
Activation of MT1/MT2 to Protect Testes and Leydig Cells against Cisplatin-Induced Oxidative Stress through the SIRT1/Nrf2 Signaling Pathway

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Study Participants
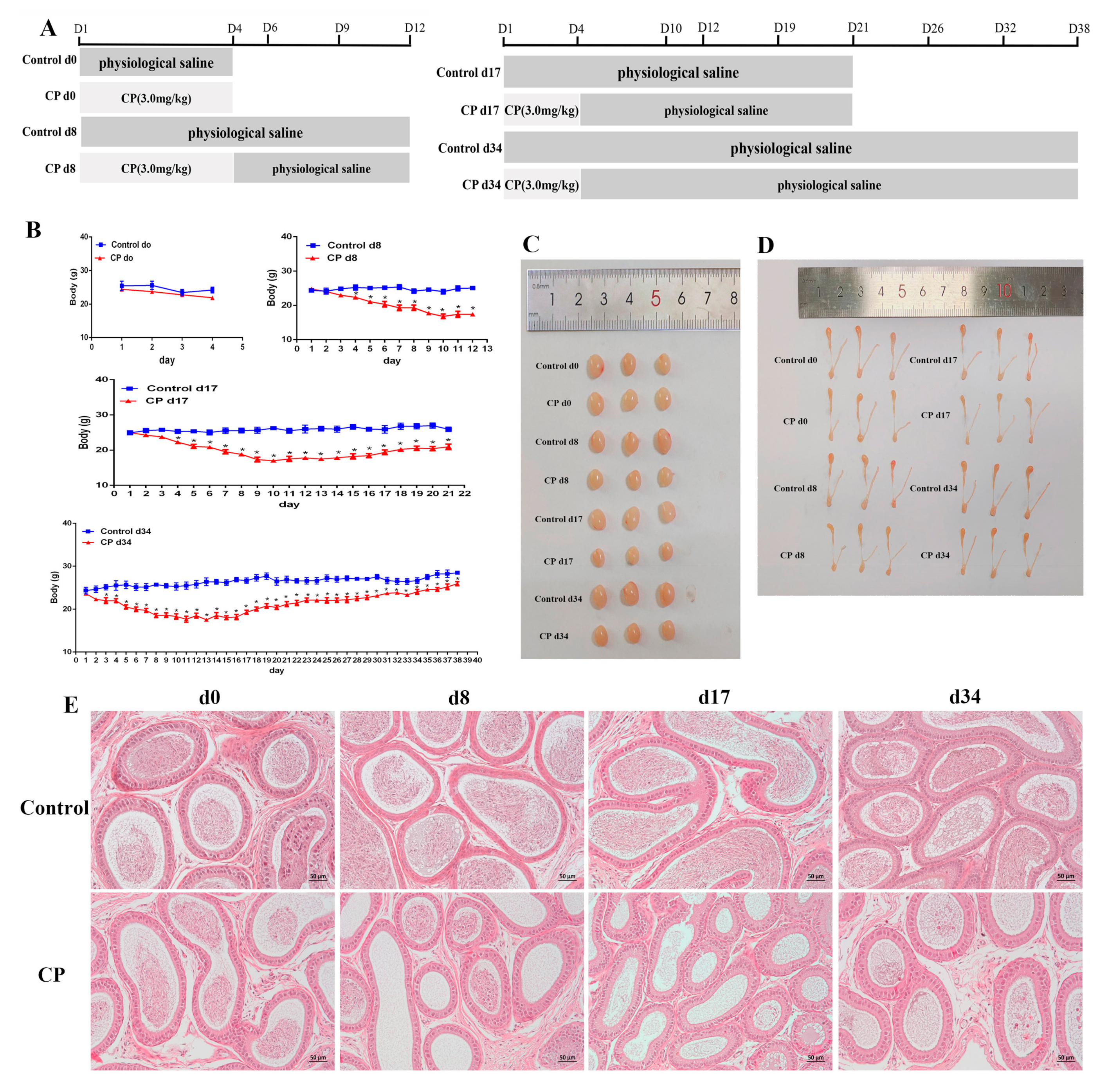
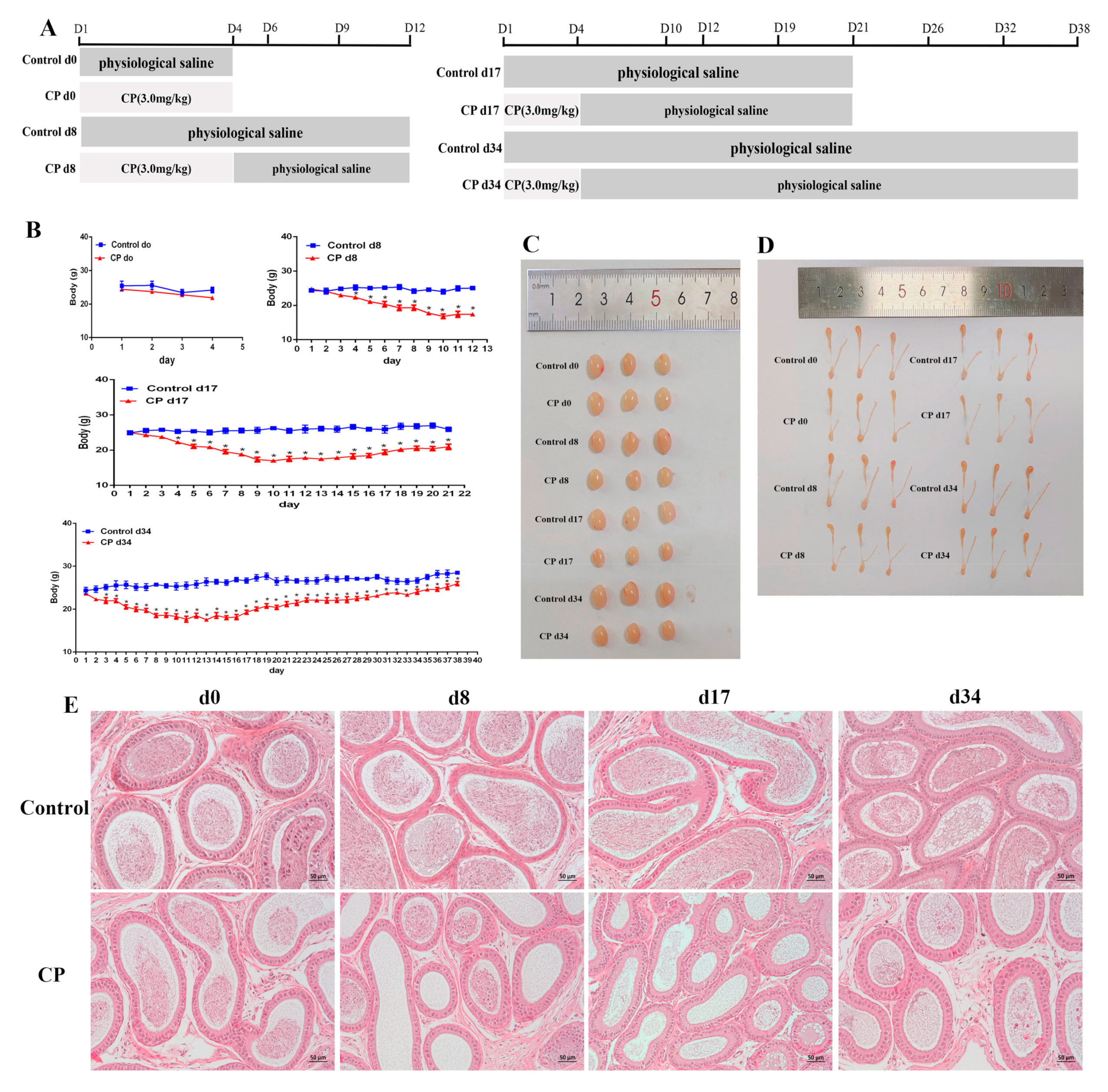
2.3. Animal Experiments and Design
- Control d0 group and CP d0 group: The mice were administered an intraperitoneal injection of physiological saline (same volume as for the CP d0 group) and CP (3 mg/kg) for 4 consecutive days [1]. Then, the mice were sacrificed;
- Control d8 group and CP d8 group: The mice were administered an intraperitoneal injection of physiological saline (same volume as for the CP d8 group) and CP (3 mg/kg) for 4 consecutive days. Then, the mice were sacrificed on the 12th day;
- Control d17 group and CP d17 group: The mice were administered an intraperitoneal injection of physiological saline (same volume as for the CP d17 group) and CP (3 mg/kg) for 4 consecutive days. Then, the mice were sacrificed on the 21st day;
- Control d34 group and CP d34 group: The mice were administered an intraperitoneal injection of physiological saline (same volume as for the CP d34 group) and CP (3 mg/kg) for 4 consecutive days. Then, the mice were sacrificed on the 37th day.
2.4. Blood and Tissue Collection
2.5. Sperm Count
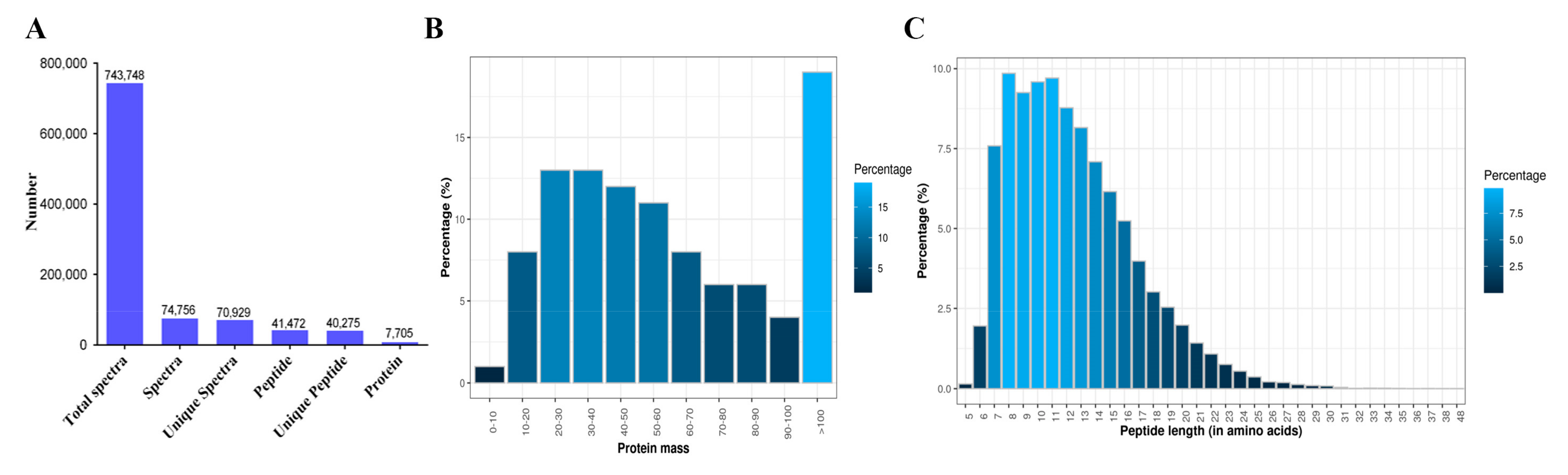
2.6. Protein Extraction, iTRAQ Labeling, and Proteomic Analysis
2.7. Isolation and Culture of Testicular Leydig Cells
2.8. Histological Evaluation of Testes
2.9. ELISA Assay
2.10. Biochemical Markers of Oxidative Damage Assay
2.11. TUNEL Staining
2.12. Immunohistochemistry
2.13. Immunofluorescence
2.14. RT-PCR
2.15. Annexin V-FITC Apoptosis Detection
2.16. Immunoblotting
2.17. Statistical Analysis
3. Results
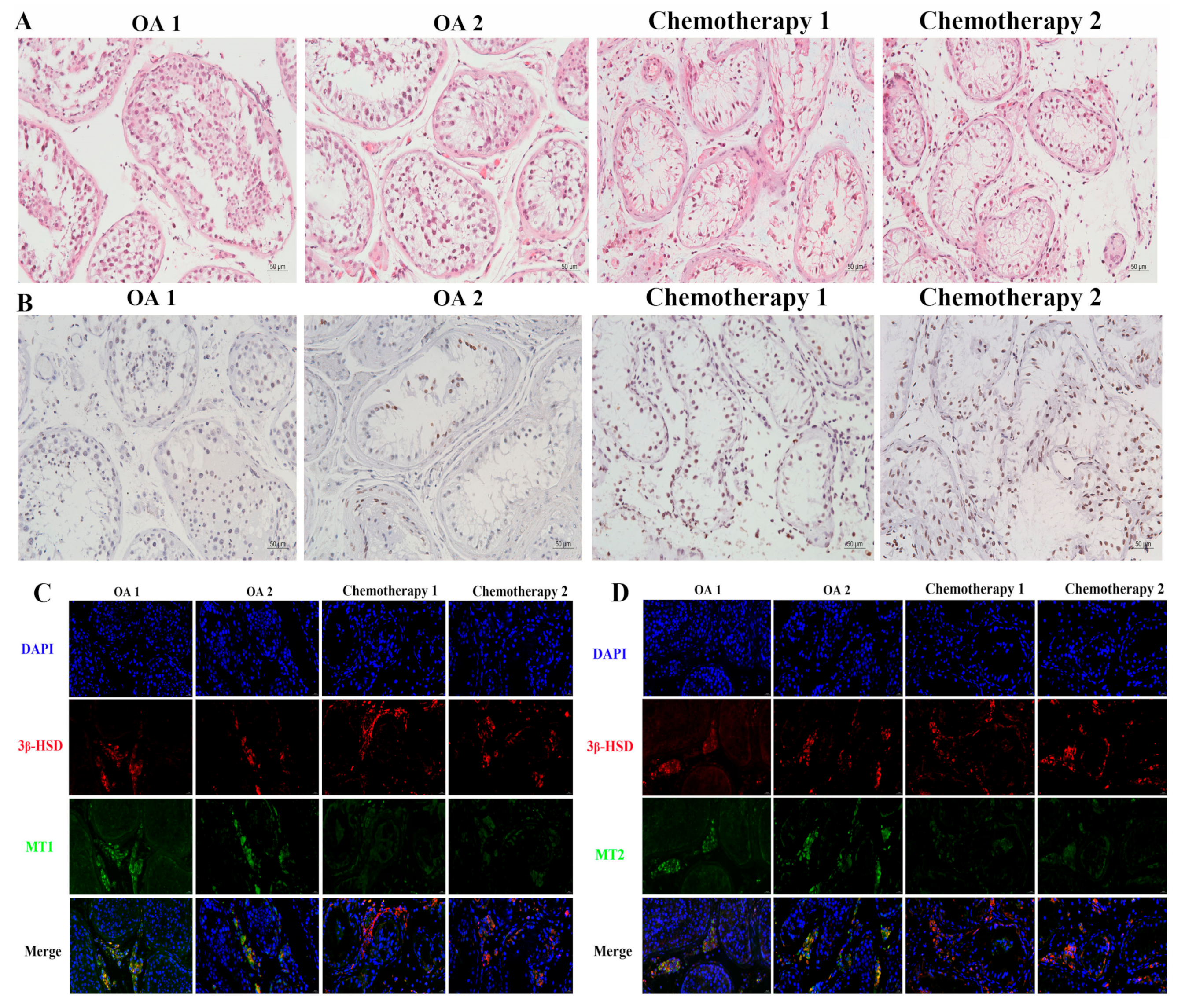
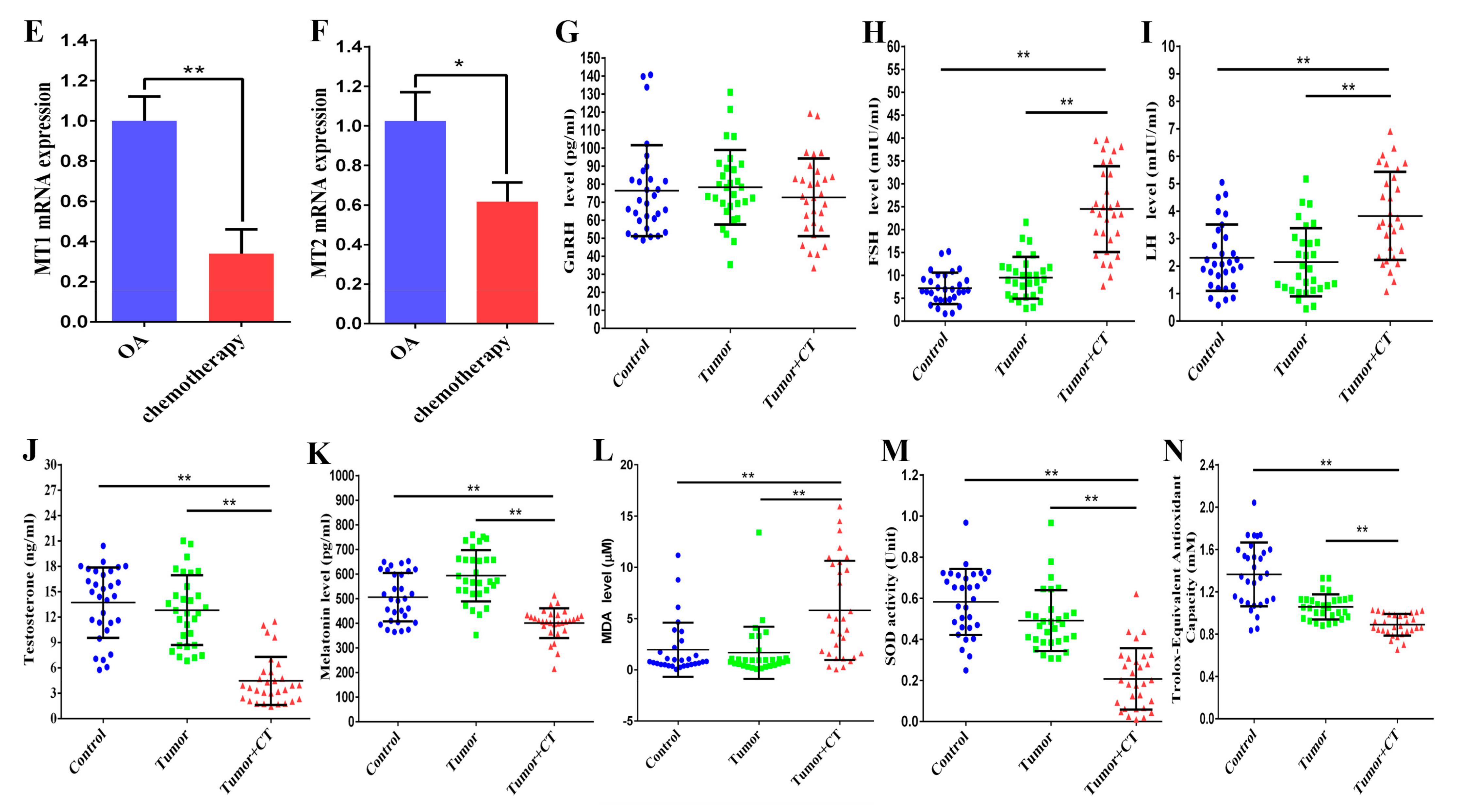
3.1. Effects of Platinum Chemotherapeutic Drugs on the Levels of Gonadotropin, Melatonin, and Oxidative Stress in the Peripheral Blood of Male Gastrointestinal Tumor Patients, and Reducing the Expression of MT1/MT2 in the Testis
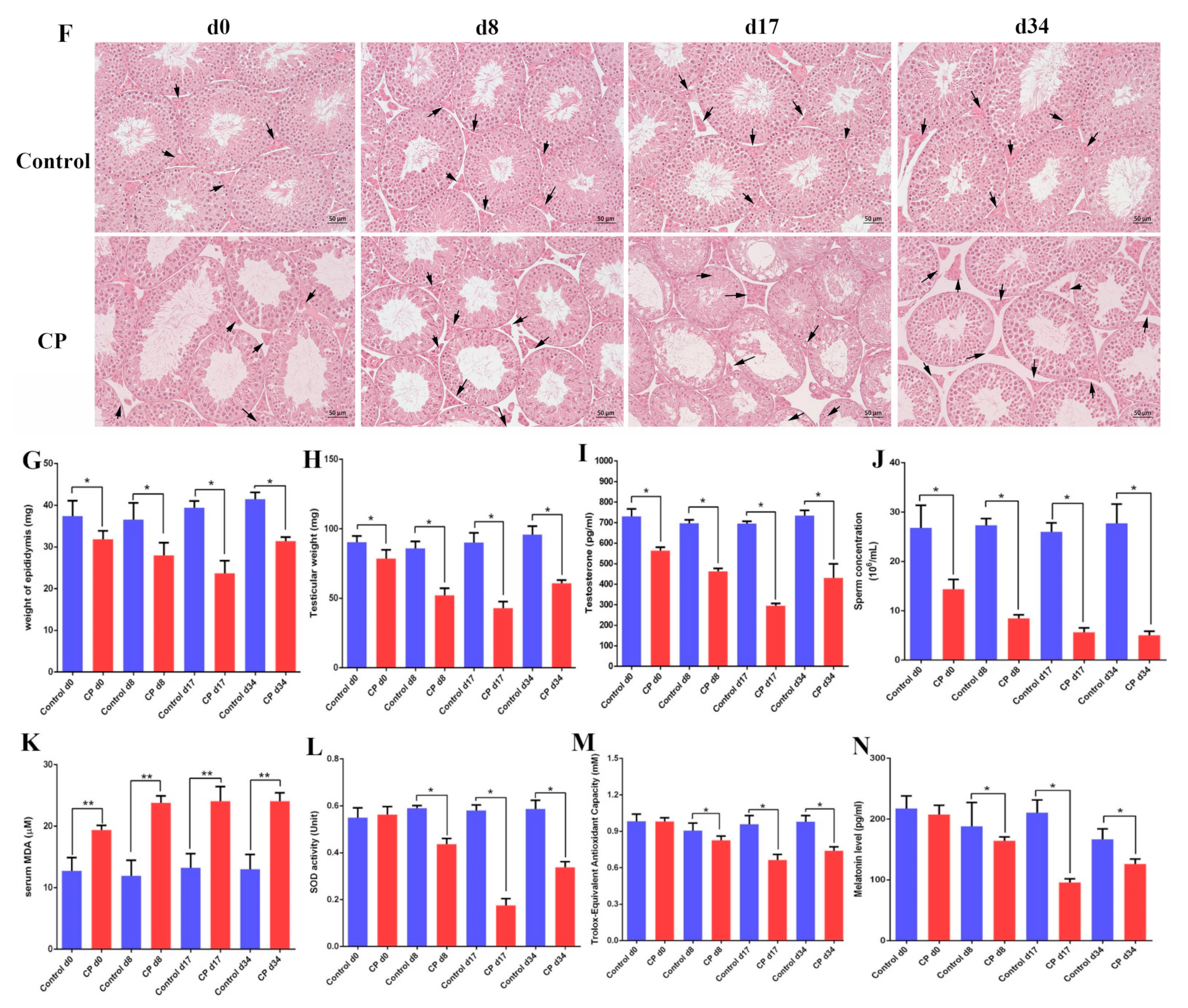
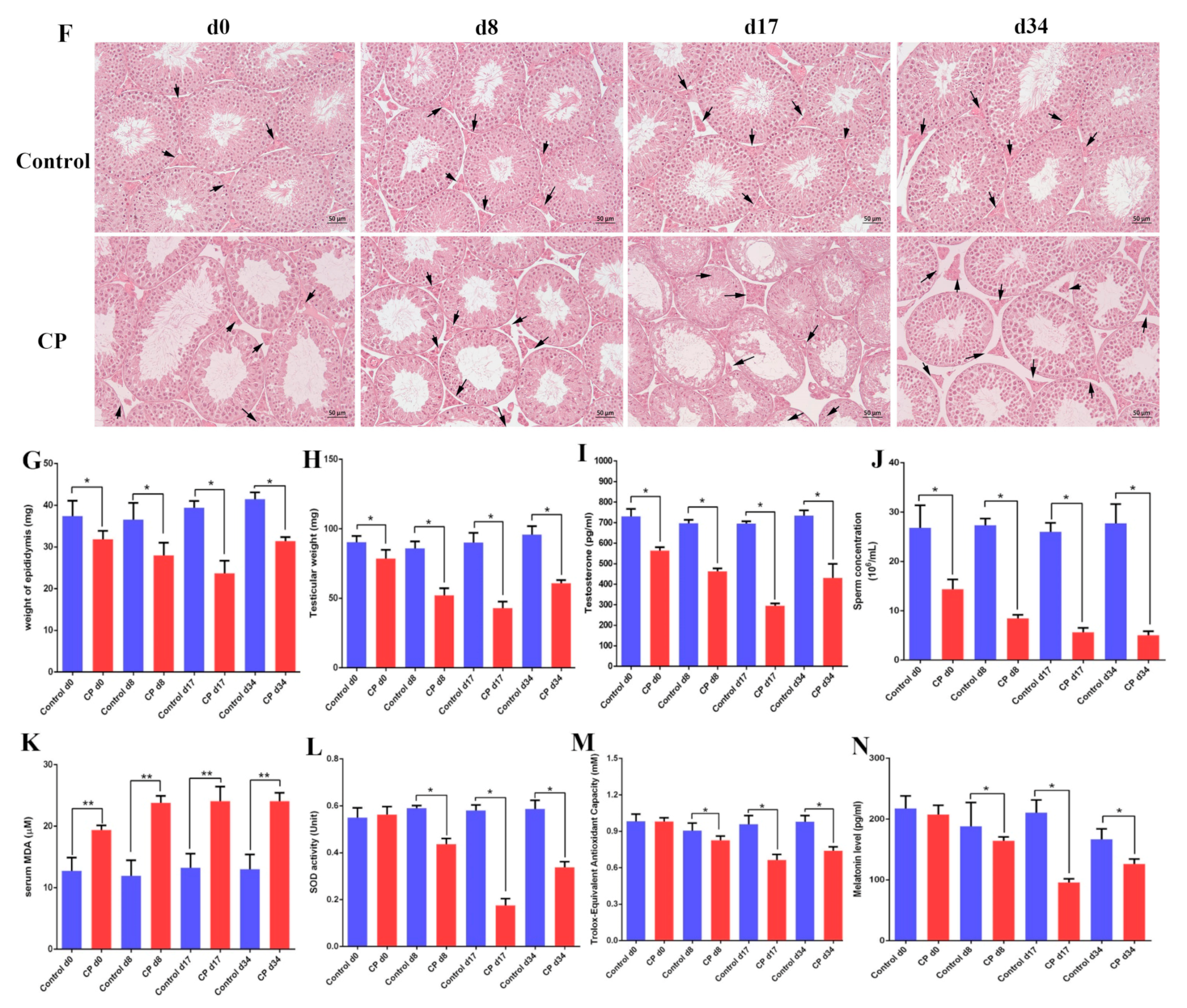
3.2. Cisplatin Treatment Resulted in Damage to the Testes of Male Mice and Increased Oxidative Stress
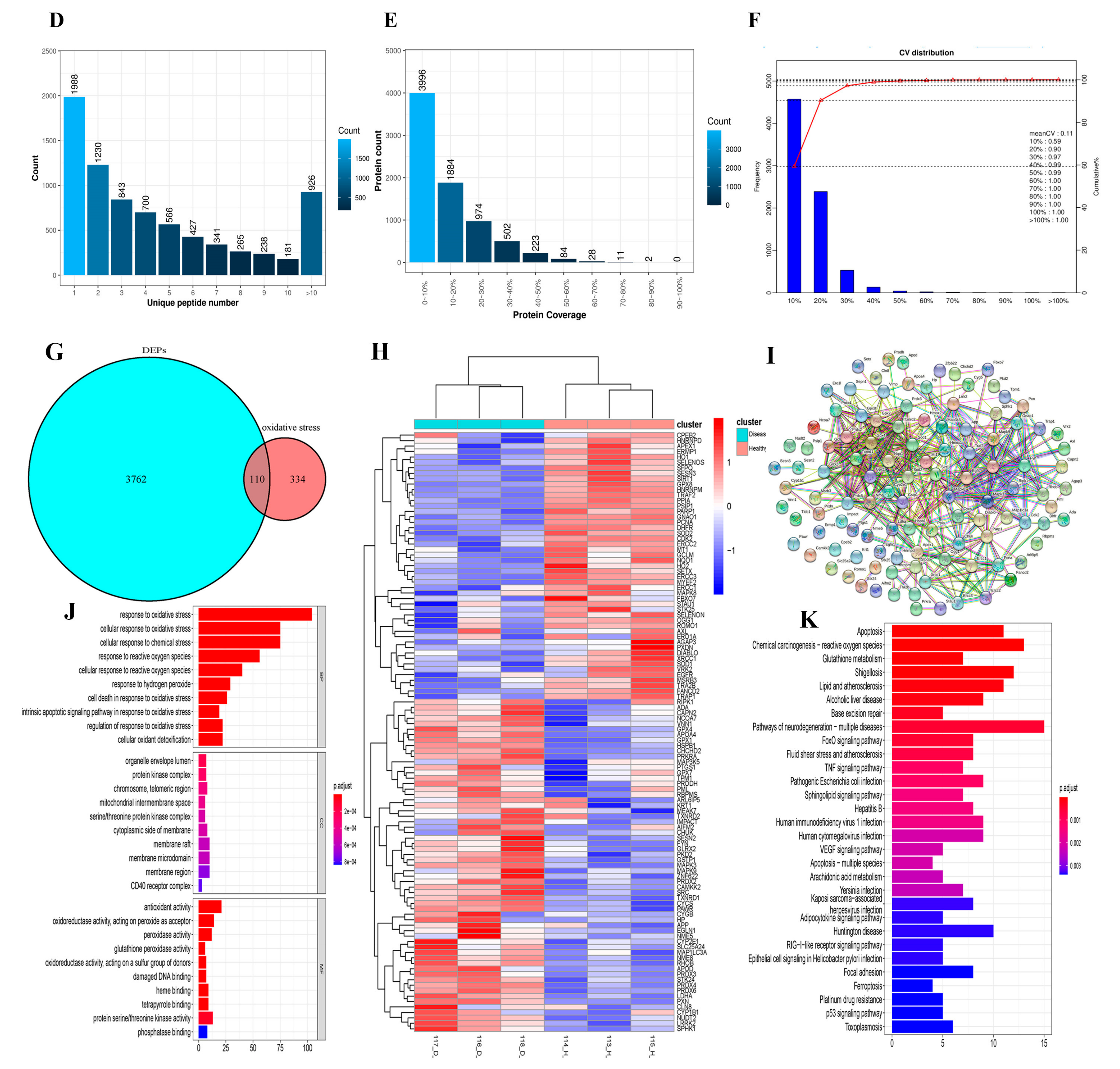
3.3. iTRAQ Quantitative Proteomics of Mouse Testes and Expression of Oxidative-Stress-Related Proteins
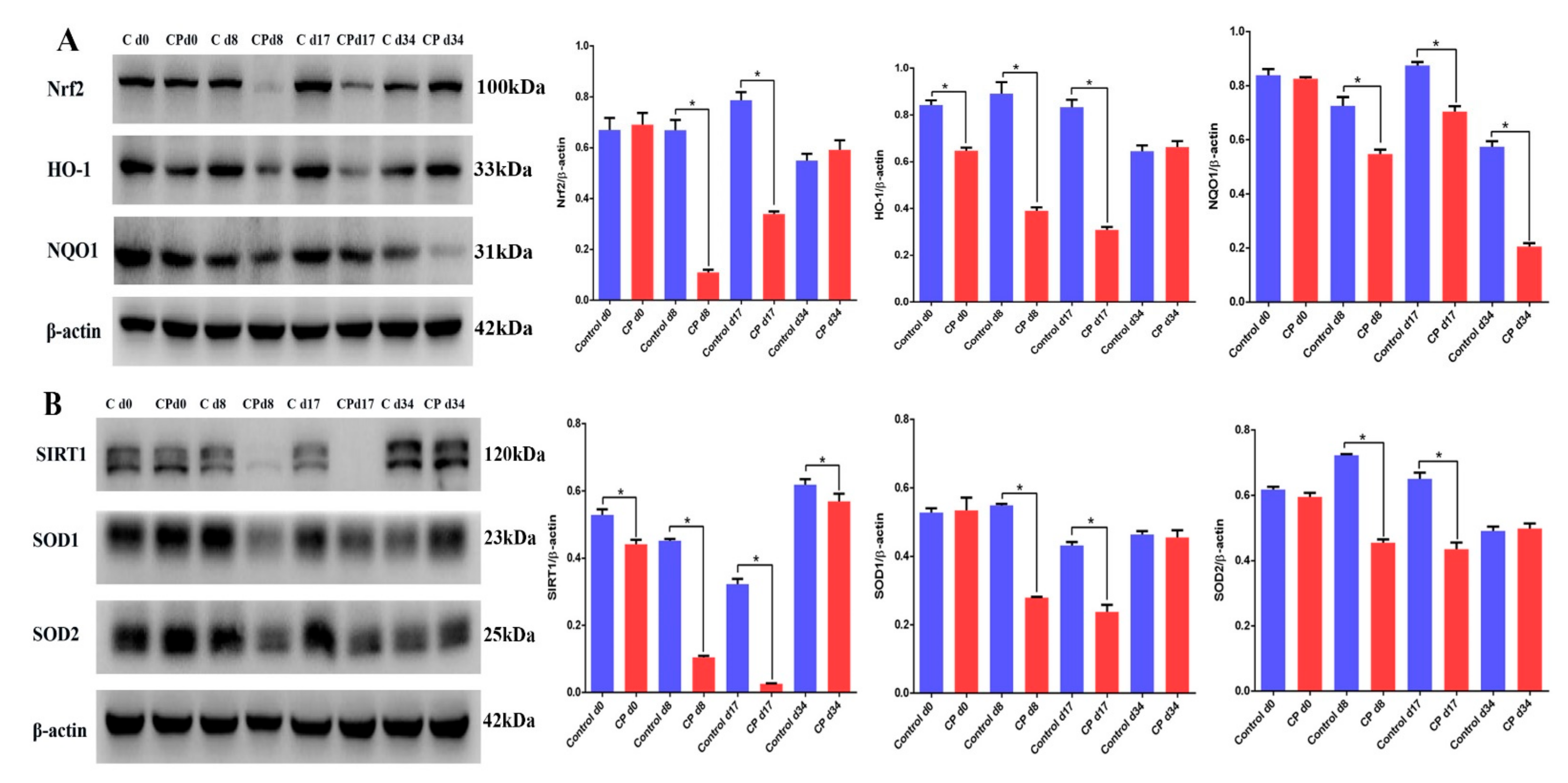
3.4. Cisplatin Attenuates the Expression of SIRT1/Nrf2 Signaling at Different Recovery Times
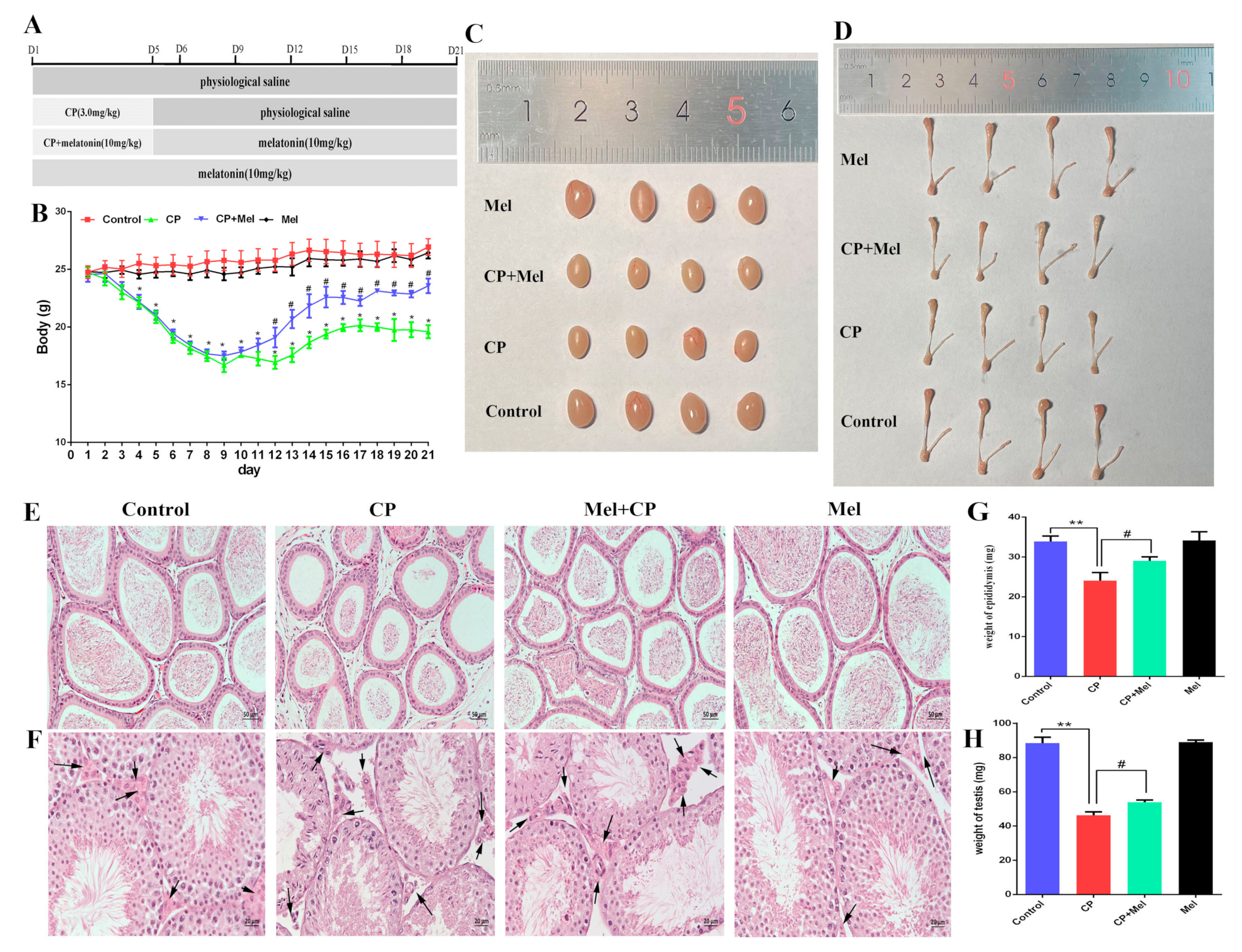
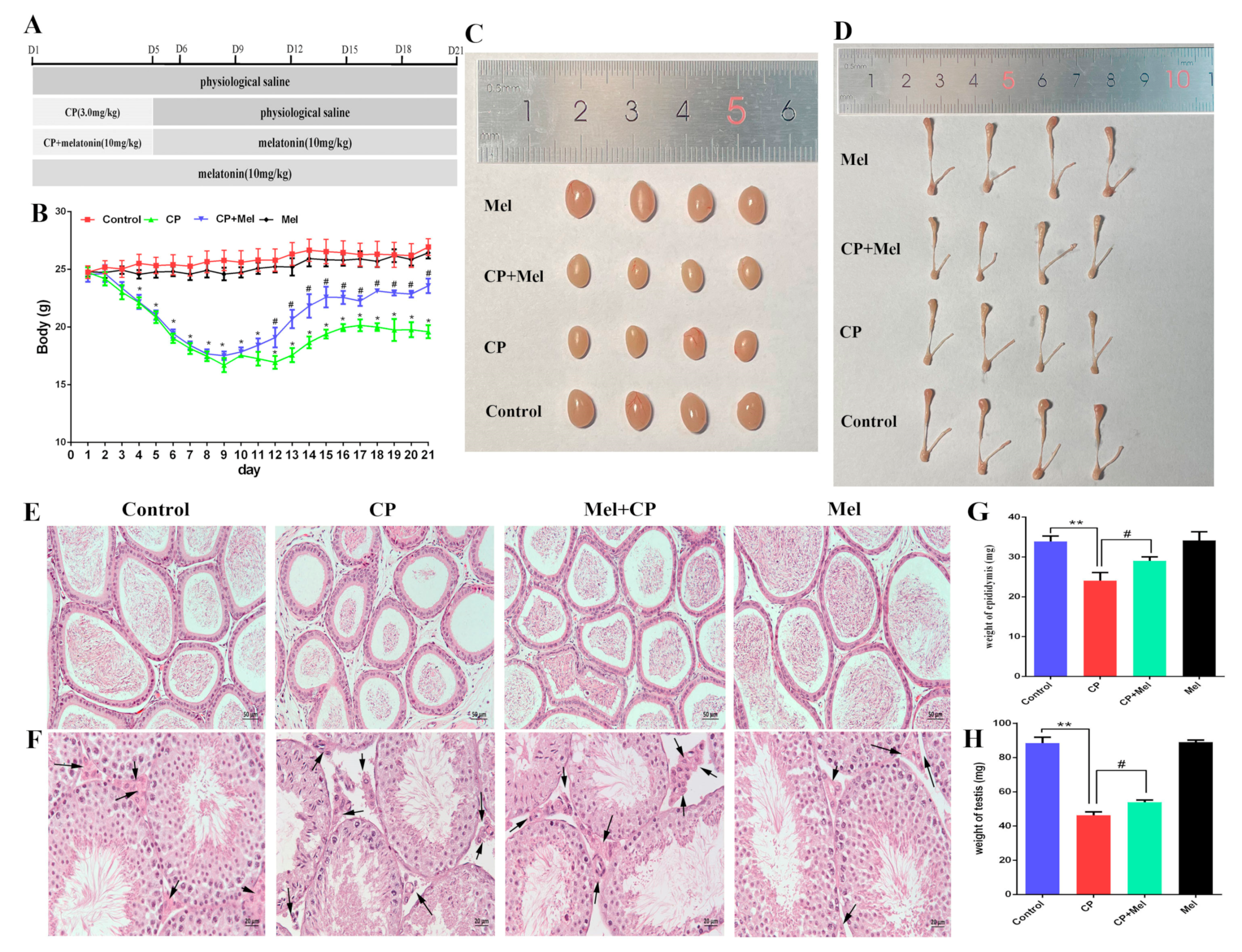
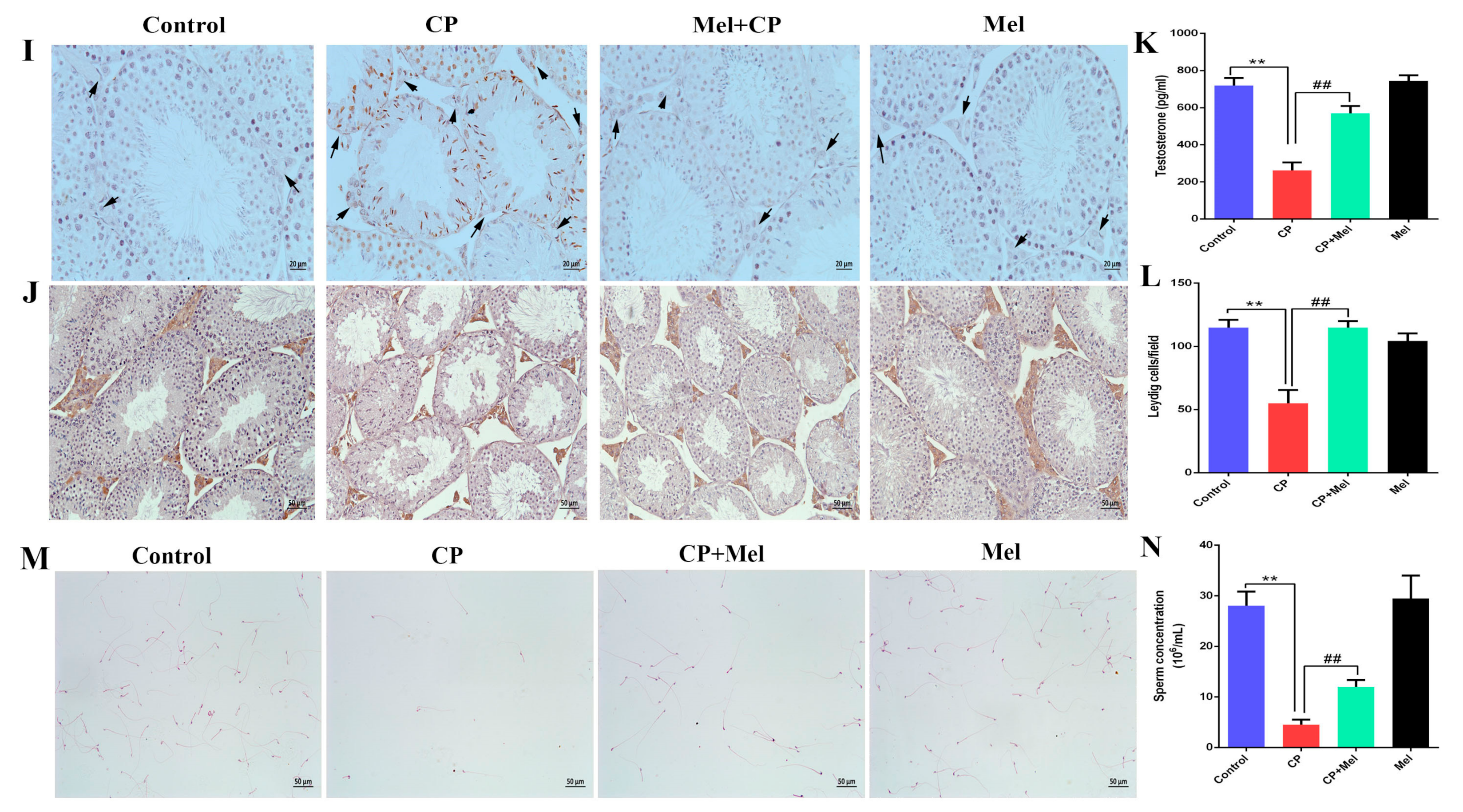
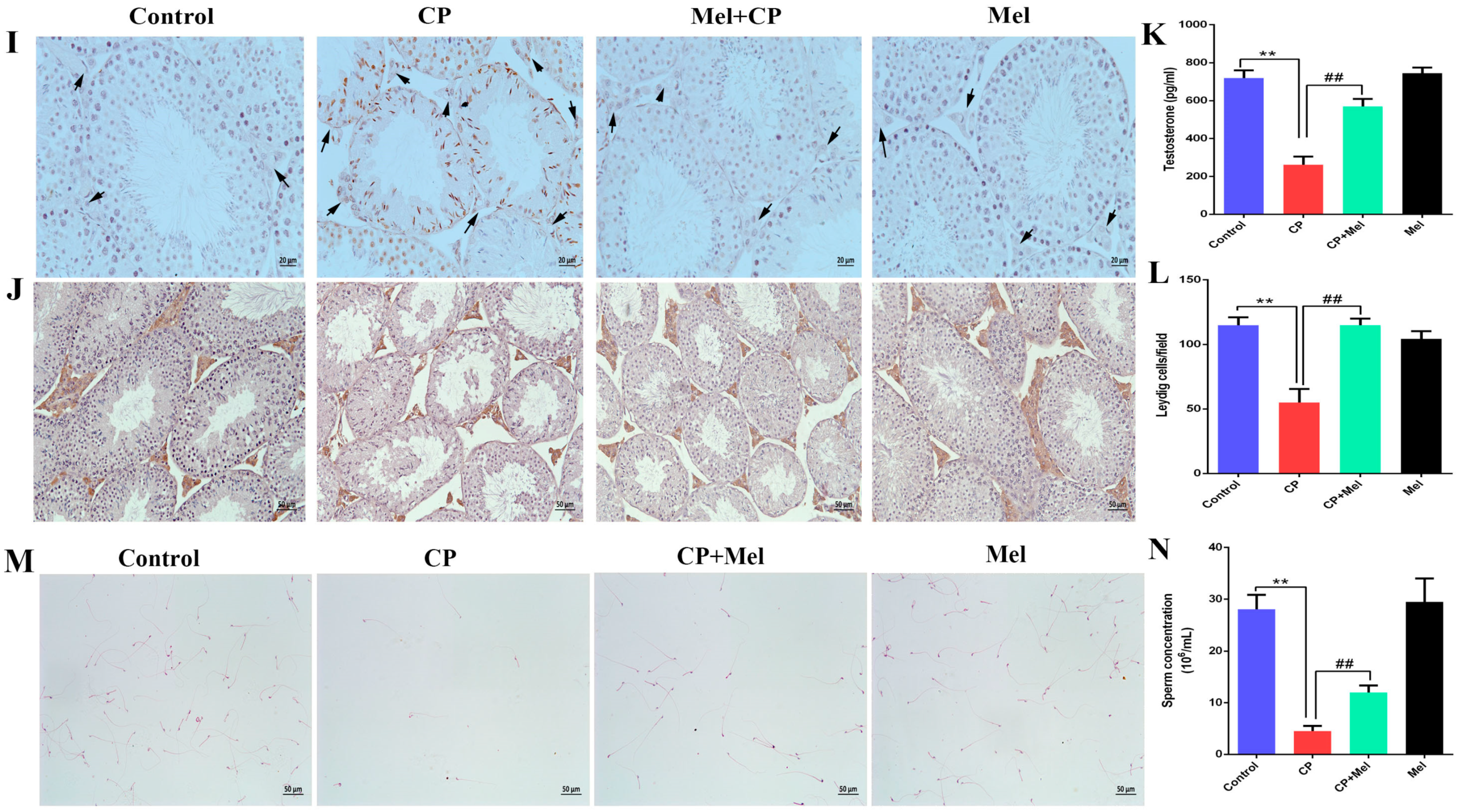
3.5. Melatonin Can Alleviate the Damage to Mouse Testes and LEYDIG Cells Caused by Cisplatin
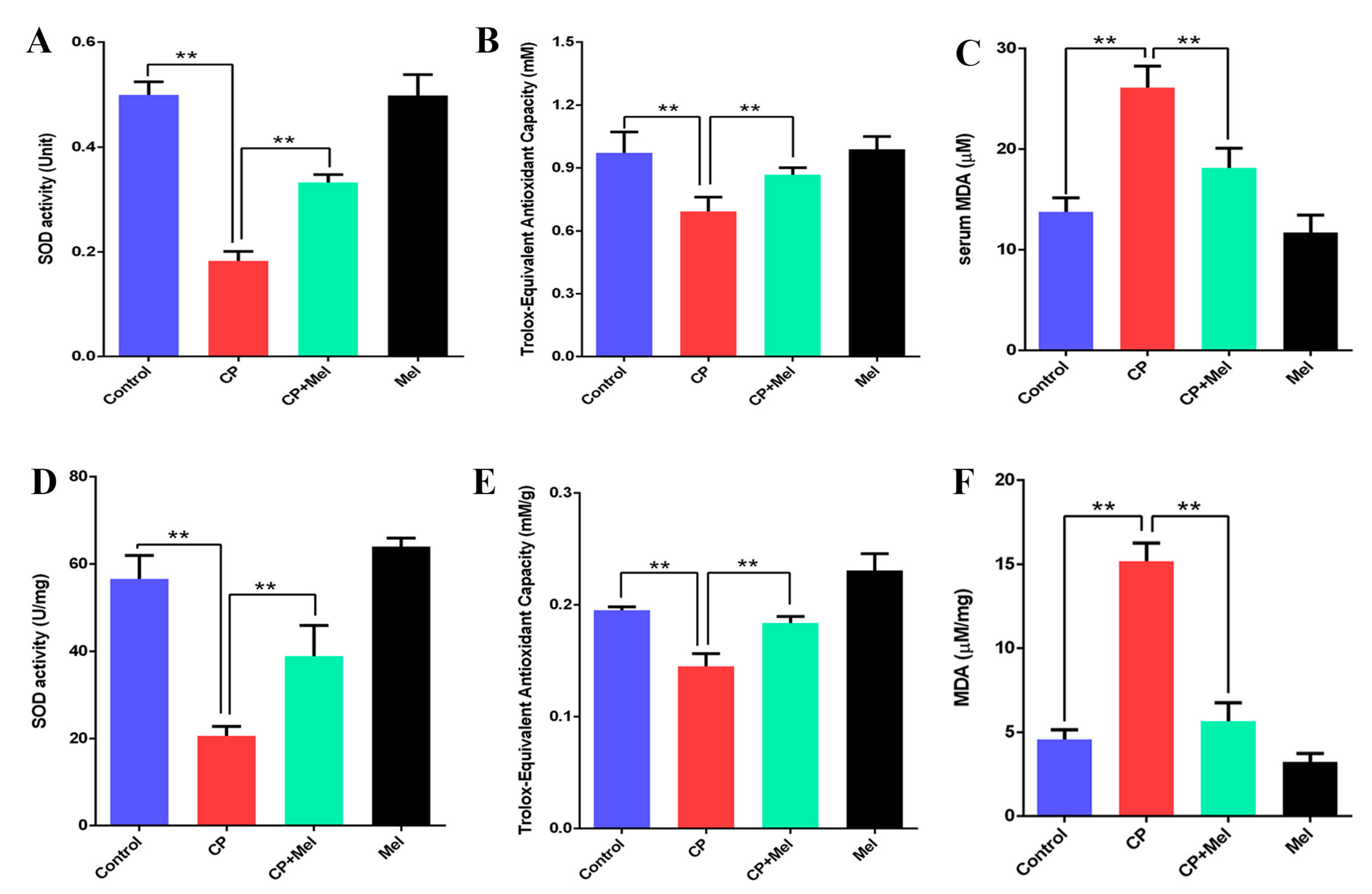
3.6. Melatonin Can Attenuate Oxidative Stress Caused by Cisplatin in Mouse Sera and Testes
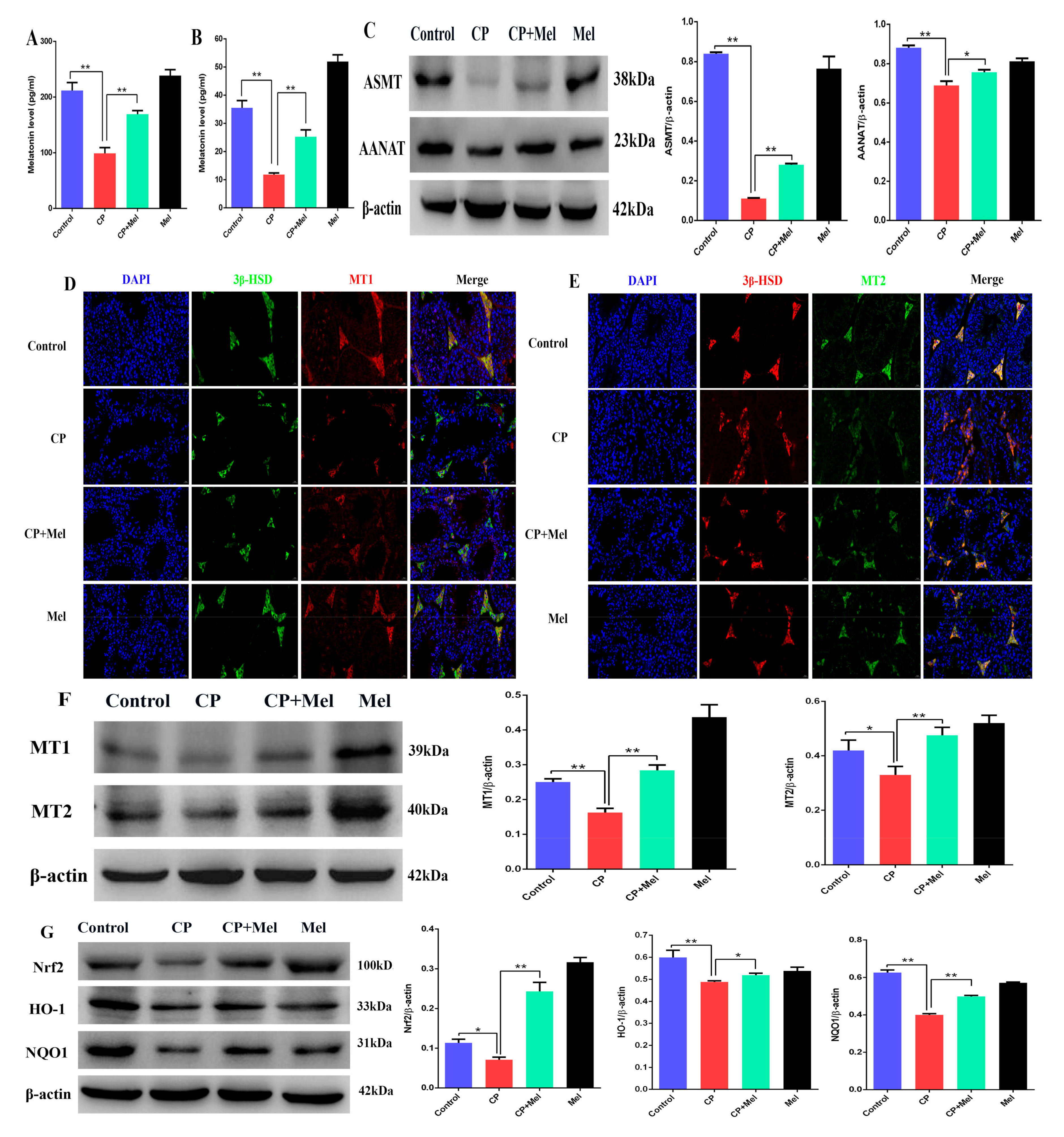
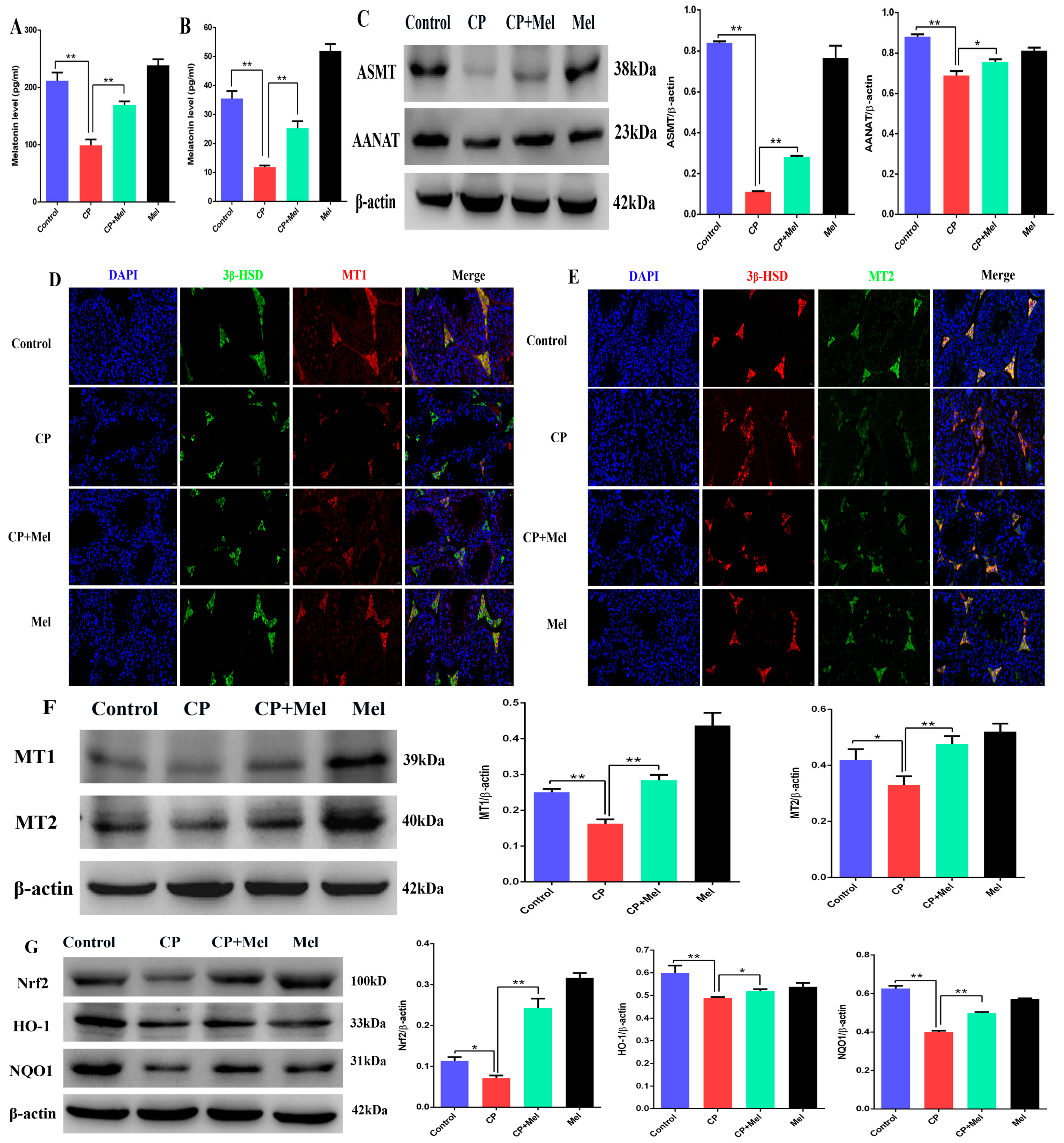
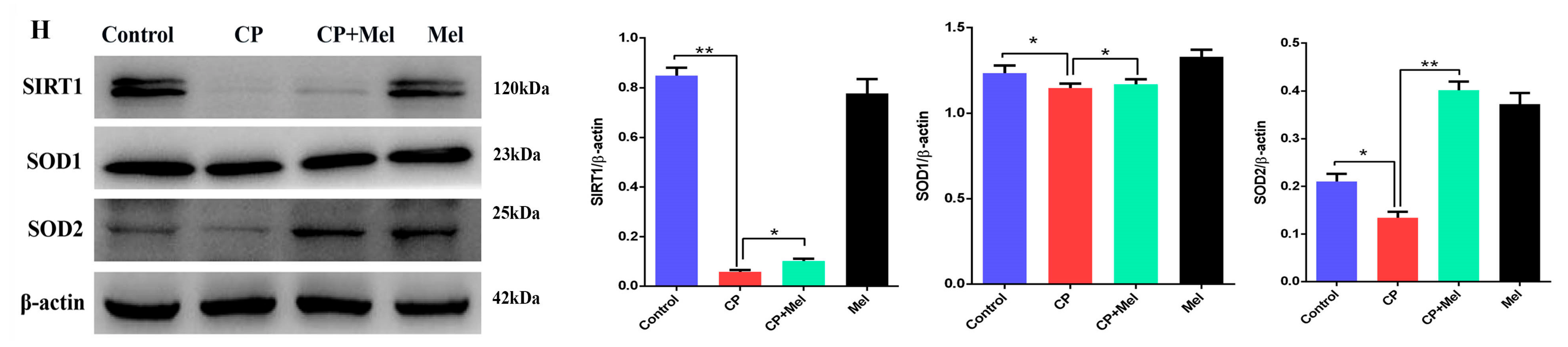
3.7. Melatonin Could Attenuate the Downregulation of MT1, MT2, and SIRT1/Nrf2 Antioxidant Signaling by Cisplatin in Mouse Testes
