
The Cell Wall Proteome of Craterostigma plantagineum Cell Cultures Habituated to Dichlobenil and Isoxaben

 ,
,  , , ,
, , ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Establishment of Calli and Cell Suspension Cultures
2.2. Effects of DCB and IXB and Habituation of Cell Suspension Cultures to the Inhibitors
2.3. Quantification of Cell Wall Monosaccharides and Total Lignin
2.4. Gel-Free and Gel-Based Proteomics
2.5. RNA Extraction, cDNA Synthesis, and qPCR
2.6. Statistical Analyses
3. Results
3.1. Phenotypes of Control and Habituated Cells in Response to DCB and IXB
3.2. Biochemical Analyses: Cell Wall Monosaccharide Composition, Total Lignin Content, and Proteomics
3.3. Gene Expression Analysis of Some Candidates Identified with Proteomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartels, D.; Hussain, S.S. Resurrection Plants: Physiology and molecular biology. In Plant Desiccation Tolerance; Ecological Studies; Lüttge, U., Beck, E., Bartels, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 139–156. ISBN 978-3-642-19106-0. [Google Scholar]
- Chen, P.; Jung, N.U.; Giarola, V.; Bartels, D. The Dynamic Responses of Cell Walls in Resurrection Plants during Dehydration and Rehydration. Front. Plant Sci. 2020, 10, 1698. [Google Scholar] [CrossRef] [Green Version]
- Bartels, D.; Schneider, K.; Terstappen, G.; Piatkowski, D.; Salamini, F. Molecular cloning of abscisic acid-modulated genes which are induced during desiccation of the resurrection plant Craterostigma plantagineum. Planta 1990, 181, 27–34. [Google Scholar] [CrossRef]
- Vicré, M.; Sherwin, H.; Driouich, A.; Jaffer, M.; Farrant, J. Cell Wall Characteristics and Structure of Hydrated and Dry leaves of the Resurrection Plant Craterostigma wilmsii, a Microscopical Study. J. Plant Physiol. 1999, 155, 719–726. [Google Scholar] [CrossRef]
- Vicre, M.; Lerouxel, O.; Farrant, J.; Lerouge, P.; Driouich, A. Composition and desiccation-induced alterations of the cell wall in the resurrection plant Craterostigma wilmsii. Physiol. Plant. 2004, 120, 229–239. [Google Scholar] [CrossRef]
- Moore, J.; Nguema-Ona, E.; Chevalier, L.; Lindsey, G.G.; Brandt, W.F.; Lerouge, P.; Farrant, J.; Driouich, A. Response of the Leaf Cell Wall to Desiccation in the Resurrection Plant Myrothamnus flabellifolius. Plant Physiol. 2006, 141, 651–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, J.P.; Farrant, J.; Driouich, A. A role for pectin-associated arabinans in maintaining the flexibility of the plant cell wall during water deficit stress. Plant Signal. Behav. 2008, 3, 102–104. [Google Scholar] [CrossRef] [Green Version]
- Jung, N.U.; Giarola, V.; Chen, P.; Knox, J.P.; Bartels, D. Craterostigma plantagineum cell wall composition is remodelled during desiccation and the glycine-rich protein CpGRP1 interacts with pectins through clustered arginines. Plant J. 2019, 100, 661–676. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.P.; Nguema-Ona, E.E.; Vicré-Gibouin, M.; Sørensen, I.; Willats, W.; Driouich, A.; Farrant, J. Arabinose-rich polymers as an evolutionary strategy to plasticize resurrection plant cell walls against desiccation. Planta 2012, 237, 739–754. