
Coupling DNA Replication and Spindle Function in Saccharomyces cerevisiae


{kind=link}
{kind=link}
Abstract
:1. Introduction
- To what extent are DNA replication and spindle function coordinated in yeast cells?
- Which molecular mechanisms control spindle dynamics upon replication failure?
- What is the influence of spindle-dependent forces on genome stability?
2. Coordinating Replication with Anaphase Spindle Elongation
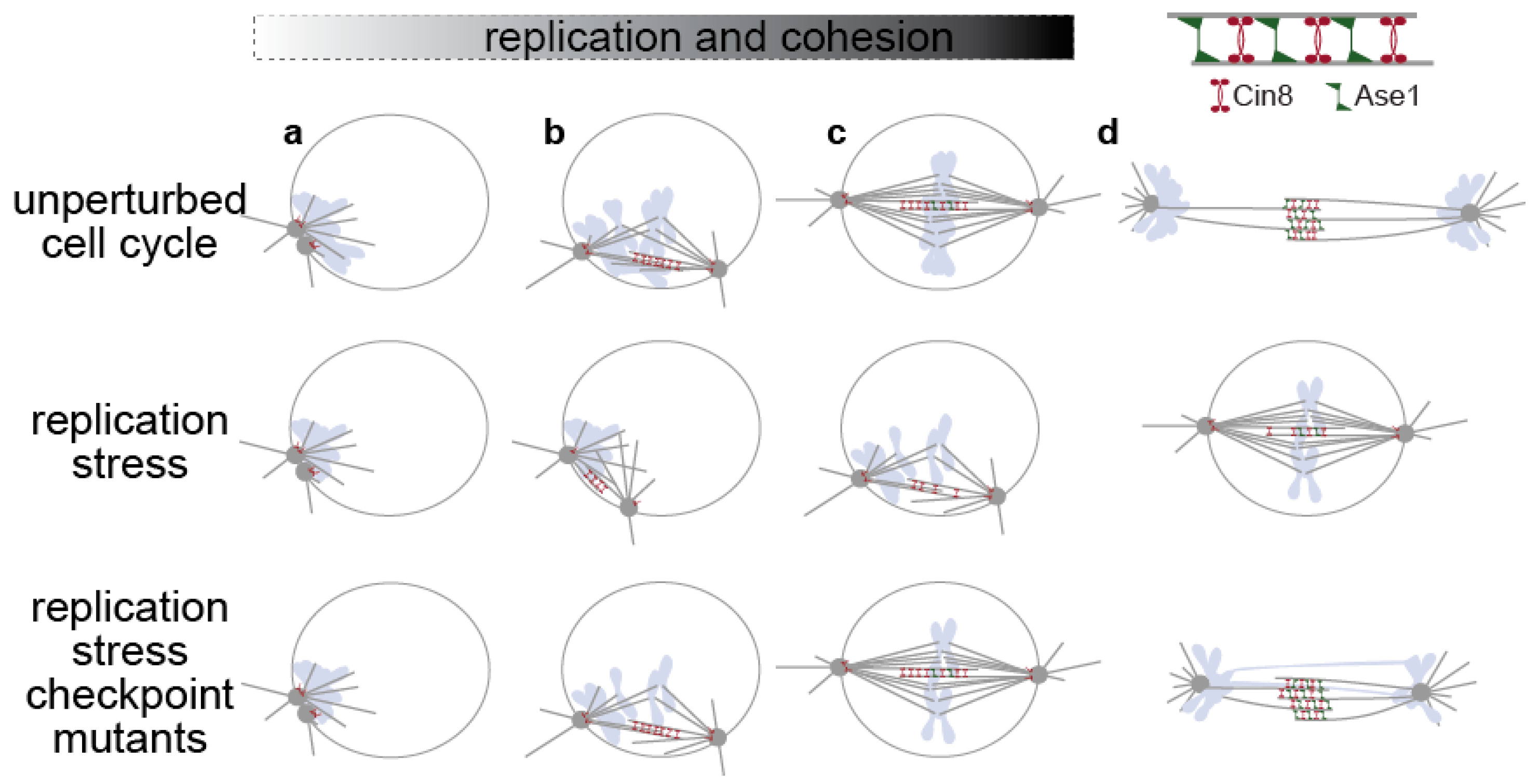
2.1. The Order of Replication and Spindle Formation in Yeast Cells
2.2. Cells under Replication Stress: Inhibition of Spindle Elongation
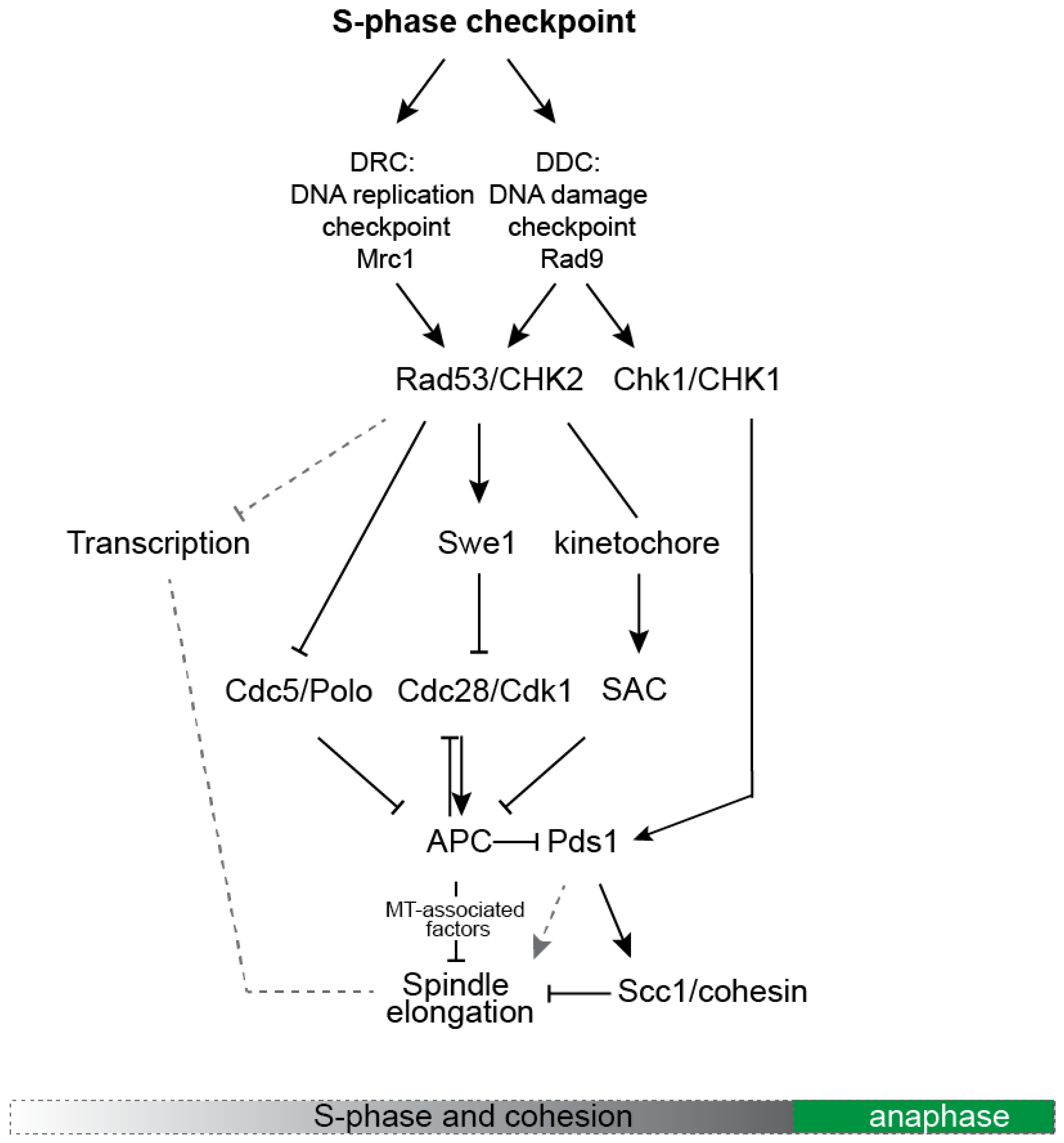
3. Mechanisms Coordinating Replication with Spindle Dynamics
3.1. Kinetochore Integrity Links Replication to Spindle Function
3.2. MT-Associated Targets of the S-Phase Checkpoint and Their Modes of Control
4. Influence of Spindle-Dependent Forces on Genome Stability
5. Conclusions and Future Prospects
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Farrell, P.H.; Stumpff, J.; Tin Su, T. Embryonic Cleavage Cycles: How Is a Mouse Like a Fly? Curr. Biol. 2004, 14, R35–R45. [Google Scholar] [CrossRef]
- Moreno, A.; Carrington, J.T.; Albergante, L.; Al Mamun, M.; Haagensen, E.J.; Komseli, E.-S.; Gorgoulis, V.G.; Newman, T.J.; Blow, J.J. Unreplicated DNA remaining from unperturbed S-phases passes through mitosis for resolution in daughter cells. Proc. Natl. Acad. Sci. USA 2016, 113, E5757–E5764. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, T.; Maier, M.; Missarova, A.; Ziegler-Birling, C.; Dam, M.; Gomar-Alba, M.; Carey, L.B.; Mendoza, M. Budding yeast complete DNA synthesis after chromosome segregation begins. Nat. Commun. 2020, 11, 2267. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.P.; Dutta, A. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 2002, 71, 333–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crasta, K.; Huang, P.; Morgan, G.; Winey, M.; Surana, U. Cdk1 regulates centrosome separation by restraining proteolysis of microtubule-associated proteins. EMBO J. 2006, 25, 2551–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broach, J.R.; Pringle, J.R.; Jones, E.W. The Molecular and Cellular Biology of the Yeast Saccharomyces; Cold Spring Harbor Monograph Series 21; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1991; ISBN 9780879693558. [Google Scholar]
