
Could Experimental Inflammation Provide Better Understanding of Migraines?


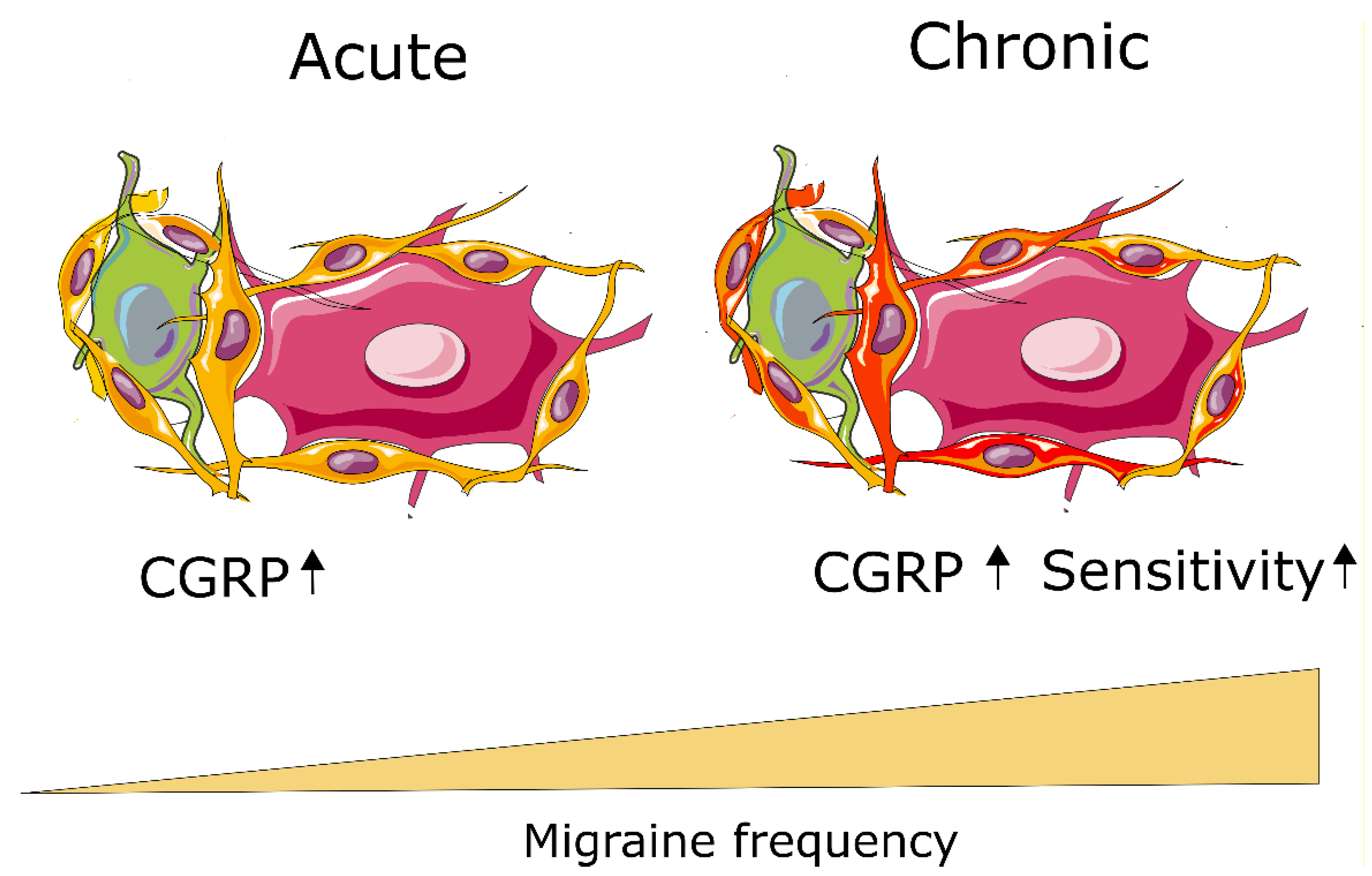{kind=link}
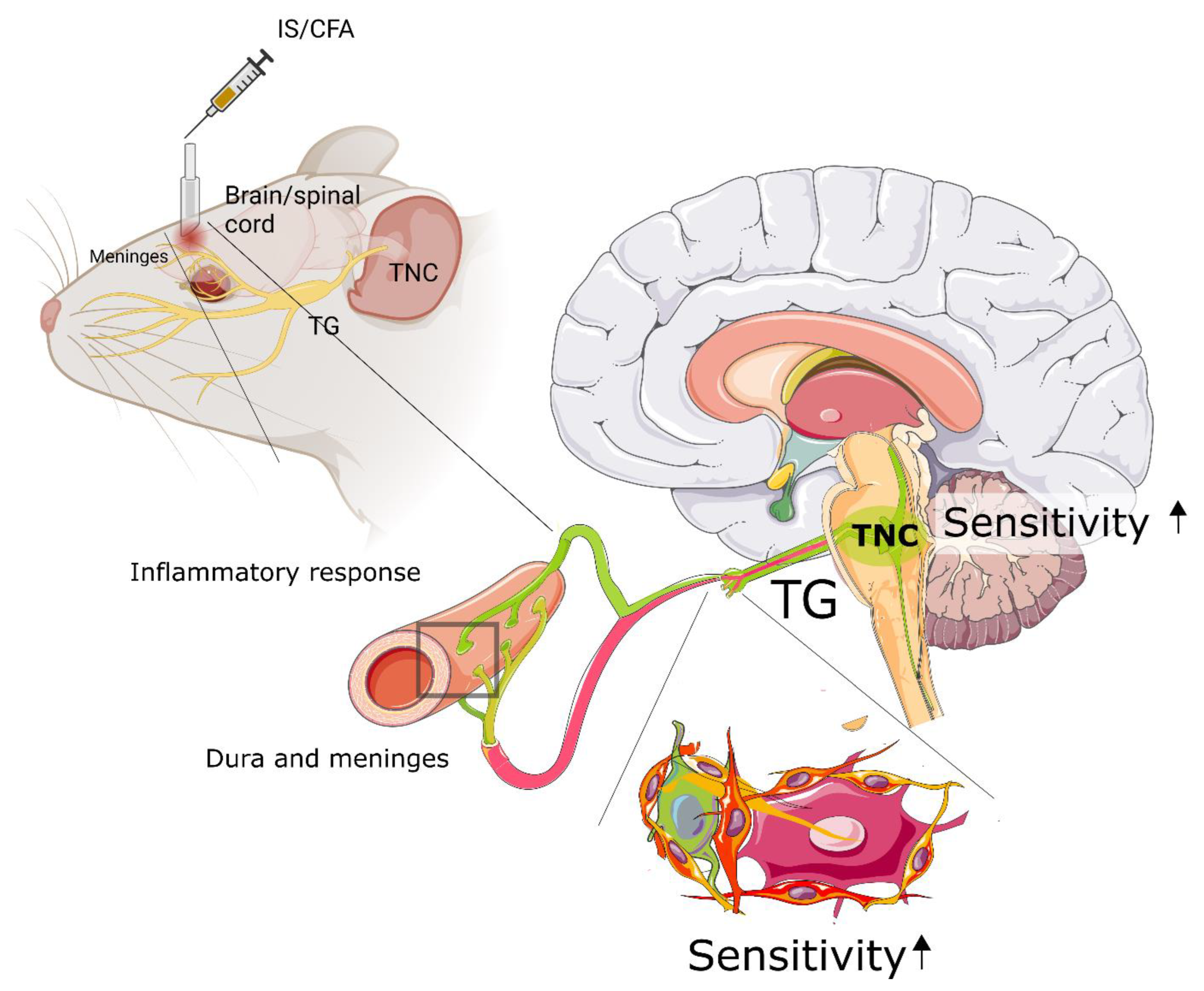{kind=link}
Abstract
:1. Pathophysiology of Migraine
2. Inflammation in Migraine
3. The Inflammatory Soup (IS) Model and Its Variants
4. Dural Activation by Complete Freund’s Adjuvant (CFA)
5. IS vs. CFA
6. Evaluating Outcomes in Animal Inflammation Models
7. Concluding on CGRP and Inflammation—A Perspective
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bigal, M.E.; Lipton, R.B. The epidemiology, burden, and comorbidities of migraine. Neurol. Clin. 2009, 27, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Ashina, M.; Katsarava, Z.; Do, T.P.; Buse, D.C.; Pozo-Rosich, P.; Özge, A.; Krymchantowski, A.V.; Lebedeva, E.R.; Ravishankar, K.; Yu, S.; et al. Migraine: Epidemiology and systems of care. Lancet 2021, 397, 1485–1495. [Google Scholar] [CrossRef]
- Bron, C.; Sutherland, H.G.; Griffiths, L.R. Exploring the Hereditary Nature of Migraine. Neuropsychiatr. Dis. Treat. 2021, 17, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Evans, R.W. Wolff’s "Headache and Other Head Pain": Happy 70th Anniversary! Headache 2019, 59, 443. [Google Scholar] [CrossRef]
- Haanes, K.A.; Edvinsson, L. Pathophysiological Mechanisms in Migraine and the Identification of New Therapeutic Targets. CNS Drugs 2019, 33, 525–537. [Google Scholar] [CrossRef]
- Matsuda, M.; Huh, Y.; Ji, R.R. Roles of inflammation, neurogenic inflammation, and neuroinflammation in pain. J. Anesth. 2019, 33, 131–139. [Google Scholar] [CrossRef]
- Riesco, N.; Cernuda-Morollón, E.; Pascual, J. Neuropeptides as a Marker for Chronic Headache. Curr. Pain Headache Rep. 2017, 21, 18. [Google Scholar] [CrossRef]
- Torres-Ferrus, M.; Ursitti, F.; Alpuente, A.; Brunello, F.; Chiappino, D.; Vries, T.D.; di Marco, S.; Ferlisi, S.; Guerritore, L.; Gonzalez-Garcia, N.; et al. School of Advanced Studies of European Headache, From transformation to chronification of migraine: Pathophysiological and clinical aspects. J. Headache Pain 2020, 21, 42. [Google Scholar] [CrossRef]
- Dodick, D.W. A Phase-by-Phase Review of Migraine Pathophysiology. Headache 2018, 58, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Bernstein, C.; Burstein, R. Sensitization of the trigeminovascular pathway: Perspective and implications to migraine pathophysiology. J. Clin. Neurol. 2012, 8, 89–99. [Google Scholar] [CrossRef] [Green Version]
- Ashina, M.; Hansen, J.M.; O, A.D.B.; Olesen, J. Human models of migraine—short-term pain for long-term gain. Nat. Rev. Neurol. 2017, 13, 713–724. [Google Scholar] [CrossRef]
- Al-Karagholi, M.A.; Ghanizada, H.; Nielsen, C.A.W.; Hougaard, A.; Ashina, M. Opening of ATP sensitive potassium channels causes migraine attacks with aura. Brain 2021, 144, 2322–2332. [Google Scholar] [CrossRef] [PubMed]
- Schulte, L.H.; May, A. The migraine generator revisited: Continuous scanning of the migraine cycle over 30 days and three spontaneous attacks. Brain 2016, 139, 1987–1993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikkeri, N.S.; Nagalli, S. Migraine with Aura. Treasure Island (FL); StatPearls Publishing LLC: Treasure Island, FL, USA, 2022.
- Borgdorff, P. Arguments against the role of cortical spreading depression in migraine. Neurol. Res. 2018, 40, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Bolay, H.; Reuter, U.; Dunn, A.K.; Huang, Z.; Boas, D.A.; Moskowitz, M.A. Intrinsic brain activity triggers trigeminal meningeal afferents in a migraine model. Nat. Med. 2002, 8, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Levy, D.; Kainz, V.; Noseda, R.; Jakubowski, M.; Burstein, R. Activation of central trigeminovascular neurons by cortical spreading depression. Ann. Neurol. 2011, 69, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Goadsby, P.J.; Edvinsson, L.; Ekman, R. Vasoactive peptide release in the extracerebral circulation of humans during migraine headache. Ann. Neurol. 1990, 28, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine therapies—successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [Google Scholar] [CrossRef]
- Ramos, M.L.; Pascual, J. AMG 334 CGRP antibody for migraine: Time to celebrate? Lancet Neurol. 2016, 15, 347–349. [Google Scholar] [CrossRef]
- Negro, A.; Martelletti, P. Gepants for the treatment of migraine. Expert. Opin. Investig. Drugs 2019, 28, 555–567. [Google Scholar] [CrossRef]
- Russo, A.F. Calcitonin gene-related peptide (CGRP): A new target for migraine. Annu. Rev. Pharmacol. Toxicol. 2015, 55, 533–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haanes, K.A.; Edvinsson, L. Hyperpolarization through ATP-sensitive potassium channels; relevance to migraine pathology. Brain 2020, 143, e13. [Google Scholar] [CrossRef] [PubMed]
- Mulderry, P.K.; Ghatei, M.A.; Bishop, A.E.; Allen, Y.S.; Polak, J.M.; Bloom, S.R. Distribution and chromatographic characterisation of CGRP-like immunoreactivity in the brain and gut of the rat. Regul. Pept. 1985, 12, 133–143. [Google Scholar] [CrossRef]
