC)
5.4. Ovarian Stem Cells (OSC)
5.5. Spermatogonial Stem Cell (SSC)
6. Stem Cell Therapy in Some Known Syndromes
6.1. Asherman Syndrome (AS)
6.2. Premature Ovarian Insufficiency (POI)
6.3. Polycystic Ovarian Syndrome (PCOS)
6.4. Endometriosis
6.5. Azoospermia
7. Clinical Trials
8. Future Prospects
8.1. Very Small Embryonic Like Stem Cells (VSELs)
8.2. Micro RNA and Stem Cell-Based Therapy
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; De Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D. The international glossary on infertility and fertility care, 2017. Hum. Reprod. 2017, 32, 1786–1801. [Google Scholar] [CrossRef] [Green Version]
- Inhorn, M.C.; Patrizio, P. Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. Hum. Reprod. Update 2015, 21, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Volarevic, V.; Bojic, S.; Nurkovic, J.; Volarevic, A.; Ljujic, B.; Arsenijevic, N.; Lako, M.; Stojkovic, M. Stem cells as new agents for the treatment of infertility: Current and future perspectives and challenges. BioMed Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Chandra, A.; Copen, C.E.; Stephen, E.H. Infertility and Impaired Fecundity in the United States, 1982–2010: Data from the National Survey of Family Growth; US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics: Atlanta, GA, USA, 2013.
- Cabella, W.; Nathan, M. Challenges Posed by Low Fertility in Latin America and the Caribbean; United Nations Population Fund: New York, NY, USA, 2018. [Google Scholar]
- Esfandyari, S.; Chugh, R.M.; Park, H.-S.; Hobeika, E.; Ulin, M.; Al-Hendy, A. Mesenchymal stem cells as a bio organ for treatment of female infertility. Cells 2020, 9, 2253. [Google Scholar] [CrossRef]
- Zhao, Y.-X.; Chen, S.-R.; Su, P.-P.; Huang, F.-H.; Shi, Y.-C.; Shi, Q.-y.; Lin, S. Using mesenchymal stem cells to treat female infertility: An update on female reproductive diseases. Stem Cells Int. 2019. [Google Scholar] [CrossRef]
- de Miguel-Gómez, L.; López-Martínez, S.; Francés-Herrero, E.; Rodríguez-Eguren, A.; Pellicer, A.; Cervelló, I. Stem cells and the endometrium: From the discovery of adult stem cells to pre-clinical models. Cells 2021, 10, 595. [Google Scholar] [CrossRef] [PubMed]
- Rossi, B.V.; Abusief, M.; Missmer, S.A. Modifiable risk factors and infertility: What are the connections? Am. J. Lifestyle Med. 2016, 10, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, T.J.; Vitrikas, K. Evaluation and treatment of infertility. Am. Fam. Physician 2015, 91, 308–314. [Google Scholar]
- Skakkebæk, N.-E.; Meyts, R.-D.; Main, K.M. Testicular dysgenesis syndrome: An increasingly common developmental disorder with environmental aspects: Opinion. Hum. Reprod. 2001, 16, 972–978. [Google Scholar] [CrossRef]
- Bhartiya, D.; Hinduja, I.; Patel, H.; Bhilawadikar, R. Making gametes from pluripotent stem cells–a promising role for very small embryonic-like stem cells. Reprod. Biol. Endocrinol. 2014, 12, 114. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Minase, G.; Okabe, K.; Ueda, H.; Sengoku, K. Male infertility and its genetic causes. J. Obstet. Gynaecol. Res. 2015, 41, 1501–1505. [Google Scholar] [CrossRef] [PubMed]
- Ferlin, A.; Raicu, F.; Gatta, V.; Zuccarello, D.; Palka, G.; Foresta, C. Male infertility: Role of genetic background. Reprod. Biomed. Online 2007, 14, 734–745. [Google Scholar] [CrossRef]
- O’brien, K.L.F.; Varghese, A.C.; Agarwal, A. The genetic causes of male factor infertility: A review. Fertil. Steril. 2010, 93, 1–12. [Google Scholar]
- National Collaborating Centre for Women’s; Children’s Health (UK). Fertility: Assessment and Treatment for People with Fertility Problems; National Collaborating Centre for Women’s; Children’s Health (UK): London, UK, 2013. [Google Scholar]
- Burns, L.H. Psychiatric aspects of infertility and infertility treatments. Psychiatr. Clin. N. Am. 2007, 30, 689–716. [Google Scholar] [CrossRef] [PubMed]
- Kroese, A.C.; de Lange, N.M.; Collins, J.; Evers, J.L. Surgery or embolization for varicoceles in subfertile men. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, A.M.; Abou-Setta, A.M.; Al-Inany, H.G. Gonadotrophins for idiopathic male factor subfertility. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Showell, M.G.; Mackenzie-Proctor, R.; Brown, J.; Yazdani, A.; Stankiewicz, M.T.; Hart, R.J. Antioxidants for male subfertility. Cochrane Database Syst. Rev. 2014. [Google Scholar] [CrossRef]
- Ayeleke, R.O.; Asseler, J.D.; Cohlen, B.J.; Veltman, S.M. Intra-uterine insemination for unexplained subfertility. Cochrane Database Syst. Rev. 2020. [Google Scholar] [CrossRef]
- Shapiro, A.; Kashani, B.; Seungdamrong, A. Oral ovulation induction agents. Top. Obstet. Gynecol. 2017, 37, 1–7. [Google Scholar]
- Wang, A.T.; Mullan, R.J.; Lane, M.A.; Hazem, A.; Prasad, C.; Gathaiya, N.W.; Fernández-Balsells, M.M.; Bagatto, A.; Coto-Yglesias, F.; Carey, J. Treatment of hyperprolactinemia: A systematic review and meta-analysis. Syst. Rev. 2012, 1, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodson, W.C.; Haney, A. Controlled ovarian hyperstimulation and intrauterine insemination for treatment of infertility. Fertil. Steril. 1991, 55, 457–467. [Google Scholar] [CrossRef]
- Tsen, L.C.; Vincent, R.D., Jr. In Vitro fertilization and other assisted reproductive technology. Chestnut’s Obstet. Anesth.: Princ. Pract. E-Book 2014, 326, 1328. [Google Scholar]
- Pearlstone, A.C.; Fournet, N.; Gambone, J.C.; Pang, S.C.; Buyalos, R.P. Ovulation induction in women age 40 and older: The importance of basal follicle-stimulating hormone level and chronological age. Fertil. Steril. 1992, 58, 674–679. [Google Scholar] [CrossRef]
- Wikland, M.; Enk, L.; Hammarberg, K.; Nilsson, L. Use of a vaginal transducer for oocyte retrieval in an IVF/ET program. J. Clin. Ultrasound 1987, 15, 245–251. [Google Scholar] [CrossRef]
- Veeck, L. Preembryo grading. Atlas Hum. Oocyte Early Conceptus 1991, 2, 121–149. [Google Scholar]
- Van Steirteghem, A.C.; Nagy, Z.; Joris, H.; Liu, J.; Staessen, C.; Smitz, J.; Wisanto, A.; Devroey, P. High fertilization and implantation rates after intracytoplasmic sperm injection. Hum. Reprod. 1993, 8, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.; Klonoff-Cohen, H. What factors affect intracytoplasmic sperm injection outcomes? Obstet. Gynecol. Surv. 2005, 60, 111–123. [Google Scholar] [CrossRef]