- Hoyt, M.A.; He, L.; Loo, K.K.; Saunders, W.S. Two Saccharomyces cerevisiae kinesin-related gene products required for mitotic spindle assembly. J. Cell Biol. 1992, 118, 109–120. [Google Scholar] [CrossRef]
- Magiera, M.M.; Gueydon, E.; Schwob, E. DNA replication and spindle checkpoints cooperate during S-phase to delay mitosis and preserve genome integrity. J. Cell Biol. 2014, 204, 165–175. [Google Scholar] [CrossRef]
- Musacchio, A.; Salmon, E.D. The spindle-assembly checkpoint in space and time. Nat. Rev. Mol. Cell Biol. 2007, 8, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Stegmeier, F.; Amon, A. Closing Mitosis: The Functions of the Cdc14 Phosphatase and Its Regulation. Annu. Rev. Genet. 2004, 38, 203–232. [Google Scholar] [CrossRef]
- Torres-Rosell, J.; De Piccoli, G.; Aragón, L. Can eukaryotic cells monitor the presence of unreplicated DNA? Cell Div. 2007, 2, 19. [Google Scholar] [CrossRef] [Green Version]
- Pardo, B.; Crabbé, L.; Pasero, P. Signaling Pathways of Replication Stress in Yeast. FEMS Yeast Res. 2016, 17, fow101. [Google Scholar] [CrossRef] [PubMed]
- Bacal, J.; Moriel-Carretero, M.; Pardo, B.; Barthe, A.; Sharma, S.; Chabes, A.; Lengronne, A.; Pasero, P. Mrc1 and Rad9 cooperate to regulate initiation and elongation of DNA replication in response to DNA damage. EMBO J. 2018, 37, e99319. [Google Scholar] [CrossRef]
- Moriel-Carretero, M.; Pasero, P.; Pardo, B. DDR Inc., one business, two associates. Curr. Genet. 2019, 65, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Piatti, S.; Lengauer, C.; Nasmyth, K. Cdc6 is an unstable protein whose de novo synthesis in G1 is important for the onset of S-phase and for preventing a “reductional” anaphase in the budding yeast Saccharomyces cerevisiae. EMBO J. 1995, 14, 3788–3799. [Google Scholar] [CrossRef] [PubMed]
- Severin, F.; Hyman, A.A.; Piatti, S. Correct spindle elongation at the metaphase/anaphase transition is an APC-dependent event in budding yeast. J. Cell Biol. 2001, 155, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Rosell, J.; De Piccoli, G.; Cordon-Preciado, V.; Farmer, S.; Jarmuz, A.; Machin, F.; Pasero, P.; Lisby, M.; Haber, J.E.; Aragon, L. Anaphase Onset Before Complete DNA Replication with Intact Checkpoint Responses. Science 2007, 315, 1411–1415. [Google Scholar] [CrossRef]