- Warfvinge, K.; Edvinsson, L. Distribution of CGRP and CGRP receptor components in the rat brain. Cephalalgia 2019, 39, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, J.C.; Reducha, P.V.; Sheykhzade, M.; Warfvinge, K.; Haanes, K.A.; Edvinsson, L. Neurokinins and their receptors in the rat trigeminal system: Differential localization and release with implications for migraine pain. Mol. Pain 2021, 17, 17448069211059400. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, J.C.A.; Warfvinge, K.; Krause, D.N.; Blixt, F.W.; Sheykhzade, M.; Edvinsson, L.; Haanes, K.A. C-fibers may modulate adjacent Adelta-fibers through axon-axon CGRP signaling at nodes of Ranvier in the trigeminal system. J. Headache Pain 2019, 20, 105. [Google Scholar] [CrossRef] [Green Version]
- Walker, H.K. Cranial Nerve V, Clinical Methods: The Trigeminal Nerve, the History, Physical, and Laboratory Examinations; Walker, H.K., Hall, W.D., Hurst, J.W., Eds.; Butterworth Publishers: Boston, MA, USA, 1990. [Google Scholar]
- Cottrell, G.S. CGRP Receptor Signalling Pathways. Handb. Exp. Pharmacol. 2019, 255, 37–64. [Google Scholar]
- Moskowitz, M.A. The neurobiology of vascular head pain. Ann. Neurol. 1984, 16, 157–168. [Google Scholar] [CrossRef]
- Markowitz, S.; Saito, K.; Moskowitz, M.A. Neurogenically mediated plasma extravasation in dura mater: Effect of ergot alkaloids. A possible mechanism of action in vascular headache. Cephalalgia 1988, 8, 83–91. [Google Scholar] [CrossRef]
- Shepherd, S.L.; Williamson, D.J.; Beer, M.S.; Hill, R.G.; Hargreaves, R.J. Differential effects of 5-HT1B/1D receptor agonists on neurogenic dural plasma extravasation and vasodilation in anaesthetized rats. Neuropharmacology 1997, 36, 525–533. [Google Scholar] [CrossRef]
- Buzzi, M.G.; Moskowitz, M.A. The antimigraine drug, sumatriptan (GR43175), selectively blocks neurogenic plasma extravasation from blood vessels in dura mater. Br. J. Pharmacol. 1990, 99, 202–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskowitz, M.A. Neurogenic versus vascular mechanisms of sumatriptan and ergot alkaloids in migraine. Trends Pharmacol. Sci. 1992, 13, 307–311. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Spanos, C.; Pang, X.; Alferes, L.; Ligris, K.; Letourneau, R.; Rozniecki, J.J.; Webster, E.; Chrousos, G.P. Stress-induced intracranial mast cell degranulation: A corticotropin-releasing hormone-mediated effect. Endocrinology 1995, 136, 5745–5750. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Donelan, J.; Kandere-Grzybowska, K.; Konstantinidou, A. The role of mast cells in migraine pathophysiology. Brain Res. Rev. 2005, 49, 65–76. [Google Scholar] [CrossRef]
- Dalkara, T.; Zervas, N.T.; Moskowitz, M.A. From spreading depression to the trigeminovascular system. Neurol. Sci. 2006, 27, S86–S90. [Google Scholar] [CrossRef]
- Levy, D. Endogenous mechanisms underlying the activation and sensitization of meningeal nociceptors: The role of immuno-vascular interactions and cortical spreading depression. Curr. Pain Headache Rep. 2012, 16, 270–277. [Google Scholar] [CrossRef]
- Johnson KW, B.H. Neurogenic Inflammatory Mechanisms; Lipincott Williams & Wilkins: Philadelphia, PA, USA, 2006. [Google Scholar]
- Lassen, L.H.; Ashina, M.; Christiansen, I.; Ulrich, V.; Olesen, J. Nitric oxide synthase inhibition in migraine. Lancet 1997, 349, 401–402. [Google Scholar] [CrossRef]
- Covelli, V.; Munno, I.; Pellegrino, N.M.; Altamura, M.; Decandia, P.; Marcuccio, C.; di Venere, A.; Jirillo, E. Are TNF-alpha and IL-1 beta relevant in the pathogenesis of migraine without aura? Acta Neurol. Napoli 1991, 13, 205–211. [Google Scholar]
- Perini, F.; D’Andrea, G.; Galloni, E.; Pignatelli, F.; Billo, G.; Alba, S.; Bussone, G.; Toso, V. Plasma cytokine levels in migraineurs and controls. Headache 2005, 45, 926–931. [Google Scholar] [CrossRef]
- Franceschini, A.; Vilotti, S.; Ferrari, M.D.; van den Maagdenberg, A.M.; Nistri, A.; Fabbretti, E. TNFalpha levels and macrophages expression reflect an inflammatory potential of trigeminal ganglia in a mouse model of familial hemiplegic migraine. PLoS ONE 2013, 8, e52394. [Google Scholar] [CrossRef]
- Tanure, M.T.; Gomez, R.S.; Hurtado, R.C.; Teixeira, A.L.; Domingues, R.B. Increased serum levels of brain-derived neurotropic factor during migraine attacks: A pilot study. J. Headache Pain 2010, 11, 427–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yucel, M.; Kotan, D.; Ciftci, G.G.; Ciftci, I.H.; Cikriklar, H.I. Serum levels of endocan, claudin-5 and cytokines in migraine. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 930–936. [Google Scholar] [PubMed]
- Sarchielli, P.; Alberti, A.; Baldi, A.; Coppola, F.; Rossi, C.; Pierguidi, L.; Floridi, A.; Calabresi, P. Proinflammatory cytokines, adhesion molecules, and lymphocyte integrin expression in the internal jugular blood of migraine patients without aura assessed ictally. Headache 2006, 46, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K. Does inflammation have a role in migraine? Nat. Rev. Neurol. 2019, 15, 483–490. [Google Scholar] [CrossRef]
