
Selection on Sperm Count, but Not on Sperm Morphology or Velocity, in a Wild Population of Anolis Lizards

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Individuals and Sperm Samples
2.2. Measuring Sperm Traits
2.3. Genotyping and Parentage Assignment
2.4. Estimating Linear and Non-Linear Selection
3. Results
3.1. Parentage
3.2. Estimating Linear and Non-Linear Selection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersson, M.; Simmons, L.W. Sexual selection and mate choice. Trends Ecol. Evol. 2006, 21, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S.; Manier, M.K.; Puniamoorthy, N.; Schoff, C.; Starmer, W.T.; Luepold, S.H.B.; Belote, J.M.; Pitnick, S. How sexual selection can drive the evolution of costly sperm ornamentation. Nature 2016, 533, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Pitnick, S.; Hosken, D.J.; Birkhead, T.R. 2009 Sperm morphological diversity. In Sperm Biology: An Evolutionary Perspective; Academic Press: Cambridge, MA, USA, 2009; pp. 69–149. [Google Scholar]
- Eberhard, W. Female Control: Sexual Selection by Cryptic Female Choice; Princeton University Press: Princeton, NJ, USA, 1996. [Google Scholar]
- Lüpold, S.; Pitnick, S. Sperm form and function: What do we know about the role of sexual selection? Reproduction 2018, 155, R229–R243. [Google Scholar] [CrossRef]
- Parker, G. The reproductive behavior and the nature of sexual selection in Scatophaga stercoraria L.(Diptera: Scatophagidae). VII. The origin and evolution of the passive phase. Evolution 1970, 24, 774–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkhead, T.R.; Møller, A.P. Sperm Competition and Sexual Selection; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Birkhead, T.R.; Pizzari, T. Postcopulatory sexual selection. Nat. Rev. Genet. 2002, 3, 262–273. [Google Scholar] [CrossRef]
- Møller, A.P. Testes size, ejaculate quality and sperm competition in birds. Biol. J. Linn. Soc. 1988, 33, 273–283. [Google Scholar] [CrossRef]
- Harcourt, A.H.; Purvis, A.; Liles, L. Sperm competition: Mating system, not breeding season, affects testes size of primates. Funct. Ecol. 1995, 9, 468–476. [Google Scholar] [CrossRef]
- Stockley, P.; Gage, M.; Parker, G.A.; Møller, A.P. Sperm competition in fishes: The evolution of testis size and ejaculate characteristics. Am. Nat. 1997, 149, 933–954. [Google Scholar] [CrossRef] [Green Version]
- Byrne, P.G.; Roberts, J.D.; Simmons, L.W. Sperm competition selects for increased testes mass in Australian frogs. J. Evol. Biol. 2002, 15, 347–355. [Google Scholar] [CrossRef]
- Parker, G.A. The evolution of expenditure on testes. J. Zool. 2016, 298, 3–19. [Google Scholar] [CrossRef]
- Lüpold, S.; Linz, G.M.; Rivers, J.W.; Westneat, D.F.; Birkhead, T.R. Sperm competition selects beyond relative testes size in birds. Evolution 2009, 63, 391–402. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S.; Wistuba, J.; Damm, O.S.; Rivers, J.W.; Birkhead, T.R. Sperm competition leads to functional adaptations in avian testes to maximize sperm quantity and quality. Reproduction 2011, 141, 595–605. [Google Scholar] [CrossRef] [Green Version]
- Ramm, S.A.; Schärer, L. The evolutionary ecology of testicular function: Size isn’t everything. Biol. Rev. 2014, 89, 874–888. [Google Scholar] [CrossRef] [Green Version]
- Lüpold, S.; de Boer, R.A.; Evans, J.P.; Tomkins, J.L.; Fitzpatrick, J.L. How sperm competition shapes the evolution of testes and sperm: A meta-analysis. Philos. Trans. R. Soc. Lond. B 2020, 375, 20200064. [Google Scholar] [CrossRef]
- Simmons, L.W.; Fitzpatrick, J.L. Sperm wars and the evolution of male fertility. Reproduction 2012, 144, 519–534. [Google Scholar] [CrossRef]
- Fitzpatrick, J.L.; Lüpold, S. Sexual selection and the evolution of sperm quality. Mol. Hum. Reprod. 2014, 20, 1180–1189. [Google Scholar] [CrossRef] [Green Version]
