
Bacterial Lymphatic Metastasis in Infection and Immunity


Abstract
:1. The Dogma of Bacterial Invasion
2. Barriers to Studying Bacteria in the Lymphatics
3. Bacteria in the Lymphatics
4. Extracellular Transit of Intracellular Bacteria
5. Role of Virulence in Bacterial Dissemination
6. Importance of Bacterial Dissemination
7. Challenging the Dogma of Bacterial Invasion
8. Initial Events in Infection
9. Anatomy of Bacterial Invasion I: Vessels and Fluid Flow in the Interstitium
10. Effect of Damage on Vessel Permeability and Fluid Flow
11. Anatomy of Bacterial Invasion II: Interactions with Interstitial Extracellular Matrix
12. How Bacterial Properties Influence Invasion
13. Anatomy of Bacterial Invasion III: Path of Lymph Return to the Blood
14. Anatomy of Bacterial Invasion IV: Path of Lymph through the Lymph Node
15. Immune Defence in Lymph Nodes
16. Lymph Nodes as Bacterial Filters
17. Role and Recruitment of Neutrophils in Lymph Nodes
18. Subcapsular Sinus Macrophages in the Lymph Node
19. Medullary Sinus Macrophages in the Lymph Node
20. Bacterial Survival in the Lymph Node
21. Impact of Bacteria in Lymph Nodes on Immunity
22. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fardini, Y.; Wang, X.; Temoin, S.; Nithianantham, S.; Lee, D.; Shoham, M.; Han, Y.W. Fusobacterium nucleatum adhesin FadA binds vascular endothelial cadherin and alters endothelial integrity. Mol. Microbiol. 2011, 82, 1468–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemichez, E.; Lecuit, M.; Nassif, X.; Bourdoulous, S. Breaking the wall: Targeting of the endothelium by pathogenic bacteria. Nat. Rev. Microbiol. 2010, 8, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Spadoni, I.; Zagato, E.; Bertocchi, A.; Paolinelli, R.; Hot, E.; Di Sabatino, A.; Caprioli, F.; Bottiglieri, L.; Oldani, A.; Viale, G.; et al. A gut-vascular barrier controls the systemic dissemination of bacteria. Science 2015, 350, 830–834. [Google Scholar] [CrossRef]
- Rangel, S.M.; Diaz, M.H.; Knoten, C.A.; Zhang, A.; Hauser, A.R. The Role of ExoS in Dissemination of Pseudomonas aeruginosa during Pneumonia. PLoS Pathog. 2015, 11, e1004945. [Google Scholar] [CrossRef] [Green Version]
- Clement, C.C.; Wang, W.; Dzieciatkowska, M.; Cortese, M.; Hansen, K.C.; Becerra, A.; Thangaswamy, S.; Nizamutdinova, I.; Moon, J.Y.; Stern, L.J.; et al. Quantitative Profiling of the Lymph Node Clearance Capacity. Sci. Rep. 2018, 8, 11253. [Google Scholar] [CrossRef]
- Drinker, C.K.; Field, M.E.; Ward, H.K. The Filtering Capacity of Lymph Nodes. J. Exp. Med. 1934, 59, 393–405. [Google Scholar] [CrossRef] [Green Version]
- DeLong, T.G.; Simmons, R.L. Role of lymphatic vessels in bacterial clearance from early soft-tissue infection. Arch. Surg. 1982, 117, 123–128. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Uhr, T. Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 2004, 303, 1662–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribet, D.; Cossart, P. How bacterial pathogens colonize their hosts and invade deeper tissues. Microbes Infect. 2015, 17, 173–183. [Google Scholar] [CrossRef]
- Lubkin, A.; Torres, V.J. Bacteria and endothelial cells: A toxic relationship. Curr. Opin. Microbiol. 2017, 35, 58–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casadevall, A.; Fang, F.C. The intracellular pathogen concept. Mol. Microbiol. 2020, 113, 541–545. [Google Scholar] [CrossRef] [Green Version]
- Banerji, S.; Ni, J.; Wang, S.X.; Clasper, S.; Su, J.; Tammi, R.; Jones, M.; Jackson, D.G. LYVE-1, a new homologue of the CD44 glycoprotein, is a lymph-specific receptor for hyaluronan. J. Cell Biol. 1999, 144, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Oliver, G.; Kipnis, J.; Randolph, G.J.; Harvey, N.L. The Lymphatic Vasculature in the 21(st) Century: Novel Functional Roles in Homeostasis and Disease. Cell 2020, 182, 270–296. [Google Scholar] [CrossRef]
- Oliver, G.; Detmar, M. The rediscovery of the lymphatic system: Old and new insights into the development and biological function of the lymphatic vasculature. Genes Dev. 2002, 16, 773–783. [Google Scholar] [CrossRef] [Green Version]
- St John, A.L.; Ang, W.X.G.; Huang, M.N.; Kunder, C.A.; Chan, E.W.; Gunn, M.D.; Abraham, S.N. S1P-Dependent trafficking of intracellular yersinia pestis through lymph nodes establishes Buboes and systemic infection. Immunity 2014, 41, 440–450. [Google Scholar] [CrossRef] [Green Version]
- Bonneau, M.; Epardaud, M.; Payot, F.; Niborski, V.; Thoulouze, M.I.; Bernex, F.; Charley, B.; Riffault, S.; Guilloteau, L.A.; Schwartz-Cornil, I. Migratory monocytes and granulocytes are major lymphatic carriers of Salmonella from tissue to draining lymph node. J. Leukoc. Biol. 2006, 79, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Abadie, V.; Badell, E.; Douillard, P.; Ensergueix, D.; Leenen, P.J.; Tanguy, M.; Fiette, L.; Saeland, S.; Gicquel, B.; Winter, N. Neutrophils rapidly migrate via lymphatics after Mycobacterium bovis BCG intradermal vaccination and shuttle live bacilli to the draining lymph nodes. Blood 2005, 106, 1843–1850. [Google Scholar] [CrossRef] [Green Version]
- Junt, T.; Moseman, E.A.; Iannacone, M.; Massberg, S.; Lang, P.A.; Boes, M.; Fink, K.; Henrickson, S.E.; Shayakhmetov, D.M.; Di Paolo, N.C.; et al. Subcapsular sinus macrophages in lymph nodes clear lymph-borne viruses and present them to antiviral B cells. Nature 2007, 450, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Iannacone, M.; Moseman, E.A.; Tonti, E.; Bosurgi, L.; Junt, T.; Henrickson, S.E.; Whelan, S.P.; Guidotti, L.G.; von Andrian, U.H. Subcapsular sinus macrophages prevent CNS invasion on peripheral infection with a neurotropic virus. Nature 2010, 465, 1079–1083. [Google Scholar] [CrossRef] [Green Version]
- Elomaa, O.; Kangas, M.; Sahlberg, C.; Tuukkanen, J.; Sormunen, R.; Liakka, A.; Thesleff, I.; Kraal, G.; Tryggvason, K. Cloning of a novel bacteria-binding receptor structurally related to scavenger receptors and expressed in a subset of macrophages. Cell 1995, 80, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Siggins, M.K.; Lynskey, N.N.; Lamb, L.E.; Johnson, L.A.; Huse, K.K.; Pearson, M.; Banerji, S.; Turner, C.E.; Woollard, K.; Jackson, D.G.; et al. Extracellular bacterial lymphatic metastasis drives Streptococcus pyogenes systemic infection. Nat. Commun. 2020, 11, 4697. [Google Scholar] [CrossRef]
- Lynskey, N.N.; Banerji, S.; Johnson, L.A.; Holder, K.A.; Reglinski, M.; Wing, P.A.; Rigby, D.; Jackson, D.G.; Sriskandan, S. Rapid Lymphatic Dissemination of Encapsulated Group A Streptococci via Lymphatic Vessel Endothelial Receptor-1 Interaction. PLoS Pathog. 2015, 11, e1005137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siggins, M.K.; Lynskey, N.N.; Lamb, L.E.; Johnson, L.A.; Huse, K.K.; Pearson, M.; Banerji, S.; Turner, C.E.; Woollard, K.; Jackson, D.G.; et al. Lymphatic Metastasis of Virulent Extracellular Bacteria Drives Systemic Infection. SSRN Cell Sneak Peek 2019. Available online: https://papers.ssrn.com/sol3/papers.cfm?abstract_id=3380255 (accessed on 10 December 2021).