- Hammadeh, M.E.; Fischer-Hammadeh, C.; Ali, K.R. Assisted hatching in assisted reproduction: A state of the art. J. Assist. Reprod. Genet. 2011, 28, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Hellebaut, S.; de Sutter, P.; Dozortsev, D.; Onghena, A.; Qian, C.; Dhont, M. Does assisted hatching improve implantation rates after in vitro fertilization or intracytoplasmic sperm injection in all patients? A prospective randomized study. J. Assist. Reprod. Genet. 1996, 13, 19–22. [Google Scholar] [CrossRef]
- Sallam, H.N.; Sadek, S.S.; Agameya, A.F. Assisted hatching—A meta-analysis of randomized controlled trials. J. Assist. Reprod. Genet. 2003, 20, 332–342. [Google Scholar] [CrossRef]
- Fang, F.; Li, Z.; Zhao, Q.; Li, H.; Xiong, C. Human induced pluripotent stem cells and male infertility: An overview of current progress and perspectives. Hum. Reprod. 2018, 33, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Abd-Allah, S.H.; Shalaby, S.M.; Pasha, H.F.; Amal, S.; Raafat, N.; Shabrawy, S.M.; Awad, H.A.; Amer, M.G.; Gharib, M.A.; El Gendy, E.A. Mechanistic action of mesenchymal stem cell injection in the treatment of chemically induced ovarian failure in rabbits. Cytotherapy 2013, 15, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, S.A.; Shalaby, S.M.; Abdelaziz, M.; Brakta, S.; Hill, W.D.; Ismail, N.; Al-Hendy, A. Human mesenchymal stem cells partially reverse infertility in chemotherapy-induced ovarian failure. Reprod. Sci. 2018, 25, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, X.; Cabanillas, S.; Cervello, I.; Arbona, C.; Raga, F.; Ferro, J.; Palmero, J.; Remohí, J.; Pellicer, A.; Simón, C. Autologous cell therapy with CD133+ bone marrow-derived stem cells for refractory Asherman’s syndrome and endometrial atrophy: A pilot cohort study. Hum. Reprod. 2016, 31, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manshadi, M.D.; Navid, S.; Hoshino, Y.; Daneshi, E.; Noory, P.; Abbasi, M. The effects of human menstrual blood stem cells-derived granulosa cells on ovarian follicle formation in a rat model of premature ovarian failure. Microsc. Res. Tech. 2019, 82, 635–642. [Google Scholar] [CrossRef]
- Domnina, A.; Novikova, P.; Obidina, J.; Fridlyanskaya, I.; Alekseenko, L.; Kozhukharova, I.; Lyublinskaya, O.; Zenin, V.; Nikolsky, N. Human mesenchymal stem cells in spheroids improve fertility in model animals with damaged endometrium. Stem Cell Res. Ther. 2018, 9, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Yu, L.; Sun, M.; Mu, S.; Wang, C.; Wang, D.; Yao, Y. The therapeutic potential of umbilical cord mesenchymal stem cells in mice premature ovarian failure. Biomed Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Wu, S.; Ye, S.; Wang, W.; Guo, X.; Liu, Z. Umbilical cord mesenchyme stem cell local intramuscular injection for treatment of uterine niche: Protocol for a prospective, randomized, double-blinded, placebo-controlled clinical trial. Medicine 2017, 96, e8480. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, M.; Zhang, Y.; Li, W.; Yang, B. Mesenchymal stem cells derived from Wharton jelly of the human umbilical cord ameliorate damage to human endometrial stromal cells. Fertil. Steril. 2011, 96, 1029–1036.E4. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.F.; Hu, H.B.; Xu, H.Y.; Fu, X.F.; Peng, D.X.; Su, W.Y.; He, Y.L. Human umbilical cord mesenchymal stem cell transplantation restores damaged ovaries. J. Cell. Mol. Med. 2015, 19, 2108–2117. [Google Scholar] [CrossRef]
- Wang, J.; Liu, C.; Fujino, M.; Tong, G.; Zhang, Q.; Li, X.-K.; Yan, H. Stem cells as a resource for treatment of infertility-related diseases. Curr. Mol. Med. 2019, 19, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, C.E.; Telfer, E.E.; Anderson, R.A. Ovarian stem cells—Potential roles in infertility treatment and fertility preservation. Maturitas 2013, 76, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Desai, N.; Rambhia, P.; Gishto, A. Human embryonic stem cell cultivation: Historical perspective and evolution of xeno-free culture systems. Reprod. Biol. Endocrinol. 2015, 13, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kehler, J.; Hübner, K.; Garrett, S.; Schöler, H.R. Generating oocytes and sperm from embryonic stem cells. Semin. Reprod. Med. 2005, 23, 222–233. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhou, Q.; Wan, H.; Shen, B.; Wang, X.; Wang, M.; Feng, C.; Xie, M.; Gu, T.; Zhou, T. Generation of fertile offspring from Kit w/Kit wv mice through differentiation of gene corrected nuclear transfer embryonic stem cells. Cell Res. 2015, 25, 851–863. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Hou, J.; Yang, S.; Yang, H.; Liu, Y.; Liu, Y.; Hai, Y.; Chen, Z.; Guo, Y.; Gong, Y.; Gao, W.-Q. Generation of male differentiated germ cells from various types of stem cells. Reproduction 2014, 147, R179–R188. [Google Scholar] [CrossRef] [Green Version]
- Eguizabal, C.; Montserrat, N.; Vassena, R.; Barragan, M.; Garreta, E.; Garcia-Quevedo, L.; Vidal, F.; Giorgetti, A.; Veiga, A.; Belmonte, J.I. Complete meiosis from human induced pluripotent stem cells. Stem Cells 2011, 29, 1186–1195. [Google Scholar] [CrossRef]
- Ramathal, C.; Durruthy-Durruthy, J.; Sukhwani, M.; Arakaki, J.E.; Turek, P.J.; Orwig, K.E.; Pera, R.A.R. Fate of iPSCs derived from azoospermic and fertile men following xenotransplantation to murine seminiferous tubules. Cell Rep. 2014, 7, 1284–1297. [Google Scholar] [CrossRef] [Green Version]
- Irie, N.; Weinberger, L.; Tang, W.W.; Kobayashi, T.; Viukov, S.; Manor, Y.S.; Dietmann, S.; Hanna, J.H.; Surani, M.A. SOX17 is a critical specifier of human primordial germ cell fate. Cell 2015, 160, 253–268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, K.; Yokobayashi, S.; Nakamura, T.; Okamoto, I.; Yabuta, Y.; Kurimoto, K.; Ohta, H.; Moritoki, Y.; Iwatani, C.; Tsuchiya, H. Robust In Vitro induction of human germ cell fate from pluripotent stem cells. Cell Stem Cell 2015, 17, 178–194. [Google Scholar] [CrossRef] [Green Version]
- Easley, C.A., IV; Phillips, B.T.; McGuire, M.M.; Barringer, J.M.; Valli, H.; Hermann, B.P.; Simerly, C.R.; Rajkovic, A.; Miki, T.; Orwig, K.E. Direct differentiation of human pluripotent stem cells into haploid spermatogenic cells. Cell Rep. 2012, 2, 440–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashiro, C.; Sasaki, K.; Yabuta, Y.; Kojima, Y.; Nakamura, T.; Okamoto, I.; Yokobayashi, S.; Murase, Y.; Ishikura, Y.; Shirane, K.; et al. Generation of human oogonia from induced pluripotent stem cells In Vitro. Science 2018, 362, 356–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gell, J.J.; Clark, A.T. Restoring fertility with human induced pluripotent stem cells: Are we there yet? Cell Stem Cell 2018, 23, 777–779. [Google Scholar] [CrossRef] [Green Version]