- Parker, G.A.; Pizzari, T. Sperm competition and ejaculate economics. Biol. Rev. 2010, 85, 897–934. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A. Sperm competition games: Raffles and roles. Proc. Biol. Sci. Lond. B 1990, 242, 120–126. [Google Scholar]
- Parker, G.A. Marginal value theorem with exploitation time costs: Diet, sperm reserves, and optimal copula duration in dung flies. Am. Nat. 1992, 139, 1237–1256. [Google Scholar] [CrossRef]
- Ball, M.A.M.; Parker, G.A.G. Sperm competition games: Inter- and intra-species results of a continuous external fertilization model. J. Theor. Biol. 1997, 186, 8. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A.; Ball, M.A.; Stockley, P.; Gage, M.J. Sperm competition games: A prospective analysis of risk assessment. Proc. Biol. Sci. Lond. B 1997, 264, 1793–1802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, G.A.; Ball, M.A.; Stockley, P.; Gage, M. Sperm competition games: Individual assessment of sperm competition intensity by group spawners. Proc. Biol. Sci. Lond. B 1996, 263, 1291–1297. [Google Scholar]
- Parker, G.A.; Lessells, C.M.; Simmons, L.W. Sperm competition games: A general model for precopulatory male–male competition. Evolution 2013, 67, 95–109. [Google Scholar] [CrossRef]
- Parker, G.A.; Immler, S.; Pitnick, S.; Birkhead, T.R. Sperm competition games: Sperm size (mass) and number under raffle and displacement, and the evolution of P2. J. Theor. Biol. 2010, 264, 1003–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Immler, S.; Pitnick, S.; Parker, G.A.; Durrant, K.L.; Lüpold, S.; Calhim, S.; Birkhead, T.R. Resolving variation in the reproductive tradeoff between sperm size and number. Proc. Natl. Acad. Sci. USA 2011, 108, 5325–5330. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, J.L.; Simmons, L.W.; Evans, J.P. Complex patterns of multivariate selection on the ejaculate of a broadcast spawning invertebrate. Evolution 2012, 66, 2451–2460. [Google Scholar] [CrossRef]
- Johnson, D.W.; Monro, K.; Marshall, D.J. The maintenance of sperm variability: Context-dependent selection on sperm morphology in a broadcast spawning invertebrate. Evolution 2012, 67, 1383–1395. [Google Scholar] [CrossRef]
- Monro, K.; Marshall, D.J. Unravelling anisogamy: Egg size and ejaculate size mediate selection on morphology in free-swimming sperm. Proc. Biol. Sci. Lond. B 2016, 283, 20160671. [Google Scholar] [CrossRef]
- Lymbery, R.A.; Kennington, W.J.; Evans, J.P. Multivariate sexual selection on ejaculate traits under sperm competition. Am. Nat. 2018, 192, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Chirgwin, E.; Marshall, D.J.; Monro, K. Physical and physiological impacts of ocean warming alter phenotypic selection on sperm morphology. Funct. Ecol. 2020, 34, 646–657. [Google Scholar] [CrossRef]
- Hadlow, J.H.; Evans, J.P.; Lymbery, R.A. Egg-induced changes to sperm phenotypes shape patterns of multivariate selection on ejaculates. J. Evol. Biol. 2020, 33, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Brooks, R.; Hunt, J.; Blows, M.W.; Smith, M.J.; Bussière, L.F.; Jennions, M.D. Experimental evidence for multivariate stabilizing sexual selection. Evolution 2005, 59, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Roff, D.A.; Fairbairn, D.J. The evolution of trade-offs under directional and correlational selection. Evolution 2012, 66, 2461–2474. [Google Scholar] [CrossRef] [PubMed]