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.-M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell Wall Metabolism in Response to Abiotic Stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willats, W.; Orfila, C.; Limberg, G.; Buchholt, H.C.; van Alebeek, G.-J.W.M.; Voragen, A.G.J.; Marcus, S.E.; Christensen, T.M.I.E.; Mikkelsen, J.D.; Murray, B.S.; et al. Modulation of the degree and pattern of methyl-esterification of Pectic Homogalacturonan in plant cell walls: Implications for pectin methyl esterase action, matrix properties, and cell adhesion. J. Biol. Chem. 2001, 276, 19404–19413. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.-C.; Bulgakov, V.; Jinn, T.-L. Pectin Methylesterases: Cell Wall Remodeling Proteins Are Required for Plant Response to Heat Stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moura, J.C.M.S.; Bonine, C.A.V.; Viana, J.D.O.F.; Dornelas, M.C.; Mazzafera, P. Abiotic and Biotic Stresses and Changes in the Lignin Content and Composition in Plants. J. Integr. Plant Biol. 2010, 52, 360–376. [Google Scholar] [CrossRef] [PubMed]
- Hamann, T. The Plant Cell Wall Integrity Maintenance Mechanism—Concepts for Organization and Mode of Action. Plant Cell Physiol. 2014, 56, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, Y.; Dinneny, J.R. A wall with integrity: Surveillance and maintenance of the plant cell wall under stress. New Phytol. 2019, 225, 1428–1439. [Google Scholar] [CrossRef] [Green Version]
- Manfield, I.; Orfila, C.; McCartney, L.; Harholt, J.; Bernal, A.; Scheller, H.; Gilmartin, P.M.; Mikkelsen, J.D.; Knox, P.; Willats, W.G.T. Novel cell wall architecture of isoxaben-habituated Arabidopsis suspension-cultured cells: Global transcript profiling and cellular analysis. Plant J. 2004, 40, 260–275. [Google Scholar] [CrossRef] [PubMed]
- Duval, I.; Beaudoin, N. Transcriptional profiling in response to inhibition of cellulose synthesis by thaxtomin A and isoxaben in Arabidopsis thaliana suspension cells. Plant Cell Rep. 2009, 28, 811–830. [Google Scholar] [CrossRef]
- Wormit, A.; Butt, S.M.; Chairam, I.; McKenna, J.; Nunes-Nesi, A.; Kjaer, L.; O’Donnelly, K.; Fernie, A.R.; Woscholski, R.; Barter, M.L.; et al. Osmosensitive Changes of Carbohydrate Metabolism in Response to Cellulose Biosynthesis Inhibition. Plant Physiol. 2012, 159, 105–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mélida, H.; Encina, A.; Álvarez, J.; Acebes, J.-L.; Caparrós-Ruiz, D. Unraveling the Biochemical and Molecular Networks Involved in Maize Cell Habituation to the Cellulose Biosynthesis Inhibitor Dichlobenil. Mol. Plant 2010, 3, 842–853. [Google Scholar] [CrossRef]
- De Castro, M.; Largo-Gosens, A.; Alvarez, J.M.; García-Angulo, P.; Acebes, J.L. Early cell-wall modifications of maize cell cultures during habituation to dichlobenil. J. Plant Physiol. 2014, 171, 127–135. [Google Scholar] [CrossRef]
- Encina, A.; Moral, R.M.; Acebes, J.-L.; Álvarez, J.M. Characterization of cell walls in bean (Phaseolus vulgaris L.) callus cultures tolerant to dichlobenil. Plant Sci. 2001, 160, 331–339. [Google Scholar] [CrossRef]
