
Parthenogenesis and the Evolution of Anisogamy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Model
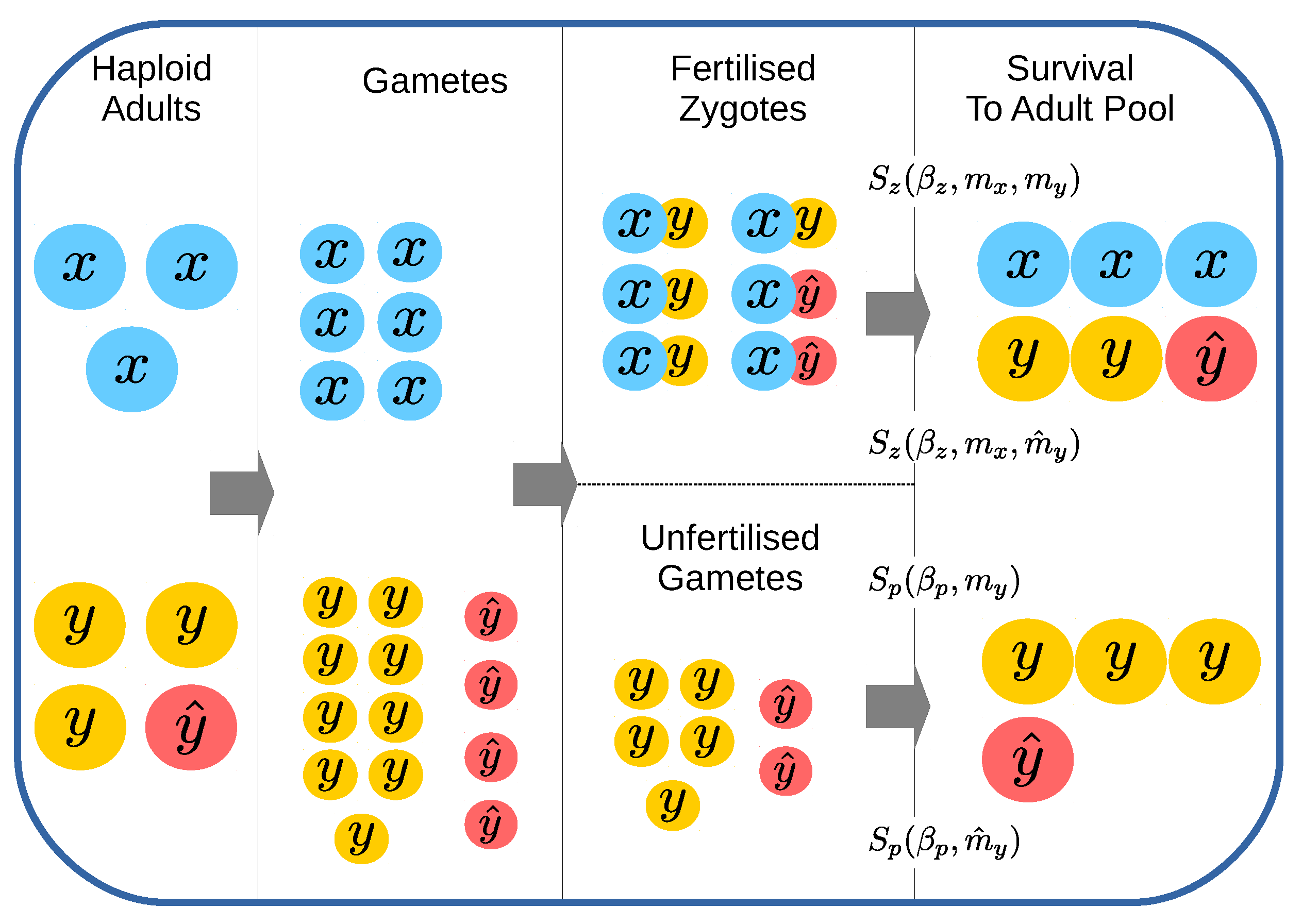
2.1. Dynamics within Each Generation
2.1.1. Gamete Formation/Gametogenesis
2.1.2. Zygote Formation
2.1.3. Zygote and Unfertilized Gamete Survival
2.1.4. Forming the Next Adult Generation
2.2. Invasion Dynamics
2.2.1. Evolution of Gamete Sizes
2.2.2. Mutations to Obligate Asexuality
2.3. Evolutionary Dynamics
2.3.1. Mutations to a Different Gamete Size
2.3.2. Mutations to Asexuality
3. Results
3.1. Examples of Evolutionary Strategies
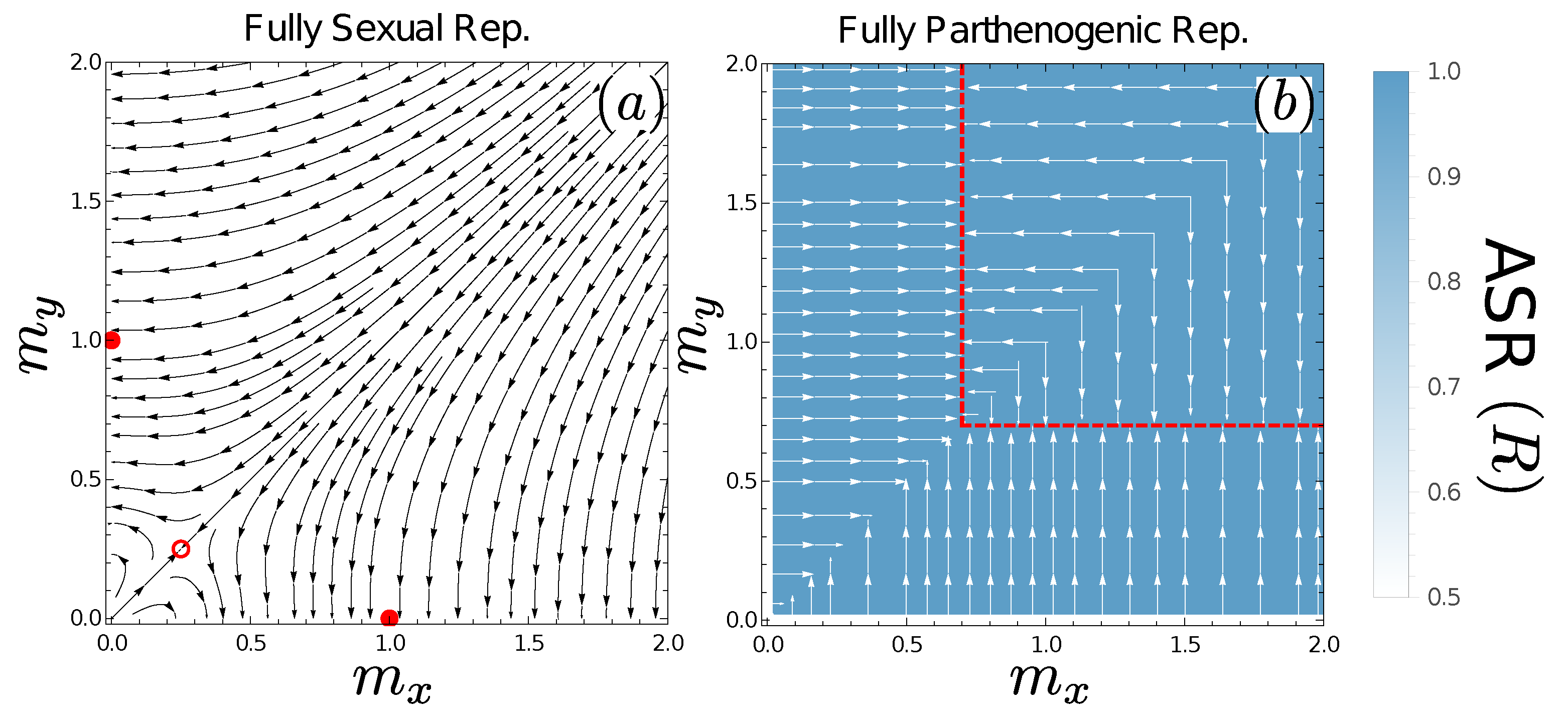
3.2. Evolutionary Dynamics under Obligately Sexual Reproduction
3.3. Evolutionary Dynamics of Progeny Size under Asexuality
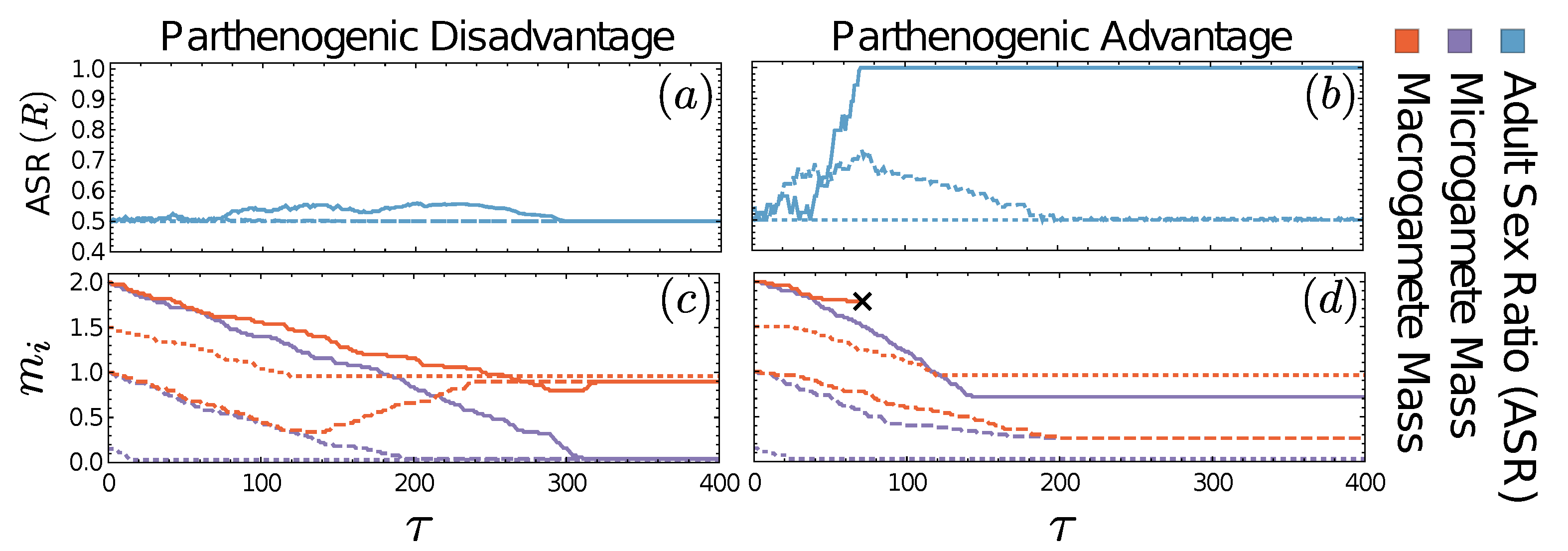
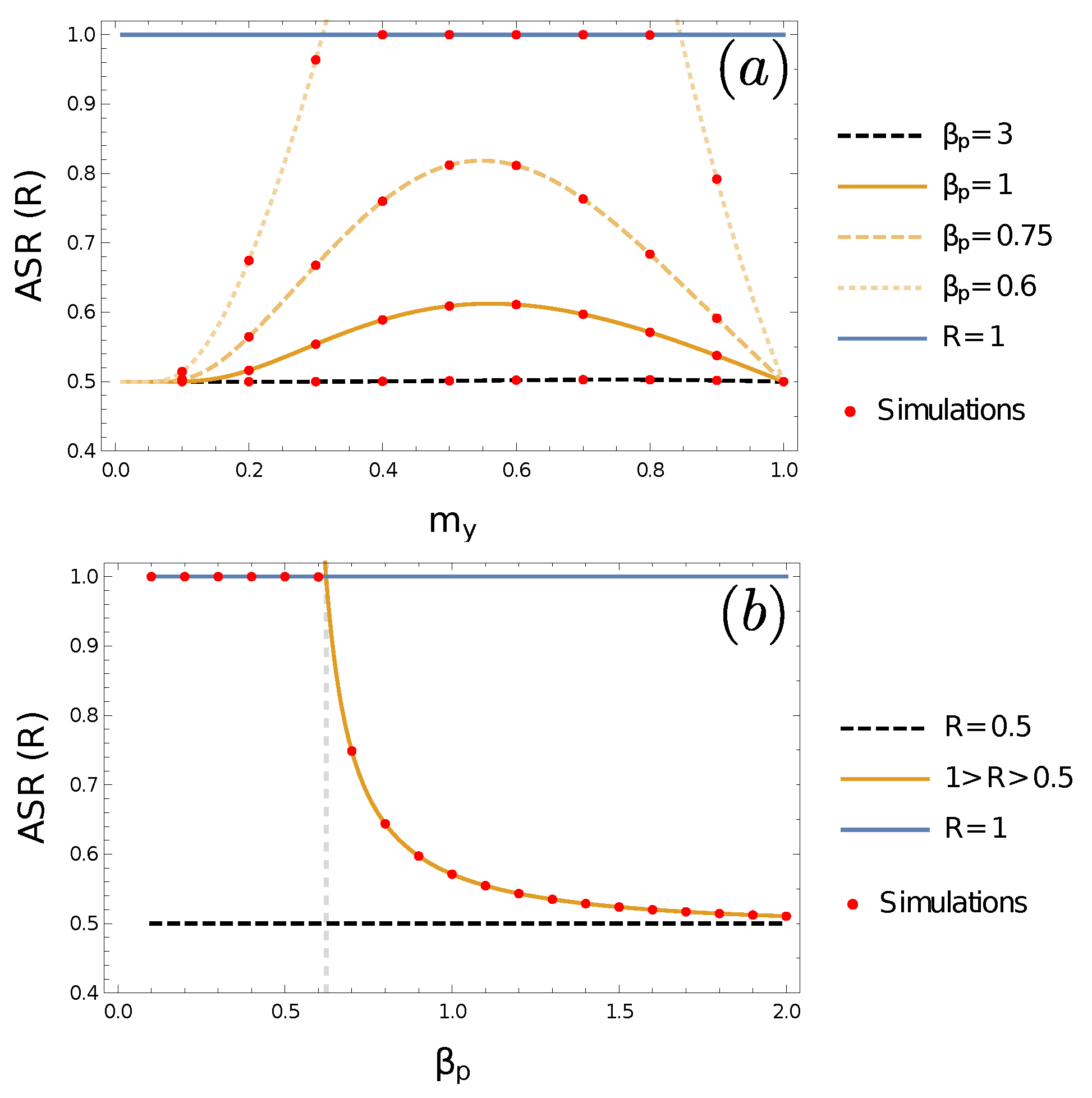
3.4. Adult Sex Ratio
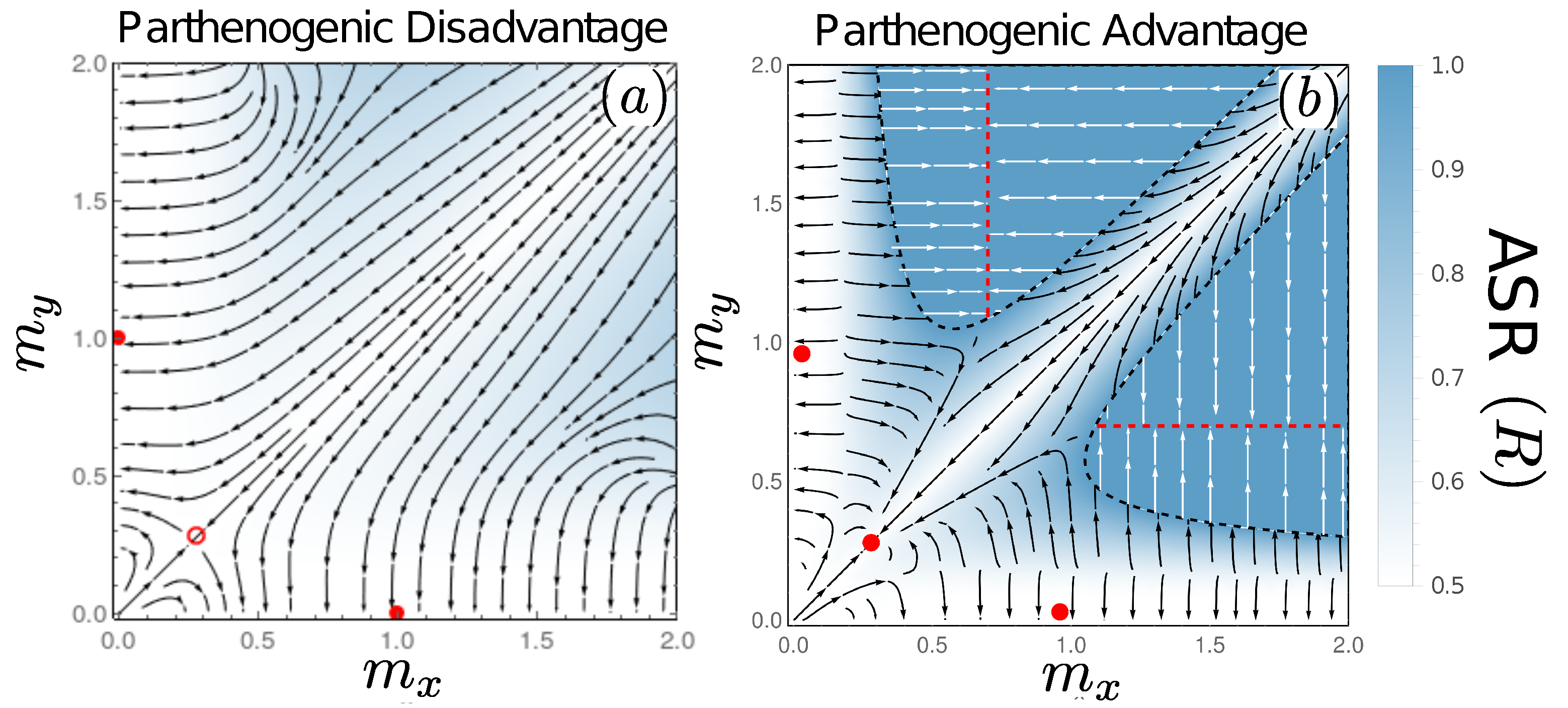
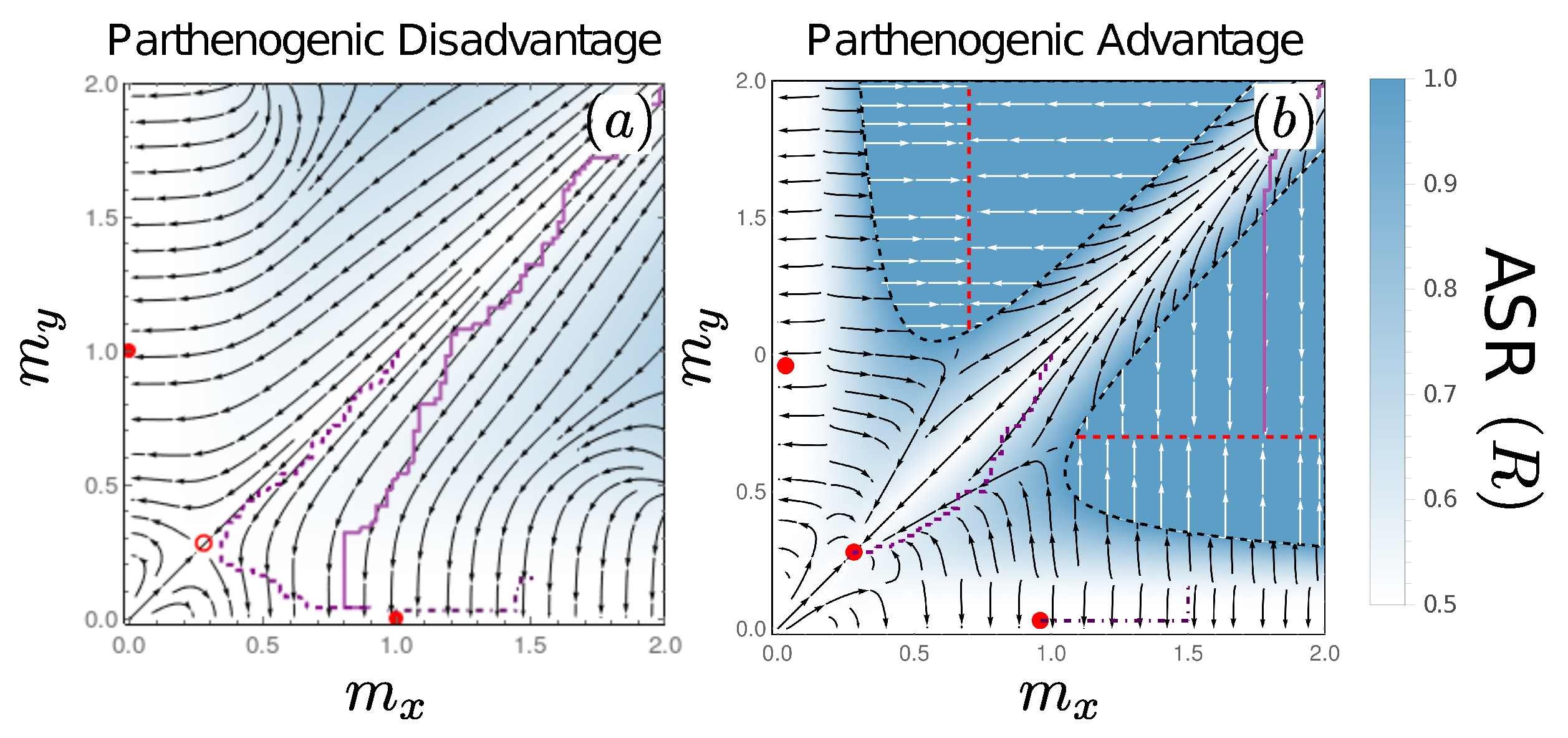
3.5. Evolutionary Dynamics: Mixed Sexual and Parthenogenic Reproduction
3.5.1. De Facto Asexuality due to Extinction of Macrogametes
3.5.2. A Novel Route to Stable Isogamy
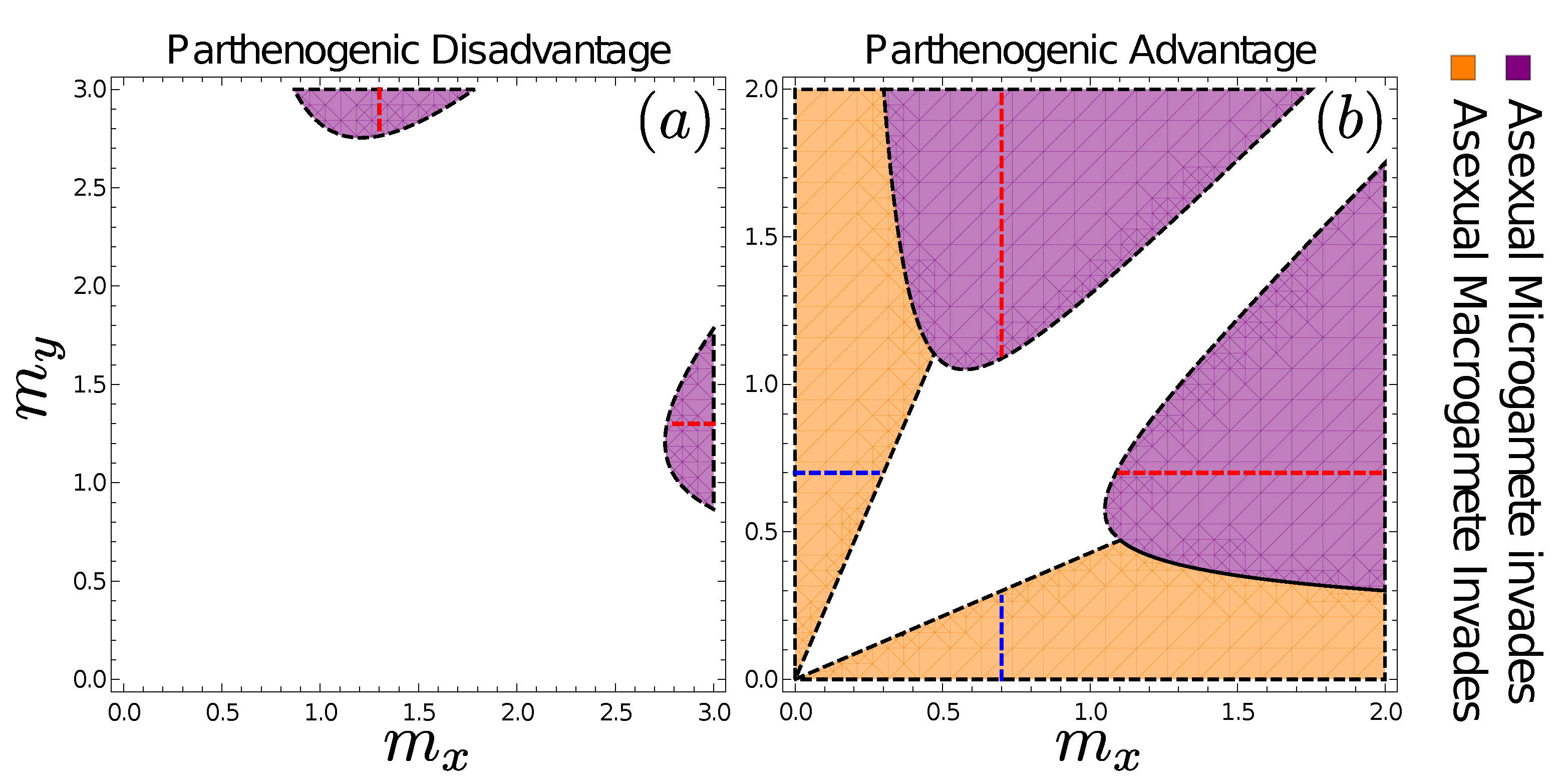
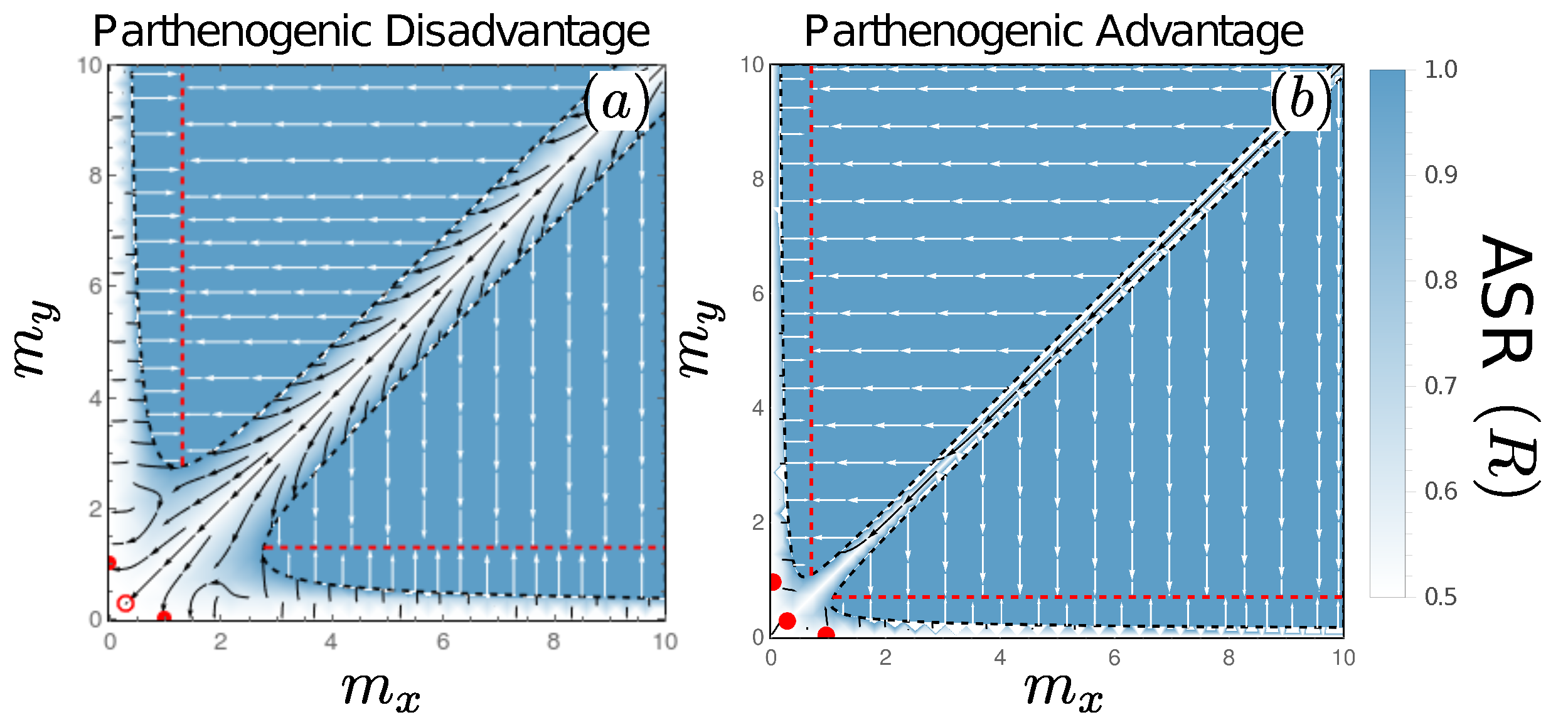
3.6. Fates of Mutations to Asexuality Depend on the Mating Type They Arise in
4. Discussion
4.1. Relation to “Evolution of Anisogamy in Organisms with Parthenogenetic Gametes”
4.2. Two Routes to Asexuality
4.3. Relation to Empirical Systems
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A. Fertilisation Kinetics: Mutant Gamete Size
Appendix B. Fertilisation Kinetics: Mutant Asexual
Appendix C. Derivation of Invasion Dynamics of a Different Sized Gamete
Appendix D. Derivation of Adult Sex Ratio
Appendix E. Derivation of Evolutionary Dynamics of Gamete Size