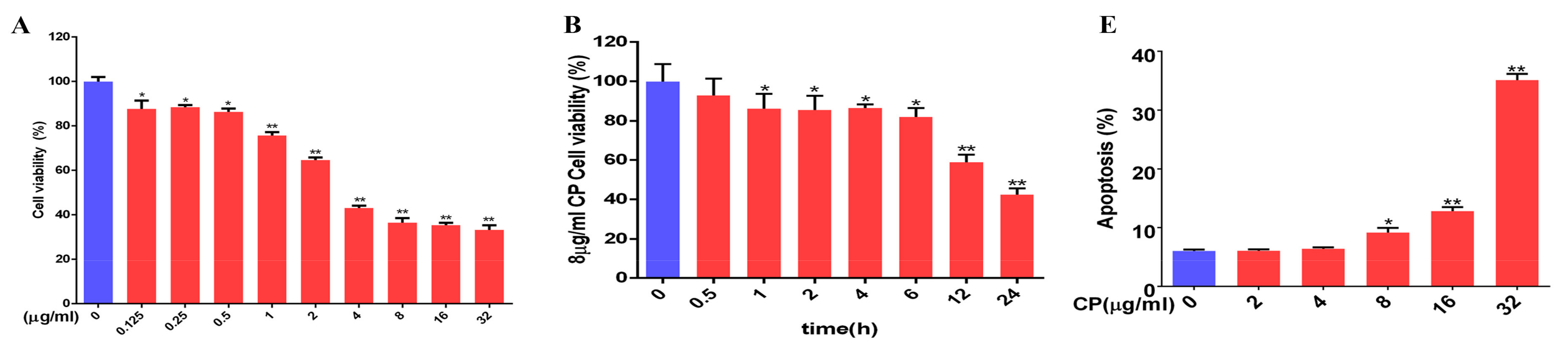
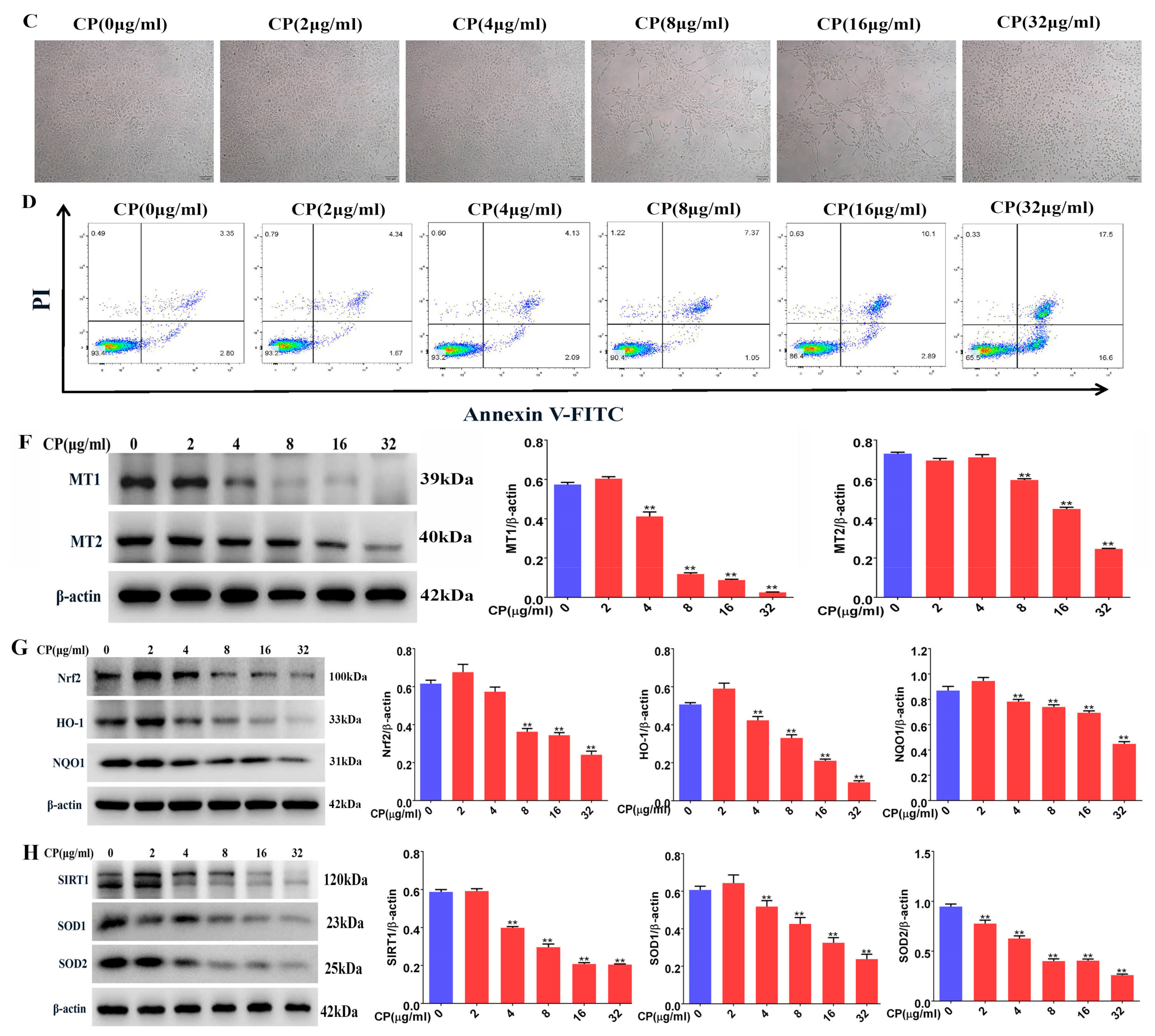
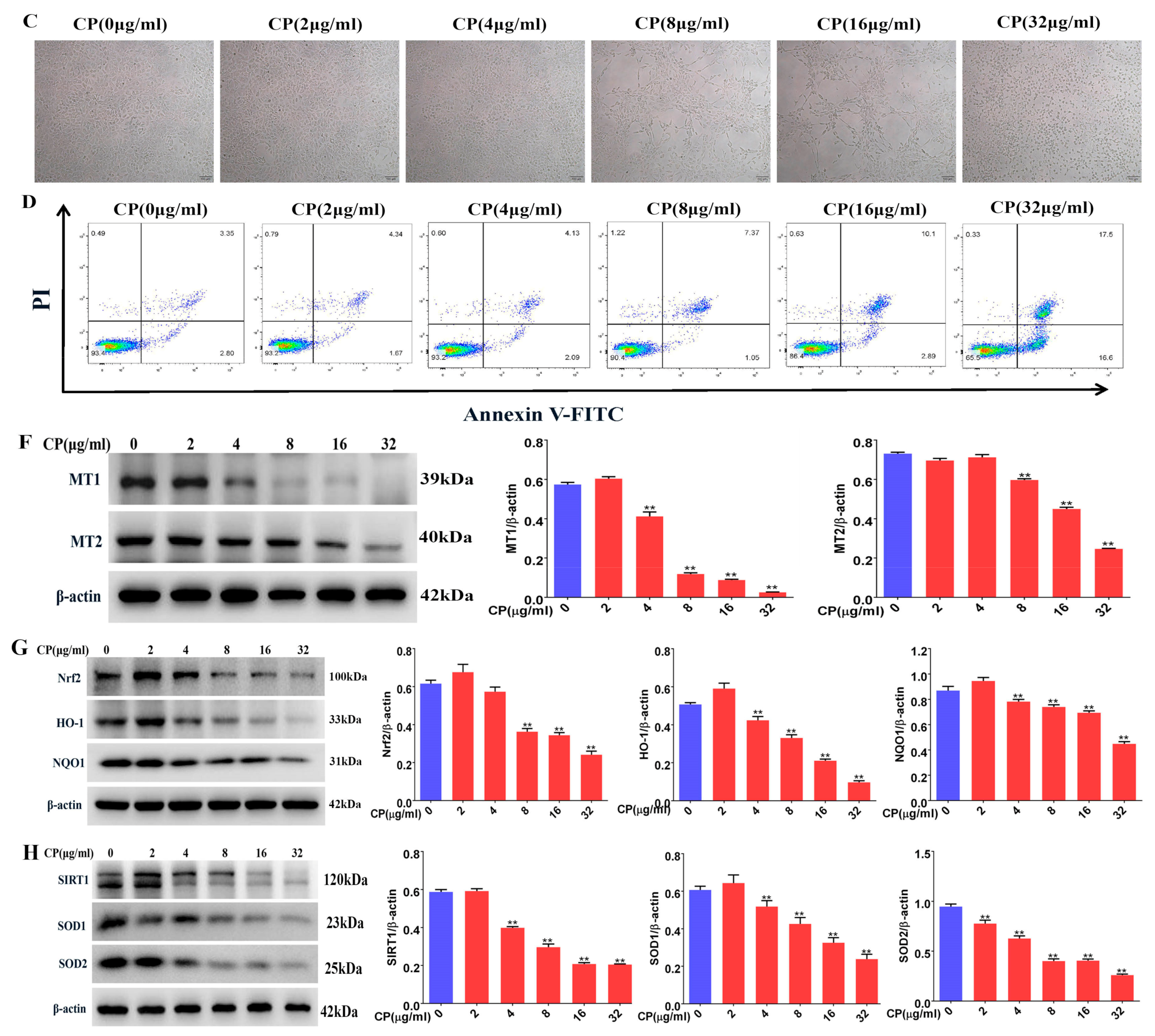
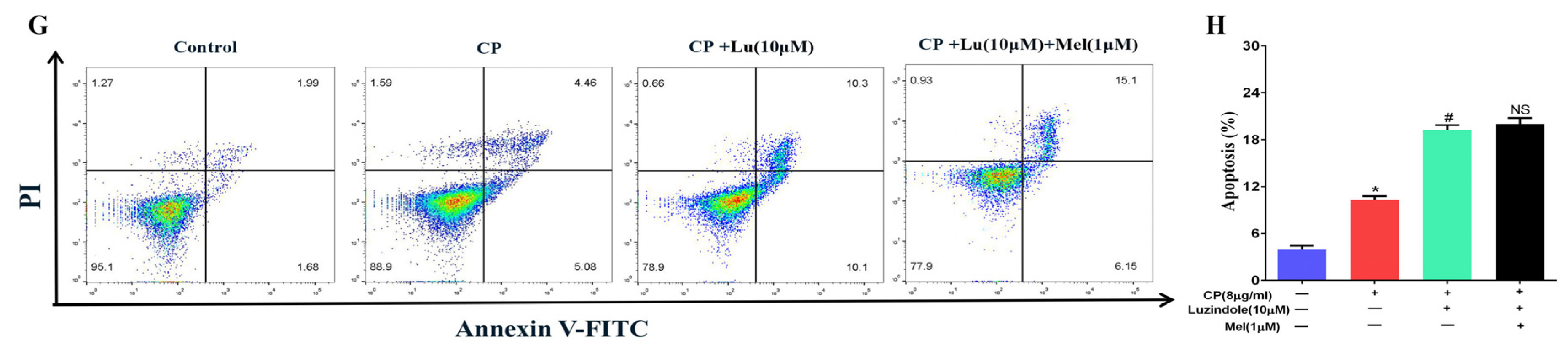
3.8. Cisplatin Increased Apoptosis and Downregulated MT1, MT2, and SIRT1/Nrf2 Signaling Pathways in Leydig Cells
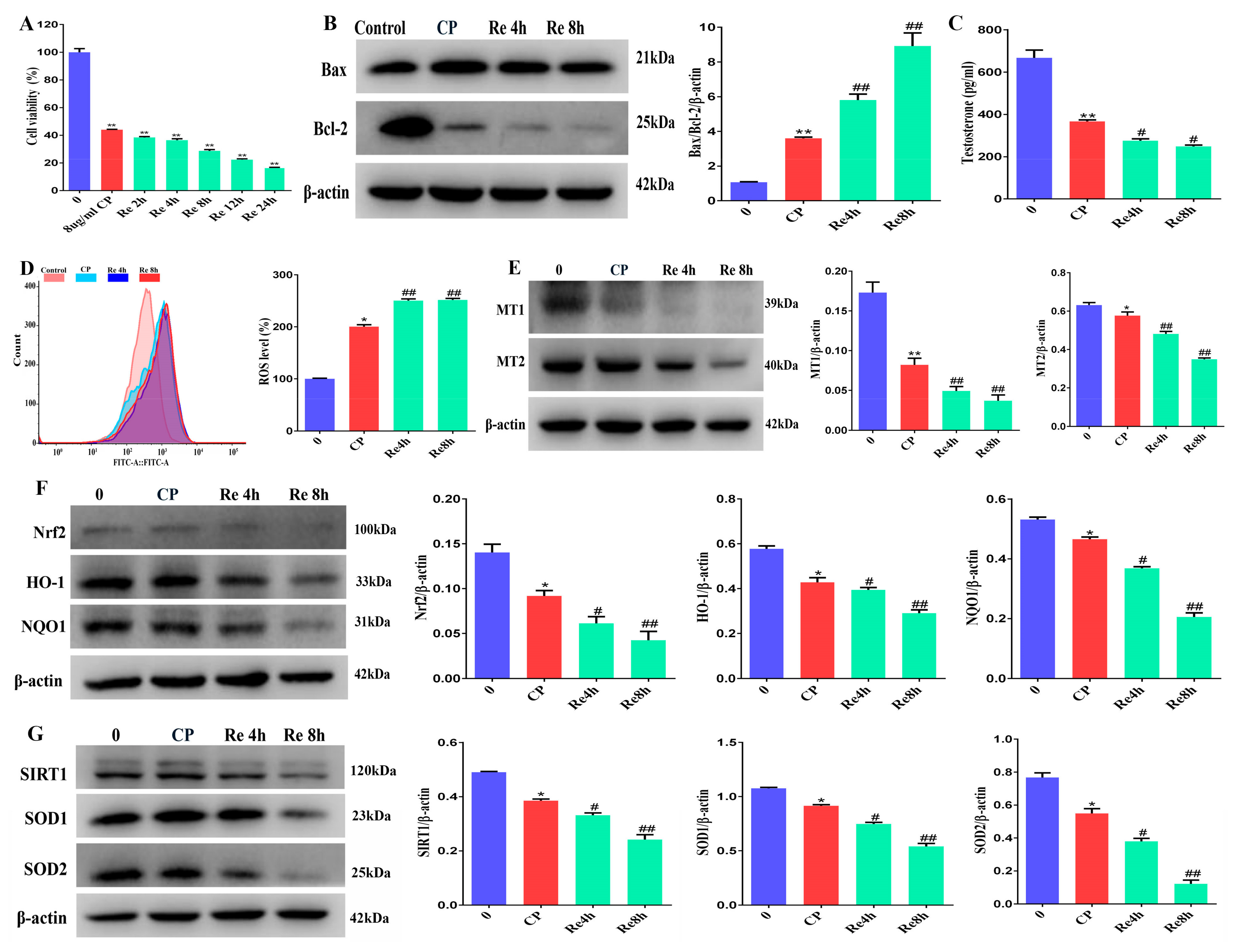
3.9. Leydig Cells Failed to Self-Recovery after Removing Cisplatin
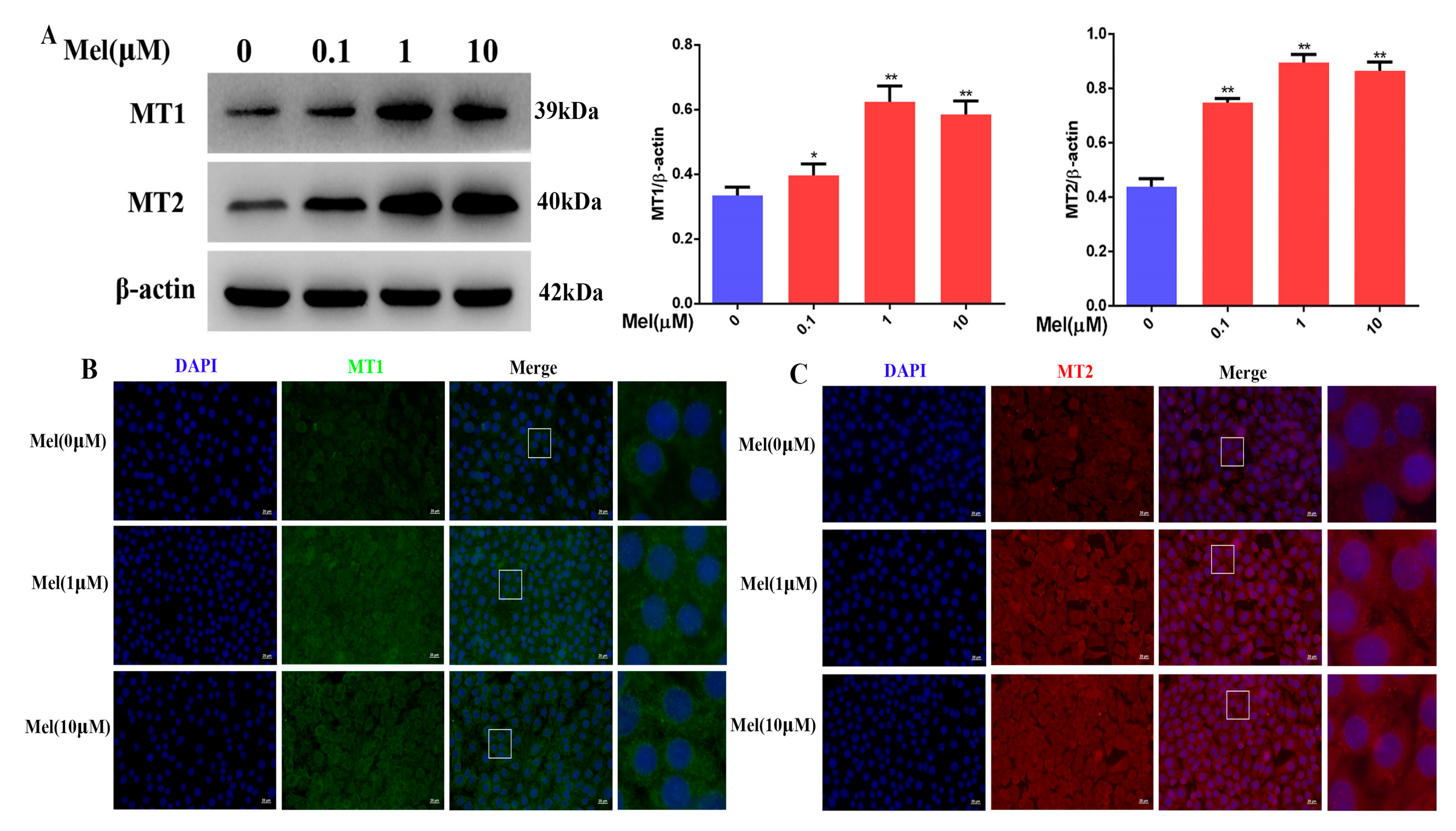
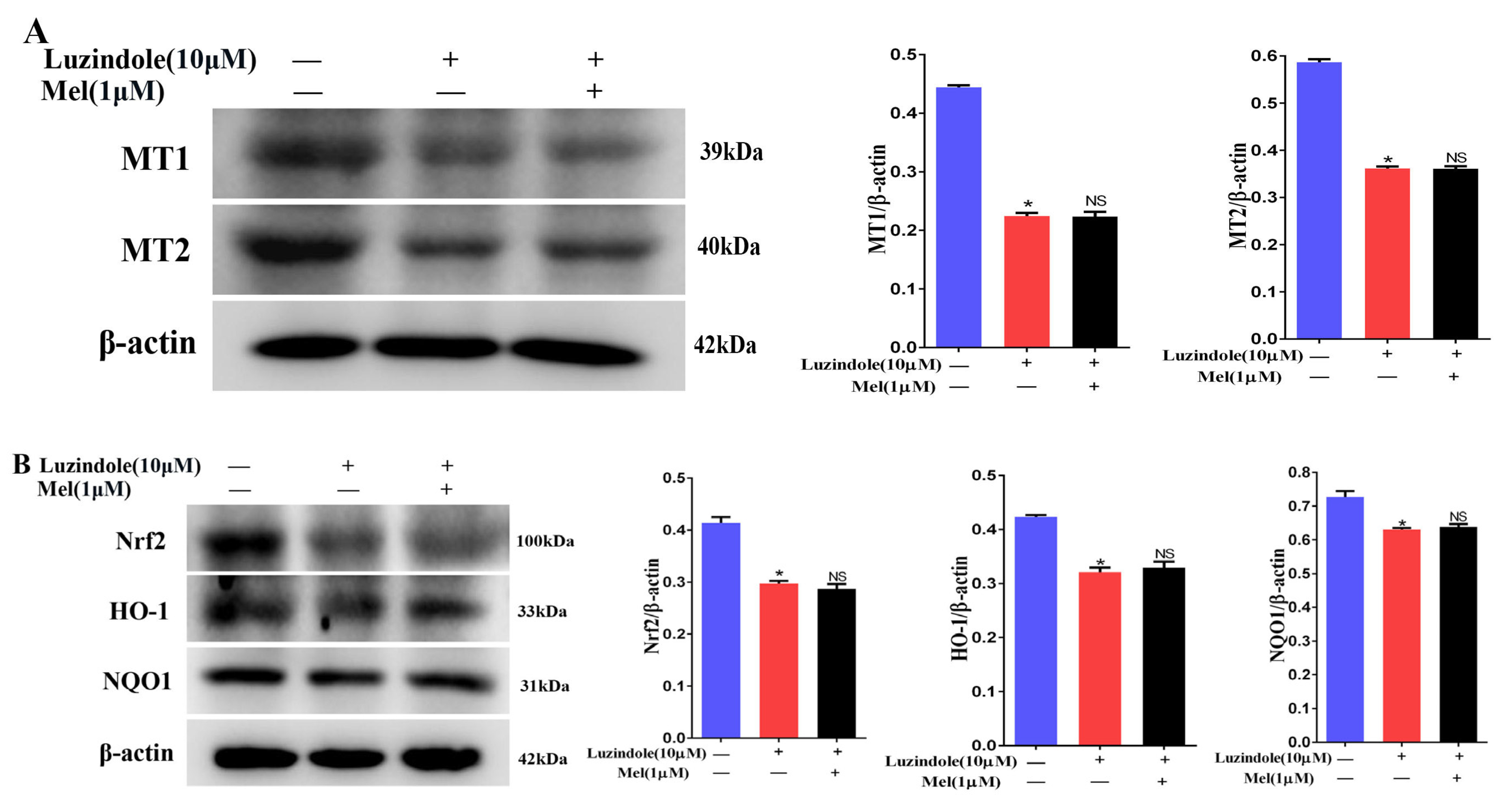
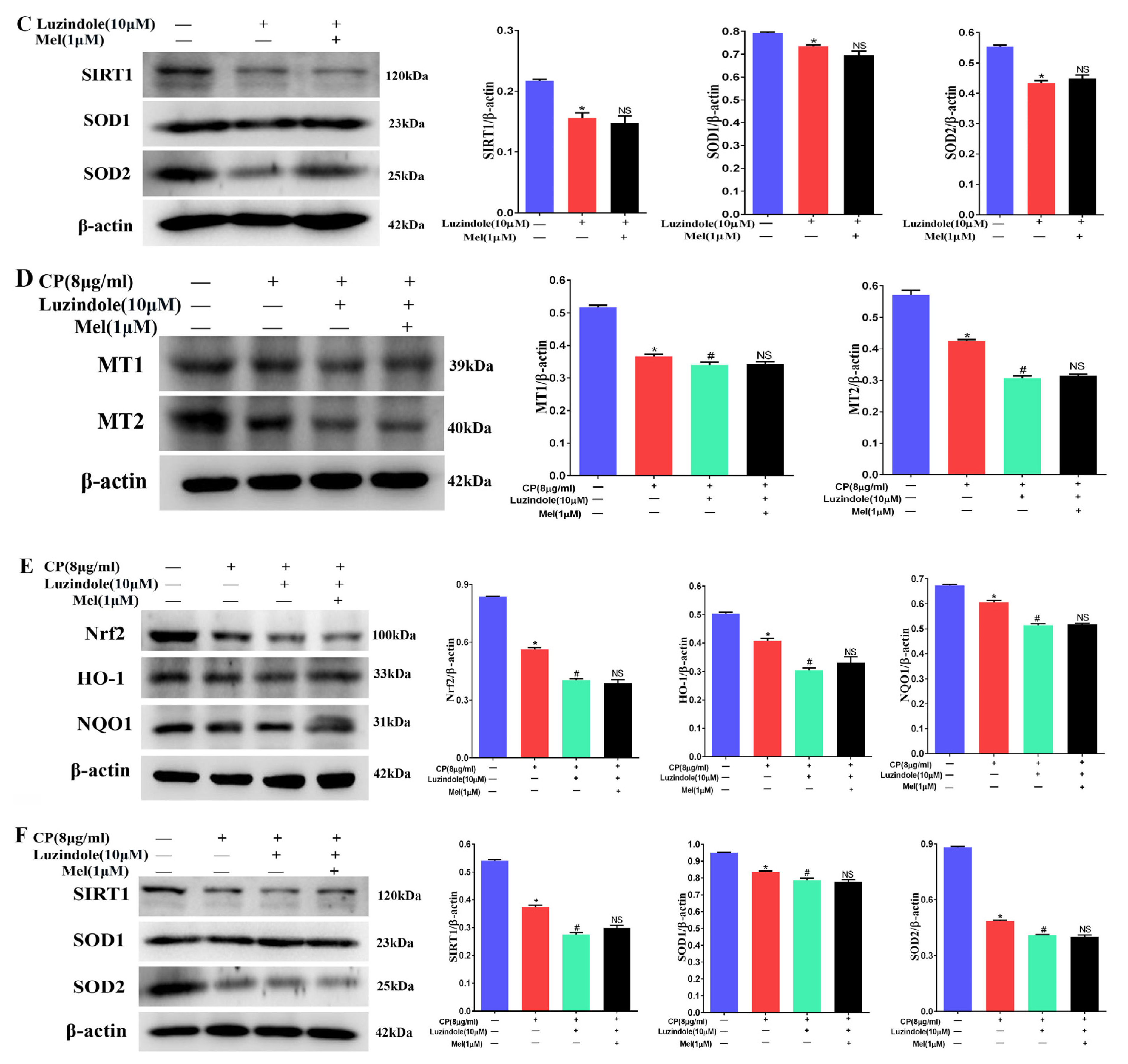
3.10. Melatonin Could Attenuate the Downregulation of the MT1, MT2, and SIRT1/Nrf2 Signaling Pathways by Cisplatin in Leydig Cells
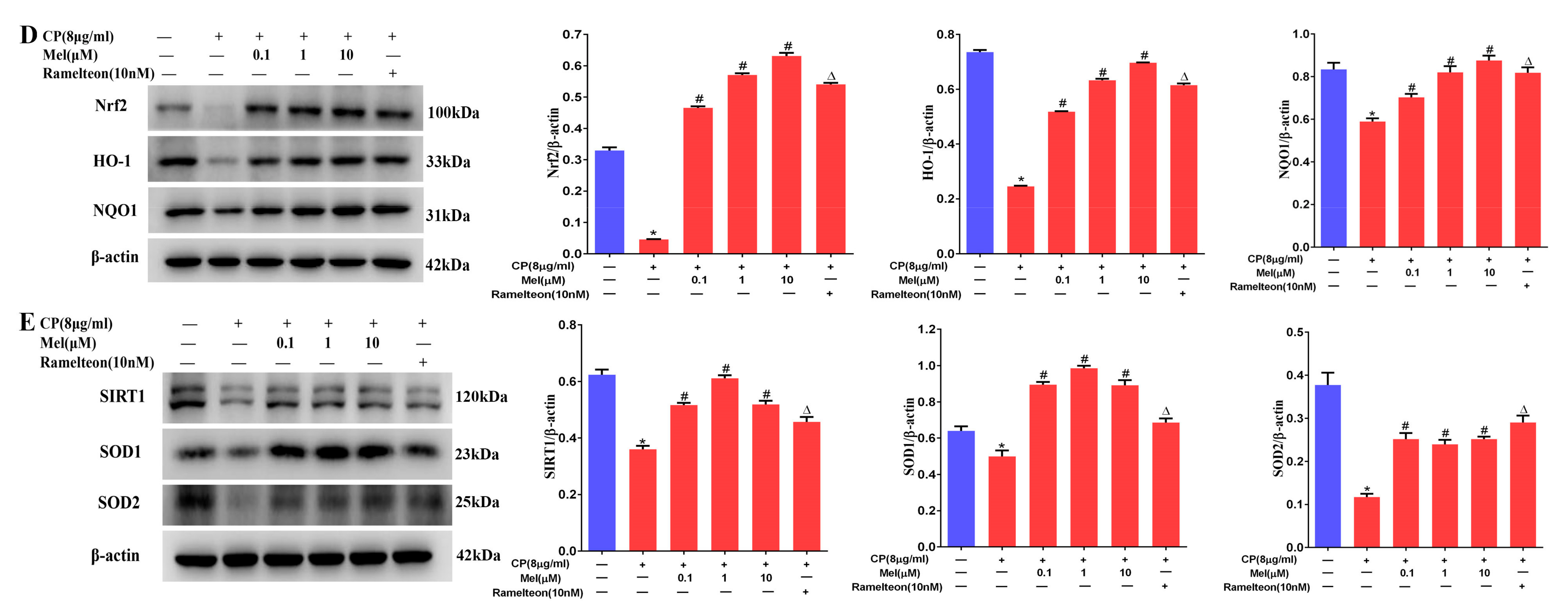
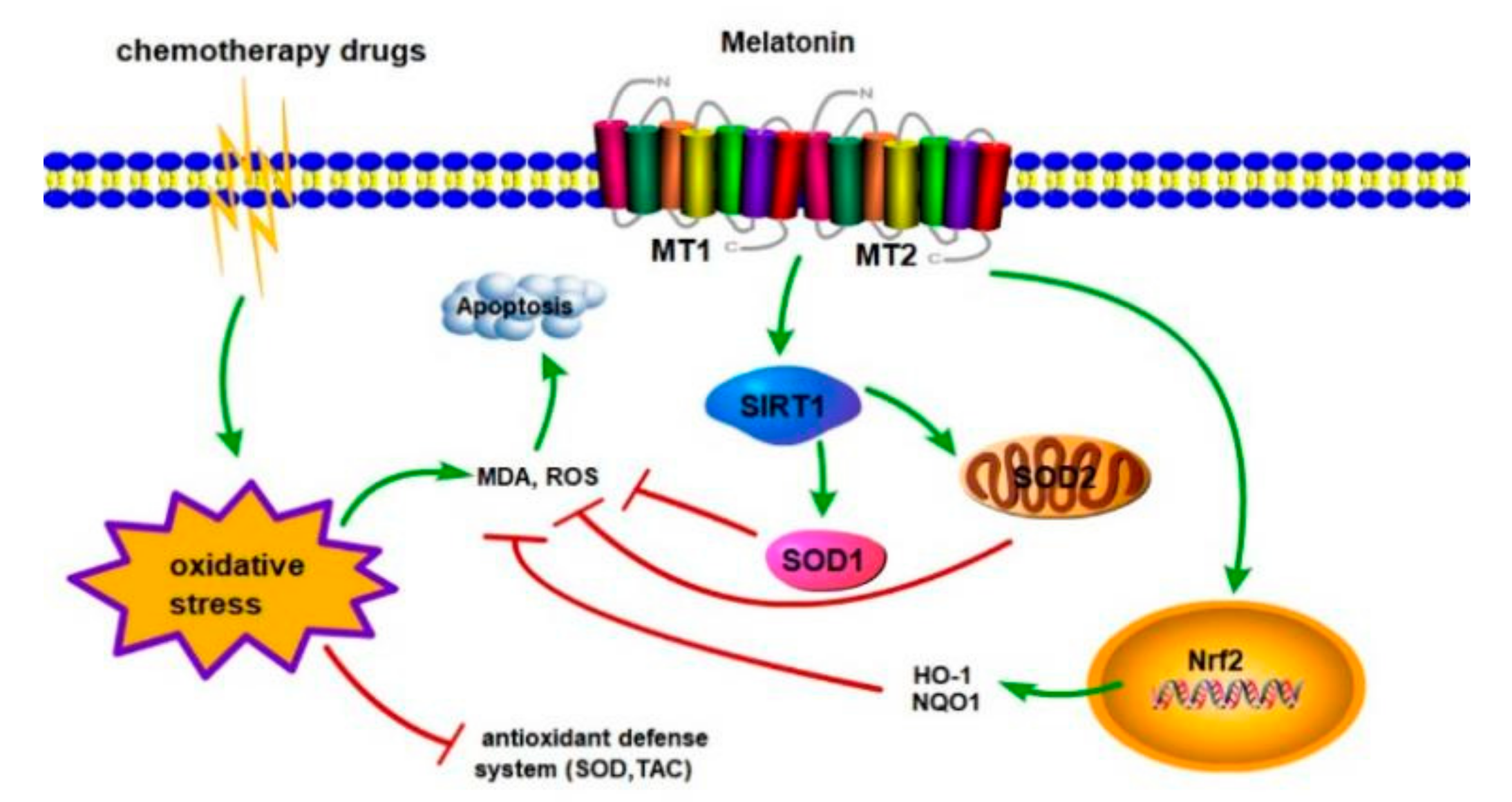
3.11. Melatonin Protects Leydig Cells from Cisplatin-Induced Oxidative Damage by Activating the MT1/MT2-Mediated SIRT1/Nrf2 Signaling Pathway
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Zhang, K.; Weng, H.; Yang, J.; Wu, C. Protective effect of Liuwei Dihuang Pill on cisplatin-induced reproductive toxicity and genotoxicity in male mice. J. Ethnopharmacol. 2020, 247, 112269. [Google Scholar] [CrossRef] [PubMed]
- Nna, V.U.; Ujah, G.A.; Suleiman, J.B.; Mohamed, M.; Nwokocha, C.; Akpan, T.J.; Ekuma, H.C.; Fubara, V.V.; Kekung-Asu, C.B.; Osim, E.E. Tert-butylhydroquinone preserve testicular steroidogenesis and spermatogenesis in cisplatin-intoxicated rats by targeting oxidative stress, inflammation and apoptosis. Toxicology 2020, 441, 152528. [Google Scholar] [CrossRef] [PubMed]
- Bandak, M.; Jørgensen, N.; Juul, A.; Lauritsen, J.; Kier, M.G.G.; Mortensen, M.S.; Daugaard, G. Longitudinal changes in serum levels of testosterone and luteinizing hormone in testicular cancer patients after orchiectomy alone or bleomycin, etoposide, and cisplatin. Eur. Urol. Focus 2018, 4, 591–598. [Google Scholar] [CrossRef]
- Fung, C.; Dinh, P., Jr.; Ardeshir-Rouhani-Fard, S.; Schaffer, K.; Fossa, S.D.; Travis, L.B. Toxicities associated with cisplatin-based chemotherapy and radiotherapy in long-term testicular cancer survivors. Adv. Urol. 2018, 2018, 8671832. [Google Scholar] [CrossRef] [PubMed]
- Shati, A.A. Resveratrol improves sperm parameter and testicular apoptosis in cisplatin-treated rats: Effects on ERK1/2, JNK, and Akt pathways. Syst. Biol. Reprod. Med. 2019, 65, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Bhartiya, D.; Sriraman, K.; Patel, H.; Manjramkar, D.; Bakshi, G.; Dhamankar, V.; Kurkure, P. Quiescent very small embryonic-like stem cells resist oncotherapy and can restore spermatogenesis in germ cell depleted mammalian testis. Stem Cells Dev. 2013, 23, 1428. [Google Scholar] [CrossRef] [PubMed]
- Barak, S. Fertility preservation in male patients with cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2019, 55, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Lackner, J.E.; Märk, I.; Schatzl, G.; Marberger, M.; Kratzik, C. Hypogonadism and androgen deficiency symptoms in testicular cancer survivors. Urology 2007, 69, 754–758. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Couture, R.; Touaibia, M.; Martin, L.J. Transcriptome modulation following administration of luteolin to bleomycin-etoposide-cisplatin chemotherapy on rat LC540 tumor Leydig cells. Andrologia 2021, 53, e13960. [Google Scholar] [CrossRef]