- Encina, A.; Sevillano, J.M.; Acebes, J.-L.; Alvarez, J. Cell wall modifications of bean (Phaseolus vulgaris) cell suspensions during habituation and dehabituation to dichlobenil. Physiol. Plant. 2002, 114, 182–191. [Google Scholar] [CrossRef]
- Tateno, M.; Brabham, C.; DeBolt, S. Cellulose biosynthesis inhibitors—A multifunctional toolbox. J. Exp. Bot. 2015, 67, 533–542. [Google Scholar] [CrossRef] [Green Version]
- DeBolt, S.; Gutierrez, R.; Ehrhardt, D.W.; Somerville, C. Nonmotile Cellulose Synthase Subunits Repeatedly Accumulate within Localized Regions at the Plasma Membrane in Arabidopsis Hypocotyl Cells following 2,6-Dichlorobenzonitrile Treatment. Plant Physiol. 2007, 145, 334–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, J.R.; Barnes, W.S.; Bedinger, P. 2,6-Dichlorobenzonitrile, a cellulose biosynthesis inhibitor, affects morphology and structural integrity of Petunia and lily pollen tubes. J. Plant Physiol. 2002, 159, 61–67. [Google Scholar] [CrossRef]
- Lazzaro, M.D.; Donohue, J.M.; Soodavar, F.M. Disruption of cellulose synthesis by isoxaben causes tip swelling and disorganizes cortical microtubules in elongating conifer pollen tubes. Protoplasma 2003, 220, 201–207. [Google Scholar] [CrossRef]
- Díaz-Cacho, P.; Moral, R.; Encina, A.; Acebes, J.-L.; Alvarez, J. Cell wall modifications in bean (Phaseolus vulgaris) callus cultures tolerant to isoxaben. Physiol. Plant. 1999, 107, 54–59. [Google Scholar] [CrossRef]
- Mélida, H.; García-Angulo, P.; Alonso-Simón, A.; Encina, A.; Alvarez, J.; Acebes, J.L. Novel type II cell wall architecture in dichlobenil-habituated maize calluses. Planta 2008, 229, 617–631. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Toldi, O.; Tóth, S.; Pónyi, T.; Scott, P. An effective and reproducible transformation protocol for the model resurrection plant Craterostigma plantagineum Hochst. Plant Cell Rep. 2002, 21, 63–69. [Google Scholar] [CrossRef]
- Pettolino, F.; Walsh, C.; Fincher, G.B.; Bacic, A. Determining the polysaccharide composition of plant cell walls. Nat. Protoc. 2012, 7, 1590–1607. [Google Scholar] [CrossRef]
- Xu, X.; Backes, A.; Legay, S.; Berni, R.; Faleri, C.; Gatti, E.; Hausman, J.-F.; Cai, G.; Guerriero, G. Cell wall composition and transcriptomics in stem tissues of stinging nettle (Urtica dioica L.): Spotlight on a neglected fibre crop. Plant Direct 2019, 3, e00151. [Google Scholar] [CrossRef] [Green Version]
- Behr, M.; Sergeant, K.; Leclercq, C.C.; Planchon, S.; Guignard, C.; Lenouvel, A.; Renaut, J.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Insights into the molecular regulation of monolignol-derived product biosynthesis in the growing hemp hypocotyl. BMC Plant Biol. 2018, 18, 1. [Google Scholar] [CrossRef] [Green Version]
- Backes, A.; Behr, M.; Xu, X.; Gatti, E.; Legay, S.; Predieri, S.; Hausman, J.-F.; Deyholos, M.K.; Cai, G.; Guerriero, G. Sucrose synthase gene expression analysis in the fibre nettle (Urtica dioica L.) cultivar “clone 13”. Ind. Crop. Prod. 2018, 123, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Sergeant, K.; Printz, B.; Guerriero, G.; Renaut, J.; Lutts, S.; Hausman, J.-F. The Dynamics of the Cell Wall Proteome of Developing Alfalfa Stems. Biology 2019, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Legay, S.; Sergeant, K.; Zorzan, S.; Leclercq, C.C.; Charton, S.; Giarola, V.; Liu, X.; Challabathula, D.; Renaut, J.; et al. Molecular insights into plant desiccation tolerance: Transcriptomics, proteomics and targeted metabolite profiling in Craterostigma plantagineum. Plant J. 2021. [Google Scholar] [CrossRef] [PubMed]