- Lincoln, R.E.; Hodges, D.R.; Klein, F.; Mahlandt, B.G.; Jones, W.I., Jr.; Haines, B.W.; Rhian, M.A.; Walker, J.S. Role of the lymphatics in the pathogenesis of anthrax. J. Infect. Dis. 1965, 115, 481–494. [Google Scholar] [CrossRef]
- Bravo-Blas, A.; Utriainen, L.; Clay, S.L.; Kastele, V.; Cerovic, V.; Cunningham, A.F.; Henderson, I.R.; Wall, D.M.; Milling, S.W.F. Salmonella enterica Serovar Typhimurium Travels to Mesenteric Lymph Nodes Both with Host Cells and Autonomously. J. Immunol. 2019, 202, 260–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, R.J.; Lane, M.C.; Wagner, N.J.; Weening, E.H.; Miller, V.L. Dissemination of a highly virulent pathogen: Tracking the early events that define infection. PLoS Pathog. 2015, 11, e1004587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, I.L., Jr.; Beeson, P.B. Bacteremia: A consideration of some experimental and clinical aspects. Yale J. Biol. Med. 1954, 26, 241–262. [Google Scholar] [PubMed]
- Shannon, J.G.; Hasenkrug, A.M.; Dorward, D.W.; Nair, V.; Carmody, A.B.; Hinnebusch, B.J. Yersinia pestis subverts the dermal neutrophil response in a mouse model of bubonic plague. MBio 2013, 4, e00170-13. [Google Scholar] [CrossRef] [Green Version]
- Balmer, M.L.; Slack, E.; de Gottardi, A.; Lawson, M.A.; Hapfelmeier, S.; Miele, L.; Grieco, A.; Van Vlierberghe, H.; Fahrner, R.; Patuto, N.; et al. The liver may act as a firewall mediating mutualism between the host and its gut commensal microbiota. Sci. Transl. Med. 2014, 6, 237ra266. [Google Scholar] [CrossRef] [Green Version]
- Bar-Haim, E.; Gat, O.; Markel, G.; Cohen, H.; Shafferman, A.; Velan, B. Interrelationship between dendritic cell trafficking and Francisella tularensis dissemination following airway infection. PLoS Pathog. 2008, 4, e1000211. [Google Scholar] [CrossRef] [PubMed]
- Bou Ghanem, E.N.; Jones, G.S.; Myers-Morales, T.; Patil, P.D.; Hidayatullah, A.N.; D’Orazio, S.E. InlA promotes dissemination of Listeria monocytogenes to the mesenteric lymph nodes during food borne infection of mice. PLoS Pathog. 2012, 8, e1003015. [Google Scholar] [CrossRef] [PubMed]
- St John, A.L.; Abraham, S.N. Salmonella disrupts lymph node architecture by TLR4-mediated suppression of homeostatic chemokines. Nat. Med. 2009, 15, 1259–1265. [Google Scholar] [CrossRef]
- Chackerian, A.A.; Alt, J.M.; Perera, T.V.; Dascher, C.C.; Behar, S.M. Dissemination of Mycobacterium tuberculosis is influenced by host factors and precedes the initiation of T-cell immunity. Infect. Immun. 2002, 70, 4501–4509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganchua, S.K.C.; Cadena, A.M.; Maiello, P.; Gideon, H.P.; Myers, A.J.; Junecko, B.F.; Klein, E.C.; Lin, P.L.; Mattila, J.T.; Flynn, J.L. Lymph nodes are sites of prolonged bacterial persistence during Mycobacterium tuberculosis infection in macaques. PLoS Pathog. 2018, 14, e1007337. [Google Scholar] [CrossRef] [PubMed]
- Kastenmuller, W.; Torabi-Parizi, P.; Subramanian, N.; Lammermann, T.; Germain, R.N. A spatially-organized multicellular innate immune response in lymph nodes limits systemic pathogen spread. Cell 2012, 150, 1235–1248. [Google Scholar] [CrossRef] [Green Version]
- Bogoslowski, A.; Butcher, E.C.; Kubes, P. Neutrophils recruited through high endothelial venules of the lymph nodes via PNAd intercept disseminating Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2018, 115, 2449–2454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulz, R.Z.; Warren, M.F.; Drinker, C.K. The Passage of Rabbit Virulent Type Iii Pneumococci from the Respiratory Tract of Rabbits into the Lymphatics and Blood. J. Exp. Med. 1938, 68, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Pullinger, G.D.; Paulin, S.M.; Charleston, B.; Watson, P.R.; Bowen, A.J.; Dziva, F.; Morgan, E.; Villarreal-Ramos, B.; Wallis, T.S.; Stevens, M.P. Systemic translocation of Salmonella enterica serovar Dublin in cattle occurs predominantly via efferent lymphatics in a cell-free niche and requires type III secretion system 1 (T3SS-1) but not T3SS-2. Infect. Immun. 2007, 75, 5191–5199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynskey, N.N.; Lawrenson, R.A.; Sriskandan, S. New understandings in Streptococcus pyogenes. Curr. Opin. Infect. Dis. 2011, 24, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.J.; Miller, V.L. A Deadly Path: Bacterial Spread During Bubonic Plague. Trends Microbiol. 2016, 24, 239–241. [Google Scholar] [CrossRef] [Green Version]
- Shannon, J.G.; Bosio, C.F.; Hinnebusch, B.J. Dermal neutrophil, macrophage and dendritic cell responses to Yersinia pestis transmitted by fleas. PLoS Pathog. 2015, 11, e1004734. [Google Scholar] [CrossRef] [Green Version]
- Sebbane, F.; Gardner, D.; Long, D.; Gowen, B.B.; Hinnebusch, B.J. Kinetics of disease progression and host response in a rat model of bubonic plague. Am. J. Pathol. 2005, 166, 1427–1439. [Google Scholar] [CrossRef] [Green Version]
- Hughes, R.; May, A.J.; Widdicombe, J.G. The role of the lymphatic system in the pathogenesis of anthrax. Br. J. Exp. Pathol. 1956, 37, 343–349. [Google Scholar] [PubMed]
- Reddick, L.E.; Alto, N.M. Bacteria fighting back: How pathogens target and subvert the host innate immune system. Mol. Cell 2014, 54, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Thammavongsa, V.; Kim, H.K.; Missiakas, D.; Schneewind, O. Staphylococcal manipulation of host immune responses. Nat. Rev. Microbiol. 2015, 13, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Angevine, D.M. The Fate of a Virulent Hemolytic Streptococcus Injected into the Skin of Normal and Immunized Rabbits. J. Exp. Med. 1936, 64, 131–147. [Google Scholar] [CrossRef] [Green Version]
- Moxon, E.R.; Kroll, J.S. The role of bacterial polysaccharide capsules as virulence factors. Curr. Top. Microbiol. Immunol. 1990, 150, 65–85. [Google Scholar] [CrossRef]
- Kim, H.K.; Missiakas, D.; Schneewind, O. Mouse models for infectious diseases caused by Staphylococcus aureus. J. Immunol. Methods 2014, 410, 88–99. [Google Scholar] [CrossRef]
- Weinstein, M.P.; Towns, M.L.; Quartey, S.M.; Mirrett, S.; Reimer, L.G.; Parmigiani, G.; Reller, L.B. The clinical significance of positive blood cultures in the 1990s: A prospective comprehensive evaluation of the microbiology, epidemiology, and outcome of bacteremia and fungemia in adults. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1997, 24, 584–602. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Cerda, J.; Cossart, P. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Ashida, H.; Ogawa, M.; Yoshikawa, Y.; Mimuro, H.; Sasakawa, C. Bacterial interactions with the host epithelium. Cell Host Microbe 2010, 8, 20–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.N.; Hagan, E.C.; Lane, M.C.; Mobley, H.L. Dissemination and systemic colonization of uropathogenic Escherichia coli in a murine model of bacteremia. MBio 2010, 1, e00262-10. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, S.M.; Bayer, C.R.; Stevens, D.L.; Lieber, R.L.; Bryant, A.E. Muscle injury, vimentin expression, and nonsteroidal anti-inflammatory drugs predispose to cryptic group A streptococcal necrotizing infection. J. Infect. Dis. 2008, 198, 1692–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.K.K.; Reglinski, M.; Teo, D.; Reza, N.; Lamb, L.E.M.; Nageshwaran, V.; Turner, C.E.; Wikstrom, M.; Frick, I.M.; Bjorck, L.; et al. Vaccine-induced, but not natural immunity, against the Streptococcal inhibitor of complement protects against invasive disease. npj Vaccines 2021, 6, 62. [Google Scholar] [CrossRef] [PubMed]