- Devigili, A.; Evans, J.P.; Di Nisio, A.; Pilastro, A. Multivariate selection drives concordant patterns of pre- and postcopulatory sexual selection in a livebearing fish. Nat. Commun. 2015, 6, 8291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, G.A.; Ball, M.A. Sperm competition, mating rate and the evolution of testis and ejaculate sizes: A population model. Biol. Lett. 2005, 1, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Vahed, K.; Parker, D.J. The evolution of large testes: Sperm competition or male mating rate? Ethology 2011, 118, 107–117. [Google Scholar] [CrossRef]
- Fromhage, L.; McNamara, J.M.; Houston, A.I. Sperm allocation strategies and female resistance: A unifying perspective. Am. Nat. 2008, 172, 25–33. [Google Scholar] [CrossRef]
- Preston, B.T.; Stevenson, I.R.; Pemberton, J.M.; Wilson, K. Dominant rams lose out by sperm depletion. Nature 2001, 409, 681–682. [Google Scholar] [CrossRef]
- Laskemoen, T.; Kleven, O.; Fossøy, F.; Robertson, R.J.; Rudolfsen, G.; Lifjeld, J.T. Sperm quantity and quality effects on fertilization success in a highly promiscuous passerine, the tree swallow Tachycineta bicolor. Behav. Ecol. Sociobiol. 2010, 64, 1473–1483. [Google Scholar] [CrossRef]
- Cramer, E.R.; Laskemoen, T.; Kleven, O.; LaBarbera, K.; Lovette, I.J.; Lifjeld, J.T. No evidence that sperm morphology predicts paternity success in wild house wrens. Behav. Ecol. Sociobiol. 2013, 67, 1845–1853. [Google Scholar] [CrossRef]
- Sætre, C.L.C.; Johnsen, A.; Stensrud, E.; Cramer, E.R. Sperm morphology, sperm motility and paternity success in the bluethroat (Luscinia svecica). PLoS ONE 2018, 13, e0192644. [Google Scholar] [CrossRef] [Green Version]
- Calsbeek, R.; Bonneaud, C. Postcopulatory fertilization bias as a form of cryptic sexual selection. Evolution 2008, 62, 1137–1148. [Google Scholar] [CrossRef]
- Kamath, A.; Losos, J.B. Estimating encounter rates as the first step of sexual selection in the lizard Anolis sagrei. Proc. Biol. Sci. Lond. B 2018, 285, 20172244. [Google Scholar] [CrossRef] [Green Version]
- Kahrl, A.F.; Cox, R.M. Diet affects ejaculate traits in a lizard with condition-dependent fertilization success. Behav. Ecol. 2015, 26, 1502–1511. [Google Scholar] [CrossRef] [Green Version]
- Cox, R.; Calsbeek, R. Cryptic sex-ratio bias provides indirect genetic benefits despite sexual conflict. Science 2010, 328, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Schoener, T.W.; Schoener, A. Densities, sex ratios, and population structure in four species of Bahamian Anolis lizards. J. Anim. Ecol. 1980, 19–53. [Google Scholar] [CrossRef]
- Tokarz, R.R.; Paterson, A.V.; McMann, S. Importance of dewlap display in male mating success in free-ranging brown anoles (Anolis sagrei). J. Herpetol. 2005, 39, 174–177. [Google Scholar] [CrossRef]
- Simon, V.B. Communication signal rates predict interaction outcome in the brown anole lizard, Anolis sagrei. Copeia 2011, 2011, 38–45. [Google Scholar] [CrossRef]
- Kahrl, A.F.; Johnson, M.A.; Cox, R.M. Rapid evolution of testis size relative to sperm morphology suggests that post-copulatory selection targets sperm number in Anolis lizards. J. Evol. Biol. 2019, 32, 302–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahrl, A.F.; Cox, R.M. Consistent differences in sperm morphology and testis size between native and introduced populations of three Anolis lizard species. J. Herpetol. 2017, 51, 532–537. [Google Scholar] [CrossRef]
- Cramer, E.R.; Rowe, M.; Eroukhmanoff, F.; Lifjeld, J.T.; Sætre, G.-P.; Johnsen, A. Measuring sperm swimming performance in birds: Effects of dilution, suspension medium, mechanical agitation, and sperm number. J. Ornithol. 2019, 160, 1053–1063. [Google Scholar] [CrossRef]
- Campbell, N.R.; Harmon, S.A.; Narum, S.R. Genotyping-in-Thousands by sequencing (GT-seq): A cost effective SNP genotyping method based on custom amplicon sequencing. Mol. Ecol. Resour. 2015, 15, 855–867. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: https://f1000research.com/articles/8-1874/v2 (accessed on 5 March 2018).