- Sugawa, F.; Araúzo-Bravo, M.J.; Yoon, J.; Kim, K.P.; Aramaki, S.; Wu, G.; Stehling, M.; Psathaki, O.E.; Hübner, K.; Schöler, H.R. Human primordial germ cell commitment in vitro associates with a unique PRDM14 expression profile. EMBO J. 2015, 34, 1009–1024. [Google Scholar] [CrossRef]
- Bharti, D.; Jang, S.-J.; Lee, S.-Y.; Lee, S.-L.; Rho, G.-J. In Vitro generation of oocyte like cells and their In VIvo efficacy: How far we have been succeeded. Cells 2020, 9, 557. [Google Scholar] [CrossRef] [Green Version]
- Lee, A.S.; Tang, C.; Rao, M.S.; Weissman, I.L.; Wu, J.C. Tumorigenicity as a clinical hurdle for pluripotent stem cell therapies. Nat. Med. 2013, 19, 998–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Deng, G.; Sai, X.; Guo, H.; Huang, H.; Zhu, P. Maturation strategies and limitations of induced pluripotent stem cell-derived cardiomyocytes. Biosci. Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, T.d.S.; de Souza Fernandez, C.; Mencalha, A.L. Human induced pluripotent stem cells from basic research to potential clinical applications in cancer. BioMed Res. Int. 2013. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical trials with mesenchymal stem cells: An update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galipeau, J.; Sensébé, L. Mesenchymal stromal cells: Clinical challenges and therapeutic opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trounson, A.; McDonald, C. Stem cell therapies in clinical trials: Progress and challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehrer, C.; Lepperdinger, G. Mesenchymal stem cell aging. Exp. Gerontol. 2005, 40, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.-M.; Yen, M.-L.; Liu, K.-J.; Sytwu, H.-K.; Yen, B.-L. Immunomodulatory properties of human adult and fetal multipotent mesenchymal stem cells. J. Biomed. Sci. 2011, 18, 49. [Google Scholar] [CrossRef] [Green Version]
- Guillot, P.V.; Gotherstrom, C.; Chan, J.; Kurata, H.; Fisk, N.M. Human first-trimester fetal MSC express pluripotency markers and grow faster and have longer telomeres than adult MSC. Stem Cells 2007, 25, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.; Dickinson, S.C.; Guillot, P.V.; Polak, J.; Blom, A.W.; Kafienah, W.; Hollander, A.P. Human fetal and adult bone marrow-derived mesenchymal stem cells use different signaling pathways for the initiation of chondrogenesis. Stem Cells Dev. 2014, 23, 541–554. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.; Qian, C.; Ding, C.; Meng, Q.; Zou, Q.; Li, H. Fetal liver mesenchymal stem cells restore ovarian function in premature ovarian insufficiency by targeting MT1. Stem Cell Res. Ther. 2019, 10, 362. [Google Scholar] [CrossRef]
- Owen, M.; Friedenstein, A. Stromal stem cells: Marrow-derived osteogenic precursors. Ciba Found. Symp. 1988, 136, 42–60. [Google Scholar]
- Gnecchi, M.; Melo, L.G. Bone marrow-derived mesenchymal stem cells: Isolation, expansion, characterization, viral transduction, and production of conditioned medium. In Stem Cells in Regenerative Medicine; Springer: Berlin/Heidelberg, Germany, 2009; pp. 281–294. [Google Scholar]
- Gao, L.; Huang, Z.; Lin, H.; Tian, Y.; Li, P.; Lin, S. Bone marrow mesenchymal stem cells (BMSCs) restore functional endometrium in the rat model for severe Asherman syndrome. Reprod. Sci. 2019, 26, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Besikcioglu, H.E.; Sarıbas, G.S.; Ozogul, C.; Tiryaki, M.; Kilic, S.; Pınarlı, F.A.; Gulbahar, O. Determination of the effects of bone marrow derived mesenchymal stem cells and ovarian stromal stem cells on follicular maturation in cyclophosphamide induced ovarian failure in rats. Taiwan. J. Obstet. Gynecol. 2019, 58, 53–59. [Google Scholar] [CrossRef]
- Jing, Z.; Qiong, Z.; Yonggang, W.; Yanping, L. Rat bone marrow mesenchymal stem cells improve regeneration of thin endometrium in rat. Fertil. Steril. 2014, 101, 587–594.E3. [Google Scholar] [CrossRef]
- Wang, J.; Ju, B.; Pan, C.; Gu, Y.; Zhang, Y.; Sun, L.; Zhang, B.; Zhang, Y. Application of bone marrow-derived mesenchymal stem cells in the treatment of intrauterine adhesions in rats. Cell. Physiol. Biochem. 2016, 39, 1553–1560. [Google Scholar] [CrossRef]
- Liu, T.; Huang, Y.; Zhang, J.; Qin, W.; Chi, H.; Chen, J.; Yu, Z.; Chen, C. Transplantation of human menstrual blood stem cells to treat premature ovarian failure in mouse model. Stem Cells Dev. 2014, 23, 1548–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, S.X.; Wang, J.; Wang, X.L.; Ali, A.; Wu, L.M.; Liu, Y.S. Feasibility analysis of treating severe intrauterine adhesions by transplanting menstrual blood-derived stem cells. Int. J. Mol. Med. 2018, 41, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, P.; Yuan, Z.; Tan, J. Platelet-rich plasma improves therapeutic effects of menstrual blood-derived stromal cells in rat model of intrauterine adhesion. Stem Cell Res. Ther. 2019, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Guo, F.; Yuan, Q.; Shao, Y.; Zhang, Y.; Wang, H.; Hao, S.; Du, X. Endometrial mesenchymal stem cells isolated from menstrual blood repaired epirubicin-induced damage to human ovarian granulosa cells by inhibiting the expression of Gadd45b in cell cycle pathway. Stem Cell Res. Ther. 2019, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, P.; Wang, Q.; Li, Y.; Li, X.; Zhao, D.; Xu, X.; Kong, L. Autologous menstrual blood-derived stromal cells transplantation for severe Asherman’s syndrome. Hum. Reprod. 2016, 31, 2723–2729. [Google Scholar] [CrossRef] [Green Version]
- Prianishnikov, V. On the concept of stem cell and a model of functional-morphological structure of the endometrium. Contraception 1978, 18, 213–223. [Google Scholar] [CrossRef]
- Chan, R.W.; Schwab, K.E.; Gargett, C.E. Clonogenicity of human endometrial epithelial and stromal cells. Biol. Reprod. 2004, 70, 1738–1750. [Google Scholar] [CrossRef]
- Gargett, C.E.; Schwab, K.E.; Zillwood, R.M.; Nguyen, H.P.; Wu, D. Isolation and culture of epithelial progenitors and mesenchymal stem cells from human endometrium. Biol. Reprod. 2009, 80, 1136–1145. [Google Scholar] [CrossRef] [Green Version]
- Lapidot, T.; Petit, I. Current understanding of stem cell mobilization: The roles of chemokines, proteolytic enzymes, adhesion molecules, cytokines, and stromal cells. Exp. Hematol. 2002, 30, 973–981. [Google Scholar] [CrossRef]
- Azizi, R.; Aghebati-Maleki, L.; Nouri, M.; Marofi, F.; Negargar, S.; Yousefi, M. Stem cell therapy in Asherman syndrome and thin endometrium: Stem cell-based therapy. Biomed. Pharmacother. 2018, 102, 333–343. [Google Scholar] [CrossRef]