- Zhao, L.; Xiao, Y.; Li, C.; Zhang, J.; Zhang, Y.; Wu, M.; Ma, T.; Yang, L.; Wang, X.; Jiang, H.; et al. Zearalenone perturbs the circadian clock and inhibits testosterone synthesis in mouse Leydig cells. J. Toxicol. Environ. Health A 2021, 84, 112–124. [Google Scholar] [CrossRef]
- Hennies, S.; Wolff, H.A.; Jung, K.; Rave-Fränk, M.; Gaedcke, J.; Ghadimi, M.; Hess, C.F.; Becker, H.; Hermann, R.M.; Christiansen, H.J.S.U.O. Testicular radiation dose after multimodal curative therapy for locally advanced rectal cancer. Strahlenther Onkol. 2012, 188, 926–932. [Google Scholar] [CrossRef]
- Riddell, I.A. Cisplatin and oxaliplatin: Our current understanding of their actions. Met. Ions. Life Sci. 2018, 18, 1–42. [Google Scholar] [CrossRef]
- Tharmalingam, M.D.; Matilionyte, G.; Wallace, W.H.B.; Stukenborg, J.B.; Jahnukainen, K.; Oliver, E.; Goriely, A.; Lane, S.; Guo, J.; Cairns, B.; et al. Cisplatin and carboplatin result in similar gonadotoxicity in immature human testis with implications for fertility preservation in childhood cancer. BMC Med. 2020, 18, 374. [Google Scholar] [CrossRef]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. Correction: The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton. Trans. 2018, 47, 7848. [Google Scholar] [CrossRef]
- Wang, T.E.; Lai, Y.H.; Yang, K.C.; Lin, S.J.; Chen, C.L.; Tsai, P.S. Counteracting Cisplatin-Induced Testicular Damages by Natural Polyphenol Constituent Honokiol. Antioxidants 2020, 9, 723. [Google Scholar] [CrossRef]
- Altındağ, F.; Meydan, İ. Evaluation of protective effects of gallic acid on cisplatin-induced testicular and epididymal damage. Andrologia 2021, 53, e14189. [Google Scholar] [CrossRef]
- Kohsaka, T.; Minagawa, I.; Morimoto, M.; Yoshida, T.; Sasanami, T.; Yoneda, Y.; Ikegaya, N.; Sasada, H. Efficacy of relaxin for cisplatin-induced testicular dysfunction and epididymal spermatotoxicity. Basic Clin. Androl. 2020, 30, 3. [Google Scholar] [CrossRef]
- Eren, H.; Mercantepe, T.; Tumkaya, L.; Mercantepe, F.; Dil, E.; Horsanali, M.O.; Yilmaz, A. Evaluation of the protective effects of amifostine and melatonin against cisplatin induced testis injury via oxidative stress and apoptosis in rats. Exp. Mol. Pathol. 2020, 112, 104324. [Google Scholar] [CrossRef]
- Madhu, P.; Reddy, K.P.; Reddy, P.S. Role of melatonin in mitigating chemotherapy-induced testicular dysfunction in Wistar rats. Drug Chem. Toxicol. 2016, 39, 137–146. [Google Scholar] [CrossRef]
- Ilbey, Y.O.; Ozbek, E.; Simsek, A.; Otunctemur, A.; Cekmen, M.; Somay, A. Potential chemoprotective effect of melatonin in cyclophosphamide- and cisplatin-induced testicular damage in rats. Fertil. Steril. 2009, 92, 1124–1132. [Google Scholar] [CrossRef]
- Haghi-Aminjan, H.; Asghari, M.H.; Farhood, B.; Rahimifard, M.; Hashemi Goradel, N.; Abdollahi, M. The role of melatonin on chemotherapy-induced reproductive toxicity. J. Pharm. Pharmacol. 2018, 70, 291–306. [Google Scholar] [CrossRef] [Green Version]
- Balmik, A.A.; Chinnathambi, S. Multi-Faceted Role of Melatonin in Neuroprotection and Amelioration of Tau Aggregates in Alzheimer’s Disease. J. Alzheimers Dis. 2018, 62, 1481–1493. [Google Scholar] [CrossRef]