Appendix F. Derivation of Asexual Invasion Conditions
Appendix G. Derivation of Evolutionary Dynamics of Parthenosporophyte Size in Absence of Syngamy
Appendix H. Figures



References
- Parker, G.A.; Baker, R.; Smith, V.G. The origin and evolution of gamete dimorphism and the male-female phenomenon. J. Theor. Biol. 1972, 36, 529–553. [Google Scholar] [CrossRef]
- Bulmer, M.G.; Parker, G.A. The evolution of anisogamy: A game-theoretic approach. Proc. R. Soc. Lond. B. 2002, 269, 2381–2388. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.; Drysdale, V.L. Isogamy in large and complex volvocine algae is consistent with the gamete competition theory of the evolution of anisogamy. Proc. R. Soc. B 2018, 285, 20181954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanschen, E.R.; Herron, M.D.; Wiens, J.J.; Nozaki, H.; Michod, R.E. Multicellularity drives the evolution of sexual traits. Am. Nat. 2018, 192, E93–E105. [Google Scholar] [CrossRef]
- Evans, J.P.; Lymbery, R.A. Sexual selection after gamete release in broadcast spawning invertebrates. Philos. Trans. R. Soc. B 2020, 375, 20200069. [Google Scholar] [CrossRef] [PubMed]
- Lessells, C.M.; Snook, R.R.; Hosken, D.J. The evolutionary origin and maintenance of sperm: Selection for a small, motile gamete mating type. In Sperm biology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 43–67. [Google Scholar]
- Lehtonen, J.; Kokko, H. Two roads to two sexes: Unifying gamete competition and gamete limitation in a single model of anisogamy evolution. Behav. Ecol. Sociobiol. 2011, 65, 445–459. [Google Scholar] [CrossRef]
- Lehtonen, J.; Parker, G.A. Evolution of the two sexes under internal fertilization and alternative evolutionary pathways. Am. Nat. 2019, 193, 702–716. [Google Scholar] [CrossRef]