- Kato, K.; Yoshimoto, M.; Kato, K.; Adachi, S.; Yamayoshi, A.; Arima, T.; Asanoma, K.; Kyo, S.; Nakahata, T.; Wake, N. Characterization of side-population cells in human normal endometrium. Hum. Reprod. 2007, 22, 1214–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Ichim, T.E.; Zhong, J.; Rogers, A.; Yin, Z.; Jackson, J.; Wang, H.; Ge, W.; Bogin, V.; Chan, K.W. Endometrial regenerative cells: A novel stem cell population. J. Transl. Med. 2007, 5, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargett, C.E.; Rogers, P.A. Human endometrial angiogenesis. Reprod.-Camb. 2001, 121, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, K.; Hosono, K.; Numao, A.; Honda, M.; Matsuda, H.; Amano, H.; Kawauchi, H.; Shibuya, M.; Unno, N.; Majima, M. Vascular endothelial growth factor regulates growth of endometrial tissues and angiogenesis in a mouse transplantation model. Kitasato Med. J. 2016, 46, 15–23. [Google Scholar]
- Wolff, E.F.; Mutlu, L.; Massasa, E.E.; Elsworth, J.D.; Eugene Redmond, D., Jr.; Taylor, H.S. Endometrial stem cell transplantation in MPTP-exposed primates: An alternative cell source for treatment of P arkinson’s disease. J. Cell. Mol. Med. 2015, 19, 249–256. [Google Scholar] [CrossRef]
- Santamaria, X.; Massasa, E.E.; Feng, Y.; Wolff, E.; Taylor, H.S. Derivation of insulin producing cells from human endometrial stromal stem cells and use in the treatment of murine diabetes. Mol. Ther. 2011, 19, 2065–2071. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, S.A.; Shalaby, S.; Brakta, S.; Elam, L.; Elsharoud, A.; Al-Hendy, A. Umbilical cord blood mesenchymal stem cells as an infertility treatment for chemotherapy induced premature ovarian insufficiency. Biomedicines 2019, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Song, D.; Zhong, Y.; Qian, C.; Zou, Q.; Ou, J.; Shi, Y.; Gao, L.; Wang, G.; Liu, Z.; Li, H. Human umbilical cord mesenchymal stem cells therapy in cyclophosphamide-induced premature ovarian failure rat model. BioMed Res. Int. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elfayomy, A.K.; Almasry, S.M.; El-Tarhouny, S.A.; Eldomiaty, M.A. Human umbilical cord blood-mesenchymal stem cells transplantation renovates the ovarian surface epithelium in a rat model of premature ovarian failure: Possible direct and indirect effects. Tissue Cell 2016, 48, 370–382. [Google Scholar] [CrossRef]
- Xie, Q.; Xiong, X.; Xiao, N.; He, K.; Chen, M.; Peng, J.; Su, X.; Mei, H.; Dai, Y.; Wei, D. Mesenchymal stem cells alleviate DHEA-Induced polycystic ovary syndrome (PCOS) by inhibiting inflammation in mice. Stem Cells Int. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C. The Roles of Different Stem Cells in Premature Ovarian Failure. Curr. Stem Cell Res. Ther. 2020, 15, 473–481. [Google Scholar] [CrossRef]
- Shi, Q.; Gao, J.; Jiang, Y.; Sun, B.; Lu, W.; Su, M.; Xu, Y.; Yang, X.; Zhang, Y. Differentiation of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells into endometrial cells. Stem Cell Res. Ther. 2017, 8, 246. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, Y.; Guan, C.-Y.; Tian, S.; Lv, X.-D.; Li, J.-H.; Ma, X.; Xia, H.-F. Therapeutic effect of human umbilical cord-derived mesenchymal stem cells on injured rat endometrium during its chronic phase. Stem Cell Res. Ther. 2018, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Ding, L.; Wang, L.; Cao, Y.; Zhu, H.; Lu, J.; Song, T.; Hu, Y.; Dai, J. Umbilical cord-derived mesenchymal stem cells on scaffolds facilitate collagen degradation via upregulation of MMP-9 in rat uterine scars. Stem Cell Res. Ther. 2017, 8, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, L.; Yan, G.; Wang, B.; Xu, L.; Gu, Y.; Ru, T.; Cui, X.; Lei, L.; Liu, J.; Sheng, X. Transplantation of UC-MSCs on collagen scaffold activates follicles in dormant ovaries of POF patients with long history of infertility. Sci. China Life Sci. 2018, 61, 1554–1565. [Google Scholar] [CrossRef]
- Steigman, S.A.; Ahmed, A.; Shanti, R.M.; Tuan, R.S.; Valim, C.; Fauza, D.O. Sternal repair with bone grafts engineered from amniotic mesenchymal stem cells. J. Pediatric Surg. 2009, 44, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Kunisaki, S.M.; Fuchs, J.R.; Kaviani, A.; Oh, J.-T.; LaVan, D.A.; Vacanti, J.P.; Wilson, J.M.; Fauza, D.O. Diaphragmatic repair through fetal tissue engineering: A comparison between mesenchymal amniocyte–and myoblast-based constructs. J. Pediatric Surg. 2006, 41, 34–39. [Google Scholar] [CrossRef]
- Xiao, G.-Y.; Liu, I.-H.; Cheng, C.-C.; Chang, C.-C.; Lee, Y.-H.; Cheng, W.T.-K.; Wu, S.-C. Amniotic fluid stem cells prevent follicle atresia and rescue fertility of mice with premature ovarian failure induced by chemotherapy. PLoS ONE 2014, 9, e106538. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Huang, Y.; Guo, L.; Cheng, W.; Zou, G. CD44+/CD105+ human amniotic fluid mesenchymal stem cells survive and proliferate in the ovary long-term in a mouse model of chemotherapy-induced premature ovarian failure. Int. J. Med. Sci. 2012, 9, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, L.; Feng, X.; Wei, T.; Wang, Y.; Wang, Y.; Wang, Z.; Tang, D.; Luo, Y.; Xiong, Z. Human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation improves ovarian function in rats with premature ovarian insufficiency (POI) at least partly through a paracrine mechanism. Stem Cell Res. Ther. 2019, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Ling, L.; Zhang, W.; Liu, X.; Wang, Y.; Luo, Y.; Xiong, Z. Effects of human amnion–derived mesenchymal stem cell (hAD-MSC) transplantation In Situ on primary ovarian insufficiency in SD rats. Reprod. Sci. 2020, 27, 1502–1512. [Google Scholar] [CrossRef]
- Ling, L.; Feng, X.; Wei, T.; Wang, Y.; Wang, Y.; Zhang, W.; He, L.; Wang, Z.; Zeng, Q.; Xiong, Z. Effects of low-intensity pulsed ultrasound (LIPUS)-pretreated human amnion-derived mesenchymal stem cell (hAD-MSC) transplantation on primary ovarian insufficiency in rats. Stem Cell Res. Ther. 2017, 8, 283. [Google Scholar] [CrossRef]
- Zhang, H.; Luo, Q.; Lu, X.; Yin, N.; Zhou, D.; Zhang, L.; Zhao, W.; Wang, D.; Du, P.; Hou, Y. Effects of hPMSCs on granulosa cell apoptosis and AMH expression and their role in the restoration of ovary function in premature ovarian failure mice. Stem Cell Res. Ther. 2018, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Yin, N.; Wang, Y.; Lu, X.; Liu, R.; Zhang, L.; Zhao, W.; Yuan, W.; Luo, Q.; Wu, H.; Luan, X. hPMSC transplantation restoring ovarian function in premature ovarian failure mice is associated with change of Th17/Tc17 and Th17/Treg cell ratios through the PI3K/Akt signal pathway. Stem Cell Res. Ther. 2018, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhao, W.; Wang, L.; Luo, Q.; Yin, N.; Lu, X.; Hou, Y.; Cui, J.; Zhang, H. Human placenta-derived mesenchymal stem cells inhibit apoptosis of granulosa cells induced by IRE1α pathway in autoimmune POF mice. Cell Biol. Int. 2019, 43, 899–909. [Google Scholar] [CrossRef]
