
Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements



Abstract
:1. Introduction
2. Materials and Methods
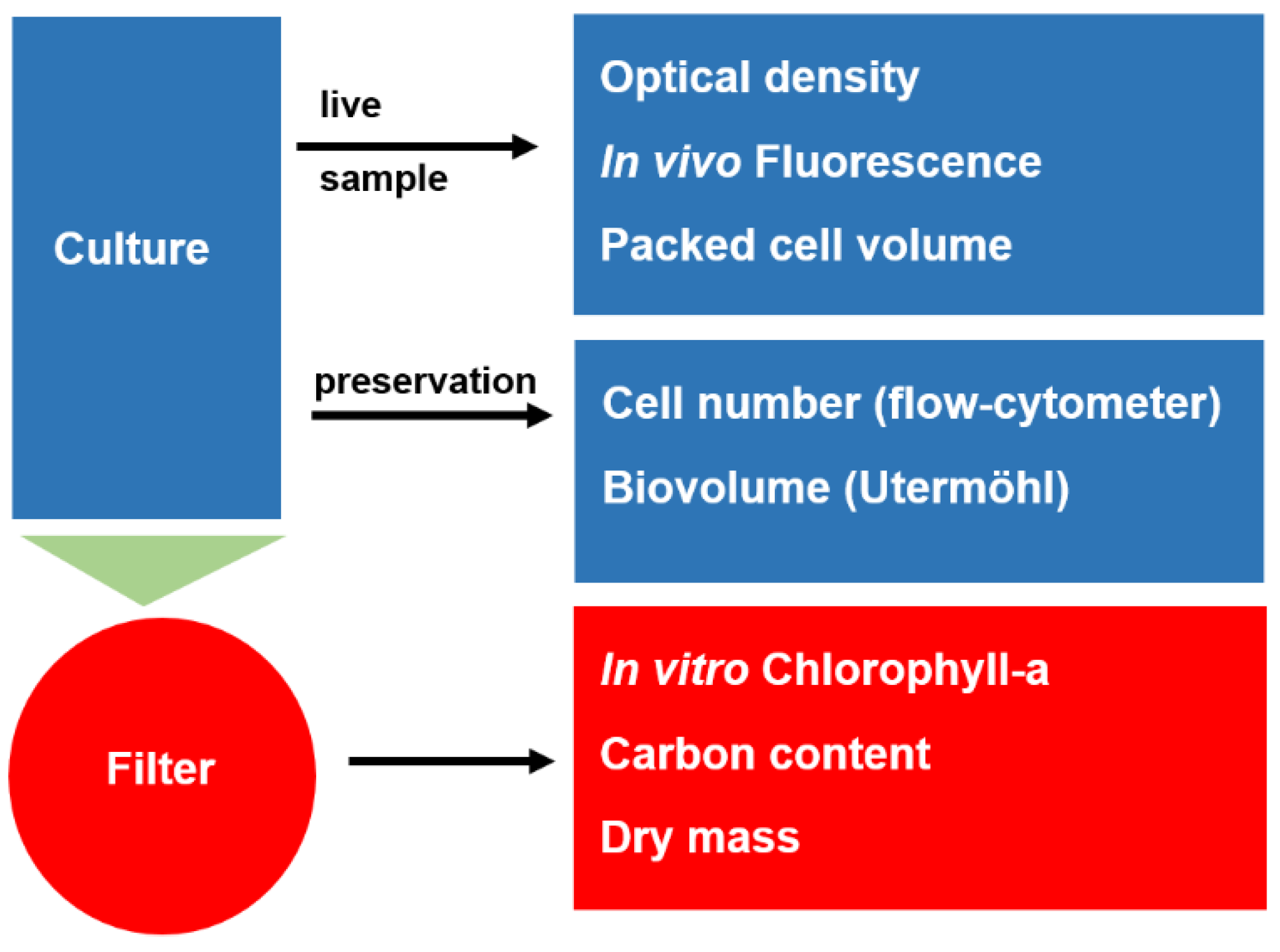
2.1. Methods Applied
2.1.1. Dry Mass
2.1.2. Packed Cell Volume (PCV)
2.1.3. Algal Cell Number and Biovolume
2.1.4. Carbon Content
2.1.5. In Vitro Chlorophyll-a
2.1.6. In Vivo Chlorophyll-a
