
Preclinical Studies on the Effect of Rucaparib in Ovarian Cancer: Impact of BRCA2 Status





Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Cell Culture
2.3. Cell Viability
2.4. Annexin V/PI Assay
2.5. PARP1 Activity Assay
2.6. Blood Samples
2.7. γ-H2AX Detection in Clinical Samples
2.8. Immunofluorescence—γ-H2AX In Vitro Assay
2.9. RNA Isolation, cDNA Synthesis and Quantitative RT-PCR
2.10. Statistical Analysis
3. Results
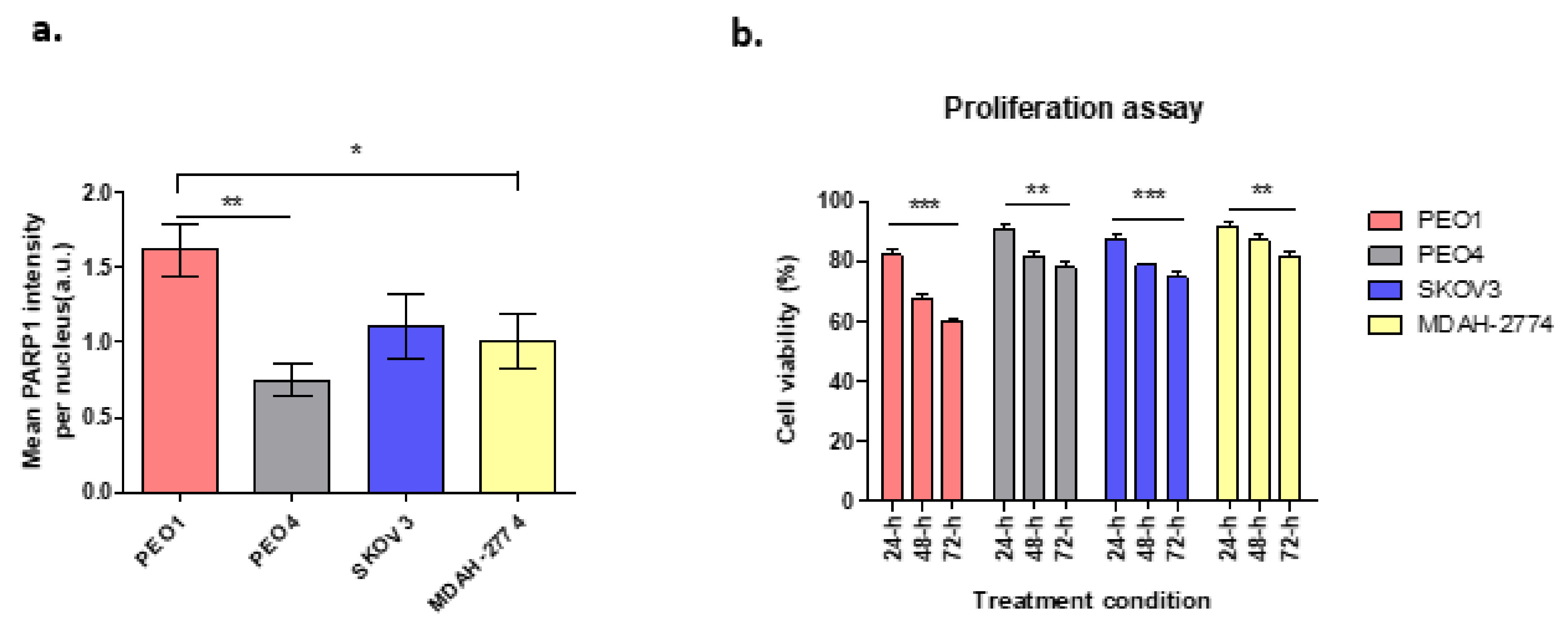
3.1. PARP1 and Cell Proliferation Assay
3.2. Wound Healing Assay
3.3. Rucaparib Induced Apoptosis in Ovarian Cancer Cells
3.4. Rucaparib Treatment Significantly Increased DNA Damage
3.5. Effect of Rucaparib on mTORC1 and mTORC2 Components
3.6. Expression of γ-H2AX in Clinical Samples
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cancer Research UK. Ovarian Cancer. Available online: https://www.cancerresearchuk.org/about-cancer/ovarian-cancer (accessed on 10 May 2021).