- Armenteros, J.J.A.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Csordas, A.; Del-Toro, N.; Dianes, J.A.; Griss, J.; Lavidas, I.; Mayer, G.; Perez-Riverol, Y.; Reisinger, F.; Ternent, T.; et al. 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 2015, 44, D447–D456. [Google Scholar] [CrossRef] [PubMed]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [Green Version]
- Giarola, V.; Challabathula, D.; Bartels, D. Quantification of expression of dehydrin isoforms in the desiccation tolerant plant Craterostigma plantagineum using specifically designed reference genes. Plant Sci. 2015, 236, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Printz, B.; Morais, R.D.S.; Wienkoop, S.; Sergeant, K.; Lutts, S.; Hausman, J.-F.; Renaut, J. An improved protocol to study the plant cell wall proteome. Front. Plant Sci. 2015, 6, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Simón, A.; García-Angulo, P.; Encina, A.; Acebes, J.L.; Álvarez, J. Habituation of Bean (Phaseolus vulgaris) Cell Cultures to Quinclorac and Analysis of the Subsequent Cell Wall Modifications. Ann. Bot. 2008, 101, 1329–1339. [Google Scholar] [CrossRef] [Green Version]
- Brochu, V.; Girard-Martel, M.; Duval, I.; Lerat, S.; Grondin, G.; Domingue, O.; Beaulieu, C.; Beaudoin, N. Habituation to thaxtomin A in hybrid poplar cell suspensions provides enhanced and durable resistance to inhibitors of cellulose synthesis. BMC Plant Biol. 2010, 10, 272. [Google Scholar] [CrossRef] [Green Version]
- Menna, A.; Fischer-Stettler, M.; Pfister, B.; Andrés, G.S.; Holbrook-Smith, D.; Sánchez-Rodríguez, C. Single-run HPLC Quantification of Plant Cell Wall Monosaccharides. Bio-Protocol 2020, 10. [Google Scholar] [CrossRef]
- Lagrimini, L.M.; Burkhart, W.; Moyer, M.; Rothstein, S. Molecular cloning of complementary DNA encoding the lignin-forming peroxidase from tobacco: Molecular analysis and tissue-specific expression. Proc. Natl. Acad. Sci. USA 1987, 84, 7542–7546. [Google Scholar] [CrossRef] [Green Version]
- Kukavica, B.M.; Veljovicć-Jovanovicć, S.D.; Menckhoff, L.; Lüthje, S. Cell wall-bound cationic and anionic class III isoperoxidases of pea root: Biochemical characterization and function in root growth. J. Exp. Bot. 2012, 63, 4631–4645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tegg, R.S.; Shabala, S.N.; Cuin, T.; Davies, N.W.; Wilson, C.R. Enhanced resistance to the cellulose biosynthetic inhibitors, thaxtomin A and isoxaben in Arabidopsis thaliana mutants, also provides specific co-resistance to the auxin transport inhibitor, 1-NPA. BMC Plant Biol. 2013, 13, 76. [Google Scholar] [CrossRef] [Green Version]
