
Activation and Migration of Human Skeletal Muscle Stem Cells In Vitro Differently Rely on Calcium Signals


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Immunofluorescence and Activation Assay
2.3. Calcium Measurements
2.4. Timelapse and Movement Tracking
2.5. Statistics
3. Results
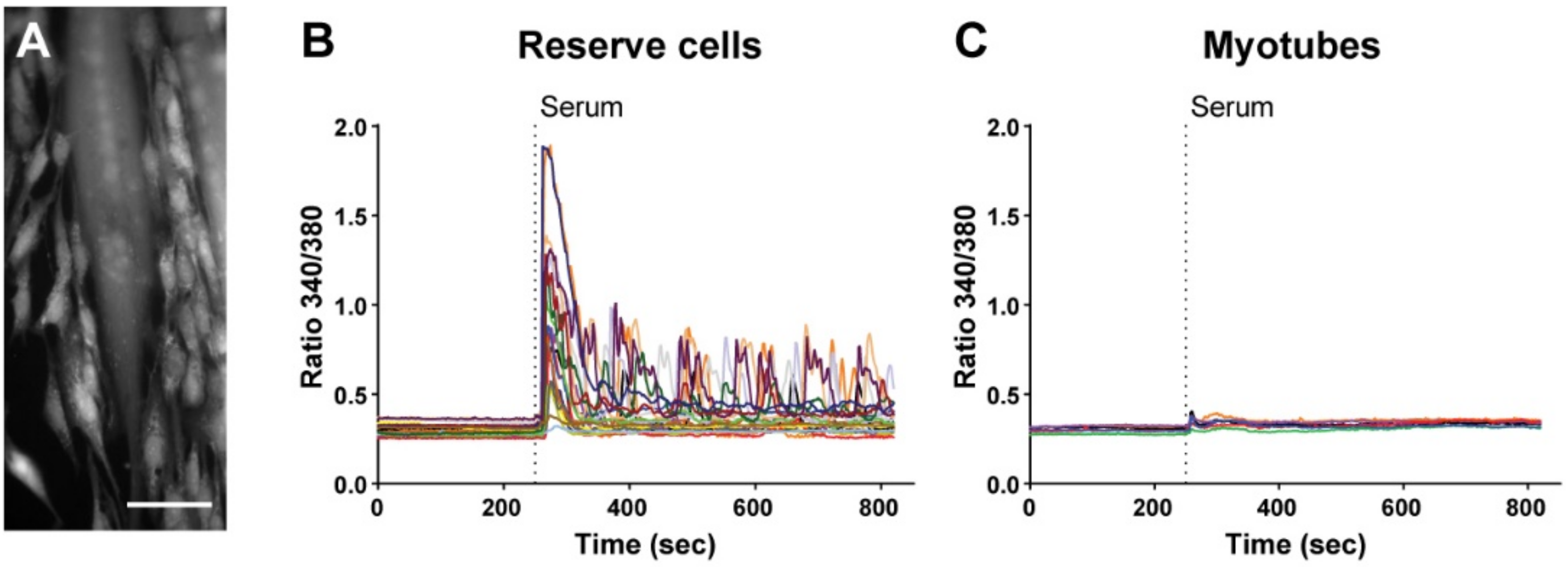
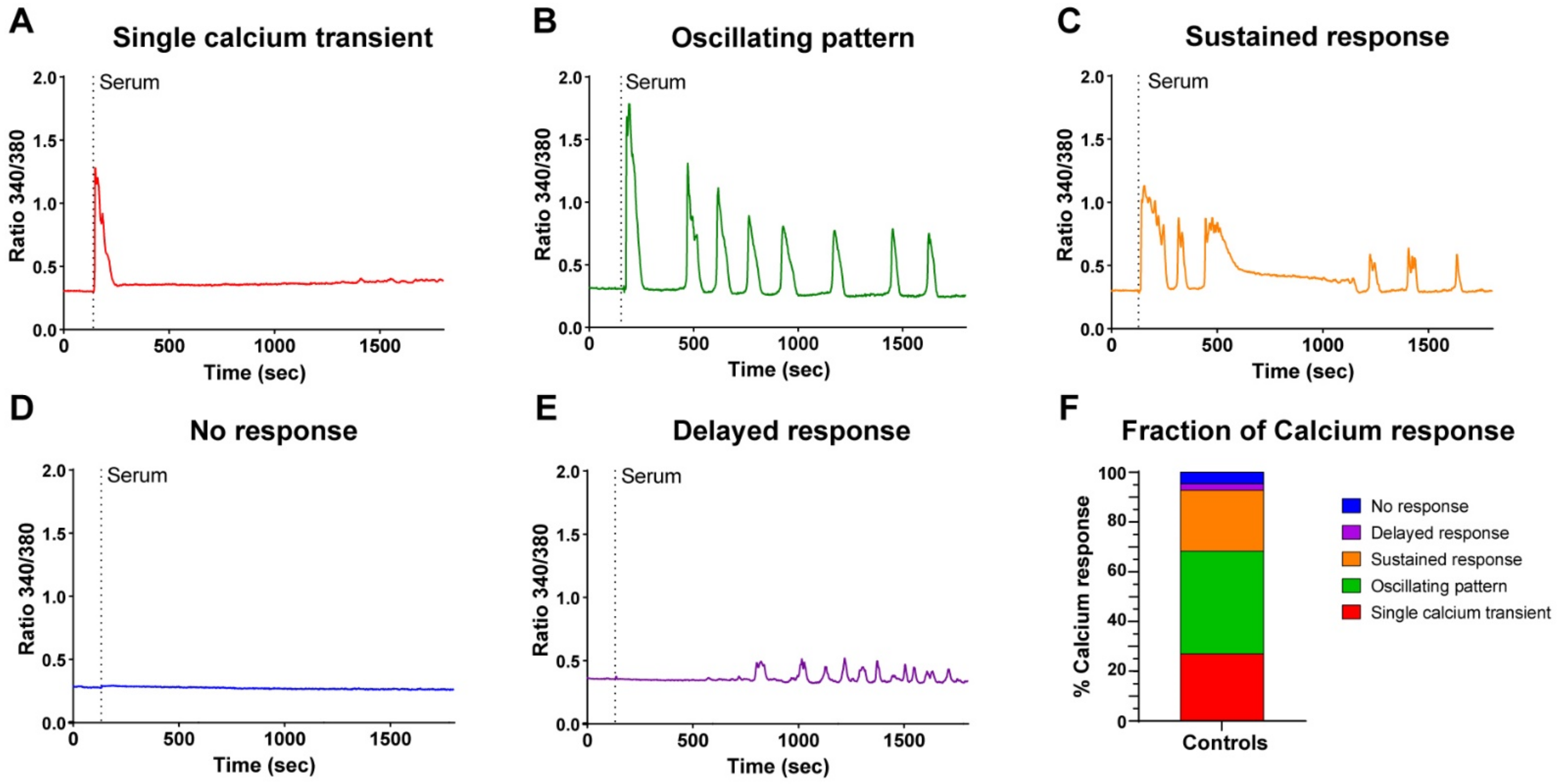
3.1. Reserve Cells Display Heterogeneous Calcium Responses upon Serum Stimulation
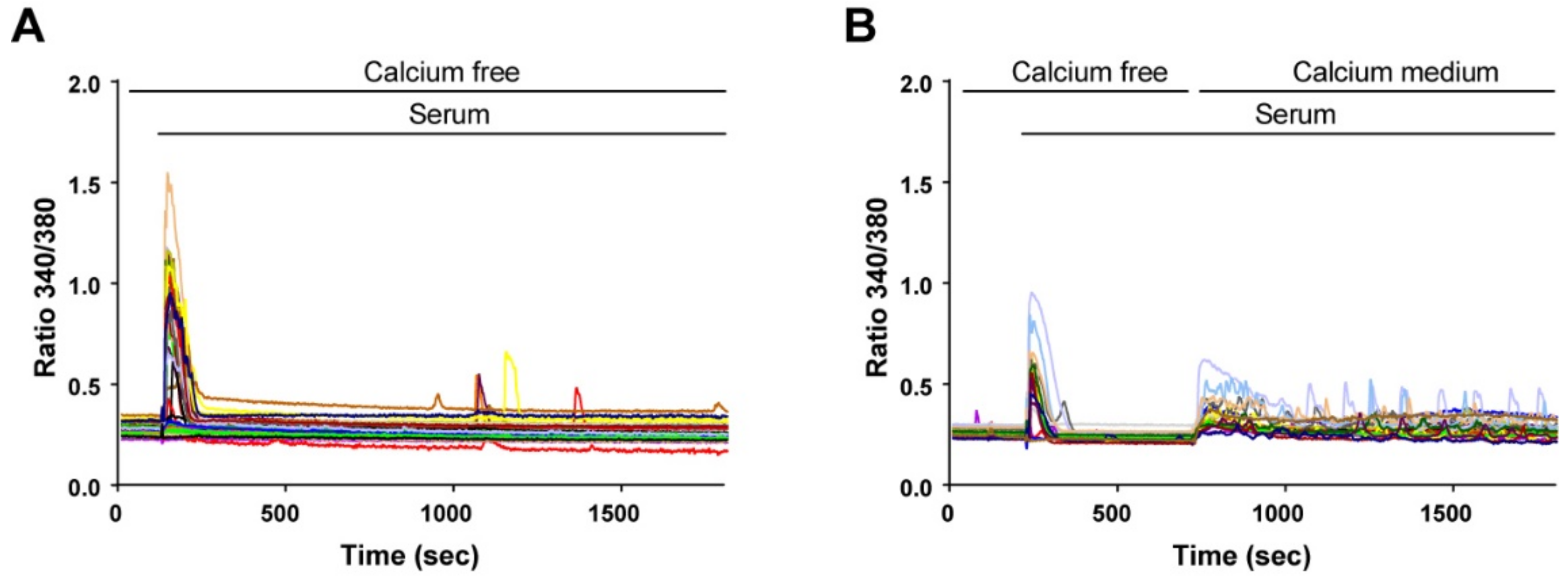
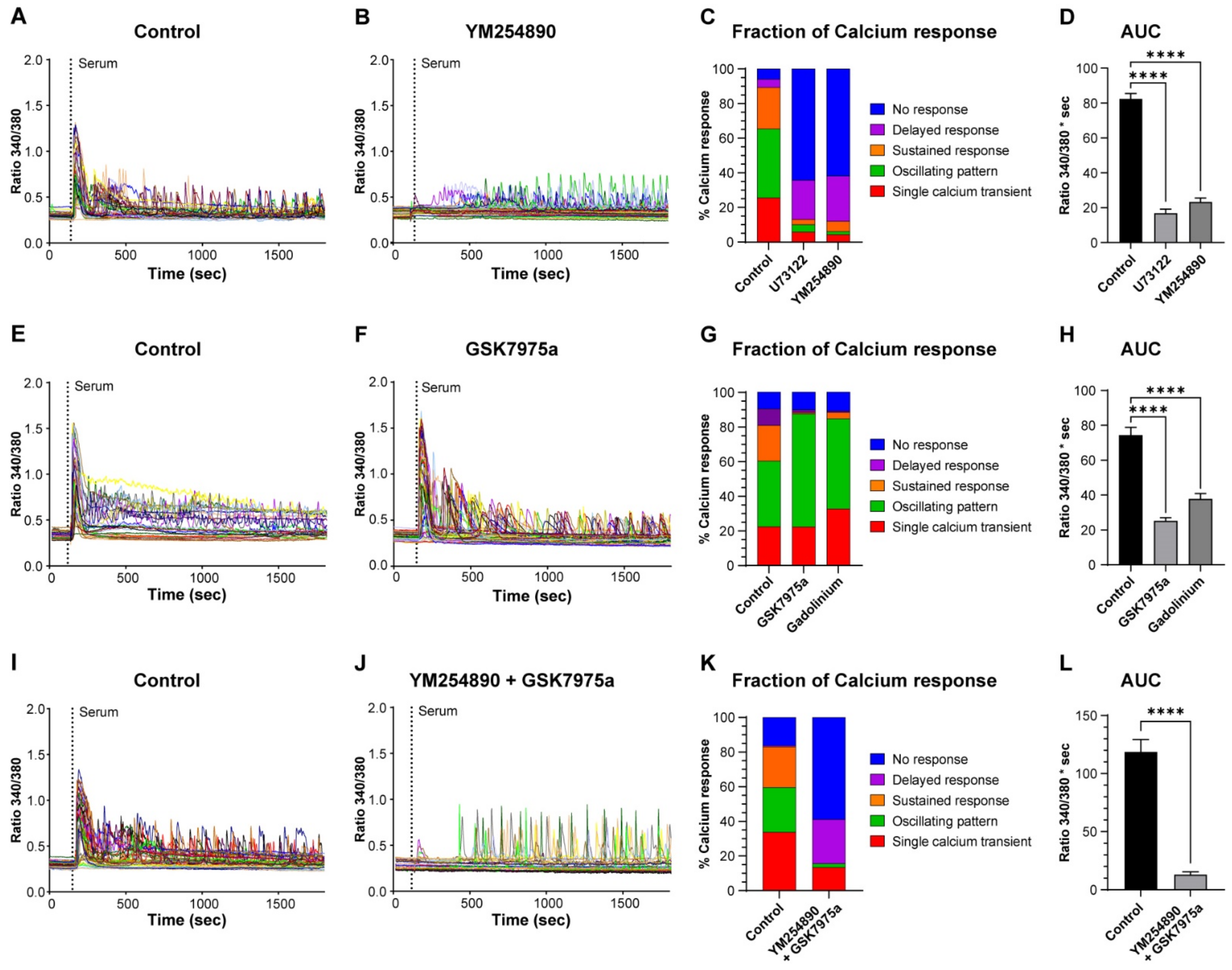
3.2. Calcium Release and Calcium Influx Are Used by Reserve Cells to Generate Calcium Signals
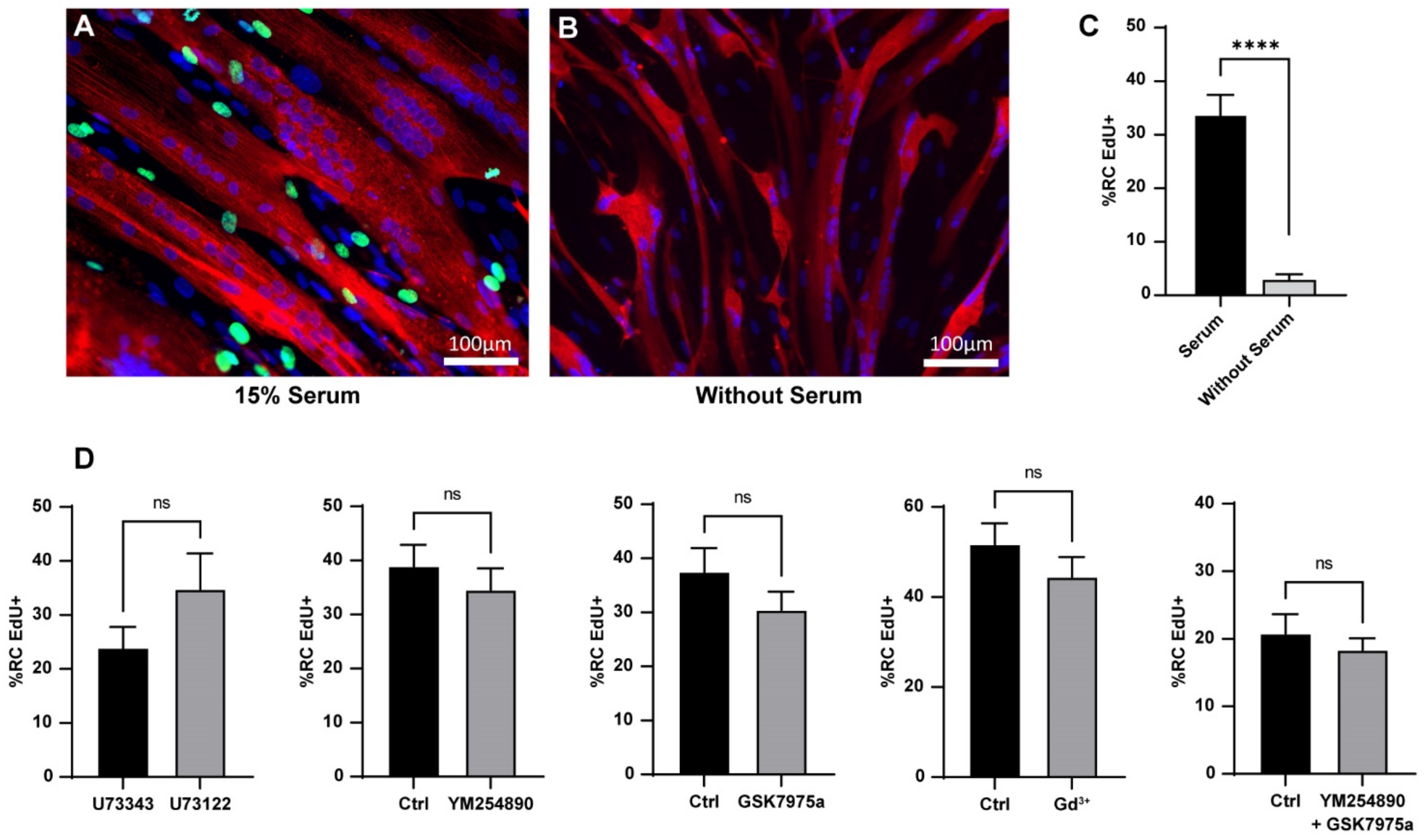
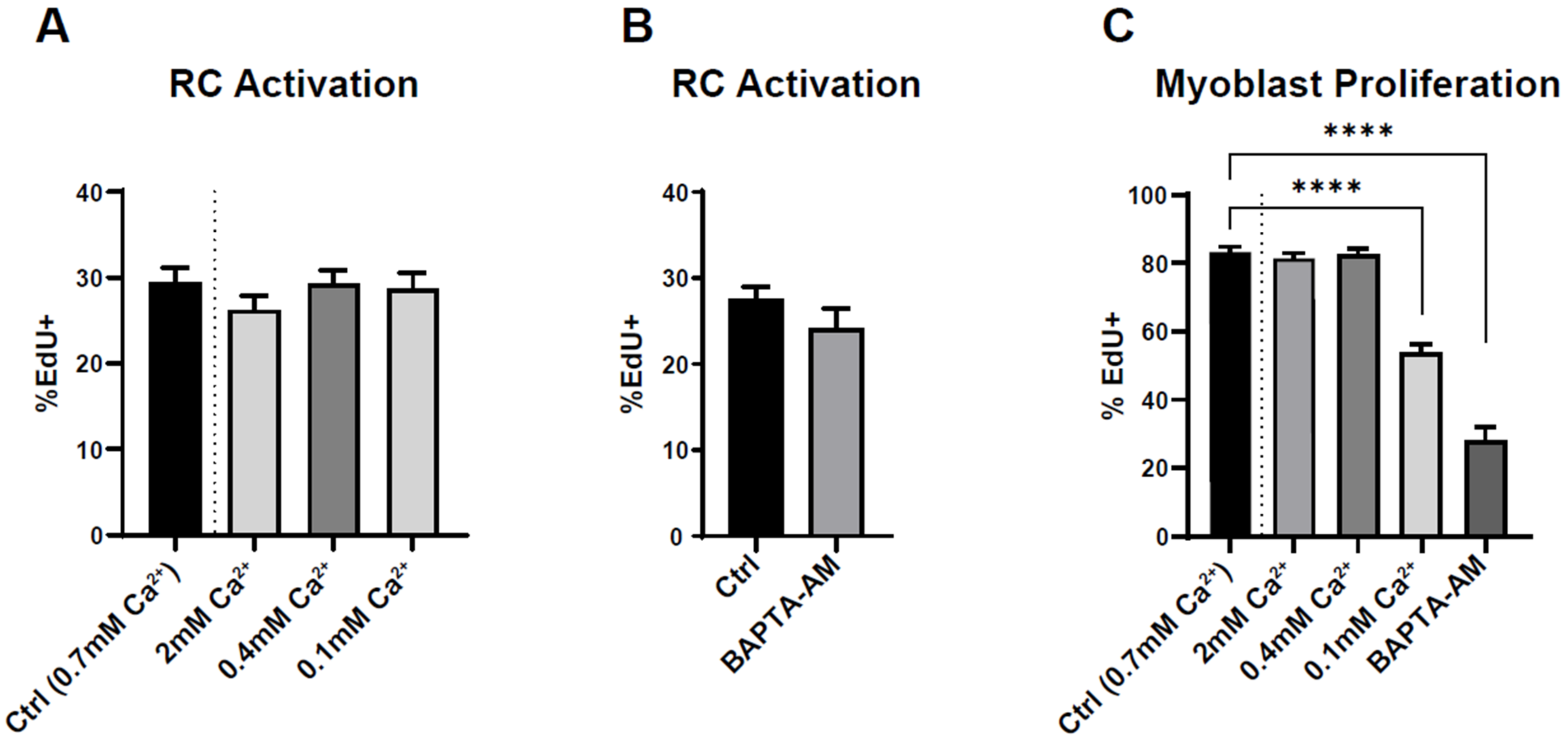
3.3. Reserve Cell Activation Is Calcium Independent
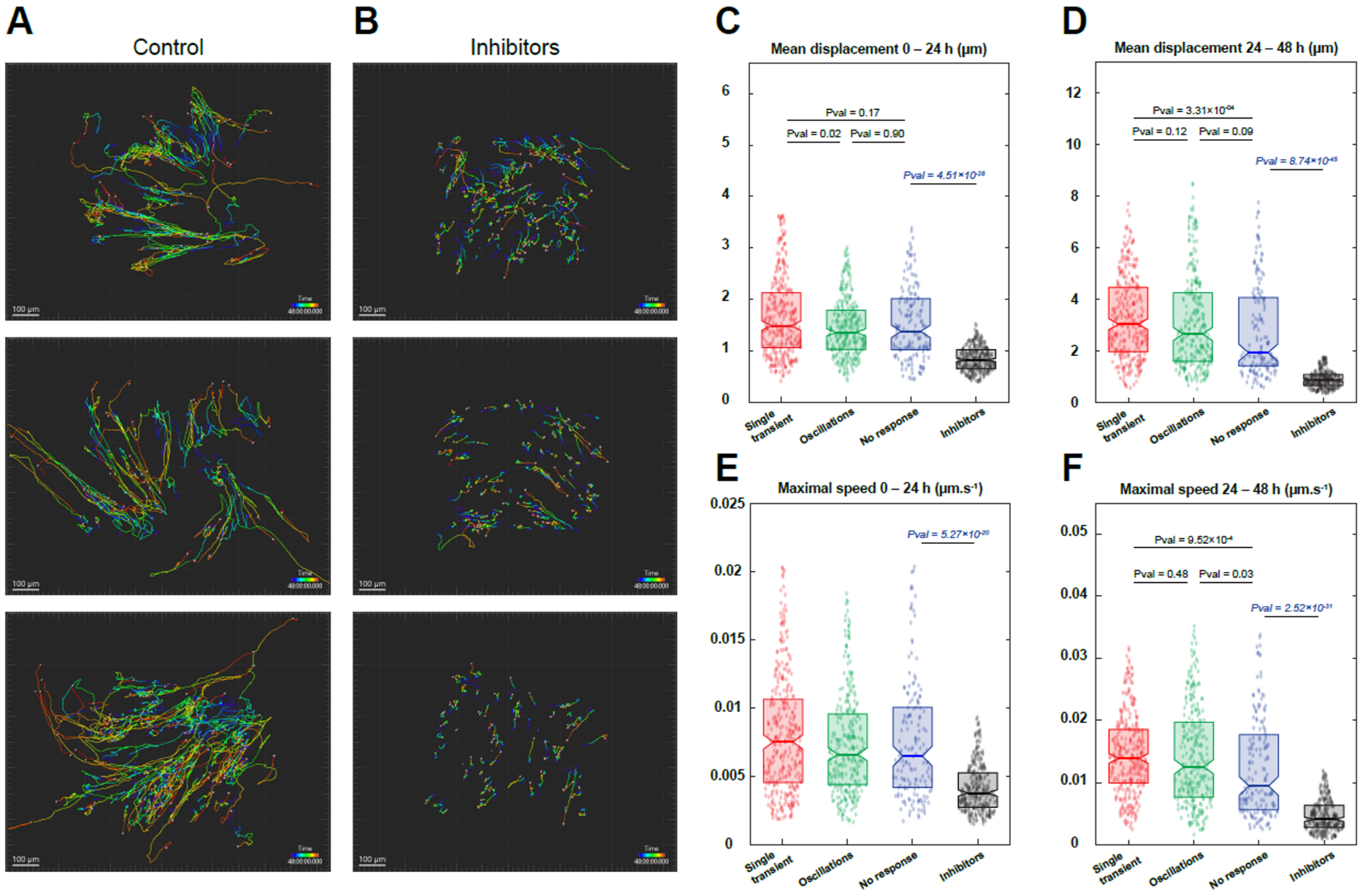
3.4. Reserve Cell Migration Depends on Calcium Signals
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, C.A.; Olsen, I.; Zammit, P.S.; Heslop, L.; Petrie, A.; Partridge, T.A.; Morgan, J.E. Stem Cell Function, Self-Renewal, and Behavioral Heterogeneity of Cells from the Adult Muscle Satellite Cell Niche. Cell 2005, 122, 289–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baghdadi, M.B.; Tajbakhsh, S. Regulation and Phylogeny of Skeletal Muscle Regeneration. Dev. Biol. 2018, 433, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Wang, H.; Hu, P. Stem Cell Activation in Skeletal Muscle Regeneration. Cell. Mol. Life Sci. 2015, 72, 1663–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumman, M.; Dhawan, J.; Kassem, M. Concise Review: Quiescence in Adult Stem Cells: Biological Significance and Relevance to Tissue Regeneration: ASC Quiescence: Role and Relevance to Tissue Regeneration. Stem Cells 2015, 33, 2903–2912. [Google Scholar] [CrossRef]