- Kim, T.-H.; Choi, J.H.; Jun, Y.; Lim, S.M.; Park, S.; Paek, J.-Y.; Lee, S.-H.; Hwang, J.-Y.; Kim, G.J. 3D-cultured human placenta-derived mesenchymal stem cell spheroids enhance ovary function by inducing folliculogenesis. Sci. Rep. 2018, 8, 15313. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.-M.; Qu, L.-P.; Pan, X.-Z.; Mu, L.-S. Upregulated miR-222 targets BCL2L11 and promotes apoptosis of mesenchymal stem cells in preeclampsia patients in response to severe hypoxia. Int. J. Clin. Exp. Pathol. 2018, 11, 110. [Google Scholar] [PubMed]
- Lee, R.H.; Kim, B.; Choi, I.; Kim, H.; Choi, H.S.; Suh, K.; Bae, Y.C.; Jung, J.S. Characterization and expression analysis of mesenchymal stem cells from human bone marrow and adipose tissue. Cell. Physiol. Biochem. 2004, 14, 311–324. [Google Scholar] [CrossRef]
- Yang, J.-A.; Chung, H.-M.; Won, C.-H.; Sung, J.-H. Potential application of adipose-derived stem cells and their secretory factors to skin: Discussion from both clinical and industrial viewpoints. Expert Opin. Biol. Ther. 2010, 10, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Ra, J.C.; Jeong, E.C.; Kang, S.K.; Lee, S.J.; Choi, K.H. A prospective, nonrandomized, no placebo-controlled, phase I/II clinical trial assessing the safety and efficacy of intramuscular injection of autologous adipose tissue-derived mesenchymal stem cells in patients with severe buerger’s disease. Cell Med. 2017, 9, 87–102. [Google Scholar] [CrossRef]
- Damous, L.L.; Nakamuta, J.S.; de Carvalho, A.E.S.; Carvalho, K.C.; Soares, J.M., Jr.; de Jesus Simões, M.; Krieger, J.E.; Baracat, E.C. Does adipose tissue-derived stem cell therapy improve graft quality in freshly grafted ovaries? Reprod. Biol. Endocrinol. 2015, 13, 108. [Google Scholar] [CrossRef] [Green Version]
- Sun, M.; Wang, S.; Li, Y.; Yu, L.; Gu, F.; Wang, C.; Yao, Y. Adipose-derived stem cells improved mouse ovary function after chemotherapy-induced ovary failure. Stem Cell Res. Ther. 2013, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Ding, L.; Cheng, J.; Yang, J.; Li, X.; Yan, G.; Sun, H.; Dai, J.; Hu, Y. Transplantation of adipose-derived stem cells combined with collagen scaffolds restores ovarian function in a rat model of premature ovarian insufficiency. Hum. Reprod. 2016, 31, 1075–1086. [Google Scholar] [CrossRef]
- Kilic, S.; Yuksel, B.; Pinarli, F.; Albayrak, A.; Boztok, B.; Delibasi, T. Effect of stem cell application on Asherman syndrome, an experimental rat model. J. Assist. Reprod. Genet. 2014, 31, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.; Canning, J.; Kaneko, T.; Pru, J.K.; Tilly, J.L. Germline stem cells and follicular renewal in the postnatal mammalian ovary. Nature 2004, 428, 145–150. [Google Scholar] [CrossRef] [PubMed]
- White, Y.A.; Woods, D.C.; Takai, Y.; Ishihara, O.; Seki, H.; Tilly, J.L. Oocyte formation by mitotically active germ cells purified from ovaries of reproductive-age women. Nat. Med. 2012, 18, 413. [Google Scholar] [CrossRef] [Green Version]
- Oatley, J.; Hunt, P.A. Of mice and (wo) men: Purified oogonial stem cells from mouse and human ovaries. Biol. Reprod. 2012, 86, 1–2. [Google Scholar] [CrossRef]
- Niikura, Y.; Niikura, T.; Tilly, J.L. Aged mouse ovaries possess rare premeiotic germ cells that can generate oocytes following transplantation into a young host environment. Aging 2009, 1, 971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, R.A.; Wallace, W.H.B. Fertility preservation in girls and young women. Clin. Endocrinol. 2011, 75, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Kita, K.; Watanabe, T.; Ohsaka, K.; Hayashi, H.; Kubota, Y.; Nagashima, Y.; Aoki, I.; Taniguchi, H.; Noce, T.; Inoue, K. Production of functional spermatids from mouse germline stem cells in ectopically reconstituted seminiferous tubules. Biol. Reprod. 2007, 76, 211–217. [Google Scholar] [CrossRef] [Green Version]
- Meistrich, M.L.; Van Beek, M. Spermatogonial Stem Cells. Cell and Molecular Biology of the Testis; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- De Kretser, D.; Baker, H. Infertility in men: Recent advances and continuing controversies. J. Clin. Endocrinol. Metab. 1999, 84, 3443–3450. [Google Scholar]
- Kanatsu-Shinohara, M.; Lee, J.; Inoue, K.; Ogonuki, N.; Miki, H.; Toyokuni, S.; Ikawa, M.; Nakamura, T.; Ogura, A.; Shinohara, T. Pluripotency of a single spermatogonial stem cell in mice. Biol. Reprod. 2008, 78, 681–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLean, D.J. Spermatogonial stem cell transplantation and testicular function. Cell Tissue Res. 2005, 322, 21–31. [Google Scholar] [CrossRef]
- Nagano, M.C. Homing efficiency and proliferation kinetics of male germ line stem cells following transplantation in mice. Biol. Reprod. 2003, 69, 701–707. [Google Scholar] [CrossRef]
- Hamra, F.K.; Schultz, N.; Chapman, K.M.; Grellhesl, D.M.; Cronkhite, J.T.; Hammer, R.E.; Garbers, D.L. Defining the spermatogonial stem cell. Dev. Biol. 2004, 269, 393–410. [Google Scholar] [CrossRef] [Green Version]
- Guan, K.; Nayernia, K.; Maier, L.S.; Wagner, S.; Dressel, R.; Lee, J.H.; Nolte, J.; Wolf, F.; Li, M.; Engel, W. Pluripotency of spermatogonial stem cells from adult mouse testis. Nature 2006, 440, 1199–1203. [Google Scholar] [CrossRef]
- Izadyar, F.; Wong, J.; Maki, C.; Pacchiarotti, J.; Ramos, T.; Howerton, K.; Yuen, C.; Greilach, S.; Zhao, H.H.; Chow, M. Identification and characterization of repopulating spermatogonial stem cells from the adult human testis. Hum. Reprod. 2011, 26, 1296–1306. [Google Scholar] [CrossRef] [Green Version]
- Hermann, B.P.; Sukhwani, M.; Winkler, F.; Pascarella, J.N.; Peters, K.A.; Sheng, Y.; Valli, H.; Rodriguez, M.; Ezzelarab, M.; Dargo, G. Spermatogonial stem cell transplantation into rhesus testes regenerates spermatogenesis producing functional sperm. Cell Stem Cell 2012, 11, 715–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koubova, J.; Menke, D.B.; Zhou, Q.; Capel, B.; Griswold, M.D.; Page, D.C. Retinoic acid regulates sex-specific timing of meiotic initiation in mice. Proc. Natl. Acad. Sci. USA 2006, 103, 2474–2479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. In Seminars in Cell & Developmental Biology; Academic Press: Cambridge, MA, USA; pp. 411–416.