- Carpin, S.; Crèvecoeur, M.; Greppin, H.; Penel, C. Molecular Cloning and Tissue-Specific Expression of an Anionic Peroxidase in Zucchini1. Plant Physiol. 1999, 120, 799–810. [Google Scholar] [CrossRef] [Green Version]
- Lagrimini, L.M.; Gingas, V.; Finger, F.; Rothstein, S.; Liu, T. Characterization of Antisense Transformed Plants Deficient in the Tobacco Anionic Peroxidase. Plant Physiol. 1997, 114, 1187–1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cosio, C.; Vuillemin, L.; De Meyer, M.; Kevers, C.; Penel, C.; Dunand, C. An anionic class III peroxidase from zucchini may regulate hypocotyl elongation through its auxin oxidase activity. Planta 2009, 229, 823–836. [Google Scholar] [CrossRef] [Green Version]
- Lehman, T.A.; Sanguinet, K. Auxin and Cell Wall Crosstalk as Revealed by the Arabidopsis thaliana Cellulose Synthase Mutant Radially Swollen 1. Plant Cell Physiol. 2019, 60, 1487–1503. [Google Scholar] [CrossRef]
- Lagrimini, L.M. Wound-Induced Deposition of Polyphenols in Transgenic Plants Overexpressing Peroxidase. Plant Physiol. 1991, 96, 577–583. [Google Scholar] [CrossRef]
- Yang, Y.; Jiang, R.; Wang, H.; Tian, Z.; Xie, C. StPOPA, encoding an anionic peroxidase, enhances potato resistance against Phytophthora infestans. Mol. Breed. 2020, 40, 16. [Google Scholar] [CrossRef]
- Gigli-Bisceglia, N.; Engelsdorf, T.; Hamann, T. Plant cell wall integrity maintenance in model plants and crop species-relevant cell wall components and underlying guiding principles. Cell. Mol. Life Sci. 2019, 77, 2049–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reem, N.T.; Pogorelko, G.; Lionetti, V.; Chambers, L.; Held, M.A.; Bellincampi, D.; Zabotina, O.A. Decreased Polysaccharide Feruloylation Compromises Plant Cell Wall Integrity and Increases Susceptibility to Necrotrophic Fungal Pathogens. Front. Plant Sci. 2016, 7, 630. [Google Scholar] [CrossRef] [Green Version]
- Rayle, D.L.; Cleland, R. The Acid Growth Theory of auxin-induced cell elongation is alive and well. Plant Physiol. 1992, 99, 1271–1274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensel, A.; Brummell, D.; Hanna, R.; MacLachlan, G. Auxin-dependent breakdown of xyloglucan in cotyledons of germinating Nasturtium seeds. Planta 1991, 183, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Du, M.; Spalding, E.P.; Gray, W. Rapid Auxin-Mediated Cell Expansion. Annu. Rev. Plant Biol. 2020, 71, 379–402. [Google Scholar] [CrossRef] [Green Version]
- Majda, M.; Robert, S. The Role of Auxin in Cell Wall Expansion. Int. J. Mol. Sci. 2018, 19, 951. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.C.S.; Edsgard, S.D.; Hussain, S.S.; Alquezar, D.; Rasmussen, M.; Gilbert, M.; Nielsen, H.B.; Bartels, D.; Mundy, J. Transcriptomes of the desiccation-tolerant resurrection plant Craterostigma plantagineum. Plant J. 2010, 63, 212–228. [Google Scholar] [CrossRef]
- Solomon, M.; Belenghi, B.; Delledonne, M.; Menachem, E.; Levine, A. The Involvement of Cysteine Proteases and Protease Inhibitor Genes in the Regulation of Programmed Cell Death in Plants. Plant Cell 1999, 11, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Giarola, V.; Krey, S.; Frerichs, A.; Bartels, D. Taxonomically restricted genes of Craterostigma plantagineum are modulated in their expression during dehydration and rehydration. Planta 2014, 241, 193–208. [Google Scholar] [CrossRef]