- Clarke, D.J.; Segal, M.; Jensen, S.; Reed, S.I. Mec1p regulates Pds1p levels in S-phase: Complex coordination of DNA replication and mitosis. Nat. Cell Biol. 2001, 3, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.J.; Segal, M.; Mondésert, G.; Reed, S.I. The Pds1 anaphase inhibitor and Mec1 kinase define distinct checkpoints coupling S-phase with mitosis in budding yeast. Curr. Biol. 1999, 9, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Liu, D.; Wang, Y.; Qin, J.; Elledge, S.J. Pds1 phosphorylation in response to DNA damage is essential for its DNA damage checkpoint function. Genes Dev. 2001, 15, 1361–1372. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, R.; Tang, Z.; Yu, H.; Cohen-Fix, O. Two Distinct Pathways for Inhibiting Pds1 Ubiquitination in Response to DNA Damage. J. Biol. Chem. 2003, 278, 45027–45033. [Google Scholar] [CrossRef] [Green Version]
- Cohen-Fix, O.; Koshland, D. The anaphase inhibitor of Saccharomyces cerevisiae Pds1p is a target of the DNA damage checkpoint pathway. Proc. Natl. Acad. Sci. USA 1997, 94, 14361–14366. [Google Scholar] [CrossRef] [Green Version]
- Yam, C.Q.X.; Chia, D.B.; Shi, I.; Lim, H.H.; Surana, U. Dun1, a Chk2-related kinase, is the central regulator of securin-separase dynamics during DNA damage signaling. Nucleic Acids Res. 2020, 48, 6092–6107. [Google Scholar] [CrossRef]
- Palou, R.; Palou, G.; Quintana, D.G. A role for the spindle assembly checkpoint in the DNA damage response. Curr. Genet. 2017, 63, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, R.; Putnam, C.W.; Weinert, T. RAD53, DUN1 and PDS1 define two parallel G2/M checkpoint pathways in budding yeast. EMBO J. 1999, 18, 3173–3185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clerici, M.; Baldo, V.; Mantiero, D.; Lottersberger, F.; Lucchini, G.; Longhese, M.P. A Tel1/MRX-dependent checkpoint inhibits the metaphase-to-anaphase transition after UV irradiation in the absence of Mec1. Mol. Cell. Biol. 2004, 24, 10126–10144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, A.; West, R.R.; McIntosh, J.R.; Hiraoka, Y. A cytoplasmic dynein heavy chain is required for oscillatory nuclear movement of meiotic prophase and efficient meiotic recombination in fission yeast. J. Cell Biol. 1999, 145, 1233–1249. [Google Scholar] [CrossRef] [Green Version]