- Machado, L.; Esteves de Lima, J.; Fabre, O.; Proux, C.; Legendre, R.; Szegedi, A.; Varet, H.; Ingerslev, L.R.; Barrès, R.; Relaix, F.; et al. In Situ Fixation Redefines Quiescence and Early Activation of Skeletal Muscle Stem Cells. Cell Rep. 2017, 21, 1982–1993. [Google Scholar] [CrossRef] [Green Version]
- Machado, L.; Geara, P.; Camps, J.; Dos Santos, M.; Teixeira-Clerc, F.; Van Herck, J.; Varet, H.; Legendre, R.; Pawlotsky, J.-M.; Sampaolesi, M.; et al. Tissue Damage Induces a Conserved Stress Response That Initiates Quiescent Muscle Stem Cell Activation. Cell Stem Cell 2021, 28, 1125–1135. [Google Scholar] [CrossRef]
- Van den Brink, S.C.; Sage, F.; Vértesy, Á.; Spanjaard, B.; Peterson-Maduro, J.; Baron, C.S.; Robin, C.; Van Oudenaarden, A. Single-Cell Sequencing Reveals Dissociation-Induced Gene Expression in Tissue Subpopulations. Nat. Methods 2017, 14, 935–936. [Google Scholar] [CrossRef]
- Baroffio, A.; Hamann, M.; Bernheim, L.; Bochaton-Piallat, M.-L.; Gabbiani, G.; Bader, C.R. Identification of Self-Renewing Myoblasts in the Progeny of Single Human Muscle Satellite Cells. Differentiation 1996, 60, 47–57. [Google Scholar] [CrossRef]
- Laumonier, T.; Koenig, S.; Saüc, S.; Frieden, M. Isolation of Human Myoblasts, Assessment of Myogenic Differentiation, and Store-Operated Calcium Entry Measurement. J. Vis. Exp. 2017, 55918. [Google Scholar] [CrossRef]