- Hamann, T.; Bennett, M.; Mansfield, J.; Somerville, C. Identification of cell-wall stress as a hexose-dependent and osmosensitive regulator of plant responses. Plant J. 2009, 57, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhao, F.; Fang, W.; Xie, D.; Hou, J.; Yang, X.; Zhao, Y.; Tang, Z.; Nie, L.; Lv, S. Identification of early salt stress responsive proteins in seedling roots of upland cotton (Gossypium hirsutum L.) employing iTRAQ-based proteomic technique. Front. Plant Sci. 2015, 6, 732. [Google Scholar] [CrossRef] [Green Version]
- Giarola, V.; Jung, N.U.; Singh, A.; Satpathy, P.; Bartels, D. Analysis of pcC13-62 promoters predicts a link between cis-element variations and desiccation tolerance in Linderniaceae. J. Exp. Bot. 2018, 69, 3773–3784. [Google Scholar] [CrossRef] [PubMed]
- Giarola, V.; Chen, P.; Dulitz, S.J.; König, M.; Manduzio, S.; Bartels, D. The dehydration- and ABA-inducible germin-like protein CpGLP1 from Craterostigma plantagineum has SOD activity and may contribute to cell wall integrity during desiccation. Planta 2020, 252, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Cortés, J.; Mrksich, M. The Platelet Integrin αIIbβ3 Binds to the RGD and AGD Motifs in Fibrinogen. Chem. Biol. 2009, 16, 990–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, N.; Sakurai, N. Increase in the amount of celA1 protein in tobacco BY-2 cells by a cellulose biosynthesis inhibitor, 2,6-dichlorobenzonitrile. Plant Cell Physiol. 1998, 39, 779–785. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, R.; Lindeboom, J.J.; Paredez, A.; Emons, A.M.C.; Ehrhardt, D.W. Arabidopsis cortical microtubules position cellulose synthase delivery to the plasma membrane and interact with cellulose synthase trafficking compartments. Nat. Cell Biol. 2009, 11, 797–806. [Google Scholar] [CrossRef]
- Wu, J.; Steinebrunner, I.; Sun, Y.; Butterfield, T.; Torres, J.; Arnold, D.; Gonzalez, A.; Jacob, F.; Reichler, S.; Roux, S.J. Apyrases (Nucleoside Triphosphate-Diphosphohydrolases) Play a Key Role in Growth Control in Arabidopsis. Plant Physiol. 2007, 144, 961–975. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.H.; Wu, J.; Yao, J.; Gallardo, I.F.; Dugger, J.; Webb, L.J.; Huang, J.; Salmi, M.; Song, J.; Clark, G.; et al. Apyrase Suppression Raises Extracellular ATP Levels and Induces Gene Expression and Cell Wall Changes Characteristic of Stress Responses. Plant Physiol. 2014, 164, 2054–2067. [Google Scholar] [CrossRef] [Green Version]
- Maeda, M.; Kimura, Y. Structural features of free N-glycans occurring in plants and functional features of de-N-glycosylation enzymes, ENGase, and PNGase: The presence of unusual plant complex type N-glycans. Front. Plant Sci. 2014, 5, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.; Kuo, M.-C.; Khoo, K.-H.; Inoue, S.; Inoue, Y. Developmentally Regulated Expression of a Peptide:N-Glycanase during Germination of Rice Seeds (Oryza sativa) and Its Purification and Characterization. J. Biol. Chem. 2000, 275, 129–134. [Google Scholar] [CrossRef] [Green Version]