- Anderson, E.C. Large-scale parentage inference with SNPs: An efficient algorithm for statistical confidence of parent pair allocations. Stat. Appl. Genet. Mol. Biol. 2012, 11, 1–26. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org/ (accessed on 18 May 2021).
- Lande, R.; Arnold, S.J. The measurement of selection on correlated characters. Evolution 1983, 37, 1210–1226. [Google Scholar] [CrossRef] [PubMed]
- Stinchcombe, J.R.; Agrawal, A.F.; Hohenlohe, P.A.; Arnold, S.J.; Blows, M.W. Estimating nonlinear selection gradients using quadratic regression coefficients: Double or nothing? Evolution 2008, 62, 2435–2440. [Google Scholar] [CrossRef]
- Brodie, E.D.; Moore, A.J.; Janzen, F.J. Visualizing and quantifying natural selection. Trends Ecol. Evol. 1995, 10, 313–318. [Google Scholar] [CrossRef]
- Schluter, D.; Nychka, D. Exploring fitness surfaces. Am. Nat. 1994, 143, 597–616. [Google Scholar] [CrossRef]
- Wood, S.; Wood, M.S. Package “mgcv”. R Package Version 2015, 1, 29. [Google Scholar]
- Cox, R.M.; Calsbeek, R. Sex-specific selection and intraspecific variation in sexual size dimorphism. Evolution 2010, 64, 798–809. [Google Scholar] [CrossRef]
- Simmons, L.W. Sperm Competition and Its Evolutionary Consequences in the Insects; Princeton University Press: Princeton, NJ, USA, 2001. [Google Scholar]
- Kahrl, A.F.; Cox, C.L.; Cox, R.M. Correlated evolution between targets of pre- and postcopulatory sexual selection across squamate reptiles. Ecol. Evol. 2016, 6, 6452–6459. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.A.; Begon, M.E. Sperm competition games: Sperm size and number under gametic control. Proc. Biol. Sci. Lond. B 1993, 253, 255–262. [Google Scholar]
- Birkhead, T.R.; Fletcher, F. Depletion determines sperm numbers in male zebra finches. Anim. Behav. 1995, 49, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Rubolini, D.; Galeotti, P.; PUPIN, F.; Sacchi, R.; NARDI, P.A.; Fasola, M. Repeated matings and sperm depletion in the freshwater crayfish Austropotamobius italicus. Freshw. Biol. 2007, 52, 1898–1906. [Google Scholar] [CrossRef]
- Dewsbury, D. Ejaculate cost and male choice. Am. Nat. 1982, 119, 601–610. [Google Scholar] [CrossRef]
- Wedell, N.; Cook, P.A. Butterflies tailor their ejaculate in response to sperm competition risk and intensity. Proc. R. Soc. Lond. B 1999, 266, 1033–1039. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.W.; Kvarnemo, C. Ejaculate expenditure by male bush crickets decreases with sperm competition intensity. Proc. Biol. Sci. Lond. B 1997, 264, 1203–1208. [Google Scholar] [CrossRef] [Green Version]