- Barberino, R.S.; Menezes, V.G.; Ribeiro, A.; Palheta, R.C., Jr.; Jiang, X.; Smitz, J.E.J.; Matos, M.H.T. Melatonin protects against cisplatin-induced ovarian damage in mice via the MT1 receptor and antioxidant activity. Biol. Reprod. 2017, 96, 1244–1255. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.; Lee, O.H.; Lee, Y.; Yoon, H.; Chang, E.M.; Park, M.; Lee, J.W.; Hong, K.; Kim, J.O.; Kim, N.K.; et al. Melatonin prevents cisplatin-induced primordial follicle loss via suppression of PTEN/AKT/FOXO3a pathway activation in the mouse ovary. J. Pineal Res. 2016, 60, 336–347. [Google Scholar] [CrossRef]
- Li, B.; He, X.; Zhuang, M.; Niu, B.; Wu, C.; Mu, H.; Tang, F.; Cui, Y.; Liu, W.; Zhao, B.; et al. Melatonin Ameliorates Busulfan-Induced Spermatogonial Stem Cell Oxidative Apoptosis in Mouse Testes. Antioxid Redox Signal 2018, 28, 385–400. [Google Scholar] [CrossRef]
- Ateşşahin, A.; Sahna, E.; Türk, G.; Ceribaşi, A.O.; Yilmaz, S.; Yüce, A.; Bulmuş, O. Chemoprotective effect of melatonin against cisplatin-induced testicular toxicity in rats. J. Pineal Res. 2006, 41, 21–27. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhang, Y. Melatonin: A well-documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef]
- Cecon, E.; Oishi, A.; Jockers, R. Melatonin receptors: Molecular pharmacology and signalling in the context of system bias. Br. J. Pharmacol. 2018, 175, 3263–3280. [Google Scholar] [CrossRef] [Green Version]
- Mayo, J.C.; Sainz, R.M.; González-Menéndez, P.; Hevia, D.; Cernuda-Cernuda, R. Melatonin transport into mitochondria. Cell. Mol. Life Sci. 2017, 74, 3927–3940. [Google Scholar] [CrossRef]
- Levoye, A.; Dam, J.; Ayoub, M.A.; Guillaume, J.L.; Couturier, C.; Delagrange, P.; Jockers, R. The orphan GPR50 receptor specifically inhibits MT1 melatonin receptor function through heterodimerization. EMBO J. 2006, 25, 3012–3023. [Google Scholar] [CrossRef]
- Baba, K.; Benleulmi-Chaachoua, A.; Journé, A.S.; Kamal, M.; Guillaume, J.L.; Dussaud, S.; Gbahou, F.; Yettou, K.; Liu, C.; Contreras-Alcantara, S.; et al. Heteromeric MT1/MT2 melatonin receptors modulate photoreceptor function. Sci. Signal 2013, 6, ra89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wu, X.; Zhao, S.; Zhang, Y.; Ma, H.; Yang, Z.; Yang, W.; Zhao, C.; Wang, L.; Zhang, Q. Melatonin receptor depletion suppressed hCG-induced testosterone expression in mouse Leydig cells. Cell. Mol. Biol. Lett. 2019, 24, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathya, M.; Moorthi, P.; Premkumar, P.; Kandasamy, M.; Jayachandran, K.S.; Anusuyadevi, M. Resveratrol Intervenes Cholesterol- and Isoprenoid-Mediated Amyloidogenic Processing of AβPP in Familial Alzheimer’s Disease. J. Alzheimers Dis. 2017, 60, S3–S23. [Google Scholar] [CrossRef]