- Bachant, J.; Jessen, S.R.; Kavanaugh, S.E.; Fielding, C.S. The yeast S-phase checkpoint enables replicating chromosomes to bi-orient and restrain spindle extension during S-phase distress. J. Cell Biol. 2005, 168, 999–1012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palou, G.; Palou, R.; Zeng, F.; Vashisht, A.A.; Wohlschlegel, J.A.; Quintana, D.G. Three Different Pathways Prevent Chromosome Segregation in the Presence of DNA Damage or Replication Stress in Budding Yeast. PLoS Genet. 2015, 11, e1005468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uhlmann, F.; Wernic, D.; Poupart, M.A.; Koonin, E.V.; Nasmyth, K. Cleavage of cohesin by the CD clan protease separin triggers anaphase in yeast. Cell 2000, 103, 375–386. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Nirantar, S.; Lim, H.H.; Sinha, I.; Surana, U. DNA damage checkpoint maintains CDH1 in an active state to inhibit anaphase progression. Dev. Cell 2009, 17, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, V.; Nirantar, S.; Crasta, K.; Cheng, A.Y.H.; Surana, U. DNA Replication Checkpoint Prevents Precocious Chromosome Segregation by Regulating Spindle Behavior. Mol. Cell 2004, 16, 687–700. [Google Scholar] [CrossRef]
- Feng, W.; Bachant, J.; Collingwood, D.; Raghuraman, M.K.; Brewer, B.J. Centromere Replication Timing Determines Different Forms of Genomic Instability in Saccharomyces cerevisiae Checkpoint Mutants During Replication Stress. Genetics 2009, 183, 1249–1260. [Google Scholar] [CrossRef] [Green Version]
- Dewar, J.M.; Lydall, D. Similarities and differences between “uncapped” telomeres and DNA double-strand breaks. Chromosoma 2012, 121, 117–130. [Google Scholar] [CrossRef]
- Li, Y.; Bachant, J.; Alcasabas, A.A.; Wang, Y.; Qin, J.; Elledge, S.J. The mitotic spindle is required for loading of the DASH complex onto the kinetochore. Genes Dev. 2002, 16, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; McQueen, J.; Cuschieri, L.; Vogel, J.; Measday, V. Spc24 and Stu2 Promote Spindle Integrity When DNA Replication Is Stalled. Mol. Biol. Cell 2007, 18, 2805–2816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Julius, J.; Peng, J.; McCulley, A.; Caridi, C.; Arnak, R.; See, C.; Nugent, C.I.; Feng, W.; Bachant, J. Inhibition of spindle extension through the yeast S-phase checkpoint is coupled to replication fork stability and the integrity of centromeric DNA. Mol. Biol. Cell 2019, 30, 2771–2789. [Google Scholar] [CrossRef]
- Popov, A.V.; Karsenti, E. Stu2p and XMAP215: Turncoat microtubule-associated proteins? Trends Cell Biol. 2003, 13, 547–550. [Google Scholar] [CrossRef]