- Kelly, C.D.; Jennions, M.D. Sexual selection and sperm quantity: Meta-analyses of strategic ejaculation. Biol. Rev. 2011, 86, 863–884. [Google Scholar] [CrossRef] [PubMed]
- Kustra, M.C.; Kahrl, A.F.; Reedy, A.M.; Warner, D.A.; Cox, R.M. Sperm morphology and count vary with fine-scale changes in local density in a wild lizard population. Oecologia 2019, 191, 555–564. [Google Scholar] [CrossRef] [Green Version]
- Wedell, N.; Gage, M.J.G.; Parker, G.A. Sperm competition, male prudence and sperm-limited females. Trends Ecol. Evol. 2002, 17, 313–320. [Google Scholar] [CrossRef]
- Duryea, M.C.; Bergeron, P.; Clare Salzler, Z.; Calsbeek, R. Field estimates of parentage reveal sexually antagonistic selection on body size in a population of Anolis lizards. Ecol. Evol. 2016, 6, 7024–7031. [Google Scholar] [CrossRef]
- Shuster, S.M.; Wade, M.J. Mating Systems and Strategies; Princeton University Press: Princeton, NJ, USA, 2003. [Google Scholar]
- Alonzo, S.H.; Warner, R.R. Allocation to mate guarding or increased sperm production in a Mediterranean wrasse. Am. Nat. 2000, 156, 266–275. [Google Scholar] [CrossRef]
- Candolin, U.; Reynolds, J.D. Adjustments of ejaculation rates in response to risk of sperm competition in a fish, the bitterling (Rhodeus sericeus). Proc. Biol. Sci. Lond. B 2002, 269, 1549–1553. [Google Scholar] [CrossRef] [Green Version]
- Neff, B.D.; Pitcher, T.E. Genetic quality and sexual selection: An integrated framework for good genes and compatible genes. Mol. Ecol. 2004, 14, 19–38. [Google Scholar] [CrossRef] [PubMed]
- Lailvaux, S.P.; Irschick, D.J. The evolution of performance-based male fighting ability in Caribbean Anolis lizards. Am. Nat. 2007, 170, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Reedy, A.M.; Pope, B.D.; Kiriazis, N.M.; Giordano, C.L.; Sams, C.L.; Warner, D.A.; Cox, R.M. Female anoles display less but attack more quickly than males in response to territorial intrusions. Behav. Ecol. 2017, 28, 1323–1328. [Google Scholar] [CrossRef]
- Kleven, O.; Fossøy, F.; Laskemoen, T.; Robertson, R.J.; Rudolfsen, G.; Lifjeld, J.T. Comparative evidence for the evolution of sperm swimming speed by sperm competition and female sperm storage duration in passerine birds. Evolution 2009, 63, 2466–2473. [Google Scholar] [CrossRef]