- Einum, S.; Fleming, I.A. Of chickens and eggs: Diverging propagule size of iteroparous and semelparous organisms. Evolution 2007, 61, 232–238. [Google Scholar] [CrossRef]
- Falster, D.S.; Moles, A.T.; Westoby, M. A general model for the scaling of offspring size and adult size. Am. Nat. 2008, 172, 299–317. [Google Scholar] [CrossRef] [Green Version]
- Bueno, J.; Lopez-Urrutia, A. The offspring-development-time/offspring-number trade-off. Am. Nat. 2012, 179, E196–E203. [Google Scholar] [CrossRef]
- Levins, R.; MacArthur, R. The maintenance of genetic polymorphism in a spatially heterogeneous environment: Variations on a theme by Howard Levene. Am. Nat. 1966, 100, 585–589. [Google Scholar] [CrossRef]
- Williams, G.C. A defense of reductionism in evolutionary biology. Oxf. Surv. Evol. Biol. 1985, 2, 1–27. [Google Scholar]
- Kokko, H. Modelling for Field Biologists and other Interesting People; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Marquet, P.A.; Allen, A.P.; Brown, J.H.; Dunne, J.A.; Enquist, B.J.; Gillooly, J.F.; Gowaty, P.A.; Green, J.L.; Harte, J.; Hubbell, S.P.; et al. On theory in ecology. BioScience 2014, 64, 701–710. [Google Scholar] [CrossRef] [Green Version]
- Smaldino, P.E. Models are stupid, and we need more of them. In Computational Social Psychology; Routledge: New York, NY, USA, 2017; pp. 311–331. [Google Scholar]
- Billiard, S.; López-Villavicencio, M.; Hood, M.; Giraud, T. Sex, outcrossing and mating types: Unsolved questions in fungi and beyond. J. Evol. Biol. 2012, 25, 1020–1038. [Google Scholar] [CrossRef] [PubMed]
- Kokko, H. Give one species the task to come up with a theory that spans them all: What good can come out of that? Proc. R. Soc. B: Biol. Sci. 2017, 284, 20171652. [Google Scholar] [CrossRef] [Green Version]