- Severin, F.; Habermann, B.; Huffaker, T.; Hyman, T. Stu2 promotes mitotic spindle elongation in anaphase. J. Cell Biol. 2001, 153, 435–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, V.; Surana, U. Taming the Spindle for Containing the Chromosomes. Cell Cycle 2005, 4, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; Roof, D.M. Degradation of the kinesin Kip1p at anaphase onset is mediated by the anaphase-promoting complex and Cdc20p. Proc. Natl. Acad. Sci. USA 2001, 98, 12515–12520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juang, Y.L.; Huang, J.; Peters, J.M.; McLaughlin, M.E.; Tai, C.Y.; Pellman, D. APC-mediated proteolysis of Ase1 and the morphogenesis of the mitotic spindle. Science 1997, 275, 1311–1314. [Google Scholar] [CrossRef]
- Hildebrandt, E.R.; Hoyt, M.A. Cell cycle-dependent degradation of the Saccharomyces cerevisiae spindle motor Cin8p requires APC(Cdh1) and a bipartite destruction sequence. Mol. Biol. Cell 2001, 12, 3402–3416. [Google Scholar] [CrossRef] [Green Version]
- Zachariae, W.; Schwab, M.; Nasmyth, K.; Seufert, W. Control of cyclin ubiquitination by CDK-regulated binding of Hct1 to the anaphase promoting complex. Science 1998, 282, 1721–1724. [Google Scholar] [CrossRef]
- Rudner, A.D.; Hardwick, K.G.; Murray, A.W. Cdc28 activates exit from mitosis in budding yeast. J. Cell Biol. 2000, 149, 1361–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valerio-Santiago, M.; de Los Santos-Velázquez, A.I.; Monje-Casas, F. Inhibition of the mitotic exit network in response to damaged telomeres. PLoS Genet. 2013, 9, e1003859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, D.J.; Segal, M.; Andrews, C.A.; Rudyak, S.G.; Jensen, S.; Smith, K.; Reed, S.I. S-phase checkpoint controls mitosis via an APC-independent Cdc20p function. Nat. Cell Biol. 2003, 5, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Khondker, S.; Kajjo, S.; Chandler-Brown, D.; Skotheim, J.; Rudner, A.; Ikui, A.E. PP2ACdc55 dephosphorylates Pds1 and inhibits spindle elongation in S. cerevisiae. J. Cell Sci. 2020, 133, jcs243766. [Google Scholar] [CrossRef]
- Jaehnig, E.J.; Kuo, D.; Hombauer, H.; Ideker, T.G.; Kolodner, R.D. Checkpoint kinases regulate a global network of transcription factors in response to DNA damage. Cell Rep. 2013, 4, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Yelamanchi, S.K.; Veis, J.; Anrather, D.; Klug, H.; Ammerer, G. Genotoxic stress prevents Ndd1-dependent transcriptional activation of G2/M-specific genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 2014, 34, 711–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKnight, K.; Liu, H.; Wang, Y. Replicative Stress Induces Intragenic Transcription of the ASE1 Gene that Negatively Regulates Ase1 Activity. Curr. Biol. 2014, 24, 1101–1106. [Google Scholar] [CrossRef] [Green Version]