- de Logu, F.; Landini, L.; Janal, M.N.; Puma, S.L.; de Cesaris, F.; Geppetti, P.; Nassini, R. Migraine-provoking substances evoke periorbital allodynia in mice. J. Headache Pain 2019, 20, 18. [Google Scholar] [CrossRef] [Green Version]
- Nakamura-Craig, M.; Gill, B.K. Effect of neurokinin A, substance P and calcitonin gene related peptide in peripheral hyperalgesia in the rat paw. Neurosci. Lett. 1991, 124, 49–51. [Google Scholar] [CrossRef]
- Birrell, G.J.; McQueen, D.S.; Iggo, A.; Coleman, R.A.; Grubb, B.D. PGI2-induced activation and sensitization of articular mechanonociceptors. Neurosci. Lett. 1991, 124, 5–8. [Google Scholar] [CrossRef]
- Wang, H.; Ehnert, C.; Brenner, G.J.; Woolf, C.J. Bradykinin and peripheral sensitization. Biol. Chem. 2006, 387, 11–14. [Google Scholar] [CrossRef]
- Schaible, H.G.; Schmidt, R.F. Excitation and sensitization of fine articular afferents from cat’s knee joint by prostaglandin E2. J. Physiol. 1988, 403, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Uddman, R.; Edvinsson, L.; Jansen, I.; Stiernholm, P.; Jensen, K.; Olesen, J.; Sundler, F. Peptide-containing nerve fibres in human extracranial tissue: A morphological basis for neuropeptide involvement in extracranial pain? Pain 1986, 27, 391–399. [Google Scholar] [CrossRef]
- Harriott, A.M.; Gold, M.S. Electrophysiological properties of dural afferents in the absence and presence of inflammatory mediators. J. Neurophysiol. 2009, 101, 3126–3134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harriott, A.M.; Scheff, N.N.; Gold, M.S. The complex actions of sumatriptan on rat dural afferents. Cephalalgia 2012, 32, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheff, N.N.; Gold, M.S. Sex differences in the inflammatory mediator-induced sensitization of dural afferents. J. Neurophysiol. 2011, 106, 1662–1668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oshinsky, M.L.; Gomonchareonsiri, S. Episodic dural stimulation in awake rats: A model for recurrent headache. Headache 2007, 47, 1026–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelmayer, R.M.; Vanderah, T.W.; Majuta, L.; Zhang, E.T.; Fioravanti, B.; de Felice, M.; Chichorro, J.G.; Ossipov, M.H.; King, T.; Lai, J.; et al. Medullary pain facilitating neurons mediate allodynia in headache-related pain. Ann. Neurol. 2009, 65, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Dong, X.; Xu, H.; Wan, Q.; Guo, Q.; Wang, J.; Xiao, H.; Yan, L. Microglial P2Y14 receptor contributes to central sensitization following repeated inflammatory dural stimulation. Brain Res. Bull. 2021, 177, 119–128. [Google Scholar] [CrossRef]
- Hu, G.; Zhang, M.; Su, M.; Zhang, Q.; Wu, H.; Wang, X.; Dong, Z.; Yu, S. Wider range of allodynia in a rat model of repeated dural nociception compared with infraorbital nerve chronic constriction injury. Neurosci. Lett. 2018, 666, 120–126. [Google Scholar] [CrossRef]
- Melo-Carrillo, A.; Lopez-Avila, A. A chronic animal model of migraine, induced by repeated meningeal nociception, characterized by a behavioral and pharmacological approach. Cephalalgia 2013, 33, 1096–1105. [Google Scholar] [CrossRef]
- Liu, D.; Chang, L.; Wang, J.; Zhu, Q.; Jiang, N.; Azhar, M.; Ahmed, A.; Zeng, G. Effect of Xiongmatang Extract on Behavioral and TRPV1-CGRP/CGRP-R Pathway in Rats with Migraine. Front. Pharmacol. 2022, 13, 835187. [Google Scholar] [CrossRef]
- Fan, S.; Xiao, Z.; Zhu, F.; He, X.; Lu, Z. A new comorbidity model and the common pathological mechanisms of migraine and epilepsy. Am. J. Transl. Res. 2017, 9, 2286–2295. [Google Scholar]
- Liu, L.; Xu, X.B.; Qu, Z.Y.; Zhao, L.P.; Zhang, C.S.; Li, Z.J.; Lyu, T.L.; Wang, X.F.; Jing, X.H.; Li, B. Determining 5HT(7)R’s Involvement in Modifying the Antihyperalgesic Effects of Electroacupuncture on Rats With Recurrent Migraine. Front. Neurosci. 2021, 15, 668616. [Google Scholar] [CrossRef] [PubMed]
- Nan, N.; Gong, M.X.; Wang, Q.; Li, M.J.; Xu, R.; Ma, Z.; Wang, S.H.; Zhao, H.; Xu, Y.S. Wuzhuyu Decoction relieves hyperalgesia by regulating central and peripheral 5-HT in chronic migraine model rats. Phytomedicine 2022, 96, 153905. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, X.; Wang, S.; Liu, C.; Fu, Q.; Qin, G.; Zhou, J.; Chen, L. Inhibition of Nerve Growth Factor Signaling Alleviates Repeated Dural Stimulation-induced Hyperalgesia in Rats. Neuroscience 2019, 398, 252–262. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, Y.; Liu, Q.; Jiang, L.; Li, M.; Wang, S.; Long, T.; He, W.; Kong, X.; Qin, G.; et al. P2X4-receptor participates in EAAT3 regulation via BDNF-TrkB signaling in a model of trigeminal allodynia. Mol. Pain 2018, 14, 1744806918795930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Tang, W.; Zhao, D.; Yu, S. Disrupted functional connectivity between the periaqueductal gray and other brain regions in a rat model of recurrent headache. Sci. Rep. 2017, 7, 3960. [Google Scholar] [CrossRef] [Green Version]