- Perroud, J.; Bernheim, L.; Frieden, M.; Koenig, S. Distinct Roles of NFATc1 and NFATc4 in Human Primary Myoblast Differentiation and in the Maintenance of Reserve Cells. J. Cell Sci. 2017, 130, 3083–3093. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Yoshida, S.; Koishi, K.; Masuda, K.; Nabeshima, Y. Cell Heterogeneity upon Myogenic Differentiation: Down-Regulation of MyoD and Myf-5 Generates ‘Reserve Cells’. J. Cell Sci. 1998, 111, 769–779. [Google Scholar] [CrossRef]
- Zammit, P.S.; Relaix, F.; Nagata, Y.; Ruiz, A.P.; Collins, C.A.; Partridge, T.A.; Beauchamp, J.R. Pax7 and Myogenic Progression in Skeletal Muscle Satellite Cells. J. Cell Sci. 2006, 119, 1824–1832. [Google Scholar] [CrossRef] [Green Version]
- Arnaudeau, S.; Holzer, N.; König, S.; Bader, C.R.; Bernheim, L. Calcium Sources Used by Post-Natal Human Myoblasts during Initial Differentiation. J. Cell. Physiol. 2006, 208, 435–445. [Google Scholar] [CrossRef]
- Darbellay, B.; Arnaudeau, S.; König, S.; Jousset, H.; Bader, C.; Demaurex, N.; Bernheim, L. STIM1- and Orai1-Dependent Store-Operated Calcium Entry Regulates Human Myoblast Differentiation. J. Biol. Chem. 2009, 284, 5370–5380. [Google Scholar] [CrossRef] [Green Version]
- Antigny, F.; Konig, S.; Bernheim, L.; Frieden, M. Inositol 1,4,5 Trisphosphate Receptor 1 Is a Key Player of Human Myoblast Differentiation. Cell Calcium 2014, 56, 513–521. [Google Scholar] [CrossRef]
- Tu, M.K.; Borodinsky, L.N. Spontaneous Calcium Transients Manifest in the Regenerating Muscle and Are Necessary for Skeletal Muscle Replenishment. Cell Calcium 2014, 56, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Stiber, J.; Hawkins, A.; Zhang, Z.