- Dotiwala, F.; Harrison, J.C.; Jain, S.; Sugawara, N.; Haber, J.E. Mad2 Prolongs DNA Damage Checkpoint Arrest Caused by a Double-Strand Break via a Centromere-Dependent Mechanism. Curr. Biol. 2010, 20, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Ayra-Plasencia, J.; Machín, F. DNA double-strand breaks in telophase lead to coalescence between segregated sister chromatid loci. Nat. Commun. 2019, 10, 2862. [Google Scholar] [CrossRef] [Green Version]
- Thu, Y.M.; Van Riper, S.K.; Higgins, L.; Zhang, T.; Becker, J.R.; Markowski, T.W.; Nguyen, H.D.; Griffin, T.J.; Bielinsky, A.K. Slx5/Slx8 Promotes Replication Stress Tolerance by Facilitating Mitotic Progression. Cell Rep. 2016, 15, 1254–1265. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.-C.; Oram, M.K.; Bielinsky, A.-K. SUMO-Targeted Ubiquitin Ligases and Their Functions in Maintaining Genome Stability. Int. J. Mol. Sci. 2021, 22, 5391. [Google Scholar] [CrossRef] [PubMed]
- Su, X.B.; Wang, M.; Schaffner, C.; Nerusheva, O.O.; Clift, D.; Spanos, C.; Kelly, D.A.; Tatham, M.; Wallek, A.; Wu, Y.; et al. SUMOylation stabilizes sister kinetochore biorientation to allow timely anaphase. J. Cell Biol. 2021, 220, e202005130. [Google Scholar] [CrossRef]
- Strecker, J.; Gupta, G.D.; Zhang, W.; Bashkurov, M.; Landry, M.-C.; Pelletier, L.; Durocher, D. DNA damage signalling targets the kinetochore to promote chromatin mobility. Nat. Cell Biol. 2016, 18, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.K.C.; Chan, J.N.Y.; Strecker, J.; Zhang, W.; Ebrahimi-Ardebili, S.; Lu, T.; Abraham, K.J.; Durocher, D.; Mekhail, K. Perinuclear tethers license telomeric DSBs for a broad kinesin- and NPC-dependent DNA repair process. Nat. Commun. 2015, 6, 7742. [Google Scholar] [CrossRef] [Green Version]
- Laflamme, G.; Sim, S.; Leary, A.; Pascariu, M.; Vogel, J.; D’Amours, D. Interphase Microtubules Safeguard Mitotic Progression by Suppressing an Aurora B-Dependent Arrest Induced by DNA Replication Stress. Cell Rep. 2019, 26, 2875–2889.e3. [Google Scholar] [CrossRef] [Green Version]