2.1.7. Optical Density
2.1.8. Pulse-Amplified Modulated Fluorescence
2.2. Experiments Conducted
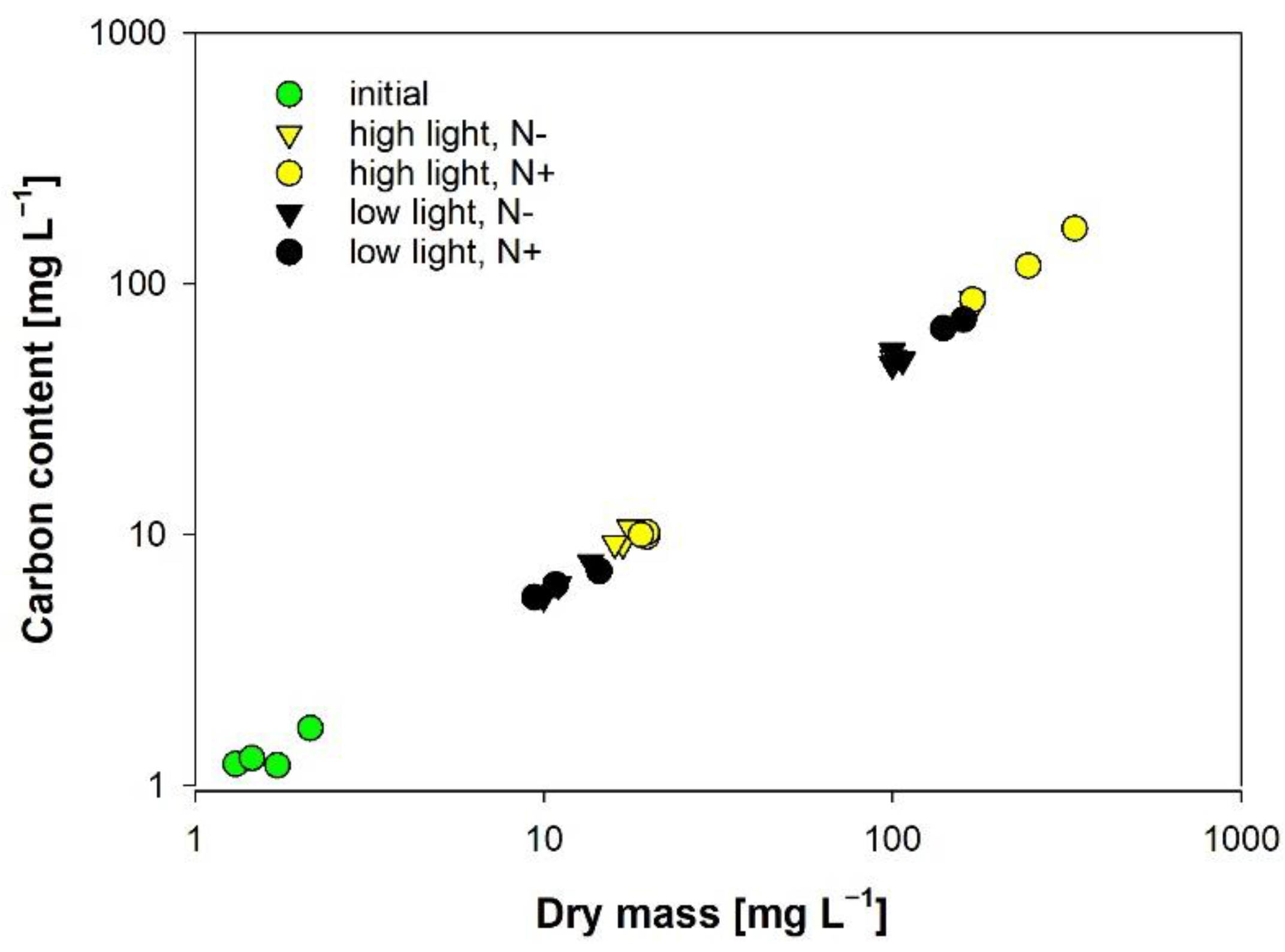
2.2.1. Carbon Content
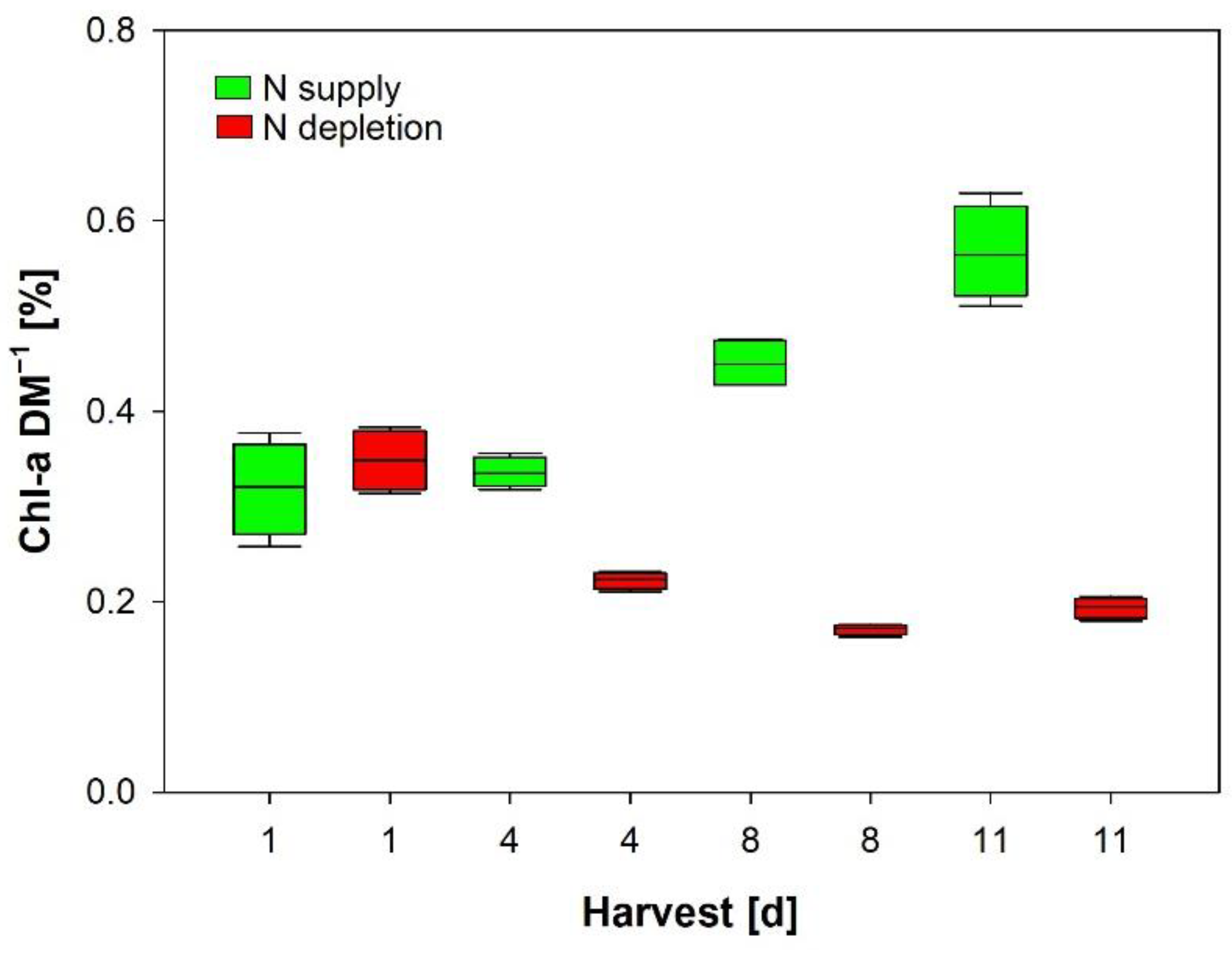
2.2.2. In Vitro Chlorophyll-a
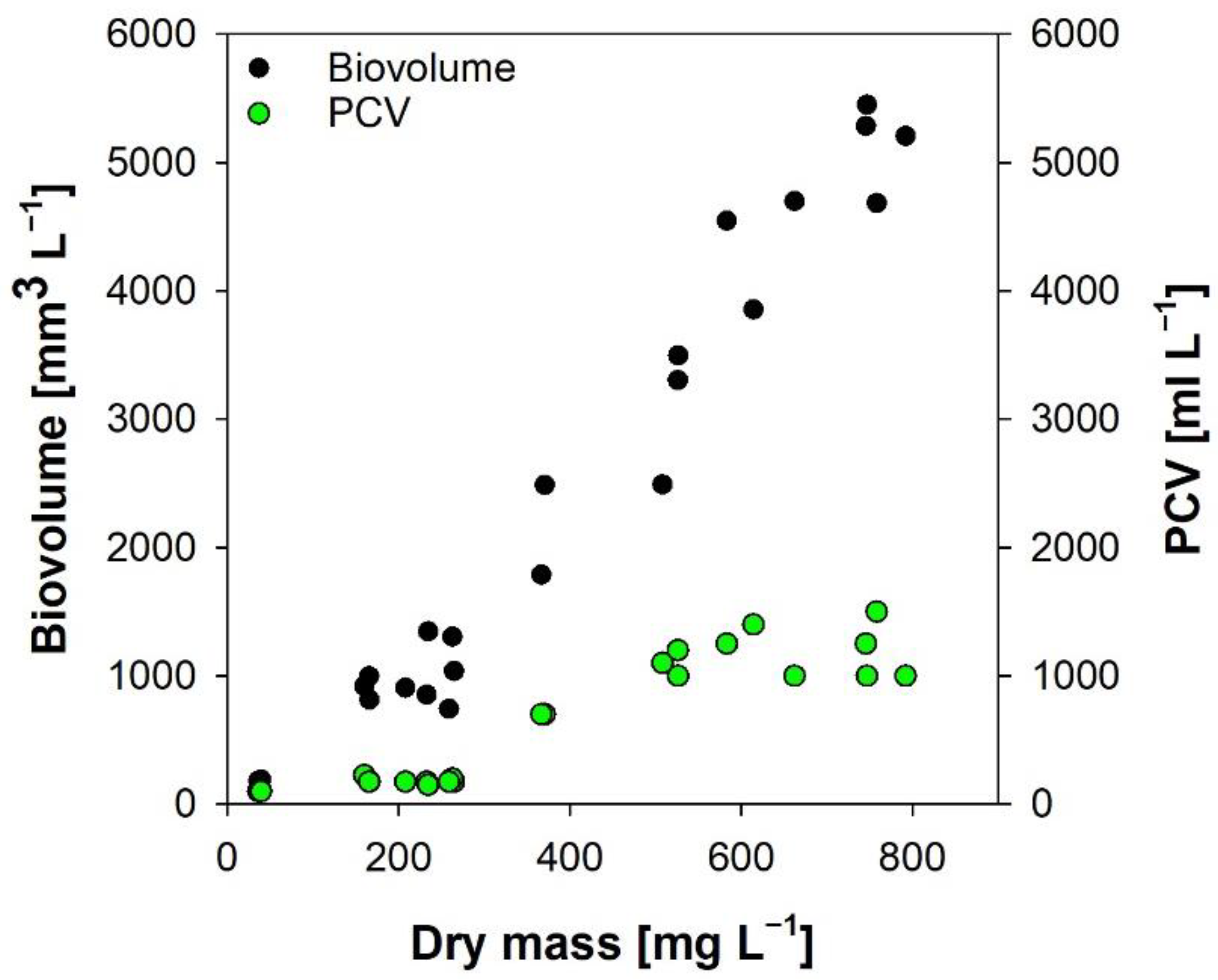
2.2.3. Packed Cell Volume and Algal Biovolume