-S.; Wang, S.; Burch, J.; Graham, V.; Ward, C.C.; Seth, M.; Finch, E.; Malouf, N.; et al. STIM1 Signalling Controls Store-Operated Calcium Entry Required for Development and Contractile Function in Skeletal Muscle. Nat. Cell Biol. 2008, 10, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Darbellay, B.; Arnaudeau, S.; Ceroni, D.; Bader, C.R.; Konig, S.; Bernheim, L. Human Muscle Economy Myoblast Differentiation and Excitation-Contraction Coupling Use the Same Molecular Partners, STIM1 and STIM2. J. Biol. Chem. 2010, 285, 22437–22447. [Google Scholar] [CrossRef] [Green Version]
- Prakriya, M.; Lewis, R.S. Store-Operated Calcium Channels. Physiol. Rev. 2015, 95, 54. [Google Scholar] [CrossRef] [Green Version]
- Hara, M.; Tabata, K.; Suzuki, T.; Do, M.-K.Q.; Mizunoya, W.; Nakamura, M.; Nishimura, S.; Tabata, S.; Ikeuchi, Y.; Sunagawa, K.; et al. Calcium Influx through a Possible Coupling of Cation Channels Impacts Skeletal Muscle Satellite Cell Activation in Response to Mechanical Stretch. Am. J. Physiol.-Cell Physiol. 2012, 302, C1741–C1750. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Schneider, M.F. FGF2 Activates TRPC and Ca2+ Signaling Leading to Satellite Cell Activation. Front. Physiol. 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Brunetti, J.; Koenig, S.; Monnier, A.; Frieden, M. Nanopattern Surface Improves Cultured Human Myotube Maturation. Skelet. Muscle 2021, 11, 12. [Google Scholar] [CrossRef]
- Yoast, R.E.; Emrich, S.M.; Zhang, X.; Xin, P.; Johnson, M.T.; Fike, A.J.; Walter, V.; Hempel, N.; Yule, D.I.; Sneyd, J.; et al. The Native ORAI Channel Trio Underlies the Diversity of Ca2+ Signaling Events. Nat. Commun. 2020, 11, 2444. [Google Scholar] [CrossRef]
- Schultz, E.; Jaryszak, D.L.; Valliere, C.R. Response of Satellite Cells to Focal Skeletal Muscle Injury. Muscle Nerve 1985, 8, 217–222. [Google Scholar] [CrossRef]