- Kokko, H. When synchrony makes the best of both worlds even better: How well do we really understand facultative sex? Am. Nat. 2020, 195, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dacks, J.; Roger, A.J. The first sexual lineage and the relevance of facultative sex. J. Mol. Evol. 1999, 48, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Schön, I.; Martens, K.; van Dijk, P. Lost sex. In The Evolutionary Biology of Parthenogenesis; Springer: Dordrecht, The Netherlands, 2009; pp. 1–615. [Google Scholar]
- Constable, G.W.A.; Kokko, H. The rate of facultative sex governs the number of expected mating types in isogamous species. Nat. Ecol. Evol. 2018, 2, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Czuppon, P.; Constable, G.W.A. Invasion and extinction dynamics of mating types under facultative sexual reproduction. Genetics 2019, 213, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Berríos-Caro, E.; Galla, T.; Constable, G.W.A. Switching environments, synchronous sex, and the evolution of mating types. Theor. Popul. Biol. 2021, 138, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Lehtonen, J.; Horinouchi, Y.; Togashi, T.; Parker, G.A. Evolution of anisogamy in organisms with parthenogenetic gametes. Am. Nat. 2021, in press. [Google Scholar] [CrossRef]
- Coelho, S.M.; Scornet, D.; Rousvoal, S.; Peters, N.T.; Dartevelle, L.; Peters, A.F.; Cock, J.M. Ectocarpus: A model organism for the brown algae. Cold Spring Harb. Protoc. 2012, 2012, pdb–emo065821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umen, J.; Coelho, S. Algal sex determination and the evolution of anisogamy. Annu. Rev. Microbiol. 2019, 73, 267–291. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Graf, L.; Peters, A.F.; Cock, J.M.; Nishitsuji, K.; Arimoto, A.; Shoguchi, E.; Nagasato, C.; Choi, C.G.; Yoon, H.S. Organelle inheritance and genome architecture variation in isogamous brown algae. Sci. Rep. 2020, 10, 2048. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, N.; Nagasato, C.; Motomura, T. Chemotactic movement in sperm of the oogamous brown algae, Saccharina japonica and Fucus distichus. Protoplasma 2017, 254, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Coelho, S.M.; Peters, A.F.; Müller, D.; Cock, J.M. Ectocarpus: An evo-devo model for the brown algae. EvoDevo 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Mignerot, L.; Avia, K.; Luthringer, R.; Lipinska, A.P.; Peters, A.F.; Cock, J.M.; Coelho, S.M. A key role for sex chromosomes in the regulation of parthenogenesis in the brown alga Ectocarpus. PLoS Genet. 2019, 15, e1008211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luthringer, R.; Cormier, A.; Ahmed, S.; Peters, A.; Cock, J.; Coelho, S. Sexual dimorphism in the brown algae. Perspect. Phycol 2014, 1, 11–25. [Google Scholar] [CrossRef]
- Heesch, S.; Serrano-Serrano, M.; Luthringer, R.; Peters, A.F.; Destombe, C.; Cock, J.M.; Valero, M.; Roze, D.; Salamin, N.; Coelho, S. Evolution of life cycles and reproductive traits: Insights from the brown algae. J. Evol. Biol. 2021, 34, 992–1009. [Google Scholar] [CrossRef]
- Phillips, N.; Burrowes, R.; Rousseau, F.; De Reviers, B.; Saunders, G.W. Resolving evolutionary relationships among the brown algae using chloroplast and nuclear genes 1. J. Phycol. 2008, 44, 394–405. [Google Scholar] [CrossRef]