- Voedisch, S.; Koenecke, C.; David, S.; Herbrand, H.; Forster, R.; Rhen, M.; Pabst, O. Mesenteric lymph nodes confine dendritic cell-mediated dissemination of Salmonella enterica serovar Typhimurium and limit systemic disease in mice. Infect. Immun. 2009, 77, 3170–3180. [Google Scholar] [CrossRef] [Green Version]
- Mellhammar, L.; Kahn, F.; Whitlow, C.; Kander, T.; Christensson, B.; Linder, A. Bacteremic sepsis leads to higher mortality when adjusting for confounders with propensity score matching. Sci. Rep. 2021, 11, 6972. [Google Scholar] [CrossRef] [PubMed]
- Triantafyllou, E.; Gudd, C.L.; Mawhin, M.A.; Husbyn, H.C.; Trovato, F.M.; Siggins, M.K.; O’Connor, T.; Kudo, H.; Mukherjee, S.K.; Wendon, J.A.; et al. PD-1 blockade improves Kupffer cell bacterial clearance in acute liver injury. J. Clin. Investig. 2021, 131, e140196. [Google Scholar] [CrossRef]
- Macpherson, A.J.; Heikenwalder, M.; Ganal-Vonarburg, S.C. The Liver at the Nexus of Host-Microbial Interactions. Cell Host Microbe 2016, 20, 561–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, aau6085. [Google Scholar] [CrossRef]
- Deniset, J.F.; Surewaard, B.G.; Lee, W.Y.; Kubes, P. Splenic Ly6G(high) mature and Ly6G(int) immature neutrophils contribute to eradication of S. pneumoniae. J. Exp. Med. 2017, 214, 1333–1350. [Google Scholar] [CrossRef]
- Zeng, Z.; Surewaard, B.G.; Wong, C.H.; Geoghegan, J.A.; Jenne, C.N.; Kubes, P. CRIg Functions as a Macrophage Pattern Recognition Receptor to Directly Bind and Capture Blood-Borne Gram-Positive Bacteria. Cell Host Microbe 2016, 20, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, P.B.; Brennan, M.T.; Sasser, H.C.; Fox, P.C.; Paster, B.J.; Bahrani-Mougeot, F.K. Bacteremia associated with toothbrushing and dental extraction. Circulation 2008, 117, 3118–3125. [Google Scholar] [CrossRef] [Green Version]
- Francois, M.; Bingen, E.H.; Lambert-Zechovsky, N.Y.; Mariani-Kurkdjian, P.; Nottet, J.B.; Narcy, P. Bacteremia during tonsillectomy. Arch. Otolaryngol. Head Neck Surg. 1992, 118, 1229–1231. [Google Scholar] [CrossRef] [PubMed]
- Ercoli, G.; Fernandes, V.E.; Chung, W.Y.; Wanford, J.J.; Thomson, S.; Bayliss, C.D.; Straatman, K.; Crocker, P.R.; Dennison, A.; Martinez-Pomares, L.; et al. Intracellular replication of Streptococcus pneumoniae inside splenic macrophages serves as a reservoir for septicaemia. Nat. Microbiol. 2018, 3, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Holland, T.L.; Baddour, L.M.; Bayer, A.S.; Hoen, B.; Miro, J.M.; Fowler, V.G., Jr. Infective endocarditis. Nat. Rev. Dis. Primers 2016, 2, 16059. [Google Scholar] [CrossRef]
- Gaya, M.; Castello, A.; Montaner, B.; Rogers, N.; Reis e Sousa, C.; Bruckbauer, A.; Batista, F.D. Host response. Inflammation-induced disruption of SCS macrophages impairs B cell responses to secondary infection. Science 2015, 347, 667–672. [Google Scholar] [CrossRef]
- Kamenyeva, O.; Boularan, C.; Kabat, J.; Cheung, G.Y.; Cicala, C.; Yeh, A.J.; Chan, J.L.; Periasamy, S.; Otto, M.; Kehrl, J.H. Neutrophil recruitment to lymph nodes limits local humoral response to Staphylococcus aureus. PLoS Pathog. 2015, 11, e1004827. [Google Scholar] [CrossRef]
- Cunningham, A.F.; Gaspal, F.; Serre, K.; Mohr, E.; Henderson, I.R.; Scott-Tucker, A.; Kenny, S.M.; Khan, M.; Toellner, K.M.; Lane, P.J.; et al. Salmonella induces a switched antibody response without germinal centers that impedes the extracellular spread of infection. J. Immunol. 2007, 178, 6200–6207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, H.; Turner, C.E.; Siggins, M.K.; El-Bahrawy, M.; Pichon, B.; Kearns, A.; Sriskandan, S. Toxic Shock Syndrome Toxin 1 Evaluation and Antibiotic Impact in a Transgenic Model of Staphylococcal Soft Tissue Infection. Msphere 2019, 4, e00665-19. [Google Scholar] [CrossRef] [Green Version]
- Griffin, A.J.; Li, L.X.; Voedisch, S.; Pabst, O.; McSorley, S.J. Dissemination of persistent intestinal bacteria via the mesenteric lymph nodes causes typhoid relapse. Infect. Immun. 2011, 79, 1479–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaud, B.; Brun-Buisson, C.; Group, I.C.-B.S. Outcomes of primary and catheter-related bacteremia. A cohort and case-control study in critically ill patients. Am. J. Respir. Crit. Care Med. 2001, 163, 1584–1590. [Google Scholar] [CrossRef]
- Moses, A.E.; Goldberg, S.; Korenman, Z.; Ravins, M.; Hanski, E.; Shapiro, M. Invasive group a streptococcal infections, Israel. Emerg. Infect. Dis. 2002, 8, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Nelson, G.E.; Pondo, T.; Toews, K.A.; Farley, M.M.; Lindegren, M.L.; Lynfield, R.; Aragon, D.; Zansky, S.M.; Watt, J.P.; Cieslak, P.R.; et al. Epidemiology of Invasive Group A Streptococcal Infections in the United States, 2005–2012. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 63, 478–486. [Google Scholar] [CrossRef]
- Lamagni, T.L.; Darenberg, J.; Luca-Harari, B.; Siljander, T.; Efstratiou, A.; Henriques-Normark, B.; Vuopio-Varkila, J.; Bouvet, A.; Creti, R.; Ekelund, K.; et al. Epidemiology of severe Streptococcus pyogenes disease in Europe. J. Clin. Microbiol. 2008, 46, 2359–2367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, E.M.; Gudmundsson, S.; Yocum, D.E.; Haselby, R.C.; Craig, W.A.; Sundstrom, W.R. Streptococcal myositis. Arch. Intern. Med. 1985, 145, 1020–1023. [Google Scholar] [CrossRef]
- Stevens, D.L.; Tanner, M.H.; Winship, J.; Swarts, R.; Ries, K.M.; Schlievert, P.M.; Kaplan, E. Severe group A streptococcal infections associated with a toxic shock-like syndrome and scarlet fever toxin A. N. Engl. J. Med. 1989, 321, 1–7. [Google Scholar] [CrossRef]
- Stevens, D.L.; Bryant, A.E. Severe Group A Streptococcal Infections. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Prudent, E.; La Scola, B.; Drancourt, M.; Angelakis, E.; Raoult, D. Molecular strategy for the diagnosis of infectious lymphadenitis. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Wells, C.L.; Jechorek, R.P.; Twiggs, L.B.; Brooker, D.C. Recovery of viable bacteria from pelvic lymph nodes of patients with gynecologic tumors. J. Infect. Dis. 1990, 162, 1216–1218. [Google Scholar] [CrossRef]