- Gong, Q.; Lin, Y.; Lu, Z.; Xiao, Z. Microglia-Astrocyte Cross Talk through IL-18/IL-18R Signaling Modulates Migraine-like Behavior in Experimental Models of Migraine. Neuroscience 2020, 451, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Wieseler, J.; Ellis, A.; McFadden, A.; Stone, K.; Brown, K.; Cady, S.; Bastos, L.F.; Sprunger, D.; Rezvani, N.; Johnson, K.; et al. Supradural inflammatory soup in awake and freely moving rats induces facial allodynia that is blocked by putative immune modulators. Brain Res. 2017, 1664, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Ran, Y.; He, Z.; Zhang, M.; Hu, G.; Tang, W.; Zhao, D.; Yu, S. Inhibition of toll-like receptor 4 alleviates hyperalgesia induced by acute dural inflammation in experimental migraine. Mol. Pain 2018, 14, 1744806918754612. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhang, L.; Hao, Y.; Yang, L.; Fan, S.; Xiao, Z. FKN/CX3CR1 axis facilitates migraine-Like behaviour by activating thalamic-cortical network microglia in status epilepticus model rats. J. Headache Pain 2022, 23, 42. [Google Scholar] [CrossRef]
- Vellani, V.; Moschetti, G.; Franchi, S.; Giacomoni, C.; Sacerdote, P.; Amodeo, G. Effects of NSAIDs on the Release of Calcitonin Gene-Related Peptide and Prostaglandin E2 from Rat Trigeminal Ganglia. Mediat. Inflamm. 2017, 2017, 9547056. [Google Scholar] [CrossRef] [Green Version]
- Fried, N.T.; Maxwell, C.R.; Elliott, M.B.; Oshinsky, M.L. Region-specific disruption of the blood-brain barrier following repeated inflammatory dural stimulation in a rat model of chronic trigeminal allodynia. Cephalalgia 2018, 38, 674–689. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Liu, C.; Jiang, L.; Li, M.; Long, T.; He, W.; Qin, G.; Chen, L.; Zhou, J. α7 Nicotinic acetylcholine receptor-mediated anti-inflammatory effect in a chronic migraine rat model via the attenuation of glial cell activation. J. Pain Res. 2018, 11, 1129–1140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.Y.; Zhou, H.R.; Wang, S.; Liu, C.Y.; Qin, G.C.; Fu, Q.Q.; Zhou, J.Y.; Chen, L.X. NR2B-Tyr phosphorylation regulates synaptic plasticity in central sensitization in a chronic migraine rat model. J. Headache Pain 2018, 19, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.; Wang, S.; Qin, G.; Xie, J.; Tan, G.; Zhou, J.; Chen, L. Protein Kinase C γ Contributes to Central Sensitization in a Rat Model of Chronic Migraine. J. Mol. Neurosci. 2017, 63, 131–141. [Google Scholar] [CrossRef]
- Stucky, N.L.; Gregory, E.; Winter, M.K.; He, Y.Y.; Hamilton, E.S.; McCarson, K.E.; Berman, N.E. Sex differences in behavior and expression of CGRP-related genes in a rodent model of chronic migraine. Headache 2011, 51, 674–692. [Google Scholar] [CrossRef]
- Liang, X.; Wang, S.; Qin, G.; Xie, J.; Tan, G.; Zhou, J.; McBride, D.W.; Chen, L. Tyrosine Phosphorylation of NR2B Contributes to Chronic Migraines via Increased Expression of CGRP in Rats. Biomed. Res. Int. 2017, 2017, 7203458. [Google Scholar] [CrossRef]
- Liang, J.; Zhou, X.; Wang, J.; Fei, Z.Y.; Qin, G.C.; Zhang, D.K.; Zhou, J.Y.; Chen, L.X. Upregulation of silent information regulator 1 alleviates mitochondrial dysfunction in the trigeminal nucleus caudalis in a rat model of chronic migraine. Neuroreport 2021, 32, 144–156. [Google Scholar] [CrossRef]
- Wang, J.; Fei, Z.; Liang, J.; Zhou, X.; Qin, G.; Zhang, D.; Zhou, J.; Chen, L. EphrinB/EphB Signaling Contributes to the Synaptic Plasticity of Chronic Migraine Through NR2B Phosphorylation. Neuroscience 2020, 428, 178–191. [Google Scholar] [CrossRef]
- Wang, S.; Wu, B.X.; Liu, C.Y.; Qin, G.C.; Yan, W.H.; Zhou, J.Y.; Chen, L.X. Expression of ASIC3 in the Trigeminal Nucleus Caudalis Plays a Role in a Rat Model of Recurrent Migraine. J. Mol. Neurosci. 2018, 66, 44–52. [Google Scholar] [CrossRef]
- Chen, N.; Su, W.; Cui, S.H.; Guo, J.; Duan, J.C.; Li, H.X.; He, L. A novel large animal model of recurrent migraine established by repeated administration of inflammatory soup into the dura mater of the rhesus monkey. Neural. Regen. Res. 2019, 14, 100–106. [Google Scholar]
- Tuka, B.; Helyes, Z.; Markovics, A.; Bagoly, T.; Szolcsányi, J.; Szabó, N.; Tóth, E.; Kincses, Z.T.; Vécsei, L.; Tajti, J. Alterations in PACAP-38-like immunoreactivity in the plasma during ictal and interictal periods of migraine patients. Cephalalgia 2013, 33, 1085–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Dong, Z.; Hou, L.; Wan, D.; Chen, M.; Tang, W.; Yu, S. Interictal plasma pituitary adenylate cyclase-activating polypeptide levels are decreased in migraineurs but remain unchanged in patients with tension-type headache. Clin. Chim. Acta 2015, 450, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Ran, Y.; Su, M.; Liu, Y.; Tang, W.; Dong, Z.; Yu, S. Chronic changes in pituitary adenylate cyclase-activating polypeptide and related receptors in response to repeated chemical dural stimulation in rats. Mol. Pain 2017, 13, 1744806917720361. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Vega, C.C.; Ahn, D.D.; Bischoff, C.; Wang, W.; Horne, D.; Wang, J.; Gavva, N.; Dussor, G. Meningeal transient receptor potential channel M8 activation causes cutaneous facial and hindpaw allodynia in a preclinical rodent model of headache. Cephalalgia 2016, 36, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Guan, X.; Chen, K.; Jin, S.; Wang, C.; Yan, L.; Shi, Z.; Zhang, X.; Chen, L.; Wan, Q. Abnormal mitochondrial dynamics and impaired mitochondrial biogenesis in trigeminal ganglion neurons in a rat model of migraine. Neurosci. Lett. 2017, 636, 127–133. [Google Scholar] [CrossRef]