- Phillips, G.D.; Hoffman, J.R.; Knighton, D.R. Migration of Myogenic Cells in the Rat Extensor Digitorum Longus Muscle Studied with a Split Autograft Model. Cell Tissue Res. 1990, 262, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Laumonier, T.; Bermont, F.; Hoffmeyer, P.; Kindler, V.; Menetrey, J. Human Myogenic Reserve Cells Are Quiescent Stem Cells That Contribute to Muscle Regeneration after Intramuscular Transplantation in Immunodeficient Mice. Sci. Rep. 2017, 7, 3462. [Google Scholar] [CrossRef] [Green Version]
- Dupont, G.; Combettes, L.; Bird, G.S.; Putney, J.W. Calcium Oscillations. Cold Spring Harb. Perspect. Biol. 2011, 3, a004226. [Google Scholar] [CrossRef] [Green Version]
- Pecze, L.; Schwaller, B. Characterization and Modeling of Ca2+ Oscillations in Mouse Primary Mesothelial Cells. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2015, 1853, 632–645. [Google Scholar] [CrossRef] [Green Version]
- Derler, I.; Schindl, R.; Fritsch, R.; Heftberger, P.; Riedl, M.C.; Begg, M.; House, D.; Romanin, C. The Action of Selective CRAC Channel Blockers Is Affected by the Orai Pore Geometry. Cell Calcium 2013, 53, 139–151. [Google Scholar] [CrossRef]
- Patel, S.; Marchant, J.S.; Brailoiu, E. Two-Pore Channels: Regulation by NAADP and Customized Roles in Triggering Calcium Signals. Cell Calcium 2010, 47, 480–490. [Google Scholar] [CrossRef] [Green Version]
- Galione, A. NAADP Receptors. Cold Spring Harb. Perspect. Biol. 2019, 11, a035071. [Google Scholar] [CrossRef] [PubMed]