- Sakamoto, H.; Naito, H.; Ohta, Y.; Tanakna, R.; Maeda, N.; Sasaki, J.; Nord, C.E. Isolation of bacteria from cervical lymph nodes in patients with oral cancer. Arch. Oral Biol. 1999, 44, 789–793. [Google Scholar] [CrossRef]
- Sakamoto, H.; Sasaki, J.; Nord, C.E. Association between bacterial colonization on the tumor, bacterial translocation to the cervical lymph nodes and subsequent postoperative infection in patients with oral cancer. Clin. Microbiol. Infect. 1999, 5, 612–616. [Google Scholar] [CrossRef] [Green Version]
- O’Boyle, C.J.; MacFie, J.; Mitchell, C.J.; Johnstone, D.; Sagar, P.M.; Sedman, P.C. Microbiology of bacterial translocation in humans. Gut 1998, 42, 29–35. [Google Scholar] [CrossRef]
- Stevens, D.L.; Bryant, A.E. Impetigo, Erysipelas and Cellulitis. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2016. [Google Scholar]
- Stevens, D.L.; Bryant, A.E. Necrotizing Soft-Tissue Infections. N. Engl. J. Med. 2017, 377, 2253–2265. [Google Scholar] [CrossRef]
- Ackermann, M.; Stecher, B.; Freed, N.E.; Songhet, P.; Hardt, W.D.; Doebeli, M. Self-destructive cooperation mediated by phenotypic noise. Nature 2008, 454, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Cywes, C.; Wessels, M.R. Group A Streptococcus tissue invasion by CD44-mediated cell signalling. Nature 2001, 414, 648–652. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern recognition receptors and inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Moore, J.E., Jr.; Bertram, C.D. Lymphatic System Flows. Annu. Rev. Fluid Mech. 2018, 50, 459–482. [Google Scholar] [CrossRef]
- Polomska, A.K.; Proulx, S.T. Imaging technology of the lymphatic system. Adv. Drug Deliv. Rev. 2021, 170, 294–311. [Google Scholar] [CrossRef] [PubMed]
- Sarin, H. Physiologic upper limits of pore size of different blood capillary types and another perspective on the dual pore theory of microvascular permeability. J. Angiogenesis Res. 2010, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Lea, A.W. Some Remarks on Puerperal Infection. Br. Med. J. 1905, 1, 581–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, J. Puerperal Infection. Postgrad. Med. J. 1931, 6, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baluk, P.; Fuxe, J.; Hashizume, H.; Romano, T.; Lashnits, E.; Butz, S.; Vestweber, D.; Corada, M.; Molendini, C.; Dejana, E.; et al. Functionally specialized junctions between endothelial cells of lymphatic vessels. J. Exp. Med. 2007, 204, 2349–2362. [Google Scholar] [CrossRef] [PubMed]
- Leak, L.V.; Burke, J.F. Ultrastructural studies on the lymphatic anchoring filaments. J. Cell Biol. 1968, 36, 129–149. [Google Scholar] [CrossRef] [PubMed]
- Ikomi, F.; Hunt, J.; Hanna, G.; Schmid-Schonbein, G.W. Interstitial fluid, plasma protein, colloid, and leukocyte uptake into initial lymphatics. J. Appl. Physiol. 1996, 81, 2060–2067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzewik, J.; Mallipattu, S.K.; Artmann, G.M.; Delano, F.A.; Schmid-Schonbein, G.W. Evidence for a second valve system in lymphatics: Endothelial microvalves. FASEB J. 2001, 15, 1711–1717. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.A.; Banerji, S.; Lawrance, W.; Gileadi, U.; Prota, G.; Holder, K.A.; Roshorm, Y.M.; Hanke, T.; Cerundolo, V.; Gale, N.W.; et al. Dendritic cells enter lymph vessels by hyaluronan-mediated docking to the endothelial receptor LYVE-1. Nat. Immunol. 2017, 18, 762–770. [Google Scholar] [CrossRef] [PubMed]
- Jamalian, S.; Jafarnejad, M.; Zawieja, S.D.; Bertram, C.D.; Gashev, A.A.; Zawieja, D.C.; Davis, M.J.; Moore, J.E., Jr. Demonstration and Analysis of the Suction Effect for Pumping Lymph from Tissue Beds at Subatmospheric Pressure. Sci. Rep. 2017, 7, 12080. [Google Scholar] [CrossRef]
- Jackson, D.G. Lymphatic Regulation of Cellular Trafficking. J. Clin. Cell Immunol 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Jackson, D.G. Leucocyte Trafficking via the Lymphatic Vasculature- Mechanisms and Consequences. Front. Immunol. 2019, 10, 471. [Google Scholar] [CrossRef]
- Oh, S.Y.; Budzik, J.M.; Garufi, G.; Schneewind, O. Two capsular polysaccharides enable Bacillus cereus G9241 to cause anthrax-like disease. Mol. Microbiol. 2011, 80, 455–470. [Google Scholar] [CrossRef] [Green Version]
- Guss, B.; Flock, M.; Frykberg, L.; Waller, A.S.; Robinson, C.; Smith, K.C.; Flock, J.I. Getting to grips with strangles: An effective multi-component recombinant vaccine for the protection of horses from Streptococcus equi infection. PLoS Pathog. 2009, 5, e1000584. [Google Scholar] [CrossRef] [Green Version]
- Harper, M.; Boyce, J.D.; Adler, B. Pasteurella multocida pathogenesis: 125 years after Pasteur. FEMS Microbiol. Lett. 2006, 265, 1–10. [Google Scholar] [CrossRef]
- McMaster, P.D.; Hudack, S.S. The Participation of Skin Lymphatics in Repair of the Lesions Due to Incisions and Burns. J. Exp. Med. 1934, 60, 479–501. [Google Scholar] [CrossRef] [Green Version]
- Barnes, J.M.; Trueta, J. Absorption of Bacteria, Toxins and Snake Venoms from the Tissues: Importance of the Lymphatic Circulation. Lancet 1941, 237, 623–626. [Google Scholar] [CrossRef]
- Claesson-Welsh, L.; Dejana, E.; McDonald, D.M. Permeability of the Endothelial Barrier: Identifying and Reconciling Controversies. Trends Mol. Med. 2021, 27, 314–331. [Google Scholar] [CrossRef]
- McDonald, D.M.; Thurston, G.; Baluk, P. Endothelial gaps as sites for plasma leakage in inflammation. Microcirculation 1999, 6, 7–22. [Google Scholar] [CrossRef]
- Miteva, D.O.; Rutkowski, J.M.; Dixon, J.B.; Kilarski, W.; Shields, J.D.; Swartz, M.A. Transmural flow modulates cell and fluid transport functions of lymphatic endothelium. Circ. Res. 2010, 106, 920–931. [Google Scholar] [CrossRef] [PubMed]
- Baluk, P.; Hirata, A.; Thurston, G.; Fujiwara, T.; Neal, C.R.; Michel, C.C.; McDonald, D.M. Endothelial gaps: Time course of formation and closure in inflamed venules of rats. Am. J. Physiol. 1997, 272, L155–L170. [Google Scholar] [CrossRef]
- Baffert, F.; Le, T.; Thurston, G.; McDonald, D.M. Angiopoietin-1 decreases plasma leakage by reducing number and size of endothelial gaps in venules. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H107–H118. [Google Scholar] [CrossRef]