- Jia, Z.; Yu, S.; Tang, W.; Zhao, D. Altered functional connectivity of the insula in a rat model of recurrent headache. Mol. Pain 2020, 16, 1744806920922115. [Google Scholar] [CrossRef] [Green Version]
- Jia, Z.; Chen, X.; Tang, W.; Zhao, D.; Yu, S. Atypical functional connectivity between the anterior cingulate cortex and other brain regions in a rat model of recurrent headache. Mol. Pain 2019, 15, 1744806919842483. [Google Scholar] [CrossRef]
- Becerra, L.; Bishop, J.; Barmettler, G.; Kainz, V.; Burstein, R.; Borsook, D. Brain network alterations in the inflammatory soup animal model of migraine. Brain Res. 2017, 1660, 36–46. [Google Scholar] [CrossRef]
- Bishop, J.; Becerra, L.; Barmettler, G.; Chang, P.C.; Kainz, V.; Burstein, R.; Borsook, D. Modulation of brain networks by sumatriptan-naproxen in the inflammatory soup migraine model. Pain 2019, 160, 2161–2171. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Hu, G.; Kang, L.; Ran, Y.; Su, M.; Yu, S. Cognitive impairment in a classical rat model of chronic migraine may be due to alterations in hippocampal synaptic plasticity and N-methyl-D-aspartate receptor subunits. Mol. Pain 2020, 16, 1744806920959582. [Google Scholar] [CrossRef]
- Zeng, X.; Niu, Y.; Qin, G.; Zhang, D.; Zhou, J.; Chen, L. Deficiency in the function of inhibitory interneurons contributes to glutamate-associated central sensitization through GABABR2-SynCAM1 signaling in chronic migraine rats. FASEB J. 2020, 34, 14780–14798. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zeng, X.; Qin, G.; Zhang, D.; Zhou, J.; Chen, L. Downregulation of metabotropic glutamate receptor 5 alleviates central sensitization by activating autophagy via inhibiting mTOR pathway in a rat model of chronic migraine. Neurosci. Lett. 2021, 743, 135552. [Google Scholar] [CrossRef] [PubMed]
- Boyer, N.; Dallel, R.; Artola, A.; Monconduit, L. General trigeminospinal central sensitization and impaired descending pain inhibitory controls contribute to migraine progression. Pain 2014, 155, 1196–1205. [Google Scholar] [CrossRef] [PubMed]
- Mi, X.; Ran, L.; Chen, L.; Qin, G. Recurrent Headache Increases Blood-Brain Barrier Permeability and VEGF Expression in Rats. Pain Physician. 2018, 21, E633–E642. [Google Scholar] [PubMed]
- Levy, D.; Zhang, X.C.; Jakubowski, M.; Burstein, R. Sensitization of meningeal nociceptors: Inhibition by naproxen. Eur. J. Neurosci. 2008, 27, 917–922. [Google Scholar] [CrossRef] [Green Version]
- Lambert, G.A.; Davis, J.B.; Appleby, J.M.; Chizh, B.A.; Hoskin, K.L.; Zagami, A.S. The effects of the TRPV1 receptor antagonist SB-705498 on trigeminovascular sensitisation and neurotransmission. Naunyn Schmiedeberg’s Arch. Pharmcol. 2009, 380, 311–325. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, L.; Yang, Y.; Zhao, L.; Xu, X.; Li, Z.; Zhu, Y.; Jing, X.; Wang, X.; Zhang, C.S.; et al. Electro-acupuncture inhibits C-fiber-evoked WDR neuronal activity of the trigeminocervical complex: Neurophysiological hypothesis of a complementary therapy for acute migraine modeled rats. Brain Res. 2020, 1730, 146670. [Google Scholar] [CrossRef]
- Boyer, N.; Signoret-Genest, J.; Artola, A.; Dallel, R.; Monconduit, L. Propranolol treatment prevents chronic central sensitization induced by repeated dural stimulation. Pain 2017, 158, 2025–2034. [Google Scholar] [CrossRef]
- Jia, Z.; Tang, W.; Zhao, D.; Hu, G.; Li, R.; Yu, S. Volumetric abnormalities of the brain in a rat model of recurrent headache. Mol. Pain 2018, 14, 1744806918756466. [Google Scholar] [CrossRef]
- Togha, M.; Jahromi, S.R.; Ghorbani, Z.; Ghaemi, A.; Rafiee, P. Evaluation of Inflammatory State in Migraineurs: A Case-control Study. Iran. J. Allergy Asthma Immunol. 2020, 19, 83–90. [Google Scholar] [CrossRef]
- Burgos-Vega, C.C.; Quigley, L.D.; Avona, A.; Price, T.; Dussor, G. Dural stimulation in rats causes brain-derived neurotrophic factor-dependent priming to subthreshold stimuli including a migraine trigger. Pain 2016, 157, 2722–2730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avona, A.; Price, T.J.; Dussor, G. Interleukin-6 induces spatially dependent whole-body hypersensitivity in rats: Implications for extracephalic hypersensitivity in migraine. J. Headache Pain 2021, 22, 70. [Google Scholar] [CrossRef] [PubMed]