- Byskov, A.G.; Fenger, M.; Westergaard, L.; Andersen, C.Y. Forskolin and the meiosis inducing substance synergistically initiate meiosis in fetal male germ cells. Mol. Reprod. Dev. 1993, 34, 47–52. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Inoue, K.; Ogonuki, N.; Miki, H.; Yoshida, S.; Toyokuni, S.; Lee, J.; Ogura, A.; Shinohara, T. Leukemia inhibitory factor enhances formation of germ cell colonies in neonatal mouse testis culture. Biol. Reprod. 2007, 76, 55–62. [Google Scholar] [CrossRef]
- Zhu, Y.; Hu, H.-L.; Li, P.; Yang, S.; Zhang, W.; Ding, H.; Tian, R.-H.; Ning, Y.; Zhang, L.-L.; Guo, X.-Z. Generation of male germ cells from induced pluripotent stem cells (iPS cells): An in vitro and in vivo study. Asian J. Androl. 2012, 14, 574. [Google Scholar] [CrossRef] [Green Version]
- Easley IV, C.A.; Simerly, C.R.; Schatten, G. Stem cell therapeutic possibilities: Future therapeutic options for male-factor and female-factor infertility? Reprod. Biomed. Online 2013, 27, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Asherman, J.G. Traumatic intra-uterine adhesions. BJOG Int. J. Obstet. Gynaecol. 1950, 57, 892–896. [Google Scholar] [CrossRef]
- Wallach, E.E.; Schenker, J.G.; Margalioth, E.J. Intrauterine adhesions: An updated appraisal. Fertil. Steril. 1982, 37, 593–610. [Google Scholar] [CrossRef]
- Valle, R.F.; Sciarra, J.J. Intrauterine adhesions: Hysteroscopic diagnosis, classification, treatment, and reproductive outcome. Am. J. Obstet. Gynecol. 1988, 158, 1459–1470. [Google Scholar] [CrossRef]
- Sugimoto, O. Diagnostic and therapeutic hysteroscopy for traumatic intrauterine adhesions. Am. J. Obstet. Gynecol. 1978, 131, 539–547. [Google Scholar] [CrossRef]
- Zikopoulos, K.A.; Kolibianakis, E.M.; Platteau, P.; de Munck, L.; Tournaye, H.; Devroey, P.; Camus, M. Live delivery rates in subfertile women with Asherman’s syndrome after hysteroscopic adhesiolysis using the resectoscope or the Versapoint system. Reprod. Biomed. Online 2004, 8, 720–725. [Google Scholar] [CrossRef]
- Alawadhi, F.; Du, H.; Cakmak, H.; Taylor, H.S. Bone marrow-derived stem cell (BMDSC) transplantation improves fertility in a murine model of Asherman’s syndrome. PLoS ONE 2014, 9, e96662. [Google Scholar] [CrossRef]
- Gan, L.; Duan, H.; Xu, Q.; Tang, Y.-Q.; Li, J.-J.; Sun, F.-Q.; Wang, S. Human amniotic mesenchymal stromal cell transplantation improves endometrial regeneration in rodent models of intrauterine adhesions. Cytotherapy 2017, 19, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Nagori, C.B.; Panchal, S.Y.; Patel, H. Endometrial regeneration using autologous adult stem cells followed by conception by in vitro fertilization in a patient of severe Asherman’s syndrome. J. Hum. Reprod. Sci. 2011, 4, 43. [Google Scholar]
- Singh, N.; Mohanty, S.; Seth, T.; Shankar, M.; Bhaskaran, S.; Dharmendra, S. Autologous stem cell transplantation in refractory Asherman’s syndrome: A novel cell based therapy. J. Hum. Reprod. Sci. 2014, 7, 93. [Google Scholar] [CrossRef]
- The ESHRE Guideline Group on POI; Webber, L.; Davies, M.; Anderson, R.; Bartlett, J.; Braat, D.; Cartwright, B.; Cifkova, R.; de Muinck Keizer-Schrama, S.; Hogervorst, E. ESHRE Guideline: Management of women with premature ovarian insufficiency. Hum. Reprod. 2016, 31, 926–937. [Google Scholar]
- Zangmo, R.; Singh, N.; Sharma, J. Diminished ovarian reserve and premature ovarian failure: A review. IVF Lite 2016, 3, 46. [Google Scholar] [CrossRef]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.-h.; Kawamura, N.; Tamura, M.; Hashimoto, S. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Yoshioka, N.; Takae, S.; Sugishita, Y.; Tamura, M.; Hashimoto, S.; Morimoto, Y.; Kawamura, K. Successful fertility preservation following ovarian tissue vitrification in patients with primary ovarian insufficiency. Hum. Reprod. 2015, 30, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Zhai, J.; Yao, G.; Dong, F.; Bu, Z.; Cheng, Y.; Sato, Y.; Hu, L.; Zhang, Y.; Wang, J.; Dai, S. In Vitro activation of follicles and fresh tissue auto-transplantation in primary ovarian insufficiency patients. J. Clin. Endocrinol. Metab. 2016, 101, 4405–4412. [Google Scholar] [CrossRef]
- Legro, R.S.; Arslanian, S.A.; Ehrmann, D.A.; Hoeger, K.M.; Murad, M.H.; Pasquali, R.; Welt, C.K. Diagnosis and treatment of polycystic ovary syndrome: An Endocrine Society clinical practice guideline. J. Clin. Endocrinol. Metab. 2013, 98, 4565–4592. [Google Scholar] [CrossRef] [Green Version]
- Yin, M.; Wang, X.; Yao, G.; Lü, M.; Liang, M.; Sun, Y.; Sun, F. Transactivation of micrornA-320 by microRNA-383 regulates granulosa cell functions by targeting E2F1 and SF-1 proteins. J. Biol. Chem. 2014, 289, 18239–18257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chugh, R.; Ashour, D.; Garcia, N.; Park, H.; Takala, H.; Ismail, N.; McAllister, J.; Al-Hendy, A.; El Andaloussi, A. Towards cell therapy of polycystic ovary syndrome (PCOS): Human mesenchymal stem cells secretome inhibits androgen production by pcos theca cells. Cytotherapy 2019, 21, S81. [Google Scholar] [CrossRef]
- Igboeli, P.; El Andaloussi, A.; Omar, M.; Ali, M.; Laknaur, A.; Al-Hendy, A. Towards stem cell therapy of polycystic ovary syndrome (PCOS): Human mesenchymal stem cells engrafment in letrozole-induced PCOS murine model. Fertil. Steril. 2017, 108, e247–e248. [Google Scholar] [CrossRef]
- Verkauf, B. Incidence, symptoms, and signs of endometriosis in fertile and infertile women. J. Fla. Med Assoc. 1987, 74, 671–675. [Google Scholar]
- The Practice Committee of the American Society for Reproductive Medicine. Endometriosis and infertility: A committee opinion. Fertil. Steril. 2012, 98, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Macer, M.L.; Taylor, H.S. Endometriosis and infertility: A review of the pathogenesis and treatment of endometriosis-associated infertility. Obstet. Gynecol. Clin. 2012, 39, 535–549. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Kodithuwakku, S.P.; Ng, P.-Y.; Chai, J.; Ng, E.H.; Yeung, W.S.; Ho, P.-C.; Lee, K.-F. Excessive ovarian stimulation up-regulates the Wnt-signaling molecule DKK1 in human endometrium and may affect implantation: An In Vitro co-culture study. Hum. Reprod. 2010, 25, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Sallam, H.N.; Garcia-Velasco, J.A.; Dias, S.; Arici, A.; Abou-Setta, A.M. Long-Term pituitary down-regulation before in vitro fertilization (IVF) for women with endometriosis. Cochrane Database Syst. Rev. 2006. [Google Scholar] [CrossRef]