- Aldrich, M.B.; Sevick-Muraca, E.M. Cytokines are systemic effectors of lymphatic function in acute inflammation. Cytokine 2013, 64, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Cromer, W.E.; Zawieja, S.D.; Tharakan, B.; Childs, E.W.; Newell, M.K.; Zawieja, D.C. The effects of inflammatory cytokines on lymphatic endothelial barrier function. Angiogenesis 2014, 17, 395–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chary, S.R.; Jain, R.K. Direct measurement of interstitial convection and diffusion of albumin in normal and neoplastic tissues by fluorescence photobleaching. Proc. Natl. Acad. Sci. USA 1989, 86, 5385–5389. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Young, A.J.; West, C.A.; Su, M.; Konerding, M.A.; Mentzer, S.J. Stimulation of regional lymphatic and blood flow by epicutaneous oxazolone. J. Appl. Physiol. 2002, 93, 966–973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohlen, H.G.; Wang, W.; Gashev, A.; Gasheva, O.; Zawieja, D. Phasic contractions of rat mesenteric lymphatics increase basal and phasic nitric oxide generation in vivo. Am. J. Physiol. Heart Circ. Physiol. 2009, 297, H1319–H1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamb, L.E.; Siggins, M.K.; Scudamore, C.; Macdonald, W.; Turner, C.E.; Lynskey, N.N.; Tan, L.K.K.; Sriskandan, S. Impact of contusion injury on intramuscular emm1 group a streptococcus infection and lymphatic spread. Virulence 2018, 9, 1074–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swartz, M.A.; Berk, D.A.; Jain, R.K. Transport in lymphatic capillaries. I. Macroscopic measurements using residence time distribution theory. Am. J. Physiol. 1996, 270, H324–H329. [Google Scholar] [CrossRef]
- Reddy, S.T.; van der Vlies, A.J.; Simeoni, E.; Angeli, V.; Randolph, G.J.; O’Neil, C.P.; Lee, L.K.; Swartz, M.A.; Hubbell, J.A. Exploiting lymphatic transport and complement activation in nanoparticle vaccines. Nat. Biotechnol. 2007, 25, 1159–1164. [Google Scholar] [CrossRef]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 2008, 38, 1404–1413. [Google Scholar] [CrossRef]
- Trevaskis, N.L.; Kaminskas, L.M.; Porter, C.J. From sewer to saviour-targeting the lymphatic system to promote drug exposure and activity. Nat. Rev. Drug Discov. 2015, 14, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Supersaxo, A.; Hein, W.R.; Steffen, H. Effect of molecular weight on the lymphatic absorption of water-soluble compounds following subcutaneous administration. Pharm. Res. 1990, 7, 167–169. [Google Scholar] [CrossRef]
- Abel, S.; Abel zur Wiesch, P.; Davis, B.M.; Waldor, M.K. Analysis of Bottlenecks in Experimental Models of Infection. PLoS Pathog. 2015, 11, e1004823. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.H.; Voedisch, S.; Wahl, B.; Rouf, S.F.; Geffers, R.; Rhen, M.; Pabst, O. Independent bottlenecks characterize colonization of systemic compartments and gut lymphoid tissue by salmonella. PLoS Pathog. 2014, 10, e1004270. [Google Scholar] [CrossRef]
- Kono, M.; Zafar, M.A.; Zuniga, M.; Roche, A.M.; Hamaguchi, S.; Weiser, J.N. Single Cell Bottlenecks in the Pathogenesis of Streptococcus pneumoniae. PLoS Pathog. 2016, 12, e1005887. [Google Scholar] [CrossRef]
- Moxon, E.R.; Murphy, P.A. Haemophilus influenzae bacteremia and meningitis resulting from survival of a single organism. Proc. Natl. Acad. Sci. USA 1978, 75, 1534–1536. [Google Scholar] [CrossRef] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef] [PubMed]
- Mouw, J.K.; Ou, G.; Weaver, V.M. Extracellular matrix assembly: A multiscale deconstruction. Nat. Rev. Mol. Cell Biol. 2014, 15, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Frantz, C.; Stewart, K.M.; Weaver, V.M. The extracellular matrix at a glance. J. Cell Sci. 2010, 123, 4195–4200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Porter, C.J. Drug delivery to the lymphatic system. Crit. Rev. Drug Carr. Syst. 1997, 14, 333–393. [Google Scholar]
- Comper, W.D.; Laurent, T.C. Physiological function of connective tissue polysaccharides. Physiol. Rev. 1978, 58, 255–315. [Google Scholar] [CrossRef]
- Wiig, H.; Swartz, M.A. Interstitial fluid and lymph formation and transport: Physiological regulation and roles in inflammation and cancer. Physiol. Rev. 2012, 92, 1005–1060. [Google Scholar] [CrossRef]
- Swartz, M.A. The physiology of the lymphatic system. Adv. Drug Deliv. Rev. 2001, 50, 3–20. [Google Scholar] [CrossRef]
- Rao, D.A.; Forrest, M.L.; Alani, A.W.; Kwon, G.S.; Robinson, J.R. Biodegradable PLGA based nanoparticles for sustained regional lymphatic drug delivery. J. Pharm. Sci. 2010, 99, 2018–2031. [Google Scholar] [CrossRef] [PubMed]
- Bagby, T.R.; Duan, S.; Cai, S.; Yang, Q.; Thati, S.; Berkland, C.; Aires, D.J.; Forrest, M.L. Lymphatic trafficking kinetics and near-infrared imaging using star polymer architectures with controlled anionic character. Eur. J. Pharm. Sci. 2012, 47, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlin, H.; Piccinini, A.M. A complex interplay between the extracellular matrix and the innate immune response to microbial pathogens. Immunology 2018, 155, 186–201. [Google Scholar] [CrossRef]
- Sorokin, L. The impact of the extracellular matrix on inflammation. Nat. Rev. Immunol. 2010, 10, 712–723. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Schembri, M.A.; Dalsgaard, D.; Klemm, P. Capsule shields the function of short bacterial adhesins. J. Bacteriol. 2004, 186, 1249–1257. [Google Scholar] [CrossRef] [Green Version]
- Hollands, A.; Pence, M.A.; Timmer, A.M.; Osvath, S.R.; Turnbull, L.; Whitchurch, C.B.; Walker, M.J.; Nizet, V. Genetic switch to hypervirulence reduces colonization phenotypes of the globally disseminated group A streptococcus M1T1 clone. J. Infect. Dis. 2010, 202, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Absolom, D.R. The role of bacterial hydrophobicity in infection: Bacterial adhesion and phagocytic ingestion. Can. J. Microbiol. 1988, 34, 287–298. [Google Scholar] [CrossRef]
- Fletcher, M.; Loeb, G.I. Influence of substratum characteristics on the attachment of a marine pseudomonad to solid surfaces. Appl. Environ. Microbiol. 1979, 37, 67–72. [Google Scholar] [CrossRef] [Green Version]
- Kuusela, P. Fibronectin binds to Staphylococcus aureus. Nature 1978, 276, 718–720. [Google Scholar] [CrossRef]
- Heilmann, C. Adhesion mechanisms of staphylococci. Adv. Exp. Med. Biol. 2011, 715, 105–123. [Google Scholar] [CrossRef]
- Siggins, M.K.; Gill, S.K.; Langford, P.R.; Li, Y.; Ladhani, S.N.; Tregoning, J.S. PHiD-CV induces anti-Protein D antibodies but does not augment pulmonary clearance of nontypeable Haemophilus influenzae in mice. Vaccine 2015, 33, 4954–4961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, P.; Evans, D.J.; Evans, E.; Duguid, I.G.; Brown, M.R. Surface characteristics and adhesion of Escherichia coli and Staphylococcus epidermidis. J. Appl. Bacteriol. 1991, 71, 72–77. [Google Scholar]
- Gryllos, I.; Cywes, C.; Shearer, M.H.; Cary, M.; Kennedy, R.C.; Wessels, M.R. Regulation of capsule gene expression by group A Streptococcus during pharyngeal colonization and invasive infection. Mol. Microbiol. 2001, 42, 61–74. [Google Scholar] [CrossRef]
- Schmid-Schonbein, G.W. Microlymphatics and lymph flow. Physiol. Rev. 1990, 70, 987–1028. [Google Scholar] [CrossRef]