- Panci, G.; Chazaud, B. Inflammation during Post-Injury Skeletal Muscle Regeneration. Semin. Cell Dev. Biol. 2021, 119, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Baryshnikov, S.G.; Pulina, M.V.; Zulian, A.; Linde, C.I.; Golovina, V.A. Orai1, a Critical Component of Store-Operated Ca 2+ Entry, Is Functionally Associated with Na +/Ca 2+ Exchanger and Plasma Membrane Ca 2+ Pump in Proliferating Human Arterial Myocytes. Am. J. Physiol.-Cell Physiol. 2009, 297, C1103–C1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, V.; Guerra, M.; Jardim, C.; Melo, F.; Silva, W.; Ortega, J.M.; Robert, M.; Nathanson, M.H.; Leite, F. Nucleoplasmic Calcium Regulates Cell Proliferation through Legumain. J. Hepatol. 2011, 55, 626–635. [Google Scholar] [CrossRef] [Green Version]
- Micallef, L.; Belaubre, F.; Pinon, A.; Jayat-Vignoles, C.; Delage, C.; Charveron, M.; Simon, A. Effects of Extracellular Calcium on the Growth-Differentiation Switch in Immortalized Keratinocyte HaCaT Cells Compared with Normal Human Keratinocytes. Exp. Dermatol. 2009, 18, 143–151. [Google Scholar] [CrossRef]
- Lee, M.N.; Hwang, H.-S.; Oh, S.-H.; Roshanzadeh, A.; Kim, J.-W.; Song, J.H.; Kim, E.-S.; Koh, J.-T. Elevated Extracellular Calcium Ions Promote Proliferation and Migration of Mesenchymal Stem Cells via Increasing Osteopontin Expression. Exp. Mol. Med. 2018, 50, 1–16. [Google Scholar] [CrossRef]
- Rossi, F.; Noren, H.; Jove, R.; Beljanski, V.; Grinnemo, K.-H. Differences and Similarities between Cancer and Somatic Stem Cells: Therapeutic Implications. Stem Cell Res. Ther. 2020, 11, 489. [Google Scholar] [CrossRef]
- Swierenga, S.H.; Whitfield, J.F.; Karasaki, S. Loss of Proliferative Calcium Dependence: Simple in Vitro Indicator of Tumorigenicity. Proc. Natl. Acad. Sci. 1978, 75, 6069–6072. [Google Scholar] [CrossRef] [Green Version]