- Jayson, G.C.; Kohn, E.C.; Kitchener, H.C.; Ledermann, J.A. Ovarian Cancer. Lancet 2014, 384, 1376–1388. [Google Scholar] [CrossRef]
- The Cancer Genome Atlas Research Network. Integrated Genomic Analyses of Ovarian Carcinoma The Cancer Genome Atlas Research Network. Nature 2011, 474, 609–615. [Google Scholar] [CrossRef]
- Gelmon, K.A.; Tischkowitz, M.; Mackay, H.; Swenerton, K.; Robidoux, A.; Tonkin, K.; Hirte, H.; Huntsman, D.; Clemons, M.; Gilks, B.; et al. Olaparib in Patients with Recurrent High-Grade Serous or Poorly Differentiated Ovarian Carcinoma or Triple-Negative Breast Cancer: A Phase 2, Multicentre, Open-Label, Non-Randomised Study. Lancet Oncol. 2011, 12, 852–861. [Google Scholar] [CrossRef]
- Ledermann, J.; Harter, P.; Gourley, C.; Friedlander, M.; Vergote, I.; Rustin, G.; Scott, C.L.; Meier, W.; Shapira-Frommer, R.; Safra, T.; et al. Olaparib Maintenance Therapy in Patients With Platinum-Sensitive Relapsed Serous Ovarian Cancer. Obstet. Gynecol. Surv. 2012, 69, 594–596. [Google Scholar] [CrossRef]
- El Botty, R.; Coussy, F.; Hatem, R.; Assayag, F.; Servely, J.; Leboucher, S.; Fouillade, C. Inhibition of MTOR Downregulates Expression of DNA Repair Proteins and Is Highly Efficient against BRCA2-Mutated Breast Cancer in Combination to PARP Inhibition. Oncotarget 2018, 9, 29587–29600. [Google Scholar] [CrossRef] [Green Version]
- Ledermann, J.; Harter, P.; Gourley, C.; Friedlander, M.; Vergote, I.; Rustin, G.; Scott, C.L.; Meier, W.; Shapira-Frommer, R.; Safra, T.; et al. Olaparib Maintenance Therapy in Patients with Platinum-Sensitive Relapsed Serous Ovarian Cancer: A Preplanned Retrospective Analysis of Outcomes by BRCA Status in a Randomised Phase 2 Trial. Lancet Oncol. 2014, 15, 852–861. [Google Scholar] [CrossRef]
- Mirza, M.R.; Monk, B.J.; Herrstedt, J.; Oza, A.M.; Mahner, S.; Redondo, A.; Fabbro, M.; Ledermann, J.A.; Lorusso, D.; Vergote, I.; et al. Niraparib Maintenance Therapy in Platinum-Sensitive, Recurrent Ovarian Cancer. N. Engl. J. Med. 2016, 375, 2154–2164. [Google Scholar] [CrossRef] [PubMed]
- McNeish, I.A.; Oza, A.; Coleman, R.L.; Scott, C.L.; Konecny, G.E.; Tinker, A.V.; O’Malley, D.M.; Brenton, J.D.; Kristeleit, R.S.; Bell-McGuin, K.; et al. Results of ARIEL2: A Phase 2 Trial to Prospectively Identify Ovarian Cancer Patients Likely to Respond to Rucaparib Using Tumor. Genetic Analysis. J. Clin. Oncol. 2015, 33, 5508. [Google Scholar] [CrossRef]
- Coleman, R.L.; Swisher, E.M.; Oza, A.M.; Scott, C.L.; Giordano, H.; Lin, K.K.; Konecny, G.E.; Tinker, A.V.; O’Malley, D.M.; Kristeleit, R.S.; et al. Refinement of Prespecified Cutoff for Genomic Loss of Heterozygosity (LOH) in ARIEL2 Part 1: A Phase II Study of Rucaparib in Patients (Pts) with High Grade Ovarian Carcinoma (HGOC). J. Clin. Oncol. 2016, 34, 5540. [Google Scholar] [CrossRef]
- Ivashkevich, A.; Redon, C.E.; Nakamura, A.J.; Martin, R.F.; Martin, O.A. Use of the γ-H2AX Assay to Monitor DNA Damage and Repair in Translational Cancer Research. Cancer Lett. 2012, 327, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Redon, C.E.; Nakamura, A.J.; Zhang, Y.W.; Ji, J.; Bonner, W.M.; Kinders, R.J.; Parchment, R.E.; Doroshow, J.H.; Pommier, Y. Histone ΓH2AX and Poly(ADP-Ribose) as Clinical Pharmacodynamic Biomarkers. Clin. Cancer Res. 2010, 16, 4532–4542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.M.; Gordon, N.; Trepel, J.B.; Lee, M.J.; Yu, M.; Kohn, E.C. Development of a Multiparameter Flow Cytometric Assay as a Potential Biomarker for Homologous Recombination Deficiency in Women with High-Grade Serous Ovarian Cancer. J. Transl. Med. 2015, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Villa, A.; Balasubramanian, P.; Miller, B.L.; Lustberg, M.B.; Ramaswamy, B.; Chalmers, J.J. Assessment of γ-H2AX Levels in Circulating Tumor Cells from Patients Receiving Chemotherapy. Front. Oncol. 2012, 2, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamm, N.; Rogers, S.; Cesare, A.J. The MTOR Pathway: Implications for DNA Replication. Prog. Biophys. Mol. Biol. 2019, 147, 17–25. [Google Scholar] [CrossRef]