- Kuan, E.L.; Ivanov, S.; Bridenbaugh, E.A.; Victora, G.; Wang, W.; Childs, E.W.; Platt, A.M.; Jakubzick, C.V.; Mason, R.J.; Gashev, A.A.; et al. Collecting lymphatic vessel permeability facilitates adipose tissue inflammation and distribution of antigen to lymph node-homing adipose tissue dendritic cells. J. Immunol. 2015, 194, 5200–5210. [Google Scholar] [CrossRef] [Green Version]
- Margaris, K.N.; Black, R.A. Modelling the lymphatic system: Challenges and opportunities. J. R. Soc. Interface 2012, 9, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willard-Mack, C.L. Normal structure, function, and histology of lymph nodes. Toxicol. Pathol. 2006, 34, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Van den Broeck, W.; Derore, A.; Simoens, P. Anatomy and nomenclature of murine lymph nodes: Descriptive study and nomenclatory standardization in BALB/cAnNCrl mice. J. Immunol. Methods 2006, 312, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Nossal, G.J.; Abbot, A.; Mitchell, J. Antigens in immunity. XIV. Electron microscopic radioautographic studies of antigen capture in the lymph node medulla. J. Exp. Med. 1968, 127, 263–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foley, M.J.; Smith, M.R.; Wood, W.B., Jr. Studies on the pathogenicity of group A Streptococci. I. Its relation to surface phagocytosis. J. Exp. Med. 1959, 110, 603–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, M.E.; Drinker, C.K.; White, J.C. Lymph Pressures in Sterile Inflammation. J. Exp. Med. 1932, 56, 363–370. [Google Scholar] [CrossRef]
- Benoit, J.N.; Zawieja, D.C. Effects of f-Met-Leu-Phe-induced inflammation on intestinal lymph flow and lymphatic pump behavior. Am. J. Physiol. 1992, 262, G199–G202. [Google Scholar] [CrossRef] [PubMed]
- Fossum, S. The architecture of rat lymph nodes. IV. Distribution of ferritin and colloidal carbon in the draining lymph nodes after foot-pad injection. Scand. J. Immunol. 1980, 12, 433–441. [Google Scholar] [CrossRef] [PubMed]
- Rantakari, P.; Auvinen, K.; Jappinen, N.; Kapraali, M.; Valtonen, J.; Karikoski, M.; Gerke, H.; Iftakhar, E.K.I.; Keuschnigg, J.; Umemoto, E.; et al. The endothelial protein PLVAP in lymphatics controls the entry of lymphocytes and antigens into lymph nodes. Nat. Immunol. 2015, 16, 386–396. [Google Scholar] [CrossRef] [PubMed]
- Sixt, M.; Kanazawa, N.; Selg, M.; Samson, T.; Roos, G.; Reinhardt, D.P.; Pabst, R.; Lutz, M.B.; Sorokin, L. The conduit system transports soluble antigens from the afferent lymph to resident dendritic cells in the T cell area of the lymph node. Immunity 2005, 22, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siggins, M.K.; Thwaites, R.S.; Openshaw, P.J.M. Durability of Immunity to SARS-CoV-2 and Other Respiratory Viruses. Trends Microbiol. 2021, 29, 648–662. [Google Scholar] [CrossRef]
- Liao, S.; von der Weid, P.Y. Lymphatic system: An active pathway for immune protection. Semin. Cell Dev. Biol. 2015, 38, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.R.; Wood, W.B., Jr. Surface phagocy.ytosis; further evidence of its destructive action upon fully encapsulated pneumococci in the absence of type-specific antibody. J. Exp. Med. 1958, 107, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.O.; Wood, W.B., Jr. Cellular mechanisms of antibacterial defense in lymph nodes; pathogenesis of acute bacterial lymphadenitis. J. Exp. Med. 1949, 90, 567–576. [Google Scholar] [CrossRef]
- Sansonetti, P. Phagocytosis of bacterial pathogens: Implications in the host response. Semin. Immunol. 2001, 13, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Garcia, Z.; Lemaitre, F.; van Rooijen, N.; Albert, M.L.; Levy, Y.; Schwartz, O.; Bousso, P. Subcapsular sinus macrophages promote NK cell accumulation and activation in response to lymph-borne viral particles. Blood 2012, 120, 4744–4750. [Google Scholar] [CrossRef] [Green Version]
- Moseman, E.A.; Iannacone, M.; Bosurgi, L.; Tonti, E.; Chevrier, N.; Tumanov, A.; Fu, Y.X.; Hacohen, N.; von Andrian, U.H. B cell maintenance of subcapsular sinus macrophages protects against a fatal viral infection independent of adaptive immunity. Immunity 2012, 36, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Takeda, A.; Hollmen, M.; Dermadi, D.; Pan, J.; Brulois, K.F.; Kaukonen, R.; Lonnberg, T.; Bostrom, P.; Koskivuo, I.; Irjala, H.; et al. Single-Cell Survey of Human Lymphatics Unveils Marked Endothelial Cell Heterogeneity and Mechanisms of Homing for Neutrophils. Immunity 2019, 51, 561–572.e5. [Google Scholar] [CrossRef]
- Xiang, M.; Grosso, R.A.; Takeda, A.; Pan, J.; Bekkhus, T.; Brulois, K.; Dermadi, D.; Nordling, S.; Vanlandewijck, M.; Jalkanen, S.; et al. A Single-Cell Transcriptional Roadmap of the Mouse and Human Lymph Node Lymphatic Vasculature. Front. Cardiovasc. Med. 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Jalkanen, S.; Salmi, M. Lymphatic endothelial cells of the lymph node. Nat. Rev. Immunol. 2020, 20, 566–578. [Google Scholar] [CrossRef] [PubMed]
- Jafarnejad, M.; Woodruff, M.C.; Zawieja, D.C.; Carroll, M.C.; Moore, J.E., Jr. Modeling Lymph Flow and Fluid Exchange with Blood Vessels in Lymph Nodes. Lymphat. Res. Biol. 2015, 13, 234–247. [Google Scholar] [CrossRef] [Green Version]
- Widdicombe, J.G.; Hughes, R.; May, A.J. The efficiency of filtration by the popliteal lymph node of the rabbit. Br. J. Exp. Pathol. 1955, 36, 473–478. [Google Scholar]
- Bogoslowski, A.; Wijeyesinghe, S.; Lee, W.Y.; Chen, C.S.; Alanani, S.; Jenne, C.; Steeber, D.A.; Scheiermann, C.; Butcher, E.C.; Masopust, D.; et al. Neutrophils Recirculate through Lymph Nodes to Survey Tissues for Pathogens. J. Immunol. 2020, 204, 2552–2561. [Google Scholar] [CrossRef] [PubMed]
- Chtanova, T.; Schaeffer, M.; Han, S.J.; van Dooren, G.G.; Nollmann, M.; Herzmark, P.; Chan, S.W.; Satija, H.; Camfield, K.; Aaron, H.; et al. Dynamics of neutrophil migration in lymph nodes during infection. Immunity 2008, 29, 487–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampton, H.R.; Bailey, J.; Tomura, M.; Brink, R.; Chtanova, T. Microbe-dependent lymphatic migration of neutrophils modulates lymphocyte proliferation in lymph nodes. Nat. Commun. 2015, 6, 7139. [Google Scholar] [CrossRef]
- Rigby, D.A.; Ferguson, D.J.; Johnson, L.A.; Jackson, D.G. Neutrophils rapidly transit inflamed lymphatic vessel endothelium via integrin-dependent proteolysis and lipoxin-induced junctional retraction. J. Leukoc. Biol. 2015, 98, 897–912. [Google Scholar] [CrossRef]
- Phan, T.G.; Green, J.A.; Gray, E.E.; Xu, Y.; Cyster, J.G. Immune complex relay by subcapsular sinus macrophages and noncognate B cells drives antibody affinity maturation. Nat. Immunol. 2009, 10, 786–793. [Google Scholar] [CrossRef]
- Cyster, J.G. B cell follicles and antigen encounters of the third kind. Nat. Immunol. 2010, 11, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Moran, I.; Grootveld, A.K.; Nguyen, A.; Phan, T.G. Subcapsular Sinus Macrophages: The Seat of Innate and Adaptive Memory in Murine Lymph Nodes. Trends Immunol. 2018, 40, 35–48. [Google Scholar] [CrossRef]