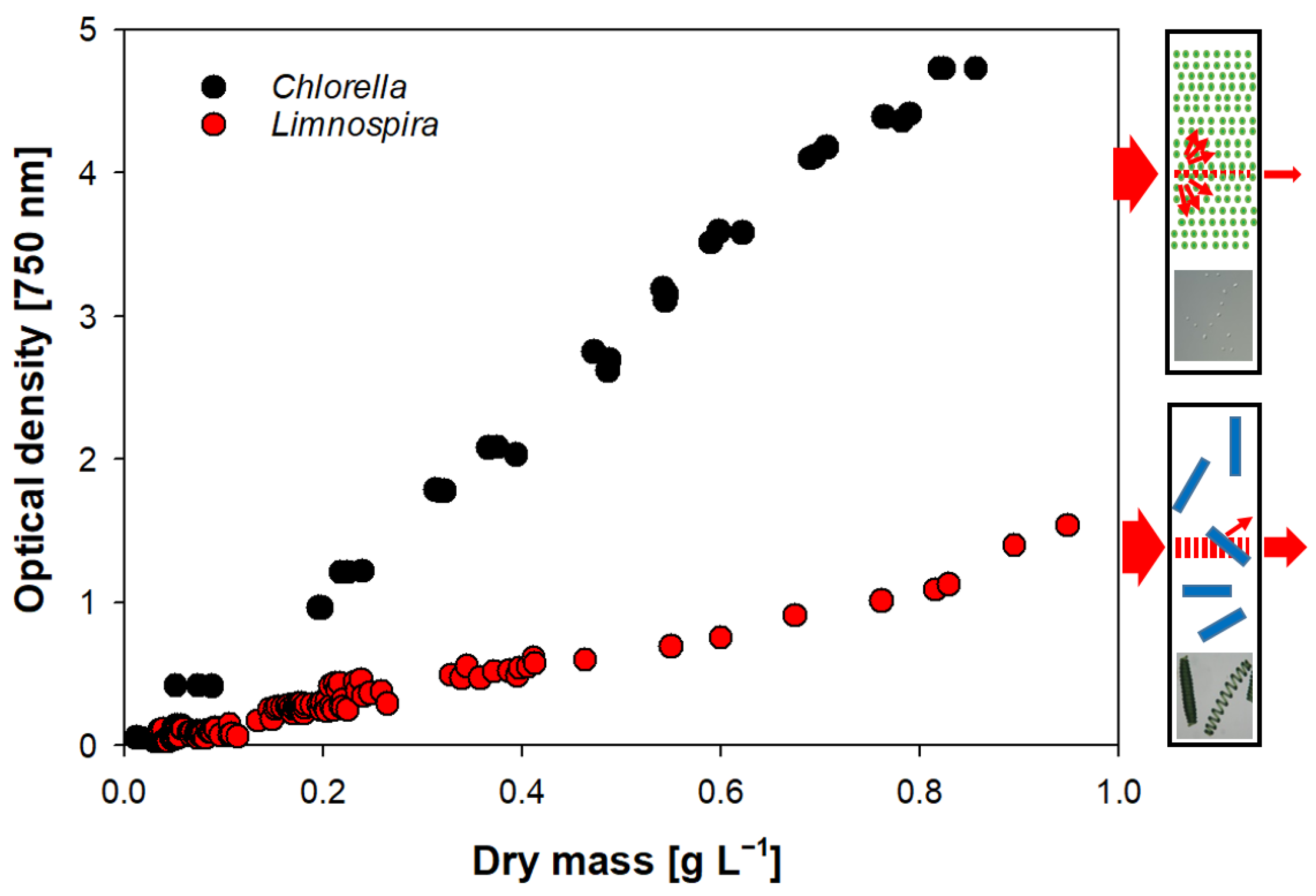
2.2.4. Optical Density
- (1)
- Unialgal batch cultures of Chlorella vulgaris were grown in 2 L cultivation flasks placed on magnet stirrers. Growth conditions were BG11 medium at 22 °C and permanent light of approximately 150 µmol photons m−2 s−1 reaching the bottle’s light-facing side (fluorescent tubes, Lumilux Cool White, 36 W/840, Osram, Munich, Germany). Sterile air enriched with 5% CO2 was delivered to the bottom of the flask via a silicone hose. Limnospira fusiformis was cultivated in Zarrouk medium in bubble column reactors run with sterile air at 27 °C and 50 µmol photons m−2 s−1 and a light:dark cycle = 12 h:12 h (warm-white LED panels).