- Müller, D.G.; Gaschet, E.; Godfroy, O.; Gueno, J.; Cossard, G.; Kunert, M.; Peters, A.F.; Westermeier, R.; Boland, W.; Cock, J.M.; et al. A partially sex-reversed giant kelp sheds light into the mechanisms of sexual differentiation in a UV sexual system. New Phytol. 2021, 232, 252–263. [Google Scholar] [CrossRef]
- Coelho, S.M.; Mignerot, L.; Cock, J.M. Origin and evolution of sex-determination systems in the brown algae. New Phytol. 2019, 222, 1751–1756. [Google Scholar] [CrossRef]
- Lehtonen, J. Models of fertilization kinetics. R. Soc. Open Sci. 2015, 2, 150175. [Google Scholar] [CrossRef]
- Vance, R.R. On reproductive strategies in marine bottom invertebrates. Am. Nat. 1973, 107, 339–352. [Google Scholar] [CrossRef]
- Levitan, D.R. The importance of sperm limitation to the evolution of egg size in marine invertebrates. Am. Nat. 1993, 141, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Cock, J.; Pessia, E.; Luthringer, R.; Cormier, A.; Robuchon, M.; Sterck, L.; Peters, A.; Dittami, S.; Corre, E.; et al. A Haploid System of Sex Determination in the Brown Alga Ectocarpus sp. Curr. Biol. 2014, 24, 1945–1957. [Google Scholar] [CrossRef] [Green Version]
- Meirmans, S.; Meirmans, P.G.; Kirkendall, L.R. The Costs Of Sex: Facing Real-world Complexities. Q. Rev. Biol. 2012, 87, 19–40. [Google Scholar] [CrossRef]
- Lehtonen, J.; Jennions, M.; Kokko, H. The many costs of sex. Trends Ecol. Evol. 2012, 27, 172–178. [Google Scholar] [CrossRef]
- Lipinska, A.P.; Serrano-Serrano, M.L.; Cormier, A.; Peters, A.F.; Kogame, K.; Cock, J.M.; Coelho, S.M. Rapid turnover of life-cycle-related genes in the brown algae. Genome Biol. 2019, 20, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siljestam, M.; Martinossi-Allibert, I. The evolution of anisogamy does not always lead to male competition. bioRxiv 2020. [Google Scholar] [CrossRef]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. The Natural History of Model Organisms: From molecular manipulation of domesticated Chlamydomonas reinhardtii to survival in nature. Elife 2018, 7, e39233. [Google Scholar] [CrossRef]