- Aiken, J.; Sept, D.; Costanzo, M.; Boone, C.; Cooper, J.A.; Moore, J.K. Genome-wide analysis reveals novel and discrete functions for tubulin carboxy-terminal tails. Curr. Biol. 2014, 24, 1295–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrem, C.; Moore, J.K. Astral microtubule forces alter nuclear organization and inhibit DNA repair in budding yeast. Mol. Biol. Cell 2019, 30, 2000–2013. [Google Scholar] [CrossRef]
- Estrem, C.; Moore, J.K. Help or hindrance: How do microtubule-based forces contribute to genome damage and repair? Curr. Genet. 2020, 66, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Oshidari, R.; Strecker, J.; Chung, D.K.C.; Abraham, K.J.; Chan, J.N.Y.; Damaren, C.J.; Mekhail, K. Nuclear microtubule filaments mediate non-linear directional motion of chromatin and promote DNA repair. Nat. Commun. 2018, 9, 2567. [Google Scholar] [CrossRef] [PubMed]
- Oza, P.; Peterson, C.L. Opening the DNA repair toolbox: Localization of DNA double strand breaks to the nuclear periphery. Cell Cycle 2010, 9, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Nagai, S.; Dubrana, K.; Tsai-Pflugfelder, M.; Davidson, M.B.; Roberts, T.M.; Brown, G.W.; Varela, E.; Hediger, F.; Gasser, S.M.; Krogan, N.J. Functional targeting of DNA damage to a nuclear pore-associated SUMO-dependent ubiquitin ligase. Science 2008, 322, 597–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villoria, M.T.; Ramos, F.; Dueñas, E.; Faull, P.; Cutillas, P.R.; Clemente-Blanco, A. Stabilization of the metaphase spindle by Cdc14 is required for recombinational DNA repair. EMBO J. 2017, 36, 79–101. [Google Scholar] [CrossRef]
- Hurst, V.; Challa, K.; Jonas, F.; Forey, R.; Sack, R.; Seebacher, J.; Schmid, C.D.; Barkai, N.; Shimada, K.; Gasser, S.M.; et al. A regulatory phosphorylation site on Mec1 controls chromatin occupancy of RNA polymerases during replication stress. EMBO J. 2021, 40, e108439. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liakopoulos, D. Coupling DNA Replication and Spindle Function in Saccharomyces cerevisiae. Cells 2021, 10, 3359. https://doi.org/10.3390/cells10123359
Liakopoulos D. Coupling DNA Replication and Spindle Function in Saccharomyces cerevisiae. Cells. 2021; 10(12):3359. https://doi.org/10.3390/cells10123359
Chicago/Turabian StyleLiakopoulos, Dimitris. 2021. "Coupling DNA Replication and Spindle Function in Saccharomyces cerevisiae" Cells 10, no. 12: 3359. https://doi.org/10.3390/cells10123359
APA StyleLiakopoulos, D. (2021). Coupling DNA Replication and Spindle Function in Saccharomyces cerevisiae. Cells, 10(12), 3359. https://doi.org/10.3390/cells10123359