- Wang, N.; Wang, L.; Yang, J.; Wang, Z.; Cheng, L. Quercetin promotes osteogenic differentiation and antioxidant responses of mouse bone mesenchymal stem cells through activation of the AMPK/SIRT1 signaling pathway. Phytother. Res. 2021, 35, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Z.; Wang, J.; Lv, D.; Zhu, T.; Wang, F.; Tian, X.; Yao, Y.; Ji, P.; Liu, G. Melatonin regulates the activities of ovary and delays the fertility decline in female animals via MT1/AMPK pathway. J. Pineal Res. 2019, 66, e12550. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Shi, Z.; Peng, Z.; Zhao, C.; Zhang, Y.; Wang, Z.; Li, X.; Liu, G.; Li, X. Acetoacetate induces hepatocytes apoptosis by the ROS-mediated MAPKs pathway in ketotic cows. J. Cell. Physiol. 2017, 232, 3296–3308. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Li, S.; Lv, Y.; Yang, D.; Li, J.; Yang, Q.; Wu, P.; Lv, Z.; Zhang, Z. Dietary melatonin attenuates chromium-induced lung injury via activating the Sirt1/Pgc-1α/Nrf2 pathway. Food Funct. 2019, 10, 5555–5565. [Google Scholar] [CrossRef]
- Liu, B.; Yu, H.; Baiyun, R.; Lu, J.; Li, S.; Bing, Q.; Zhang, X.; Zhang, Z. Protective effects of dietary luteolin against mercuric chloride-induced lung injury in mice: Involvement of AKT/Nrf2 and NF-κB pathways. Food Chem. Toxicol. 2018, 113, 296–302. [Google Scholar] [CrossRef]
- Kim, E.H.; Ridlo, M.R.; Lee, B.C.; Kim, G.A. Melatonin-Nrf2 Signaling Activates Peroxisomal Activities in Porcine Cumulus Cell-Oocyte Complexes. Antioxidants 2020, 9, 1080. [Google Scholar] [CrossRef]
- Fang, J.; Yan, Y.; Teng, X.; Wen, X.; Li, N.; Peng, S.; Liu, W.; Donadeu, F.X.; Zhao, S.; Hua, J. Melatonin prevents senescence of canine adipose-derived mesenchymal stem cells through activating NRF2 and inhibiting ER stress. Aging 2018, 10, 2954–2972. [Google Scholar] [CrossRef]
- Arioz, B.I.; Tastan, B.; Tarakcioglu, E.; Tufekci, K.U.; Olcum, M.; Ersoy, N.; Bagriyanik, A.; Genc, K.; Genc, S. Melatonin Attenuates LPS-Induced Acute Depressive-Like Behaviors and Microglial NLRP3 Inflammasome Activation Through the SIRT1/Nrf2 Pathway. Front. Immunol. 2019, 10, 1511. [Google Scholar] [CrossRef]
- Ali, T.; Hao, Q.; Ullah, N.; Rahman, S.U.; Shah, F.A.; He, K.; Zheng, C.; Li, W.; Murtaza, I.; Li, Y.; et al. Melatonin Act as an Antidepressant via Attenuation of Neuroinflammation by Targeting Sirt1/Nrf2/HO-1 Signaling. Front. Mol. Neurosci. 2020, 13, 96. [Google Scholar] [CrossRef]
- Khaksary Mahabady, M.; Najafzadeh Varzi, H.; Bakhtiari, E. The effects of cyclophosphamide, melatonin and carvedilol on neural tube and skeletal system of mice fetuses in prenatal period. Ann. Anat. 2011, 193, 459–465. [Google Scholar] [CrossRef]
- Wen, B.; Zhou, R.; Feng, Q.; Wang, Q.; Wang, J.; Liu, S. IQuant: An automated pipeline for quantitative proteomics based upon isobaric tags. Proteomics 2014, 14, 2280–2285. [Google Scholar] [CrossRef]
- Savitski, M.M.; Wilhelm, M.; Hahne, H.; Kuster, B.; Bantscheff, M. A Scalable Approach for Protein False Discovery Rate Estimation in Large Proteomic Data Sets. Mol. Cell. Proteomics 2015, 14, 2394–2404. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.T.; Cao, M.D.; Yu, R.L.; Shi, H.Y.; Yan, W.J.; Liu, J.G.; Pan, C.; Sun, J.; Wei, Q.Y.; Wang, D.Y.; et al. Leukotriene A(4) Hydrolase Is a Candidate Predictive Biomarker for Successful Allergen Immunotherapy. Front. Immunol. 2020, 11, 559746. [Google Scholar] [CrossRef]
- Klinefelter, G.R.; Hall, P.F.; Ewing, L.L. Effect of luteinizing hormone deprivation in situ on steroidogenesis of rat Leydig cells purified by a multistep procedure. Biol. Reprod. 1987, 36, 769–783. [Google Scholar] [CrossRef] [Green Version]