- Benschop, L.; Farquhar, C.; van der Poel, N.; Heineman, M.J. Interventions for women with endometrioma prior to assisted reproductive technology. Cochrane Database Syst. Rev. 2010. [Google Scholar] [CrossRef]
- Cocuzza, M.; Alvarenga, C.; Pagani, R. The epidemiology and etiology of azoospermia. Clinics 2013, 68, 15–26. [Google Scholar] [CrossRef]
- Vij, S.C.; Sabanegh, E., Jr.; Agarwal, A. Biological therapy for non-obstructive azoospermia. Expert Opin. Biol. Ther. 2018, 18, 19–23. [Google Scholar] [CrossRef]
- Kanatsu-Shinohara, M.; Shinohara, T. Spermatogonial stem cell self-renewal and development. Annu. Rev. Cell Dev. Biol. 2013, 29, 163–187. [Google Scholar] [CrossRef]
- Brinster, R.L.; Zimmermann, J.W. Spermatogenesis following male germ-cell transplantation. Proc. Natl. Acad. Sci. USA 1994, 91, 11298–11302. [Google Scholar] [CrossRef] [Green Version]
- Nayernia, K.; Nolte, J.; Michelmann, H.W.; Lee, J.H.; Rathsack, K.; Drusenheimer, N.; Dev, A.; Wulf, G.; Ehrmann, I.E.; Elliott, D.J. In Vitro-differentiated embryonic stem cells give rise to male gametes that can generate offspring mice. Dev. Cell 2006, 11, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, K.; Ohta, H.; Kurimoto, K.; Aramaki, S.; Saitou, M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell 2011, 146, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Igboeli, P.; El Andaloussi, A.; Sheikh, U.; Takala, H.; ElSharoud, A.; McHugh, A.; Gavrilova-Jordan, L.; Levy, S.; Al-Hendy, A. Intraovarian injection of autologous human mesenchymal stem cells increases estrogen production and reduces menopausal symptoms in women with premature ovarian failure: Two case reports and a review of the literature. J. Med. Case Rep. 2020, 14, 108. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Sun, H.; Zhu, H.; Zhu, X.; Tang, X.; Yan, G.; Wang, J.; Bai, D.; Wang, J.; Wang, L. Allogeneic cell therapy using umbilical cord MSCs on collagen scaffolds for patients with recurrent uterine adhesion: A phase I clinical trial. Stem Cell Res. Ther. 2018, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Mashayekhi, M.; Mirzadeh, E.; Chekini, Z.; Ahmadi, F.; Eftekhari-Yazdi, P.; Vesali, S.; Madani, T.; Aghdami, N. Evaluation of safety, feasibility and efficacy of intra-ovarian transplantation of autologous adipose derived mesenchymal stromal cells in idiopathic premature ovarian failure patients: Non-randomized clinical trial, phase I, first in human. J. Ovarian Res. 2021, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Takagi, M.; Milbourne, A.; Champlin, R.E.; Ueno, N.T. Phase II study of gonadotropin-releasing hormone analog for ovarian function preservation in hematopoietic stem cell transplantation patients. Oncologist 2012, 17, 233. [Google Scholar] [CrossRef] [Green Version]
- Haahr, M.K.; Jensen, C.H.; Toyserkani, N.M.; Andersen, D.C.; Damkier, P.; Sørensen, J.A.; Lund, L.; Sheikh, S.P. Safety and potential effect of a single intracavernous injection of autologous adipose-derived regenerative cells in patients with erectile dysfunction following radical prostatectomy: An open-label phase I clinical trial. EBioMedicine 2016, 5, 204–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhartiya, D.; Unni, S.; Parte, S.; Anand, S. Very small embryonic-like stem cells: Implications in reproductive biology. BioMed Res. Int. 2013. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, M.Z.; Kucia, M.; Majka, M.; Reca, R.; Ratajczak, J. Heterogeneous populations of bone marrow stem cells--are we spotting on the same cells from the different angles? Folia Histochem. Et Cytobiol. 2004, 42, 139–146. [Google Scholar]
- Kucia, M.; Reca, R.; Campbell, F.; Zuba-Surma, E.; Majka, M.; Ratajczak, J.; Ratajczak, M. A population of very small embryonic-like (VSEL) CXCR4+ SSEA-1+ Oct-4+ stem cells identified in adult bone marrow. Leukemia 2006, 20, 857–869. [Google Scholar] [CrossRef] [PubMed]
- Bhartiya, D. Pluripotent very small embryonic-like stem cells get discarded during cord blood and bone marrow processing. Stem Cells Dev. 2012, 21, 2563–2564. [Google Scholar] [CrossRef]
- Bhartiya, D.; Kasiviswanathan, S.; Unni, S.K.; Pethe, P.; Dhabalia, J.V.; Patwardhan, S.; Tongaonkar, H.B. Newer insights into premeiotic development of germ cells in adult human testis using Oct-4 as a stem cell marker. J. Histochem. Cytochem. 2010, 58, 1093–1106. [Google Scholar] [CrossRef] [Green Version]
- Parte, S.; Bhartiya, D.; Telang, J.; Daithankar, V.; Salvi, V.; Zaveri, K.; Hinduja, I. Detection, characterization, and spontaneous differentiation In Vitro of very small embryonic-like putative stem cells in adult mammalian ovary. Stem Cells Dev. 2011, 20, 1451–1464. [Google Scholar] [CrossRef]
- Notarianni, E. Reinterpretation of evidence advanced for neo-oogenesis in mammals, in terms of a finite oocyte reserve. J. Ovarian Res. 2011, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Zuckerman, S. The number of oocytes in the mature ovary. Recent Prog. Horm. Res. 1951, 6, 63–109. [Google Scholar]
- Zhang, H.; Zheng, W.; Shen, Y.; Adhikari, D.; Ueno, H.; Liu, K. Experimental evidence showing that no mitotically active female germline progenitors exist in postnatal mouse ovaries. Proc. Natl. Acad. Sci. USA 2012, 109, 12580–12585. [Google Scholar] [CrossRef] [Green Version]
- Kurkure, P.; Prasad, M.; Dhamankar, V.; Bakshi, G. Very small embryonic-like stem cells (VSELs) detected in azoospermic testicular biopsies of adult survivors of childhood cancer. Reprod. Biol. Endocrinol. 2015, 13, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; He, Y.; Wang, X.; Peng, D.; Chen, X.; Li, X.; Wang, Q. Overexpression of miR-21 in stem cells improves ovarian structure and function in rats with chemotherapy-induced ovarian damage by targeting PDCD4 and PTEN to inhibit granulosa cell apoptosis. Stem Cell Res. Ther. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, B.; Ma, Y.; Wang, F.; Hu, L.; Sun, Y. miR-644-5p carried by bone mesenchymal stem cell-derived exosomes targets regulation of p53 to inhibit ovarian granulosa cell apoptosis. Stem Cell Res. Ther. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lin, L.; Sha, C.; Li, T.; Zhao, D.; Wei, H.; Chen, Q.; Liu, Y.; Chen, X.; Xu, W. Bone marrow mesenchymal stem cell-derived exosomal miR-144-5p improves rat ovarian function after chemotherapy-induced ovarian failure by targeting PTEN. Lab. Investig. 2020, 100, 342–352. [Google Scholar] [CrossRef]