- Herger, A.; Dünser, K.; Kleine-Vehn, J.; Ringli, C. Leucine-Rich Repeat Extensin Proteins and Their Role in Cell Wall Sensing. Curr. Biol. 2019, 29, R851–R858. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, X.; Lu, C.; Zeng, X.; Li, Y.; Fu, D.; Wu, G. Non-specific lipid transfer proteins in plants: Presenting new advances and an integrated functional analysis. J. Exp. Bot. 2015, 66, 5663–5681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelen, F.A.; Hartog, M.V.; Thomas, T.L.; Taylor, B.; Sturm, A.; Van Kammen, A.; Vries, S.C. The carrot secreted glycoprotein gene EP1 is expressed in the epidermis and has sequence homology to Brassica S-locus glycoproteins. Plant J. 1993, 4, 855–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hove, C.A.T.; Bochdanovits, Z.; Jansweijer, V.M.A.; Koning, F.G.; Berke, L.; Sanchez-Perez, G.; Scheres, B.; Heidstra, R. Probing the roles of LRR RLK genes in Arabidopsis thaliana roots using a custom T-DNA insertion set. Plant Mol. Biol. 2011, 76, 69–83. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Kucukoglu, M.; Zhang, L.; Chen, P.; Decker, D.; Nilsson, O.; Jones, B.; Sandberg, G.; Zheng, B. The Arabidopsis LRR-RLK, PXC1, is a regulator of secondary wall formation correlated with the TDIF-PXY/TDR-WOX4 signaling pathway. BMC Plant Biol. 2013, 13, 94. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
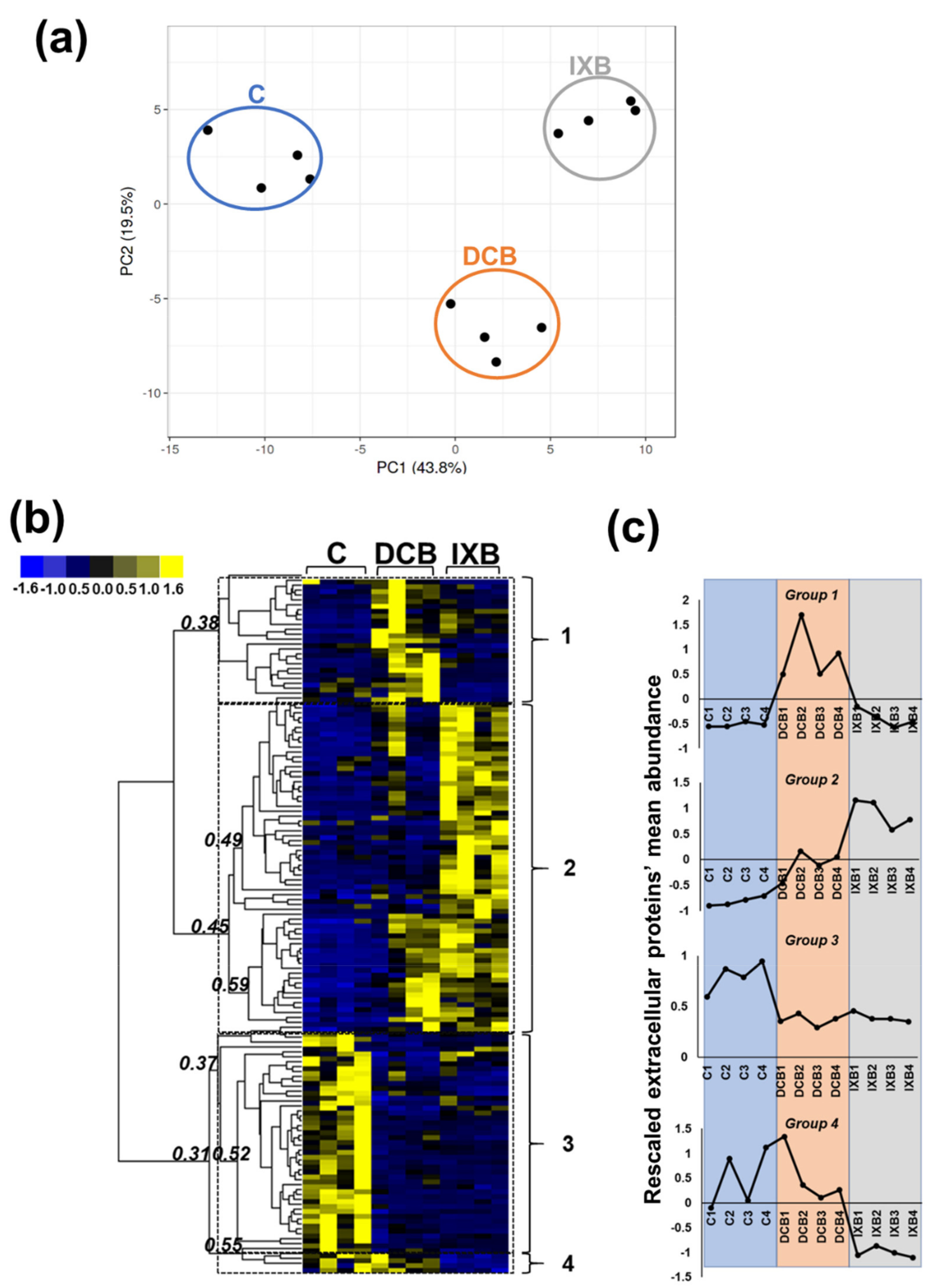{kind=link}
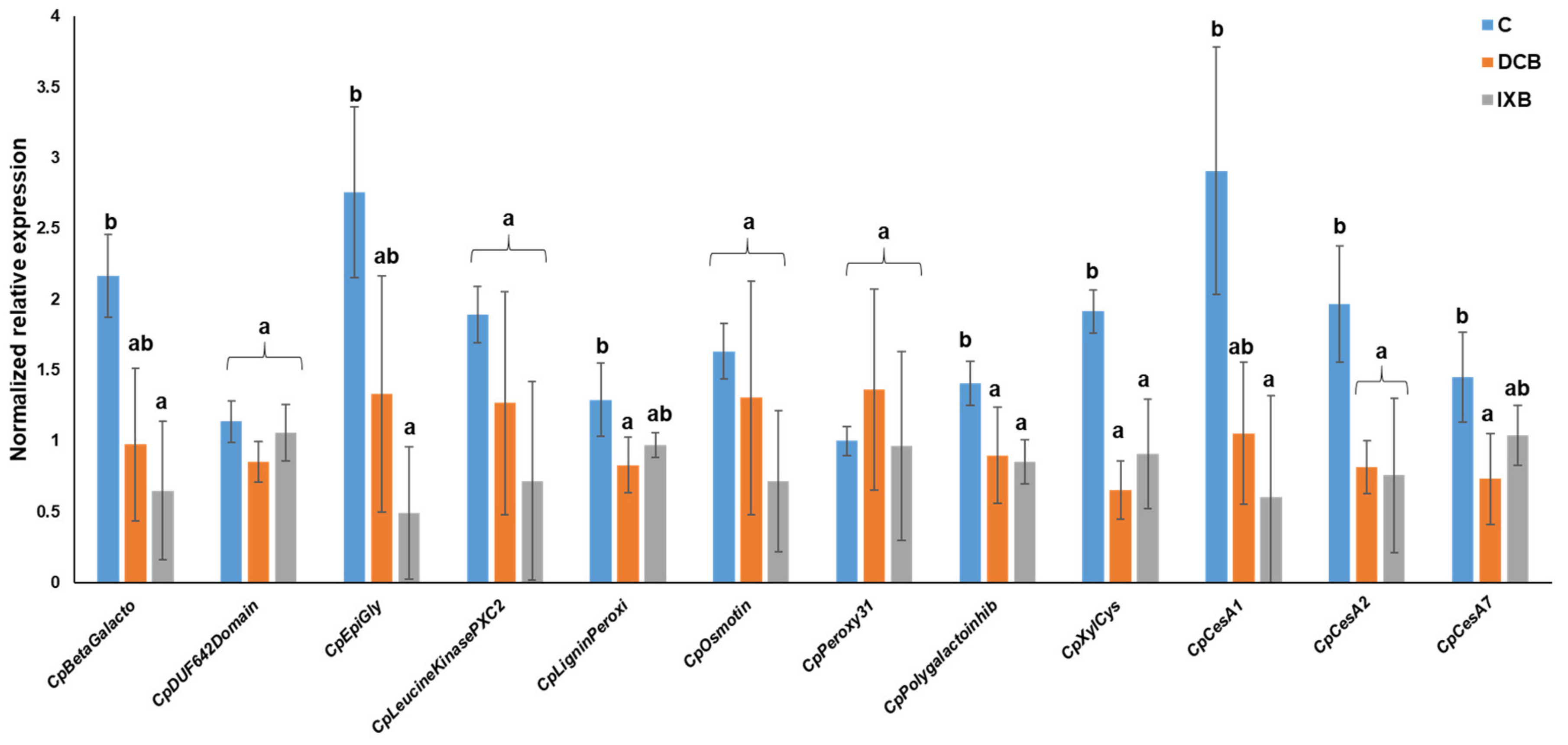{kind=link}
| Description | Contig Number | DCB/C FC | DCB/C log2 FC | Group |
|---|---|---|---|---|
| lignin-forming anionic peroxidase | Cp_V2_contig_18867 | 7656.2 | 12.9 | 2 |
| beta-glucosidase 40 | Cp_V2_contig_7909 | 896.0 | 9.8 | 1 |
| AF230277_1alpha-expansin 2 | Cp_V2_contig_38601 | 496.0 | 9.0 | 2 |
| probable beta-D-xylosidase 6 | Cp_V2_contig_42585 | 198.6 | 7.6 | 2 |
| ubiquitin-like protein | Cp_V2_contig_19299 | 74 | 6.2 | 2 |
| beta-galactosidase 3-like | Cp_V2_contig_34887 | 56.5 | 5.8 | 2 |
| beta-galactosidase 9 | Cp_V2_contig_9145 | 47.4 | 5.6 | 1 |
| F-box/kelch-repeat protein | Cp_V2_contig_38461 | 12.3 | 3.6 | 2 |
| apyrase 2-like | Cp_V2_contig_8189 | 12.1 | 3.6 | 1 |
| heparanase-like protein 2 | Cp_V2_contig_6154 | 11.4 | 3.5 | 2 |
| epidermis-specific secreted glycoprotein EP1-like | Cp_V2_contig_23179 | 11.2 | 3.4 | 2 |
| peroxidase 73-like | Cp_V2_contig_19463 | 10.9 | 3.4 | 1 |
| putative lipid-transfer protein DIR1 | Cp_V2_contig_45594 | 10.4 | 3.4 | 1 |
| beta-glucosidase 40-like | Cp_V2_contig_28315 | 10.4 | 3.4 | 1 |
| desiccation-related protein PCC13-62-like | Cp_V2_contig_37935 | 9.3 | 3.2 | 2 |
| cysteine-rich rehydration-responsive 1 | Cp_V2_contig_1534 | 8.2 | 3.0 | 1 |
| Description | Contig Number | IXB/C FC | IXB/C log2 FC | Group |
|---|---|---|---|---|
| lignin-forming anionic peroxidase | Cp_V2_contig_18867 | 7103.8 | 12.8 | 2 |
| AF230277_1alpha-expansin 2 | Cp_V2_contig_38600 | 908.6 | 9.8 | 2 |
| probable beta-D-xylosidase 6 | Cp_V2_contig_42585 | 333.8 | 8.4 | 2 |
| beta-galactosidase 3-like | Cp_V2_contig_34887 | 99.8 | 6.6 | 2 |
| ubiquitin-like protein | Cp_V2_contig_19299 | 73.3 | 6.2 | 2 |