- (2)
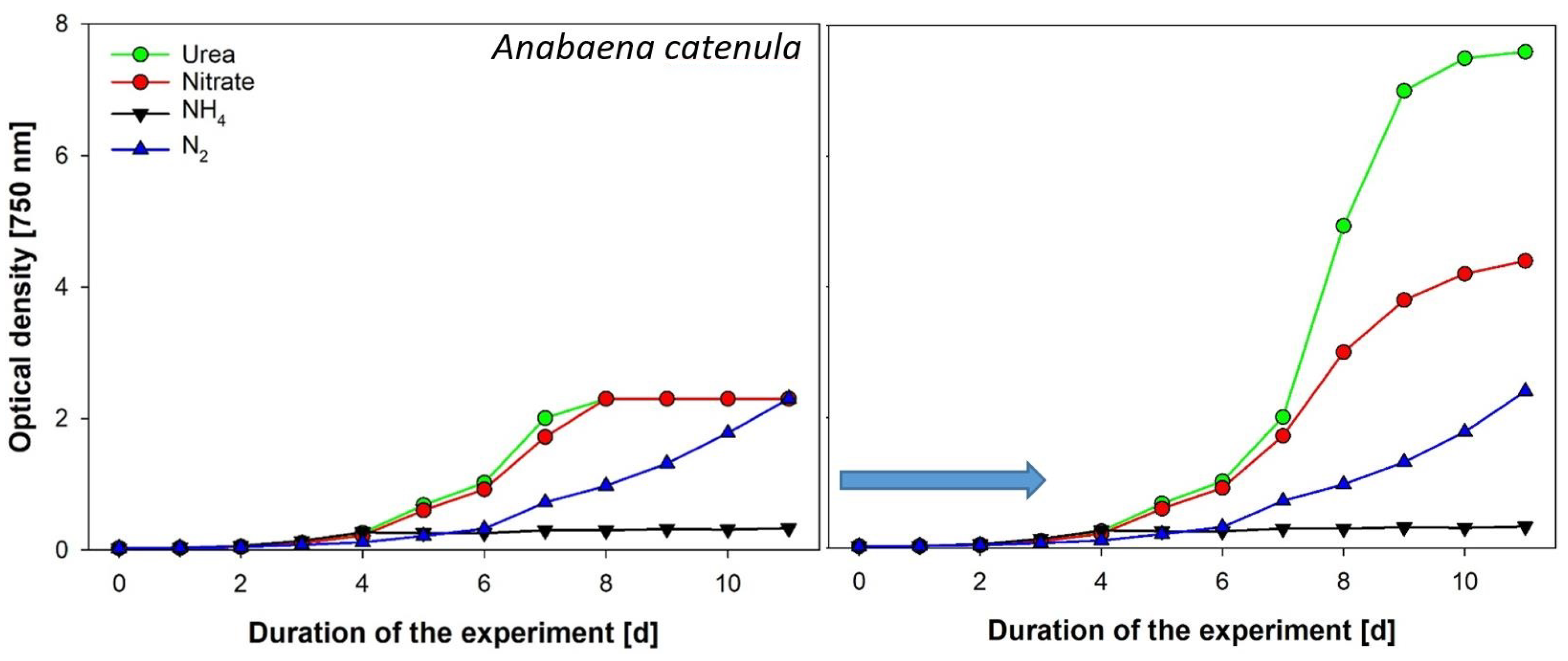
- Anabaena catenula was grown at 25 °C in bubble column reactors aerated with sterile air. Continuous light supply was provided via warm-white LED panels and 30 µmol photons m−2 s−1. Culture treatments consisted of four groups with different nitrogen supply (3 replicates each). The basic growth medium was BG11 but without inorganic nitrogen (treatment 1 with atmospheric N). Three more N sources were added to the basic growth medium for the respective treatment as “pulses” every 24 h: 1 mM NaNO3, 1 mM NH4Cl, 0.5 mM CH4N2O.