- Chemes, H.; Cigorraga, S.; Bergadá, C.; Schteingart, H.; Rey, R.; Pellizzari, E. Isolation of human Leydig cell mesenchymal precursors from patients with the androgen insensitivity syndrome: Testosterone production and response to human chorionic gonadotropin stimulation in culture. Biol. Reprod. 1992, 46, 793–801. [Google Scholar] [CrossRef] [Green Version]
- Osanai, K.; Kobayashi, Y.; Otsu, M.; Izawa, T.; Sakai, K.; Iwashita, M. Ramelteon, a selective MT1/MT2 receptor agonist, suppresses the proliferation and invasiveness of endometrial cancer cells. Hum. Cell 2017, 30, 209–215. [Google Scholar] [CrossRef]
- Estaras, M.; Marchena, A.M.; Fernandez-Bermejo, M.; Mateos, J.M.; Vara, D.; Roncero, V.; Salido, G.M.; Gonzalez, A. The melatonin receptor antagonist luzindole induces the activation of cellular stress responses and decreases viability of rat pancreatic stellate cells. J. Appl. Toxicol. 2020, 40, 1554–1565. [Google Scholar] [CrossRef]
- Muggia, F.M.; Bonetti, A.; Hoeschele, J.D.; Rozencweig, M.; Howell, S.B. Platinum Antitumor Complexes: 50 Years Since Barnett Rosenberg’s Discovery. J. Clin. Oncol. 2015, 33, 4219–4226. [Google Scholar] [CrossRef]
- Esteban-Fernández, D.; Verdaguer, J.M.; Ramírez-Camacho, R.; Palacios, M.A.; Gómez-Gómez, M.M. Accumulation, fractionation, and analysis of platinum in toxicologically affected tissues after cisplatin, oxaliplatin, and carboplatin administration. J. Anal. Toxicol. 2008, 32, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, E.A.; Omar, H.M.; Elghaffar, S.; Ragb, S.M.; Nasser, A.Y. The antioxidant activity of vitamin C, DPPD and L-cysteine against Cisplatin-induced testicular oxidative damage in rats. Food Chem. Toxicol. 2011, 49, 1115–1121. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Moradi, M.; Goodarzi, N.; Faramarzi, A.; Cheraghi, H.; Jalili, C. Melatonin protects rats testes against bleomycin, etoposide, and cisplatin-induced toxicity via mitigating nitro-oxidative stress and apoptosis. Biomed. Pharmacother. 2021, 138, 111481. [Google Scholar] [CrossRef]
- Al-Bader, M.; Kilarkaje, N. Effects of bleomycin, etoposide and cisplatin treatment on Leydig cell structure and transcription of steroidogenic enzymes in rat testis. Eur. J. Pharmacol. 2015, 747, 150–159. [Google Scholar] [CrossRef]
- Kisielewska, K.; Rytelewska, E.; Gudelska, M.; Kiezun, M.; Dobrzyn, K.; Bogus-Nowakowska, K.; Kaminska, B.; Smolinska, N.; Kaminski, T. Expression of chemerin receptors CMKLR1, GPR1 and CCRL2 in the porcine pituitary during the oestrous cycle and early pregnancy and the effect of chemerin on MAPK/Erk1/2, Akt and AMPK signalling pathways. Theriogenology 2020, 157, 181–198. [Google Scholar] [CrossRef]
- Wang, L.; He, Y.; Li, Y.; Pei, C.; Olatunji, O.J.; Tang, J.; Famurewa, A.C.; Wang, H.; Yan, B. Protective Effects of Nucleosides-Rich Extract from Cordyceps cicadae against Cisplatin Induced Testicular Damage. Chem. Biodivers. 2020, 17, e2000671. [Google Scholar] [CrossRef]
- Aly, H.A.A.; Eid, B.G. Cisplatin induced testicular damage through mitochondria mediated apoptosis, inflammation and oxidative stress in rats: Impact of resveratrol. Endocr. J. 2020, 67, 969–980. [Google Scholar] [CrossRef]