- Bennison, C.; Hemmings, N.; Slate, J.; Birkhead, T. Long sperm fertilize more eggs in a bird. Proc. Biol. Sci. Lond. B 2014, 282, 20141897. [Google Scholar] [CrossRef]
- Bakker, T.C.M.; Hollmann, M.; Mehlis, M.; Zbinden, M. Functional variation of sperm morphology in sticklebacks. Behav. Ecol. Sociobiol. 2014, 68, 617–627. [Google Scholar] [CrossRef]
- Friesen, C.R.; Kahrl, A.F.; Olsson, M. Sperm competition in squamate reptiles. Philos. Trans. R. Soc. Lond. B 2020, 375, 20200079. [Google Scholar] [CrossRef]
- Lüpold, S.; Calhim, S.; Immler, S.; Birkhead, T.R. Sperm morphology and sperm velocity in passerine birds. Proc. R. Soc. Lond. B 2009, 276, 1175–1181. [Google Scholar] [CrossRef] [Green Version]
- Firman, R.C.; Simmons, L.W. Sperm midpiece length predicts sperm swimming velocity in house mice. Biol. Lett. 2010, 6, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Crowe, S.A.; Kleven, O.; Delmore, K.E.; Laskemoen, T.; Nocera, J.J.; Lifjeld, J.T.; Robertson, R.J. Paternity assurance through frequent copulations in a wild passerine with intense sperm competition. Anim. Behav. 2009, 77, 183–187. [Google Scholar] [CrossRef]
- Carleial, R.; McDonald, G.; Spurgin, L.; Fairfield, E.; Wang, Y.; Richardson, D.; Pizzari, T. Fine-scale dynamics of competitive fertilisation in social groups of red junglefowl (Gallus gallus) shed new light on avian sperm competition. Philos. Trans. R. Soc. Lond. B 2020, 375, 20200081. [Google Scholar] [CrossRef] [PubMed]


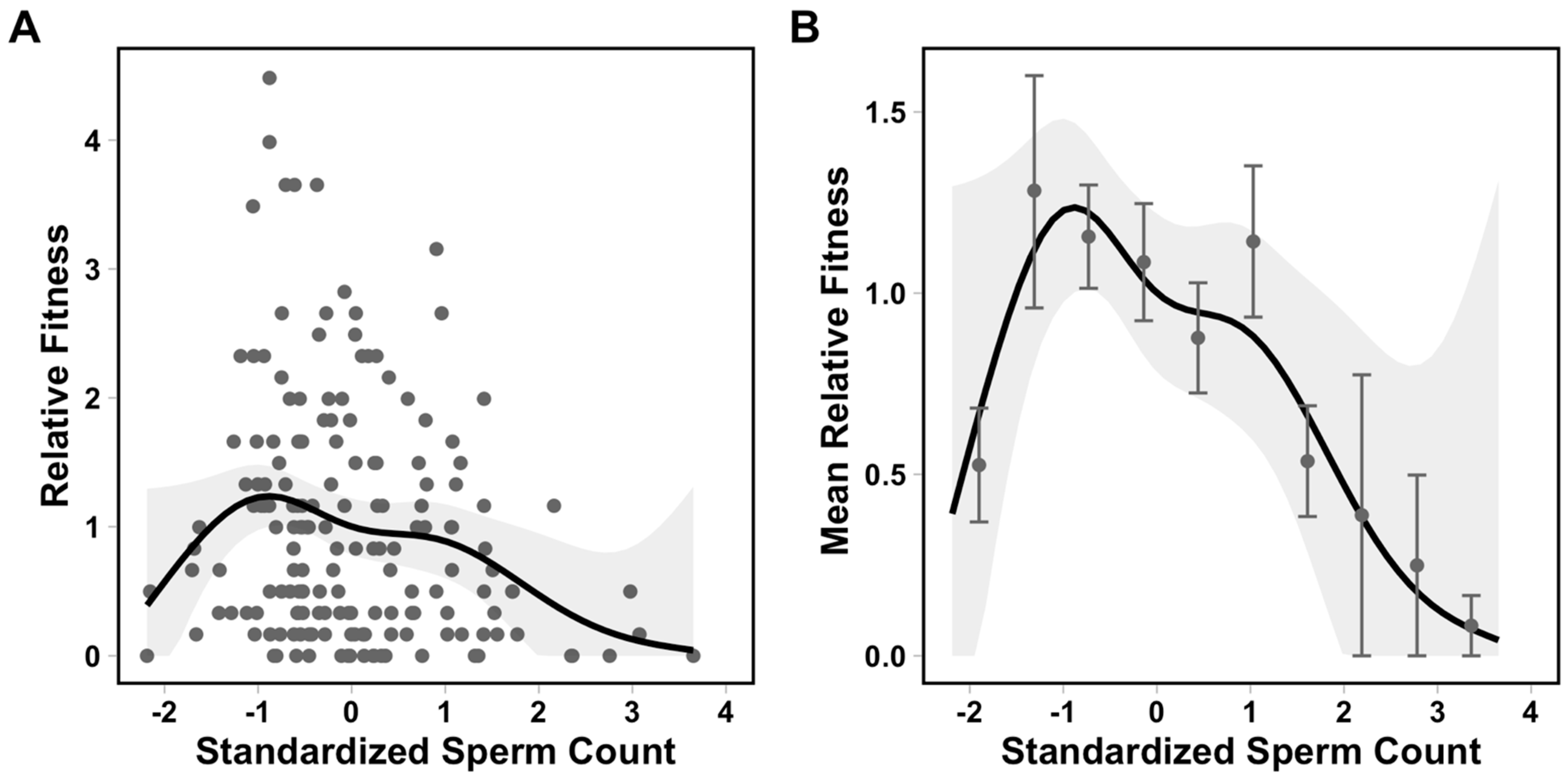{kind=link}
| s | β | c | ||||
| Traits | Linear (Univariate) | Linear (Multivariate) | Non-Linear (Univariate) | |||
| Sperm count | −0.179 (±0.07) | −0.167 (±0.07) | −0.219 (±0.08) | |||
| Velocity (VAPi) | −0.021 (±0.07) | 0.033 (±0.09) | −0.054 (±0.08) | |||
| Linearity | −0.005 (±0.07) | 0.009 (±0.09) | −0.012 (±0.08) | |||
| Head length | −0.106 (±0.07) | −0.072 (±0.08) | −0.032 (±0.11) | |||
| Midpiece length | 0.106 (±0.07) | 0.038 (±0.08) | −0.088 (±0.09) | |||
| Flagellum length | −0.069 (±0.07) | −0.047 (±0.07) | 0.024 (±0.09) | |||
| γ | ||||||
| Traits | Sperm Count (Multivariate) | Velocity (VAPi) (Multivariate) | Linearity (Multivariate) | Head Length (Multivariate) | Midpiece Length (Multivariate) | Flagellum Length (Multivariate) |
| Sperm count | −0.162 (±0.10) | −0.027 (±0.13) | −0.084 (±0.11) | −0.008 (±0.07) | −0.037 (±0.08) | 0.060 (±0.09) |