- Avona, A.; Burgos-Vega, C.; Burton, M.D.; Akopian, A.N.; Price, T.J.; Dussor, G. Dural Calcitonin Gene-Related Peptide Produces Female-Specific Responses in Rodent Migraine Models. J. Neurosci. 2019, 39, 4323–4331. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Melemedjian, O.K.; Price, T.J.; Dussor, G. Sensitization of dural afferents underlies migraine-related behavior following meningeal application of interleukin-6 (IL-6). Mol. Pain 2012, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Burgos-Vega, C.C.; Quigley, L.D.; Santos, G.T.D.; Yan, F.; Asiedu, M.; Jacobs, B.; Motina, M.; Safdar, N.; Yousuf, H.; Avona, A.; et al. Non-invasive dural stimulation in mice: A novel preclinical model of migraine. Cephalalgia 2019, 39, 123–134. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, L.; Zhao, L.; Xu, X.; Li, Z.; Zhu, Y.; Zhang, C.; Jing, X.; Wang, X.; Li, B.; et al. Prophylactic Electroacupuncture on the Upper Cervical Segments Decreases Neuronal Discharges of the Trigeminocervical Complex in Migraine-Affected Rats: An in vivo Extracellular Electrophysiological Experiment. J. Pain Res. 2020, 13, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Lackovic, J.; Price, T.J.; Dussor, G. De novo protein synthesis is necessary for priming in preclinical models of migraine. Cephalalgia 2021, 41, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, K.A.; Edvinsson, L. Neurogenic inflammation: A study of rat trigeminal ganglion. J. Headache Pain 2010, 11, 485–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuris, A.; Xu, C.B.; Zhou, M.F.; Tajti, J.; Uddman, R.; Edvinsson, L. Enhanced expression of CGRP in rat trigeminal ganglion neurons during cell and organ culture. Brain Res. 2007, 1173, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Csati, A.; Edvinsson, L.; Vecsei, L.; Toldi, J.; Fulop, F.; Tajti, J.; Warfvinge, K. Kynurenic acid modulates experimentally induced inflammation in the trigeminal ganglion. J. Headache Pain 2015, 16, 99. [Google Scholar] [CrossRef] [Green Version]
- Burstein, R.; Jakubowski, M. Analgesic triptan action in an animal model of intracranial pain: A race against the development of central sensitization. Ann. Neurol. 2004, 55, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Lukacs, M.; Haanes, K.A.; Majlath, Z.; Tajti, J.; Vecsei, L.; Warfvinge, K.; Edvinsson, L. Dural administration of inflammatory soup or Complete Freund’s Adjuvant induces activation and inflammatory response in the rat trigeminal ganglion. J. Headache Pain 2015, 16, 564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukacs, M.; Warfvinge, K.; Tajti, J.; Fulop, F.; Toldi, J.; Vecsei, L.; Edvinsson, L. Topical dura mater application of CFA induces enhanced expression of c-fos and glutamate in rat trigeminal nucleus caudalis: Attenuated by KYNA derivate (SZR72). J. Headache Pain 2017, 18, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajti, J.; Kuris, A.; Vecsei, L.; Xu, C.B.; Edvinsson, L. Organ culture of the trigeminal ganglion induces enhanced expression of calcitonin gene-related peptide via activation of extracellular signal-regulated protein kinase 1/2. Cephalalgia 2011, 31, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Tanimoto, T.; Kadoi, J.; Nasu, M.; Takahashi, M.; Kitagawa, J.; Matsumoto, S. Enhanced excitability of nociceptive trigeminal ganglion neurons by satellite glial cytokine following peripheral inflammation. Pain 2007, 129, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Takeda, M.; Takahashi, M.; Matsumoto, S. Contribution of activated interleukin receptors in trigeminal ganglion neurons to hyperalgesia via satellite glial interleukin-1beta paracrine mechanism. Brain Behav. Immun. 2008, 22, 1016–1023. [Google Scholar] [CrossRef]
- Burstein, R.; Yamamura, H.; Malick, A.; Strassman, A.M. Chemical stimulation of the intracranial dura induces enhanced responses to facial stimulation in brain stem trigeminal neurons. J. Neurophysiol. 1998, 79, 964–982. [Google Scholar] [CrossRef] [Green Version]
- Strassman, A.M.; Raymond, S.A.; Burstein, R. Sensitization of meningeal sensory neurons and the origin of headaches. Nature 1996, 384, 560–564. [Google Scholar] [CrossRef]
- Lukacs, M.; Warfvinge, K.; Kruse, L.S.; Tajti, J.; Fulop, F.; Toldi, J.; Vecsei, L.; Edvinsson, L. KYNA analogue SZR72 modifies CFA-induced dural inflammation- regarding expression of pERK1/2 and IL-1beta in the rat trigeminal ganglion. J. Headache Pain 2016, 17, 64. [Google Scholar] [CrossRef] [Green Version]
- Romero-Reyes, M.; Pardi, V.; Akerman, S. A potent and selective calcitonin gene-related peptide (CGRP) receptor antagonist, MK-8825, inhibits responses to nociceptive trigeminal activation: Role of CGRP in orofacial pain. Exp. Neurol. 2015, 271, 95–103. [Google Scholar] [CrossRef]
- Janeway, C.A., Jr. Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harb. Symp. Quant. Biol. 1989, 54, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Spekker, E.; Laborc, K.F.; Bohár, Z.; Nagy-Grócz, G.; Fejes-Szabó, A.; Szűcs, M.; Vécsei, L.; Párdutz, Á. Effect of dural inflammatory soup application on activation and sensitization markers in the caudal trigeminal nucleus of the rat and the modulatory effects of sumatriptan and kynurenic acid. J. Headache Pain 2021, 22, 17. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.R.; Paramos-de-Carvalho, D.; Certal, M.; Costa, C.; Magalhães-Cardoso, M.T.; Ferreirinha, F.; Costa, M.A.; Correia-de-Sá, P. Bradykinin-induced Ca2+ signaling in human subcutaneous fibroblasts involves ATP release via hemichannels leading to P2Y12 receptors activation. Cell Commun. Signal. 