- Carrasco, Y.R.; Batista, F.D. B cells acquire particulate antigen in a macrophage-rich area at the boundary between the follicle and the subcapsular sinus of the lymph node. Immunity 2007, 27, 160–171. [Google Scholar] [CrossRef] [Green Version]
- Farrell, H.E.; Davis-Poynter, N.; Bruce, K.; Lawler, C.; Dolken, L.; Mach, M.; Stevenson, P.G. Lymph Node Macrophages Restrict Murine Cytomegalovirus Dissemination. J. Virol. 2015, 89, 7147–7158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.C.; Olson, J.; Louie, A.; Crocker, P.R.; Varki, A.; Nizet, V. Role of macrophage sialoadhesin in host defense against the sialylated pathogen group B Streptococcus. J. Mol. Med. (Berl.) 2014, 92, 951–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heikema, A.P.; Koning, R.I.; Duarte dos Santos Rico, S.; Rempel, H.; Jacobs, B.C.; Endtz, H.P.; van Wamel, W.J.; Samsom, J.N. Enhanced, sialoadhesin-dependent uptake of Guillain-Barre syndrome-associated Campylobacter jejuni strains by human macrophages. Infect. Immun. 2013, 81, 2095–2103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, C.; Virji, M.; Crocker, P.R. Recognition of sialylated meningococcal lipopolysaccharide by siglecs expressed on myeloid cells leads to enhanced bacterial uptake. Mol. Microbiol. 2003, 49, 1213–1225. [Google Scholar] [CrossRef]
- Phan, T.G.; Grigorova, I.; Okada, T.; Cyster, J.G. Subcapsular encounter and complement-dependent transport of immune complexes by lymph node B cells. Nat. Immunol. 2007, 8, 992–1000. [Google Scholar] [CrossRef]
- Szakal, A.K.; Holmes, K.L.; Tew, J.G. Transport of immune complexes from the subcapsular sinus to lymph node follicles on the surface of nonphagocytic cells, including cells with dendritic morphology. J. Immunol. 1983, 131, 1714–1727. [Google Scholar] [PubMed]
- Gonzalez, S.F.; Lukacs-Kornek, V.; Kuligowski, M.P.; Pitcher, L.A.; Degn, S.E.; Kim, Y.A.; Cloninger, M.J.; Martinez-Pomares, L.; Gordon, S.; Turley, S.J.; et al. Capture of influenza by medullary dendritic cells via SIGN-R1 is essential for humoral immunity in draining lymph nodes. Nat. Immunol. 2010, 11, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Lynskey, N.N.; Reglinski, M.; Calay, D.; Siggins, M.K.; Mason, J.C.; Botto, M.; Sriskandan, S. Multi-functional mechanisms of immune evasion by the streptococcal complement inhibitor C5a peptidase. PLoS Pathog. 2017, 13, e1006493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rooijakkers, S.H.; Ruyken, M.; Roos, A.; Daha, M.R.; Presanis, J.S.; Sim, R.B.; van Wamel, W.J.; van Kessel, K.P.; van Strijp, J.A. Immune evasion by a staphylococcal complement inhibitor that acts on C3 convertases. Nat. Immunol. 2005, 6, 920–927. [Google Scholar] [CrossRef] [PubMed]
- Lambris, J.D.; Ricklin, D.; Geisbrecht, B.V. Complement evasion by human pathogens. Nat. Rev. Microbiol. 2008, 6, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; Groot, P.C.; Nolte, M.A.; van Vliet, S.J.; Gangaram-Panday, S.T.; van Duijnhoven, G.C.; Kraal, G.; van Oosterhout, A.J.; van Kooyk, Y. Marginal zone macrophages express a murine homologue of DC-SIGN that captures blood-borne antigens in vivo. Blood 2002, 100, 2908–2916. [Google Scholar] [CrossRef] [PubMed]
- Martens, J.H.; Kzhyshkowska, J.; Falkowski-Hansen, M.; Schledzewski, K.; Gratchev, A.; Mansmann, U.; Schmuttermaier, C.; Dippel, E.; Koenen, W.; Riedel, F.; et al. Differential expression of a gene signature for scavenger/lectin receptors by endothelial cells and macrophages in human lymph node sinuses, the primary sites of regional metastasis. J. Pathol. 2006, 208, 574–589. [Google Scholar] [CrossRef] [PubMed]
- Lanoue, A.; Clatworthy, M.R.; Smith, P.; Green, S.; Townsend, M.J.; Jolin, H.E.; Smith, K.G.; Fallon, P.G.; McKenzie, A.N. SIGN-R1 contributes to protection against lethal pneumococcal infection in mice. J. Exp. Med. 2004, 200, 1383–1393. [Google Scholar] [CrossRef]
- Kang, Y.S.; Kim, J.Y.; Bruening, S.A.; Pack, M.; Charalambous, A.; Pritsker, A.; Moran, T.M.; Loeffler, J.M.; Steinman, R.M.; Park, C.G. The C-type lectin SIGN-R1 mediates uptake of the capsular polysaccharide of Streptococcus pneumoniae in the marginal zone of mouse spleen. Proc. Natl. Acad. Sci. USA 2004, 101, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Van der Laan, L.J.; Döpp, E.A.; Haworth, R.; Pikkarainen, T.; Kangas, M.; Elomaa, O.; Dijkstra, C.D.; Gordon, S.; Tryggvason, K.; Kraal, G. Regulation and functional involvement of macrophage scavenger receptor MARCO in clearance of bacteria in vivo. J. Immunol. 1999, 162, 939–947. [Google Scholar] [PubMed]
- Doyle, S.E.; O’Connell, R.M.; Miranda, G.A.; Vaidya, S.A.; Chow, E.K.; Liu, P.T.; Suzuki, S.; Suzuki, N.; Modlin, R.L.; Yeh, W.C.; et al. Toll-like receptors induce a phagocytic gene program through p38. J. Exp. Med. 2004, 199, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Loof, T.G.; Rohde, M.; Chhatwal, G.S.; Jung, S.; Medina, E. The contribution of dendritic cells to host defenses against Streptococcus pyogenes. J. Infect. Dis. 2007, 196, 1794–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.M. The pathogenesis of anthrax following the administration of spores by the respiratory route. J. Pathol. Bacteriol. 1957, 73, 485–494. [Google Scholar] [CrossRef]
- Angevine, D.M. The Fate of Avirulent Hemolytic Streptococci Injected into the Skin of Normal and Sensitized Rabbits: Local Fixation of Bacteria. J. Exp. Med. 1934, 60, 269–285. [Google Scholar] [CrossRef]
- Wilson, A.T.; Wiley, G.G.; Bruno, P. Fate of non-virulent group A streptococci phagocytized by human and mouse neutrophils. J. Exp. Med. 1957, 106, 777–786. [Google Scholar] [CrossRef]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, A.T. The egestion of phagocytized particles by leukocytes. J. Exp. Med. 1953, 98, 305–310. [Google Scholar] [CrossRef] [Green Version]
- St Sauver, J.L.; Weaver, A.L.; Orvidas, L.J.; Jacobson, R.M.; Jacobsen, S.J. Population-based prevalence of repeated group A beta-hemolytic streptococcal pharyngitis episodes. Mayo Clin. Proc. 2006, 81, 1172–1176. [Google Scholar] [CrossRef]
- Davies, F.J.; Olme, C.; Lynskey, N.N.; Turner, C.E.; Sriskandan, S. Streptococcal superantigen-induced expansion of human tonsil T cells leads to altered T follicular helper cell phenotype, B cell death and reduced immunoglobulin release. Clin. Exp. Immunol. 2019, 197, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Dan, J.M.; Havenar-Daughton, C.; Kendric, K.; Al-Kolla, R.; Kaushik, K.; Rosales, S.L.; Anderson, E.L.; LaRock, C.N.; Vijayanand, P.; Seumois, G.; et al. Recurrent group A Streptococcus tonsillitis is an immunosusceptibility disease involving antibody deficiency and aberrant TFH cells. Sci. Transl. Med. 2019, 11, aau3776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stollerman, G.H.; Dale, J.B. The importance of the group a streptococcus capsule in the pathogenesis of human infections: A historical perspective. Clin. Infect. Dis. 2008, 46, 1038–1045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [Green Version]