- Afsar, T.; Razak, S.; Khan, M.R.; Almajwal, A. Acacia hydaspica ethyl acetate extract protects against cisplatin-induced DNA damage, oxidative stress and testicular injuries in adult male rats. BMC Cancer 2017, 17, 883. [Google Scholar] [CrossRef] [Green Version]
- Hosen, M.B.; Islam, M.R.; Begum, F.; Kabir, Y.; Howlader, M.Z.H. Oxidative stress induced sperm DNA damage, a possible reason for male infertility. Iran J. Reprod. Med. 2015, 13, 525–532. [Google Scholar] [PubMed]
- Ghasemi, F.; Faghani, M.; Khajehjahromi, S.; Bahadori, M.; Nasiri E, E.; Hemadi, M. Effect of Melatonin on Proliferative Activity and Apoptosis in Spermatogenic Cells in Mouse under Chemotherapy. J. Reprod. Contracept. 2010, 21, 79–94. [Google Scholar] [CrossRef]
- Cui, Y.; Ren, L.; Li, B.; Fang, J.; Zhai, Y.; He, X.; Du, E.; Miao, Y.; Hua, J.; Peng, S.J.C.P.; et al. Melatonin Relieves Busulfan-Induced Spermatogonial Stem Cell Apoptosis of Mouse Testis by Inhibiting Endoplasmic Reticulum Stress. Cell Physiol. Biochem. 2017, 44, 2407–2421. [Google Scholar] [CrossRef] [PubMed]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Chen, Y.T.; Yang, C.C.; Shao, P.L.; Huang, C.R.; Yip, H.K. Melatonin-mediated downregulation of ZNF746 suppresses bladder tumorigenesis mainly through inhibiting the AKT-MMP-9 signaling pathway. J. Pineal Res. 2019, 66, e12536. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.; Dauchy, R.T.; Hoffman, A.E.; Pointer, D.; Frasch, T.; Blask, D.E.; Hill, S.M. Epigenetic inhibition of the tumor suppressor ARHI by light at night-induced circadian melatonin disruption mediates STAT3-driven paclitaxel resistance in breast cancer. J. Pineal Res. 2019, 67, e12586. [Google Scholar] [CrossRef]
- Ma, Z.; Xu, L.; Liu, D.; Zhang, X.; Di, S.; Li, W.; Zhang, J.; Reiter, R.J.; Han, J.; Li, X.; et al. Utilizing Melatonin to Alleviate Side Effects of Chemotherapy: A Potentially Good Partner for Treating Cancer with Ageing. Oxid. Med. Cell. Longev. 2020, 2020, 6841581. [Google Scholar] [CrossRef]
- Ivanov, D.; Mazzoccoli, G.; Anderson, G.; Linkova, N.; Dyatlova, A.; Mironova, E.; Polyakova, V.; Kvetnoy, I.; Evsyukova, I.; Carbone, A.; et al. Melatonin, Its Beneficial Effects on Embryogenesis from Mitigating Oxidative Stress to Regulating Gene Expression. Int. J. Mol. Sci. 2021, 22, 5885. [Google Scholar] [CrossRef]
- Zonta, Y.R.; Martinez, M.; Camargo, I.C.; Domeniconi, R.F.; Lupi Júnior, L.A.; Pinheiro, P.F.; Reiter, R.J.; Martinez, F.E.; Chuffa, L.G. Melatonin Reduces Angiogenesis in Serous Papillary Ovarian Carcinoma of Ethanol-Preferring Rats. Int. J. Mol. Sci. 2017, 18, 763. [Google Scholar] [CrossRef]
- Xu, D.; Liu, L.; Zhao, Y.; Yang, L.; Cheng, J.; Hua, R.; Zhang, Z.; Li, Q. Melatonin protects mouse testes from palmitic acid-induced lipotoxicity by attenuating oxidative stress and DNA damage in a SIRT1-dependent manner. J. Pineal Res. 2020, 69, e12690. [Google Scholar] [CrossRef]
- Hayes, J.D.; Dinkova-Kostova, A.T. The Nrf2 regulatory network provides an interface between redox and intermediary metabolism. Trends Biochem. Sci. 2014, 39, 199–218. [Google Scholar] [CrossRef]