- Umen, J.G. Volvox and volvocine green algae. EvoDevo 2020, 11, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Note that under both these scenarios of gamete competition and gamete limitation, some proportion of microgametes remain unfertilised at the end of the fertilisation period. Under gamete competition, this is solely due to the greater number of microgametes relative to macrogametes; all macrogametes find a microgamete partner, leaving some microgametes unfertilised and leading to competition for fertilisation amongst the microgametes (see also sperm competition). Meanwhile, under gamete limitation, these unfertilised microgametes are also the result of the short fertilisation period, which limits the opportunities to find an unfertilised macrogametic partner, even if one is present somewhere in the population. This difficulty in finding a partner also affects macrogametes, some of which are then unfertilized under gamete limitation (but not, as we have addressed, under gamete competition).
- Parker, G.A. The sexual cascade and the rise of pre-ejaculatory (Darwinian) sexual selection, sex roles, and sexual conflict. Cold Spring Harb. Perspect. Biol. 2014, 6, a017509. [Google Scholar] [CrossRef] [Green Version]
- Engelstädter, J. Constraints on the evolution of asexual reproduction. BioEssays 2008, 30, 1138–1150. [Google Scholar] [CrossRef]
- Bengtsson, B.O. Asex and evolution: A very large-scale overview. In Lost Sex; Springer: Dordrecht, The Netherlands, 2009; pp. 1–19. [Google Scholar]