- Chopra, S.S.; Jenney, A.; Palmer, A.; Niepel, M.; Chung, M.; Mills, C.; Sivakumaren, S.C.; Liu, Q.; Chen, J.; Asara, J.; et al. Torin2 Exploits Replication and Checkpoint Vulnerabilities to Cause Death of PI3K-Activated Triple-Negative Breast Cancer Cells. Cell Syst. 2020, 10, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Rogers-Broadway, K.R.; Kumar, J.; Sisu, C.; Wander, G.; Mazey, E.; Jeyaneethi, J.; Pados, G.; Tsolakidis, D.; Klonos, E.; Grunt, T.; et al. Differential Expression of MTOR Components in Endometriosis and Ovarian Cancer: Effects of Rapalogues and Dual Kinase Inhibitors on MTORC1 and MTORC2 Stoichiometry. Int. J. Mol. Med. 2019, 43, 47–56. [Google Scholar] [CrossRef]
- Chudasama, D.; Bo, V.; Hall, M.; Anikin, V.; Jeyaneethi, J.; Gregory, J.; Pados, G.; Tucker, A.; Harvey, A.; Pink, R.; et al. Identification of Cancer Biomarkers of Prognostic Value Using Specific Gene Regulatory Networks (GRN): A Novel Role of RAD51AP1 for Ovarian and Lung Cancers. Carcinogenesis 2018, 39, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Nile, D.L.; Rae, C.; Hyndman, I.J.; Gaze, M.N.; Mairs, R.J. An Evaluation in Vitro of PARP-1 Inhibitors, Rucaparib and Olaparib, as Radiosensitisers for the Treatment of Neuroblastoma. BMC Cancer 2016, 16, 621. [Google Scholar] [CrossRef] [Green Version]
- Bryant, H.E.; Schultz, N.; Thomas, H.D.; Parker, K.M.; Flower, D.; Lopez, E.; Kyle, S.; Meuth, M.; Curtin, N.J.; Helleday, T. Specific Killing of BRCA2-Deficient Tumours with Inhibitors of Poly(ADP-Ribose) Polymerase. Nature 2005, 434, 913–917. [Google Scholar] [CrossRef]
- Coleman, R.L.; Oza, A.M.; Lorusso, D.; Aghajanian, C.; Oaknin, A.; Dean, A.; Colombo, N.; Weberpals, J.I.; Clamp, A.; Scambia, G.; et al. Rucaparib Maintenance Treatment for Recurrent Ovarian Carcinoma after Response to Platinum Therapy (ARIEL3): A Randomised, Double-Blind, Placebo-Controlled, Phase 3 Trial. Lancet 2017, 390, 1949–1961. [Google Scholar] [CrossRef] [Green Version]
- Foo, T.; George, A.; Banerjee, S. PARP Inhibitors in Ovarian Cancer: An Overview of the Practice-Changing Trials. Genes Chromosom. Cancer 2021, 60, 385–397. [Google Scholar] [CrossRef]
- Kumar, J.; Murugaiah, V.; Sotiriadis, G.; Kaur, A. Surfactant Protein D as a Potential Biomarker and Therapeutic Target in Ovarian Cancer. Front. Oncol. 2019, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jebali, A.; Dumaz, N. The Role of RICTOR Downstream of Receptor Tyrosine Kinase in Cancers. Mol. Cancer 2018, 17, 1–10. [Google Scholar] [CrossRef]
- Ramaiah, M.J.; Kumar, K.R. MTOR-Rictor-EGFR Axis in Oncogenesis and Diagnosis of Glioblastoma Multiforme. Mol. Biol. Rep. 2021, 48, 4813–4835. [Google Scholar] [CrossRef]
- Catena, V.; Fanciulli, M. Deptor: Not Only a MTOR Inhibitor. J. Exp. Clin. Cancer Res. 2017, 36, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Srinivas, K.P.; Viji, R.; Dan, V.M.; Sajitha, I.S.; Prakash, R.; Rahul, P.V.; Santhoshkumar, T.R.; Lakshmi, S.; Pillai, M.R. DEPTOR Promotes Survival of Cervical Squamous Cell Carcinoma Cells and Its Silencing Induces Apoptosis through Downregulating PI3K/AKT and by up-Regulating P38 MAP Kinase. Oncotarget 2016, 7, 24154–24171. [Google Scholar] [CrossRef] [Green Version]