2.2.5. In Vivo Chlorophyll-a
2.2.6. Comparison of Biomass Parameters
2.2.7. Algal Cell Vitality
3. Results and Discussion
3.1. Dry Mass
3.2. Carbon Content
3.3. In Vitro Chlorophyll-a
3.4. Packed Cell Volume and Algal Biovolume
3.5. Optical Density
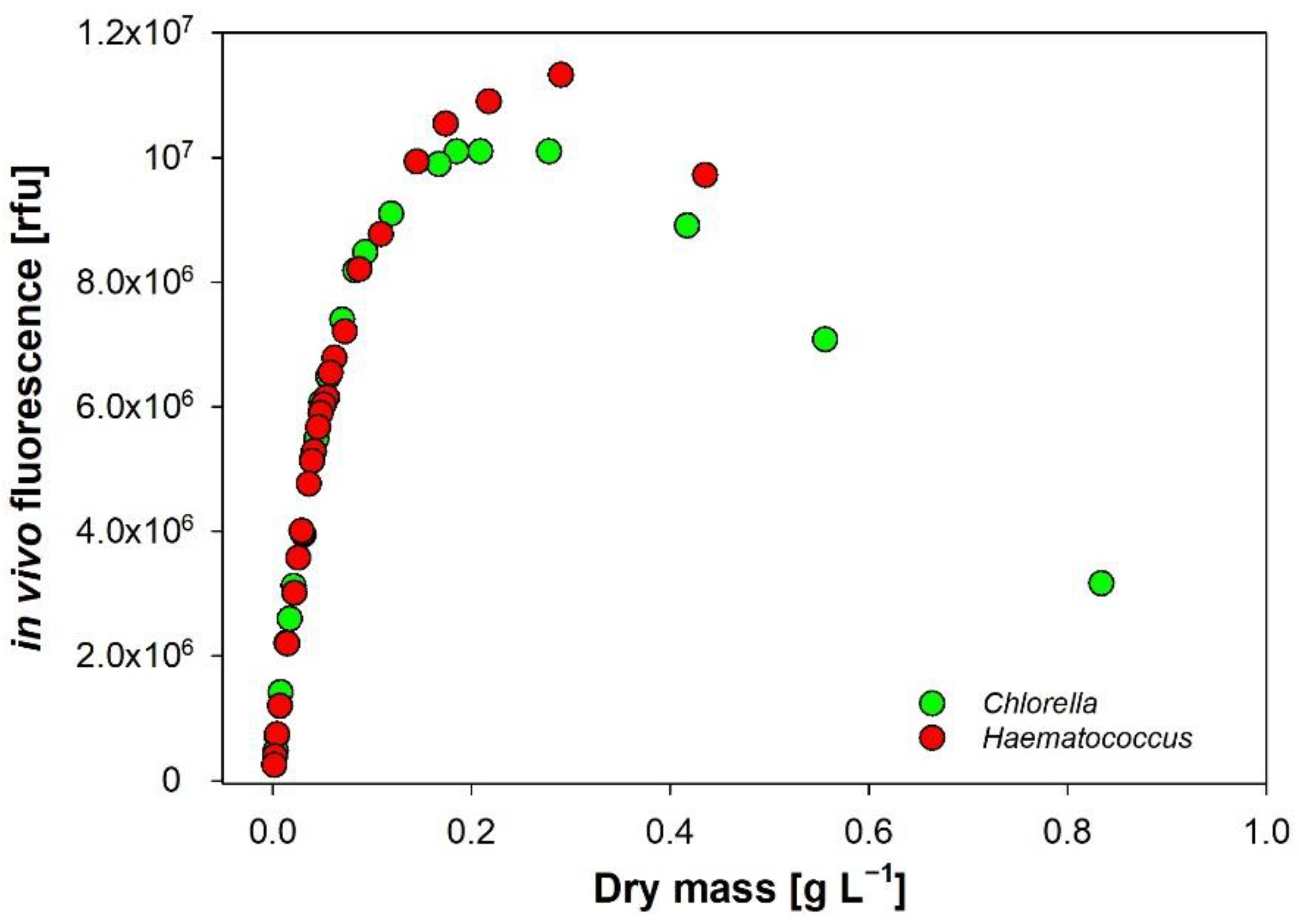
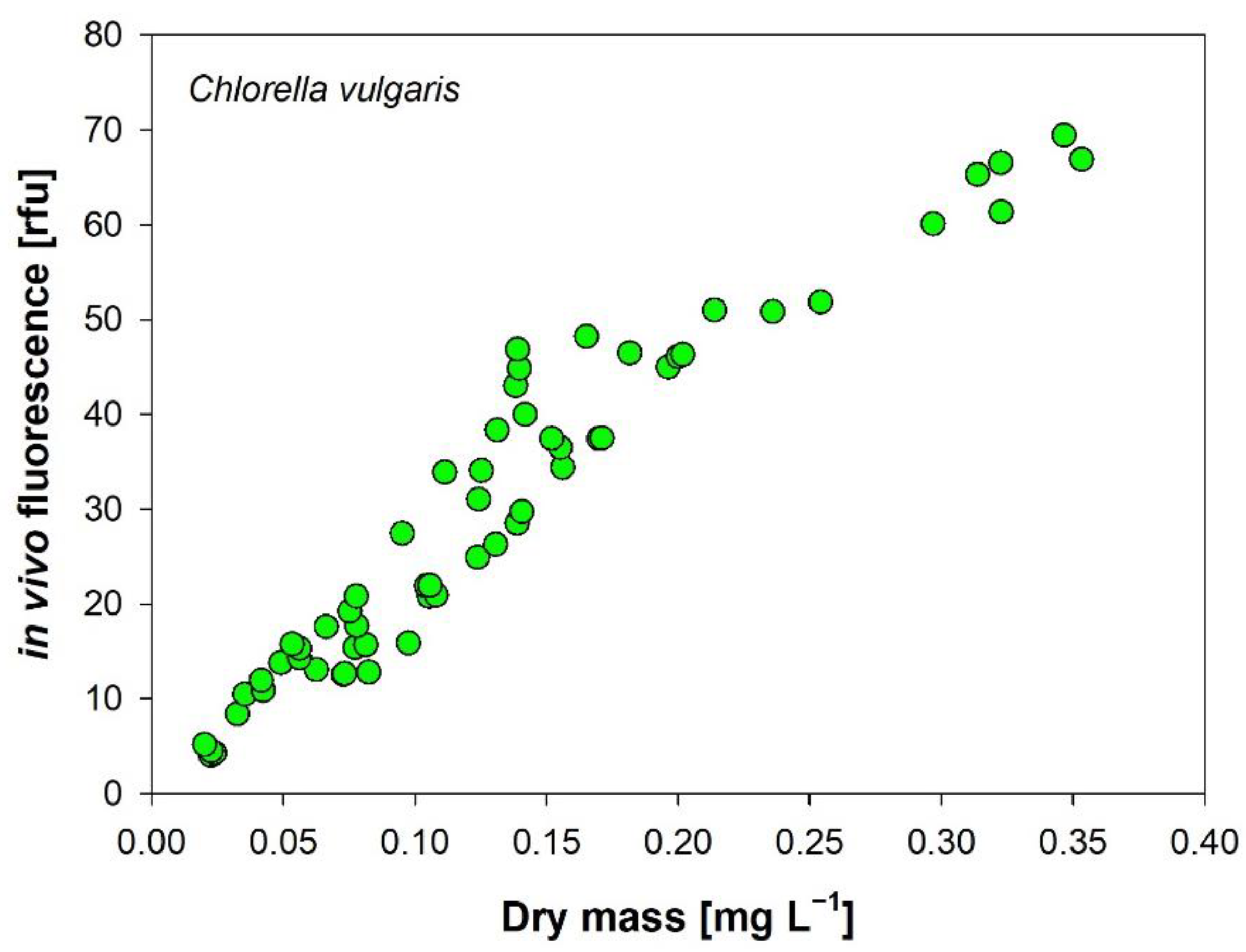