- Van der Kooi, C.J.; Schwander, T. On the fate of sexual traits under asexuality. Biol. Rev. 2014, 89, 805–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuilleumier, S.; Alcala, N.; Niculita-Hirzel, H. Transitions from reproductive systems governed by two self-incompatible loci to one in fungi. Evol. Int. J. Org. Evol. 2013, 67, 501–516. [Google Scholar] [CrossRef]
- Czuppon, P.; Rogers, D. Evolution of mating types in finite populations: The precarious advantage of being rare. J. Evol. Biol. 2019, 32, 1290–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krumbeck, Y.; Constable, G.W.A.; Rogers, T. Fitness differences suppress the number of mating types in evolving isogamous species. R. Soc. Open Sci. 2020, 7, 192126. [Google Scholar] [CrossRef] [Green Version]
- Note that in general this second assumption may also be violated when ϕ is finite; although each individual microgamete producer gives rise to more gametes than each individual macrogamete producer, if the sex ratio is R sufficiently low the number of macrogametes could still in principle exceed the number of macrogametes.





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Constable, G.W.A.; Kokko, H. Parthenogenesis and the Evolution of Anisogamy. Cells 2021, 10, 2467. https://doi.org/10.3390/cells10092467
Constable GWA, Kokko H. Parthenogenesis and the Evolution of Anisogamy. Cells. 2021; 10(9):2467. https://doi.org/10.3390/cells10092467
Chicago/Turabian StyleConstable, George W. A., and Hanna Kokko. 2021. "Parthenogenesis and the Evolution of Anisogamy" Cells 10, no. 9: 2467. https://doi.org/10.3390/cells10092467
APA StyleConstable, G. W. A., & Kokko, H. (2021). Parthenogenesis and the Evolution of Anisogamy. Cells, 10(9), 2467. https://doi.org/10.3390/cells10092467