- Boynton, A.L.; Whitfield, J.F. Different Calcium Requirements for Proliferation of Conditionally and Unconditionally Tumorigenic Mouse Cells. Proc. Natl. Acad. Sci. USA 1976, 73, 1651–1654. [Google Scholar] [CrossRef] [Green Version]
- Boynton, A.L.; Whitfield, J.F. Different Extracellular Calcium Requirements for Proliferation of Nonneoplastic, Preneoplastic, and Neoplastic Mouse Cells. Cancer Res. 1977, 6, 2657–2661. [Google Scholar]
- Borowiec, A.-S.; Bidaux, G.; Tacine, R.; Dubar, P.; Pigat, N.; Delcourt, P.; Mignen, O.; Capiod, T. Are Orai1 and Orai3 Channels More Important than Calcium Influx for Cell Proliferation? Biochim. Biophys. Acta BBA-Mol. Cell Res. 2014, 1843, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Déliot, N.; Constantin, B. Plasma Membrane Calcium Channels in Cancer: Alterations and Consequences for Cell Proliferation and Migration. Biochim. Biophys. Acta BBA-Biomembr. 2015, 1848, 2512–2522. [Google Scholar] [CrossRef] [Green Version]
- Terrié, E.; Déliot, N.; Benzidane, Y.; Harnois, T.; Cousin, L.; Bois, P.; Oliver, L.; Arnault, P.; Vallette, F.; Constantin, B.; et al. Store-Operated Calcium Channels Control Proliferation and Self-Renewal of Cancer Stem Cells from Glioblastoma. Cancers 2021, 13, 3428. [Google Scholar] [CrossRef]
- Dumont, N.A.; Bentzinger, C.F.; Sincennes, M.C.; Rudnicki, M.A. Satellite Cells and Skeletal Muscle Regeneration. Compr. Physiol. 2015, 5, 1027–1059. [Google Scholar] [CrossRef]
- Franco, S.J.; Rodgers, M.A.; Perrin, B.J.; Han, J.; Bennin, D.A.; Critchley, D.R.; Huttenlocher, A. Calpain-Mediated Proteolysis of Talin Regulates Adhesion Dynamics. Nat. Cell Biol. 2004, 6, 977–983. [Google Scholar] [CrossRef]