| Velocity (VAPi) | 0.078 (±0.20) | −0.080 (±0.17) | −0.040 (±0.11) | 0.188 (±0.13) | 0.091 (±0.11) | |
| Linearity | −0.034 (±0.17) | 0.151 (±0.11) | −0.181 (±0.13) | 0.037 (±0.11) | ||
| Head length | −0.058 (±0.12) | −0.040 (±0.08) | 0.053 (±0.09) | |||
| Midpiece length | −0.168 (±0.12) | −0.055 (±0.09) | ||||
| Flagellum length | −0.131 (±0.11) | |||||
| s | β | c | γ | ||
|---|---|---|---|---|---|
| Traits | Linear (Univariate) | Linear (Multivariate) | Non-Linear (Univariate) | Sperm Count (Multivariate) | SVL (Multivariate) |
| Sperm count | −0.179 (±0.07) | −0.168 (±0.07) | −0.220 (±0.08) | −0.225 (±0.08) | −0.022 (±0.07) |
| SVL | 0.089 (±0.07) | 0.056 (±0.07) | −0.048 (±0.08) | −0.040 (±0.08) | |
| Trait | VIF | Velocity (VAPi) | Linearity | Head Length | Midpiece Length | Flagellum Length |
|---|---|---|---|---|---|---|
| Count | 1.170 | 0.241 (0.001) | 0.041 (0.593) | 0.120 (0.117) | −0.249 (0.001) | 0.039 (0.609) |
| Velocity (VAPi) | 1.791 | 0.638 (0.001) | 0.123 (0.107) | −0.130 (0.088) | 0.135 (0.076) | |
| Linearity | 1.690 | 0.121 (0.113) | −0.118 (0.122) | 0.101 (0.185) | ||
| Head length | 1.136 | −0.287 (0.001) | 0.174 (0.023) | |||
| Midpiece length | 1.223 | −0.231 (0.003) | ||||
| Flagellum length | 1.104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kahrl, A.F.; Kustra, M.C.; Reedy, A.M.; Bhave, R.S.; Seears, H.A.; Warner, D.A.; Cox, R.M. Selection on Sperm Count, but Not on Sperm Morphology or Velocity, in a Wild Population of Anolis Lizards. Cells 2021, 10, 2369. https://doi.org/10.3390/cells10092369
Kahrl AF, Kustra MC, Reedy AM, Bhave RS, Seears HA, Warner DA, Cox RM. Selection on Sperm Count, but Not on Sperm Morphology or Velocity, in a Wild Population of Anolis Lizards. Cells. 2021; 10(9):2369. https://doi.org/10.3390/cells10092369
Chicago/Turabian StyleKahrl, Ariel F., Matthew C. Kustra, Aaron M. Reedy, Rachana S. Bhave, Heidi A. Seears, Daniel A. Warner, and Robert M. Cox. 2021. "Selection on Sperm Count, but Not on Sperm Morphology or Velocity, in a Wild Population of Anolis Lizards" Cells 10, no. 9: 2369. https://doi.org/10.3390/cells10092369
APA StyleKahrl, A. F., Kustra, M. C., Reedy, A. M., Bhave, R. S., Seears, H. A., Warner, D. A., & Cox, R. M. (2021). Selection on Sperm Count, but Not on Sperm Morphology or Velocity, in a Wild Population of Anolis Lizards. Cells, 10(9), 2369. https://doi.org/10.3390/cells10092369