- Pei, L.; Xie, P.; Zhou, E.; Yang, Q.; Luo, Y.; Tang, Z. Overexpression of DEP Domain Containing MTOR-Interacting Protein Correlates with Poor Prognosis in Differentiated Thyroid Carcinoma. Mol. Med. Rep. 2011, 4, 817–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, T.R.; Laplante, M.; Thoreen, C.C.; Sancak, Y.; Kang, S.A.; Kuehl, W.M.; Gray, N.S.; Sabatini, D.M. DEPTOR Is an MTOR Inhibitor Frequently Overexpressed in Multiple Myeloma Cells and Required for Their Survival. Cell 2009, 137, 873–886. [Google Scholar] [CrossRef] [Green Version]
- Foster, H.; Coley, H.M.; Goumenou, A.; Pados, G.; Harvey, A.; Karteris, E. Differential Expression of MTOR Signalling Components in Drug Resistance in Ovarian Cancer. Anticancer Res. 2010, 30, 3529–3534. [Google Scholar] [PubMed]
- Parvani, J.G.; Davuluri, G.; Wendt, M.K.; Espinosa, C.; Tian, M.; Danielpour, D.; Sossey-Alaoui, K.; Schiemann, W.P. Deptor Enhances Triple-Negative Breast Cancer Metastasis and Chemoresistance through Coupling to Survivin Expression. Neoplasia 2015, 17, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clamp, A.; Jayson, G. PARP Inhibitors in BRCA Mutation-Associated Ovarian Cancer. Lancet Oncol. 2015, 16, 10–12. [Google Scholar] [CrossRef]
- Wang, L.H.; Pfister, T.D.; Parchment, R.E.; Kummar, S.; Rubinstein, L.; Evrard, Y.A.; Gutierrez, M.E.; Murgo, A.J.; Tomaszewski, J.E.; Doroshow, J.H.; et al. Monitoring Drug-Induced ΓH2AX as a Pharmacodynamic Biomarker in Individual Circulating Tumor Cells. Clin. Cancer Res. 2010, 16, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | History | Mutations | Pathology |
|---|---|---|---|
| PEO1 | Derived from ascites | TP53, chemosensitive BRCA2-mutant | HGSOC |
| PEO4 | BRCA2-proficient line acquired by reversion mutation from PEO1 line following chemotherapy | TP53, BRCA2 (silent) | HGSOC |
| SKOV3 | Derived from ascites | TP53, NF1, PIK3CA, HRAS, ARID1A, ERBB2 | Thought to be HGSOC |
| MDAH-2774 | Derived from ascites | TP53, KRAS, PIK3CA, ARID1A, BRCA1/2 (silent) | Endometrioid |
| Gene name | Orientation | Sequence |
|---|---|---|
| YWHAZ | Forward | 5′-AGACGGAAGGTGCTGAGAAA-3′ |
| Reverse | 5′-GAAGCATTGGGGATCAAGAA-3′ | |
| mTOR | Forward | 5′-TGCCAACTACCTTCGGAACC-3′ |
| Reverse | 5′-GCTCGCTTCACCTCAATTC-3′ | |
| DEPTOR | Forward | 5′-CACCATGTGTGTATGAGCA-3′ |
| Reverse | 5′-TGAAGGTGCGCGCTCATTG-3′ | |
| Rictor | Forward | 5′-GGAAGCCTGTTGATGGTGAT-3′ |
| Reverse | 5′-GGCAGCCTTTTTATGGTGT-3′ | |
| Raptor | Forward | 5′-ACTGATGGAGTCCGAATGC-3′ |
| Reverse | 5′-TCATCCGATCCTTCATC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saravi, S.; Alizzi, Z.; Tosi, S.; Hall, M.; Karteris, E. Preclinical Studies on the Effect of Rucaparib in Ovarian Cancer: Impact of BRCA2 Status. Cells 2021, 10, 2434. https://doi.org/10.3390/cells10092434
Saravi S, Alizzi Z, Tosi S, Hall M, Karteris E. Preclinical Studies on the Effect of Rucaparib in Ovarian Cancer: Impact of BRCA2 Status. Cells. 2021; 10(9):2434. https://doi.org/10.3390/cells10092434
Chicago/Turabian StyleSaravi, Sayeh, Zena Alizzi, Sabrina Tosi, Marcia Hall, and Emmanouil Karteris. 2021. "Preclinical Studies on the Effect of Rucaparib in Ovarian Cancer: Impact of BRCA2 Status" Cells 10, no. 9: 2434. https://doi.org/10.3390/cells10092434
APA StyleSaravi, S., Alizzi, Z., Tosi, S., Hall, M., & Karteris, E. (2021). Preclinical Studies on the Effect of Rucaparib in Ovarian Cancer: Impact of BRCA2 Status. Cells, 10(9), 2434. https://doi.org/10.3390/cells10092434