- Chan, K.T.; Bennin, D.A.; Huttenlocher, A. Regulation of Adhesion Dynamics by Calpain-Mediated Proteolysis of Focal Adhesion Kinase (FAK). J. Biol. Chem. 2010, 285, 11418–11426. [Google Scholar] [CrossRef] [Green Version]
- Brundage, R.A.; Fogarty, K.E.; Tuft, R.A.; Fay, F.S. Calcium Gradients Underlying Polarization and Chemotaxis of Eosinophils. Science 1991, 254, 703–706. [Google Scholar] [CrossRef]
- Wei, C.; Wang, X.; Chen, M.; Ouyang, K.; Song, L.-S.; Cheng, H. Calcium Flickers Steer Cell Migration. Nature 2009, 457, 901–905. [Google Scholar] [CrossRef] [Green Version]
- Hammad, A.S.; Machaca, K. Store Operated Calcium Entry in Cell Migration and Cancer Metastasis. Cells 2021, 10, 1246. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tollance, A.; Koenig, S.; Liaudet, N.; Frieden, M. Activation and Migration of Human Skeletal Muscle Stem Cells In Vitro Differently Rely on Calcium Signals. Cells 2022, 11, 1689. https://doi.org/10.3390/cells11101689
Tollance A, Koenig S, Liaudet N, Frieden M. Activation and Migration of Human Skeletal Muscle Stem Cells In Vitro Differently Rely on Calcium Signals. Cells. 2022; 11(10):1689. https://doi.org/10.3390/cells11101689
Chicago/Turabian StyleTollance, Axel, Stéphane Koenig, Nicolas Liaudet, and Maud Frieden. 2022. "Activation and Migration of Human Skeletal Muscle Stem Cells In Vitro Differently Rely on Calcium Signals" Cells 11, no. 10: 1689. https://doi.org/10.3390/cells11101689
APA StyleTollance, A., Koenig, S., Liaudet, N., & Frieden, M. (2022). Activation and Migration of Human Skeletal Muscle Stem Cells In Vitro Differently Rely on Calcium Signals. Cells, 11(10), 1689. https://doi.org/10.3390/cells11101689