- Xiao, G.-Y.; Cheng, C.-C.; Chiang, Y.-S.; Cheng, W.T.-K.; Liu, I.-H.; Wu, S.-C. Exosomal miR-10a derived from amniotic fluid stem cells preserves ovarian follicles after chemotherapy. Sci. Rep. 2016, 6, 23120. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial Identifier | Est. # of Subjects | Status | Site | Conditions | Interventions | Outcome of Trial |
|---|---|---|---|---|---|---|
| NCT04706312 | 12 | Not yet recruiting | Nanjing Medical University | Diminished Ovarian Response | Human Amniotic Mesenchymal Stem Cells (Hamscs) Transplantation | No results posted |
| NCT04676269 | 40 | Recruiting | Indonesia University | Thin Endometrium Infertile Patients | Amnion Bilayer and Stem Cell Combination Therapy | No results posted |
| NCT03207412 | 20 | Unknown | Chongqing Medical University, China | Premature Ovarian Failure | Human Amniotic Epithelial Cells | No results posted |
| NCT02696889 | 3 | Active | University of Illinois at Chicago | Primary Ovarian Insufficiency, Low Ovarian Reserve | Autologous Stem Cell Therapy | Report of 2 cases revealed a significant improvement in clinical features related to POI. There was an increase in size as well as estrogen production in the MSC engrafted ovary [174] |
| NCT02713854 | 240 | Recruiting | The University of Hong Kong | Subfertility | Human Embryonic Stem-Cell-Derived Trophoblastic Spheroid (Bap-Eb) as a Predictive Tool | No results posted |
| NCT03592849 | 50 | Enrolling by invitation | Nanjing Drum Tower Hospital, China | Infertile Women with Thin Endometrium or Endometrial Scarring | Procedure: Collagen Scaffold Loaded with Umbilical-Cord-Derived Mesenchymal Stem Cells Therapy | No results posted |
| NCT03166189 | 46 | Completed | D.O. Ott Research Institute of Obstetrics, Gynecology, Russian Federation | Infertility of Uterine Origin Asherman Syndrome | Biological: Bone Marrow-Derived Msc and Hrt | No Results Posted |
| Other: Hormonal Replacement Therapy | ||||||
| NCT02313415 | 26 | Completed | Nanjing Drum Tower Hospital, China | Infertility with Intrauterine Adhesions | Procedure: Umbilical Cord Mesenchymal Stem Cells | Phase 1 trial revealed that transplantation of clinical grade human UC MSC could improve the proliferative and differentiation efficiency of endometrium [175] |
| NCT02025270 | 100 | Unknown | Al Azhar University, Egypt | Azoospermic Patients | Bone-Marrow-Derived Mesenchymal Stem Cells | No results posted |
| NCT02641769 | 50 | Recruiting | Stem Cells of Arabia, Amman, Jordan | Non-obstructive Azoospermia | Intratesticular Transplantation of Autologous Stem Cells | No results posted |
| NCT02414295 | 1 | Completed | Man Clinic for Andrology and male infertilit, Cairo, Egypt | Klinefelter Syndrome Azoospermia | Mesenchymal Stem Cell Injection | No Results Posted |
| NCT02062931 | 60 | Unknown | Al-Azhar University hospitals, Egypt | Premature Ovarian Failure | Biological: Stem Cell Preparation and Injection | No results posted |
| NCT02603744 | 9 | Unknown | Royan Institute | Premature Ovarian Failure | Intraovarian Injection of Adipose-Derived Stromal Cells (Adscs) | Intraovarian engrafting of ADSCs were found to be safe and feasible and linked to reduction in FSH level [176] |
| NCT02204358 | 30 | Unknown | Nanjing University Medical School | Intrauterine Adhesions, Endometrial Dysplasia | Collagen Scaffold Loaded with Autologous Bone | No results posted |
| Marrow Stem Cells | ||||||
| Testicular Injection of Autologous | ||||||
| Human Bone Marrow | ||||||
| NCT02041910 | 60 | Unknown | Hesham Saeed Elshaer, El-Rayadh Fertility Centre | Azoospermia | Derived Stem Cells | No results posted |
| NCT02151890 | 10 | Completed | Al Azhar University, Cairo, Egypt | Premature Ovarian Failure | Biological: Stem Cell | No results posted |
| NCT02372474 | 112 | Completed | Al Azhar University, Cairo, Egypt | Premature Ovarian Failure | Biological: Stem Cell | No results posted |
| NCT01742533 | 40 | Unknown | Shenzhen People’s Hospital, Shenzhen, Guangdong, China | Premature Ovarian Failure | Biological: Human Umbilical Cord Mesenchymal Stem Cells and Human Cord Blood Mononuclear Cells | No results posted |
| Drug: Hormone Replacement Therapy | ||||||
| NCT03069209 | 50 | Active, not recruiting | Stem Cells Arabia, Amman, Jordan | Premature Ovarian Failure | Biological: Stem Cells | No results posted |
| NCT00429494 | 60 | Completed | UT MD Anderson Cancer Center, United States | Amenorrhea | Procedure: Hematopoietic Stem Cell Transplantation (Hsct) | Phase II trial revealed that Leuprolide could not preserve ovarian function in HSCT patients [177] |
| Premature Ovarian Failure | Drug: Leuprolide Acetate | |||||
| Ovarian Function Insufficiency | Behavioral: Questionnaire | |||||
| NCT04009473 | 100 | Enrolling by invitation | Multicenter | Ovarian Failure | Combination Product: SEGOVA Procedure Includes Stem Cell Therapy, Growth Factor, and Platelet Plasma Rich Therapy | No results posted |
| Premature Ovarian Failure | ||||||
| NCT02240823 | 30 | Unknown | Odense University Hospital | Erectile Dysfunction After Prostatectomy | Adipose-Derived Stem Cells (ADMSC) | Intracavernous injection of ADMSC is a safe procedure and resulted in improvement of erectile function [178] |
| NCT02414308 | 20 | Unknown | Man Clinic for Andrology, Male Infertility, and Sexual Dysfunction | Erectile Dysfunction Peyronie’ Disease | Adipose Tissue Stem Cell Injection | No results posted |
| NCT02008799 | 20 | Recruiting | Man Clinic for Andrology, Male Infertility, and Sexual Dysfunction | Azoospermia | Intratesticular Artery Injection of Bone Marrow Stem Cell | No result posted |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saha, S.; Roy, P.; Corbitt, C.; Kakar, S.S. Application of Stem Cell Therapy for Infertility. Cells 2021, 10, 1613. https://doi.org/10.3390/cells10071613
Saha S, Roy P, Corbitt C, Kakar SS. Application of Stem Cell Therapy for Infertility. Cells. 2021; 10(7):1613. https://doi.org/10.3390/cells10071613
Chicago/Turabian StyleSaha, Sarama, Partha Roy, Cynthia Corbitt, and Sham S. Kakar. 2021. "Application of Stem Cell Therapy for Infertility" Cells 10, no. 7: 1613. https://doi.org/10.3390/cells10071613
APA StyleSaha, S., Roy, P., Corbitt, C., & Kakar, S. S. (2021). Application of Stem Cell Therapy for Infertility. Cells, 10(7), 1613. https://doi.org/10.3390/cells10071613