2013, 11, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinheiro, A.R.; Paramos-de-Carvalho, D.; Certal, M.; Costa, M.A.; Costa, C.; Magalhaes-Cardoso, M.T.; Ferreirinha, F.; Sevigny, J.; Correia-de-Sa, P. Histamine induces ATP release from human subcutaneous fibroblasts, via pannexin-1 hemichannels, leading to Ca2+ mobilization and cell proliferation. J. Biol. Chem. 2013, 288, 27571–27583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowal, J.M.; Haanes, K.A.; Christensen, N.M.; Novak, I. Bile acid effects are mediated by ATP release and purinergic signalling in exocrine pancreatic cells. Cell Commun. Signal 2015, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- Beckel, J.M.; Gomez, N.M.; Lu, W.; Campagno, K.E.; Nabet, B.; Albalawi, F.; Lim, J.C.; Boesze-Battaglia, K.; Mitchell, C.H. Stimulation of TLR3 triggers release of lysosomal ATP in astrocytes and epithelial cells that requires TRPML1 channels. Sci. Rep. 2018, 8, 5726. [Google Scholar] [CrossRef]
- Matsuka, Y.; Neubert, J.K.; Maidment, N.T.; Spigelman, I. Concurrent release of ATP and substance P within guinea pig trigeminal ganglia in vivo. Brain Res. 2001, 915, 248–255. [Google Scholar] [CrossRef]
- Koroleva, K.; Gafurov, O.; Guselnikova, V.; Nurkhametova, D.; Giniatullina, R.; Sitdikova, G.; Mattila, O.S.; Lindsberg, P.J.; Malm, T.M.; Giniatullin, R. Meningeal Mast Cells Contribute to ATP-Induced Nociceptive Firing in Trigeminal Nerve Terminals: Direct and Indirect Purinergic Mechanisms Triggering Migraine Pain. Front. Cell. Neurosci. 2019, 13, 195. [Google Scholar] [CrossRef]
- Haanes, K.A.; Edvinsson, L. Expression and characterization of purinergic receptors in rat middle meningeal artery-potential role in migraine. PLoS ONE 2014, 9, e108782. [Google Scholar] [CrossRef]
- Yegutkin, G.G.; Guerrero-Toro, C.; Kilinc, E.; Koroleva, K.; Ishchenko, Y.; Abushik, P.; Giniatullina, R.; Fayuk, D.; Giniatullin, R. Nucleotide homeostasis and purinergic nociceptive signaling in rat meninges in migraine-like conditions. Purinergic Signal 2016, 12, 561–574. [Google Scholar] [CrossRef] [Green Version]
- Haanes, K.A.; Labastida-Ramirez, A.; Blixt, F.W.; Rubio-Beltran, E.; Dirven, C.M.; Danser, A.H.; Edvinsson, L.; MaassenVanDenBrink, A. Exploration of purinergic receptors as potential anti-migraine targets using established pre-clinical migraine models. Cephalalgia 2019, 39, 1421–1434. [Google Scholar] [CrossRef] [PubMed]
- Dussor, G. Serotonin, 5HT1 agonists, and migraine: New data, but old questions still not answered. Curr. Opin. Support Palliat Care 2014, 8, 137–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laborc, K.F.; Spekker, E.; Bohar, Z.; Szucs, M.; Nagy-Grocz, G.; Fejes-Szabo, A.; Vecsei, L.; Pardutz, A. Trigeminal activation patterns evoked by chemical stimulation of the dura mater in rats. J. Headache Pain 2020, 21, 101. [Google Scholar] [CrossRef] [PubMed]
- Harriott, A.M.; Strother, L.C.; Vila-Pueyo, M.; Holland, P.R. Animal models of migraine and experimental techniques used to examine trigeminal sensory processing. J. Headache Pain 2019, 20, 91. [Google Scholar] [CrossRef] [PubMed]
- Vingen, J.V.; Pareja, J.A.; Støren, O.; White, L.R.; Stovner, L.J. Phonophobia in migraine. Cephalalgia 1998, 18, 243–249. [Google Scholar] [CrossRef]
- Noseda, R.; Copenhagen, D.; Burstein, R. Current understanding of photophobia, visual networks and headaches. Cephalalgia 2019, 39, 1623–1634. [Google Scholar] [CrossRef]
- Farajdokht, F.; Babri, S.; Karimi, P.; Mohaddes, G. Ghrelin attenuates hyperalgesia and light aversion-induced by nitroglycerin in male rats. Neurosci. Lett. 2016, 630, 30–37. [Google Scholar] [CrossRef]
- Vuralli, D.; Wattiez, A.S.; Russo, A.F.; Bolay, H. Behavioral and cognitive animal models in headache research. J. Headache Pain 2019, 20, 11. [Google Scholar] [CrossRef]
- Bourin, M.; Hascoët, M. The mouse light/dark box test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef]
- Parent, A.J.; Beaudet, N.; Beaudry, H.; Bergeron, J.; Bérubé, P.; Drolet, G.; Sarret, P.; Gendron, L. Increased anxiety-like behaviors in rats experiencing chronic inflammatory pain. Behav. Brain Res. 2012, 229, 160–167. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, L.; Karl, F.; Sommer, C.; Üçeyler, N. Affective and cognitive behavior in the alpha-galactosidase A deficient mouse model of Fabry disease. PLoS ONE 2017, 12, e0180601. [Google Scholar] [CrossRef] [Green Version]
- Sufka, K.J.; Staszko, S.M.; Johnson, A.P.; Davis, M.E.; Davis, R.E.; Smitherman, T.A. Clinically relevant behavioral endpoints in a recurrent nitroglycerin migraine model in rats. J. Headache Pain 2016, 17, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lantéri-Minet, M.; Radat, F.; Chautard, M.H.; Lucas, C. Anxiety and depression associated with migraine: Influence on migraine subjects’ disability and quality of life, and acute migraine management. Pain 2005, 118, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Open Field Test for Measuring Locomotor Activity and Anxiety-Like Behavior. Methods Mol. Biol. 2019, 1916, 99–103. [Google Scholar] [PubMed]