| phylloplanin-like | Cp_V2_contig_30862 | 42.3 | 5.4 | 2 |
| beta-galactosidase 9 | Cp_V2_contig_9145 | 32.6 | 5.0 | 1 |
| beta-glucosidase 40 | Cp_V2_contig_7909 | 31.0 | 5.0 | 1 |
| cysteine-rich rehydration-responsive 1 | Cp_V2_contig_1534 | 24.2 | 4.6 | 2 |
| serine carboxypeptidase-like 42 | Cp_V2_contig_44837 | 24.2 | 4.6 | 2 |
| desiccation-related protein PCC13-62-like | Cp_V2_contig_37935 | 23.5 | 4.6 | 2 |
| non-specific lipid-transfer protein 2-like | Cp_V2_contig_21060 | 22.6 | 4.5 | 2 |
| epidermis-specific secreted glycoprotein EP1-like | Cp_V2_contig_2034 | 21.7 | 4.4 | 2 |
| Elongation factor 1-alpha | Cp_V2_contig_23179 | 17.7 | 4.1 | 2 |
| leucine-rich repeat extensin-like protein 2 | Cp_V2_contig_13652 | 16.8 | 4.1 | 2 |
| probable beta-D-xylosidase 7 | Cp_V2_contig_29850 | 15.2 | 3.9 | 2 |
| low-temperature-induced cysteine proteinase-like | Cp_V2_contig_1674 | 14.0 | 3.8 | 2 |
| heparanase-like protein 2 | Cp_V2_contig_6154 | 13.1 | 3.7 | 2 |
| probable purple acid phosphatase 20 | Cp_V2_contig_43085 | 12.3 | 3.6 | 2 |
| DNA-damage-repair/toleration protein DRT100 | Cp_V2_contig_25120 | 12.0 | 3.6 | 2 |
| probable inactive purple acid phosphatase 2 | Cp_V2_contig_44352 | 11.5 | 3.5 | 2 |
| leucine-rich repeat extensin-like protein 4 | Cp_V2_contig_8997 | 10.5 | 3.4 | 2 |
| epidermis-specific secreted glycoprotein EP1-like | Cp_V2_contig_2034 | 10.5 | 3.4 | 2 |
| beta-galactosidase 1 | Cp_V2_contig_10156 | 10.1 | 3.3 | 2 |
| putative lipid-transfer protein DIR1 | Cp_V2_contig_17417 | 9.8 | 3.3 | 2 |
| low-temperature-induced cysteine proteinase-like | Cp_V2_contig_6578 | 9.6 | 3.3 | 2 |
| subtilisin-like protease SBT1.9 | Cp_V2_contig_24584 | 9.3 | 3.2 | 2 |
| cationic peroxidase 1-like | Cp_V2_contig_13540 | 8.5 | 3.1 | 2 |
| F-box/kelch-repeat protein | Cp_V2_contig_38461 | 8.4 | 3.1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerriero, G.; Achen, C.; Xu, X.; Planchon, S.; Leclercq, C.C.; Sergeant, K.; Berni, R.; Hausman, J.-F.; Renaut, J.; Legay, S. The Cell Wall Proteome of Craterostigma plantagineum Cell Cultures Habituated to Dichlobenil and Isoxaben. Cells 2021, 10, 2295. https://doi.org/10.3390/cells10092295
Guerriero G, Achen C, Xu X, Planchon S, Leclercq CC, Sergeant K, Berni R, Hausman J-F, Renaut J, Legay S. The Cell Wall Proteome of Craterostigma plantagineum Cell Cultures Habituated to Dichlobenil and Isoxaben. Cells. 2021; 10(9):2295. https://doi.org/10.3390/cells10092295
Chicago/Turabian StyleGuerriero, Gea, Charles Achen, Xuan Xu, Sébastien Planchon, Céline C. Leclercq, Kjell Sergeant, Roberto Berni, Jean-Francois Hausman, Jenny Renaut, and Sylvain Legay. 2021. "The Cell Wall Proteome of Craterostigma plantagineum Cell Cultures Habituated to Dichlobenil and Isoxaben" Cells 10, no. 9: 2295. https://doi.org/10.3390/cells10092295
APA StyleGuerriero, G., Achen, C., Xu, X., Planchon, S., Leclercq, C. C., Sergeant, K., Berni, R., Hausman, J. -F., Renaut, J., & Legay, S. (2021). The Cell Wall Proteome of Craterostigma plantagineum Cell Cultures Habituated to Dichlobenil and Isoxaben. Cells, 10(9), 2295. https://doi.org/10.3390/cells10092295