- Siggins, M.K.; Cunningham, A.F.; Marshall, J.L.; Chamberlain, J.L.; Henderson, I.R.; MacLennan, C.A. Absent bactericidal activity of mouse serum against invasive African nontyphoidal Salmonella results from impaired complement function but not a lack of antibody. J. Immunol. 2011, 186, 2365–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, H.S.; Fitting, C.; Hoff, E.; Adib-Conquy, M.; Beasley-Topliffe, L.; Tesini, B.; Liang, X.; Valentine, C.; Hellman, J.; Hayden, D.; et al. Resilience to bacterial infection: Difference between species could be due to proteins in serum. J. Infect. Dis. 2010, 201, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawash, M.B.F.; Sanz-Remon, J.; Grenier, J.C.; Kohn, J.; Yotova, V.; Johnson, Z.; Lanford, R.E.; Brinkworth, J.F.; Barreiro, L.B. Primate innate immune responses to bacterial and viral pathogens reveals an evolutionary trade-off between strength and specificity. Proc. Natl. Acad. Sci. USA 2021, 118, e2015855118. [Google Scholar] [CrossRef] [PubMed]





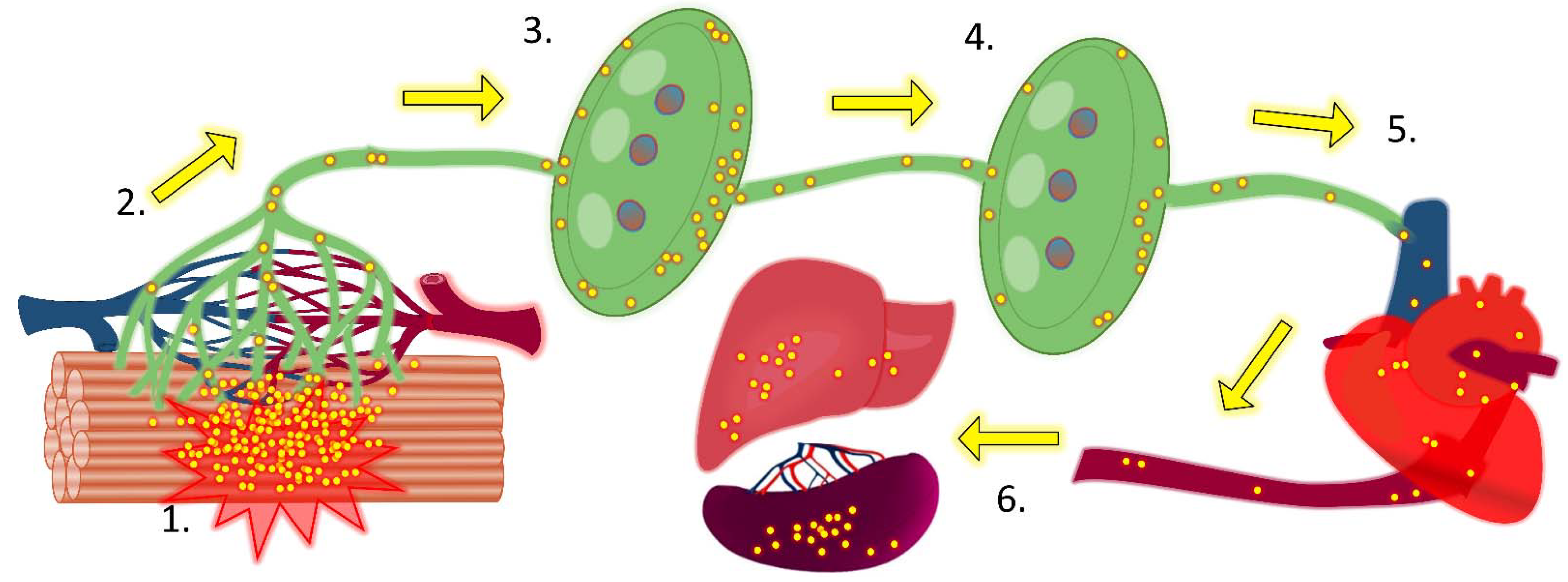{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Infection Route | Model | Mechanisms of Transit to Local Draining Lymph Nodes | Recovery of Bacteria from Other Lymphatic Sites | References |
|---|---|---|---|---|---|
| Bacillus anthracis (spores) | Aerosol, ID, IP | Rhesus macaque | Unknown | Efferent lymph | Lincoln, R.E. 1965; [24] |
| Escherichia coli | OG | Mouse | Unknown | None investigated | Balmer, M. 2014; [29] |
| Francisella tularensis | IN | Mouse | Extracellular and intracellular within dendritic cells | None investigated | Bar-Haim, E. 2008; [30] |
| Klebsiella pneumoniae | IM | Mouse | Unknown | Distant draining lymph nodes | Siggins, M.K. 2019; [23] |
| Listeria monocytogenes | IG, FB Footpad | Mouse | Unknown, influence of bacterial E-cadherin ligand Unknown | None investigated | Bou Ghanem, E. 2012; [31] St. John, A.L. 2009; [32] |
| Mycobacterium tuberculosis | Aerosol | Mouse Cynomolgus and Rhesus macaque | Unknown | None investigated | Chackerian, A.A. 2002; [33] Ganchua, S.K. 2018; [34] |
| Pseudomonas aeruginosa | IM Footpad | Mouse | Unknown | Distant draining lymph nodes None investigated | Siggins, M.K. 2019; [23] Kastenmüller, W. 2012; [35] |
| Salmonella Abortusovis | SC (oral) | Sheep | Free extracellular or associated with monocytes and granulocytes | Afferent lymph | Bonneau, M. 2006; [16] |
| Salmonella Typhimurium | OG Footpad | Sheep Mouse | Free extracellular or associated with dendritic cells Unknown | Afferent lymph None investigated | Bravo-Blas, A. 2019; [25] St. John, A.L. 2009; [32] |
| Staphylococcus aureus | IM | Mouse | Unknown | None investigated | Bogoslowski, A. 2018; [36] |
| Streptococcus agalactiae | IM | Mouse | Unknown, influence of capsule | Distant draining lymph nodes | Siggins, M.K. 2021; [unpublished] |
| Streptococcus pyogenes | IM | Mouse | Free extracellular, influence of HA capsule and LYVE-1 interaction | Distant draining lymph nodes and efferent lymph None investigated | Siggins, M.K. 2020; [21] Lynskey, N.N. 2015; [39] |
| Yersinia pestis | ID Footpad | Mouse | Unknown but not dependent on neutrophils or dendritic cells Free extracellular independent of phagocytes Free extracellular and in mononuclear phagocytes | None investigated Afferent lymphatic Distant draining lymph node and efferent lymphatic | Shannon, J.G. 2013; [28] Gonzalez, R.J. 2015; [26] St. John, A.L. 2014; [15] |
| Bacteria | Infection Route | Model | Sites of Recovery beyond Local Draining Lymph Nodes | Predominant Mechanisms of Lymphatic Transit Reported | References |
|---|---|---|---|---|---|
| Bacillus anthracis (spores) | Aerosol, IP, and SC Aerosol and SC | Rhesus macaque Rabbit | Efferent lymph | Unknown | Lincoln, R.E. 1965; [24] Hughes, R. 1956; [40] |
| Klebsiella pneumoniae | IM | Mouse | Distant draining lymph nodes | Unknown | Siggins, M.K. 2019; [23] |
| Pseudomonas aeruginosa | IM | Mouse | Distant draining lymph nodes | Unknown | Siggins, M.K. 2019; [23] |
| Salmonella Dublin | Oral | Cow | Efferent lymph | Free extracellular | Pullinger, G.D. 2007, [38] |
| Streptococcus pyogenes | IM | Mouse | Distant draining lymph nodes and efferent lymphatics | Free extracellular and extracellular association with leukocytes, contribution of HA capsule and interaction with LYVE-1 | Siggins, M.K. 2020; [21] |
| Streptococcus pneumoniae | IN and IT | Rabbit | Efferent lymph | Free extracellular | Schulz, R.Z. 1938; [37] |
| Yersinia pestis | Footpad | Mouse | Distant draining lymph nodes and efferent lymphatic | In mononuclear phagocytes | St. John, A.L. 2014; [15] |
| Term | Site | Description |
|---|---|---|
| Bacteraemia | Blood | Presence of viable bacteria in the circulating blood |
| Cellulitis | Subcutaneous tissue | Infection of the lower dermis and subcutaneous tissue causing spreading inflammation |
| Cryptic infection | - | Bloodstream or deeper tissue infections that lack a known peripheral focus |
| Erysipelas | Epidermis | Infection of the superficial epidermis |
| Fasciitis (necrotizing) | Fascia | Infection of superficial or deep fascia in association with tissue destruction. |
| Impetigo | Keratin layer of epidermis | Infection of superficial keratin layer of skin |
| Lymphadenitis/lymphadenopathy | Lymph nodes | Inflammation/enlargement of lymph nodes due to presence of infection within vessels or tissue drained by the lymph node, or the node itself |
| Lymphangitis | Lymphatic vessels | Inflammation of lymphatic vessels resulting from infection of the vessels themselves, or nearby tissue |
| Myositis/myonecrosis | Muscle | Infection of muscle leading to inflammation/necrosis |
| Puerperal sepsis | Uterus | Infection of the genital tract, particularly the uterus, occurring soon after labour |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siggins, M.K.; Sriskandan, S. Bacterial Lymphatic Metastasis in Infection and Immunity. Cells 2022, 11, 33. https://doi.org/10.3390/cells11010033
Siggins MK, Sriskandan S. Bacterial Lymphatic Metastasis in Infection and Immunity. Cells. 2022; 11(1):33. https://doi.org/10.3390/cells11010033
Chicago/Turabian StyleSiggins, Matthew K., and Shiranee Sriskandan. 2022. "Bacterial Lymphatic Metastasis in Infection and Immunity" Cells 11, no. 1: 33. https://doi.org/10.3390/cells11010033
APA StyleSiggins, M. K., & Sriskandan, S. (2022). Bacterial Lymphatic Metastasis in Infection and Immunity. Cells, 11(1), 33. https://doi.org/10.3390/cells11010033