- Ferrari, M.D. Migraine. Lancet 1998, 351, 1043–1051. [Google Scholar] [CrossRef]
- Moye, L.S.; Pradhan, A.A.A. Animal Model of Chronic Migraine-Associated Pain. Curr. Protoc. Neurosci. 2017, 80, 9.60.1–9.60.9. [Google Scholar] [CrossRef]
- Ray, J.C.; Allen, P.; Bacsi, A.; Bosco, J.J.; Chen, L.; Eller, M.; Kua, H.; Lim, L.L.; Matharu, M.S.; Monif, M.; et al. Inflammatory complications of CGRP monoclonal antibodies: A case series. J. Headache Pain 2021, 22, 121. [Google Scholar] [CrossRef]
- Borkum, J.M. Brain Energy Deficit as a Source of Oxidative Stress in Migraine: A Molecular Basis for Migraine Susceptibility. Neurochem. Res. 2021, 46, 1913–1932. [Google Scholar] [CrossRef]
- Reyngoudt, H.; Achten, E.; Paemeleire, K. Magnetic resonance spectroscopy in migraine: What have we learned so far? Cephalalgia 2012, 32, 845–859. [Google Scholar] [CrossRef] [Green Version]
- Younis, S.; Hougaard, A.; Vestergaard, M.B.; Larsson, H.B.W.; Ashina, M. Migraine and magnetic resonance spectroscopy: A systematic review. Curr. Opin. Neurol. 2017, 30, 246–262. [Google Scholar] [CrossRef]
- Tiehuis, L.H.; Koene, S.; Saris, C.G.J.; Janssen, M.C.H. Mitochondrial migraine; a prevalence, impact and treatment efficacy cohort study. Mitochondrion 2020, 53, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Dalkara, T.; Kilic, K. How does fasting trigger migraine? A hypothesis. Curr. Pain Headache Rep. 2013, 17, 368. [Google Scholar] [CrossRef]
- Stuart, S.; Griffiths, L.R. A possible role for mitochondrial dysfunction in migraine. Mol. Genet. Genom. 2012, 287, 837–844. [Google Scholar] [CrossRef] [PubMed]
- More, S.V.; Kumar, H.; Kim, I.S.; Song, S.Y.; Choi, D.K. Cellular and molecular mediators of neuroinflammation in the pathogenesis of Parkinson’s disease. Mediat. Inflamm 2013, 2013, 952375. [Google Scholar] [CrossRef] [PubMed]
- Kielian, T. Multifaceted roles of neuroinflammation: The need to consider both sides of the coin. J. Neurochem. 2016, 136, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.M.; Rothwell, N.J.; Gibson, R.M. The role of inflammation in CNS injury and disease. Br. J. Pharmacol. 2006, 147, S232–S240. [Google Scholar] [CrossRef] [Green Version]
- Winters, B.L.; Jeong, H.J.; Vaughan, C.W. Inflammation induces developmentally regulated sumatriptan inhibition of spinal synaptic transmission. Br. J. Pharmacol. 2020, 177, 3730–3743. [Google Scholar] [CrossRef]
- Cornelison, L.E.; Woodman, S.E.; Durham, P.L. Inhibition of Trigeminal Nociception by Non-invasive Vagus Nerve Stimulation: Investigating the Role of GABAergic and Serotonergic Pathways in a Model of Episodic Migraine. Front Neurol. 2020, 11, 146. [Google Scholar] [CrossRef]
- Yarwood, R.E.; Imlach, W.L.; Lieu, T.; Veldhuis, N.A.; Jensen, D.D.; Herenbrink, C.K.; Aurelio, L.; Cai, Z.; Christie, M.J.; Poole, D.P.; et al. Endosomal signaling of the receptor for calcitonin gene-related peptide mediates pain transmission. Proc. Natl. Acad. Sci. USA 2017, 114, 12309–12314. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, S.; Corradini, L.; Just, S.; Arndt, K.; Doods, H. The CGRP receptor antagonist BIBN4096BS peripherally alleviates inflammatory pain in rats. Pain 2013, 154, 700–707. [Google Scholar] [CrossRef]
- Wang, D.; Ruan, L.; Hong, Y.; Chabot, J.G.; Quirion, R. Involvement of PKA-dependent upregulation of nNOS-CGRP in adrenomedullin-initiated mechanistic pathway underlying CFA-induced response in rats. Exp. Neurol. 2013, 239, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Warfvinge, K.; Krause, D.N.; Maddahi, A.; Edvinsson, J.C.A.; Edvinsson, L.; Haanes, K.A. Estrogen receptors alpha, beta and GPER in the CNS and trigeminal system—molecular and functional aspects. J. Headache Pain 2020, 21, 131. [Google Scholar] [CrossRef] [PubMed]
- Woldeamanuel, Y.W.; Rapoport, A.M.; Cowan, R.P. The place of corticosteroids in migraine attack management: A 65-year systematic review with pooled analysis and critical appraisal. Cephalalgia 2015, 35, 996–1024. [Google Scholar] [CrossRef] [PubMed]
- Gunaydin, C.; Bilge, S.S. Effects of Nonsteroidal Anti-Inflammatory Drugs at the Molecular Level. Eurasian J. Med. 2018, 50, 116–121. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reducha, P.V.; Edvinsson, L.; Haanes, K.A. Could Experimental Inflammation Provide Better Understanding of Migraines? Cells 2022, 11, 2444. https://doi.org/10.3390/cells11152444
Reducha PV, Edvinsson L, Haanes KA. Could Experimental Inflammation Provide Better Understanding of Migraines? Cells. 2022; 11(15):2444. https://doi.org/10.3390/cells11152444
Chicago/Turabian StyleReducha, Philip Victor, Lars Edvinsson, and Kristian Agmund Haanes. 2022. "Could Experimental Inflammation Provide Better Understanding of Migraines?" Cells 11, no. 15: 2444. https://doi.org/10.3390/cells11152444
APA StyleReducha, P. V., Edvinsson, L., & Haanes, K. A. (2022). Could Experimental Inflammation Provide Better Understanding of Migraines? Cells, 11(15), 2444. https://doi.org/10.3390/cells11152444