- Kleszczyński, K.; Zillikens, D.; Fischer, T.W. Melatonin enhances mitochondrial ATP synthesis, reduces reactive oxygen species formation, and mediates translocation of the nuclear erythroid 2-related factor 2 resulting in activation of phase-2 antioxidant enzymes (γ-GCS, HO-1, NQO1) in ultraviolet radiation-treated normal human epidermal keratinocytes (NHEK). J. Pineal Res. 2016, 61, 187–197. [Google Scholar] [CrossRef]
- Dong, Y.; Zhao, J.; Zhu, Q.; Liu, H.; Wang, J.; Lu, W. Melatonin inhibits the apoptosis of rooster Leydig cells by suppressing oxidative stress via AKT-Nrf2 pathway activation. Free Radic. Biol. Med. 2020, 160, 1–12. [Google Scholar] [CrossRef]
- Favero, G.; Franceschetti, L.; Bonomini, F.; Rodella, L.F.; Rezzani, R. Melatonin as an Anti-Inflammatory Agent Modulating Inflammasome Activation. Int. J. Endocrinol. 2017, 2017, 1835195. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.; Song, J.; Yu, Z.; Cui, K.; Ruan, Y.; Wang, T.; Yang, J.; Wang, S.; Liu, J. Melatonin Treatment Ameliorates Hyperhomocysteinemia-Induced Impairment of Erectile Function in a Rat Model. J. Sex. Med. 2019, 16, 1506–1517. [Google Scholar] [CrossRef]
- Guo, Y.; Sun, J.; Li, T.; Zhang, Q.; Bu, S.; Wang, Q.; Lai, D. Melatonin ameliorates restraint stress-induced oxidative stress and apoptosis in testicular cells via NF-κB/iNOS and Nrf2/ HO-1 signaling pathway. Sci. Rep. 2017, 7, 9599. [Google Scholar] [CrossRef]
- Shao, Y.; Zhou, Q.; Xue, J.; Shen, B.; Ji, J.; Wei, Z. Melatonin ameliorates cisplatin-induced asthenozoospermia via reducing oxidative stress. J. Mens. Health 2022, 18, 16. [Google Scholar] [CrossRef]
- Xu, G.; Zhao, J.; Liu, H.; Wang, J.; Lu, W. Melatonin Inhibits Apoptosis and Oxidative Stress of Mouse Leydig Cells via a SIRT1-Dependent Mechanism. Molecules 2019, 24, 3084. [Google Scholar] [CrossRef] [Green Version]





















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Fang, Y.; Tang, D.; Xu, X.; Zhu, X.; Wu, S.; Yu, H.; Cheng, H.; Luo, T.; Shen, Q.; et al. Activation of MT1/MT2 to Protect Testes and Leydig Cells against Cisplatin-Induced Oxidative Stress through the SIRT1/Nrf2 Signaling Pathway. Cells 2022, 11, 1690. https://doi.org/10.3390/cells11101690
Zhang J, Fang Y, Tang D, Xu X, Zhu X, Wu S, Yu H, Cheng H, Luo T, Shen Q, et al. Activation of MT1/MT2 to Protect Testes and Leydig Cells against Cisplatin-Induced Oxidative Stress through the SIRT1/Nrf2 Signaling Pathway. Cells. 2022; 11(10):1690. https://doi.org/10.3390/cells11101690
Chicago/Turabian StyleZhang, Junqiang, Yuan Fang, Dongdong Tang, Xingyu Xu, Xiaoqian Zhu, Shusheng Wu, Hui Yu, Huiru Cheng, Ting Luo, Qunshan Shen, and et al. 2022. "Activation of MT1/MT2 to Protect Testes and Leydig Cells against Cisplatin-Induced Oxidative Stress through the SIRT1/Nrf2 Signaling Pathway" Cells 11, no. 10: 1690. https://doi.org/10.3390/cells11101690
APA StyleZhang, J., Fang, Y., Tang, D., Xu, X., Zhu, X., Wu, S., Yu, H., Cheng, H., Luo, T., Shen, Q., Gao, Y., Ma, C., Liu, Y., Wei, Z., Chen, X., Tao, F., He, X., & Cao, Y. (2022). Activation of MT1/MT2 to Protect Testes and Leydig Cells against Cisplatin-Induced Oxidative Stress through the SIRT1/Nrf2 Signaling Pathway. Cells, 11(10), 1690. https://doi.org/10.3390/cells11101690