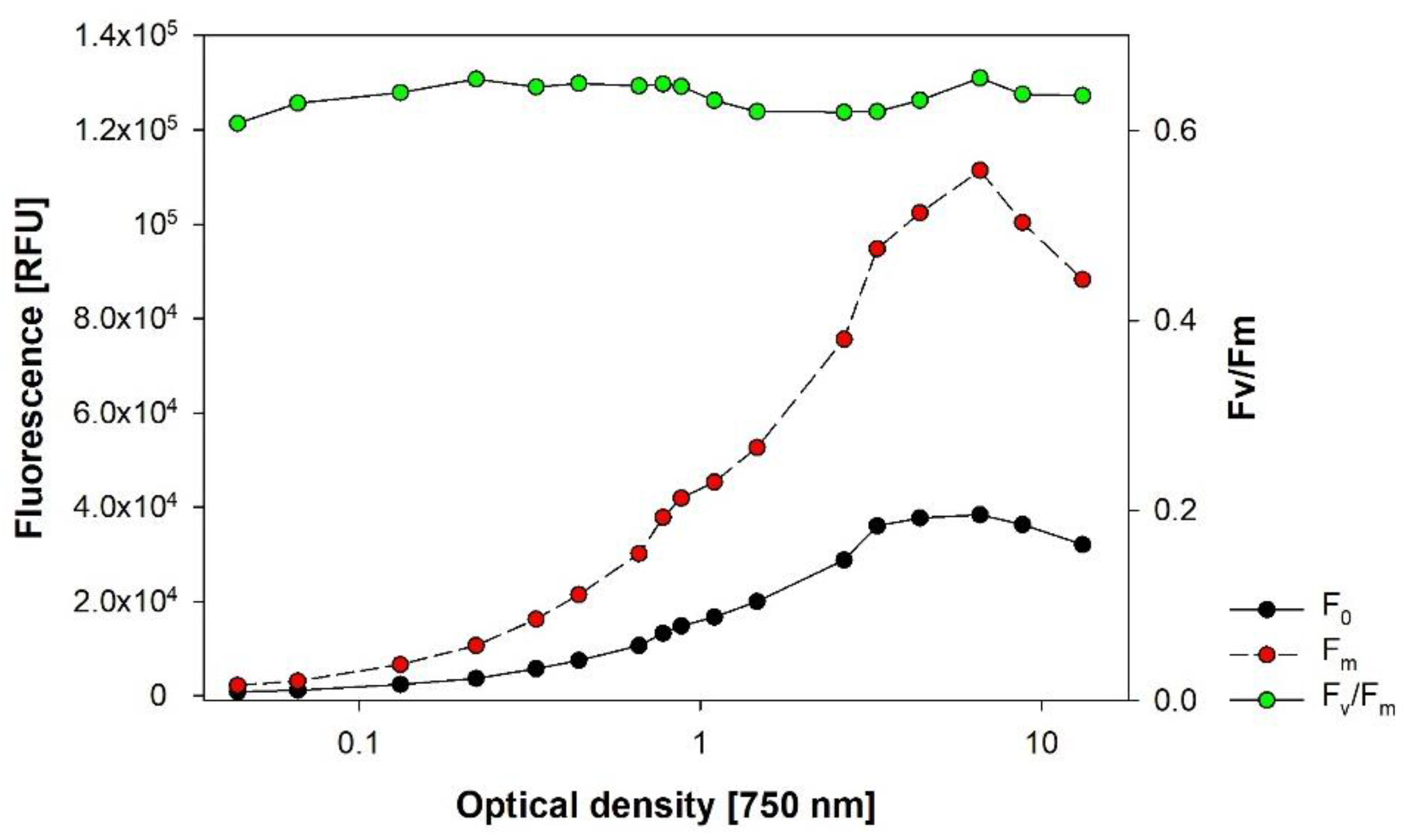
3.6. In Vivo Autofluorescence of Chlorophyll-a
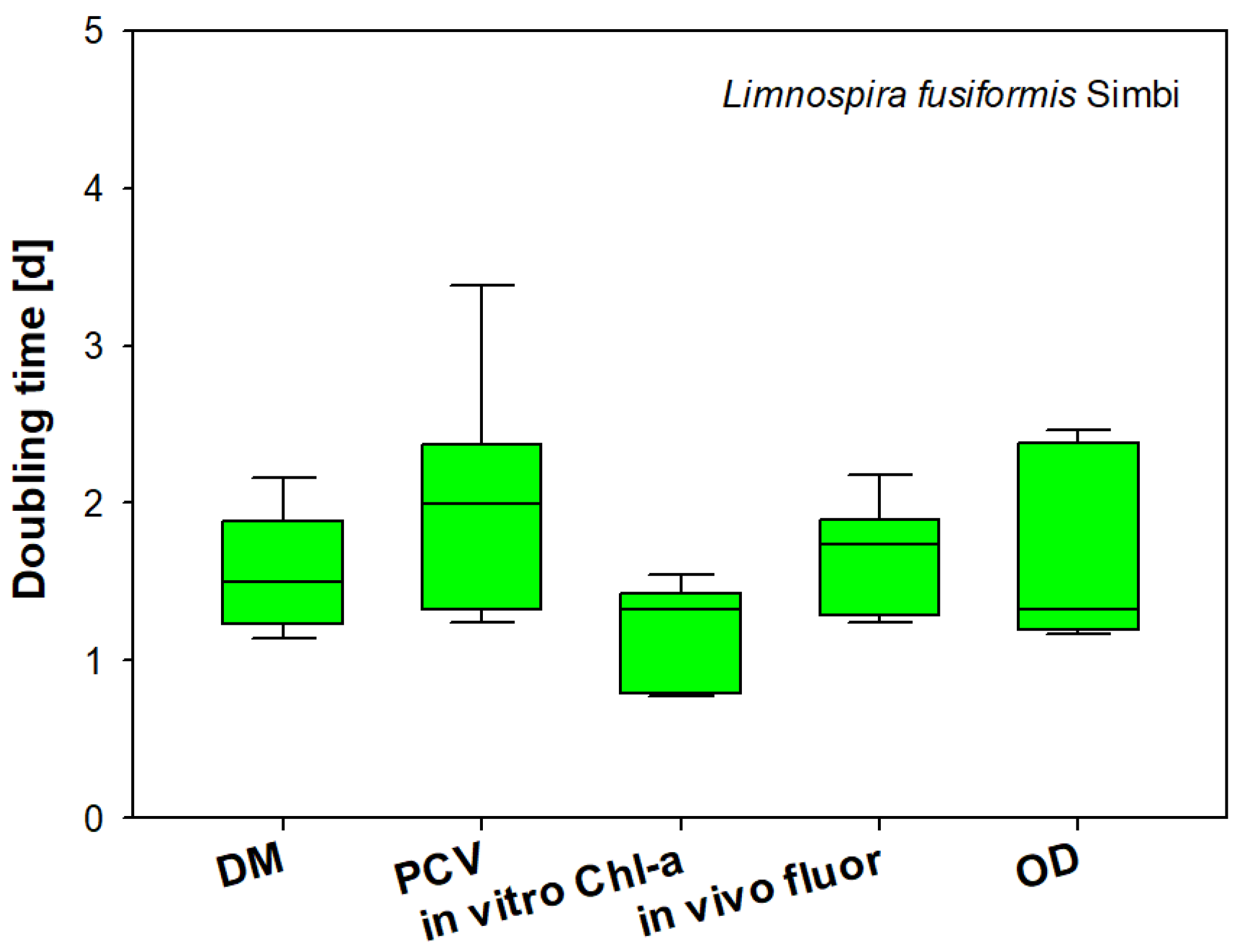
3.7. Comparison of Biomass Parameters
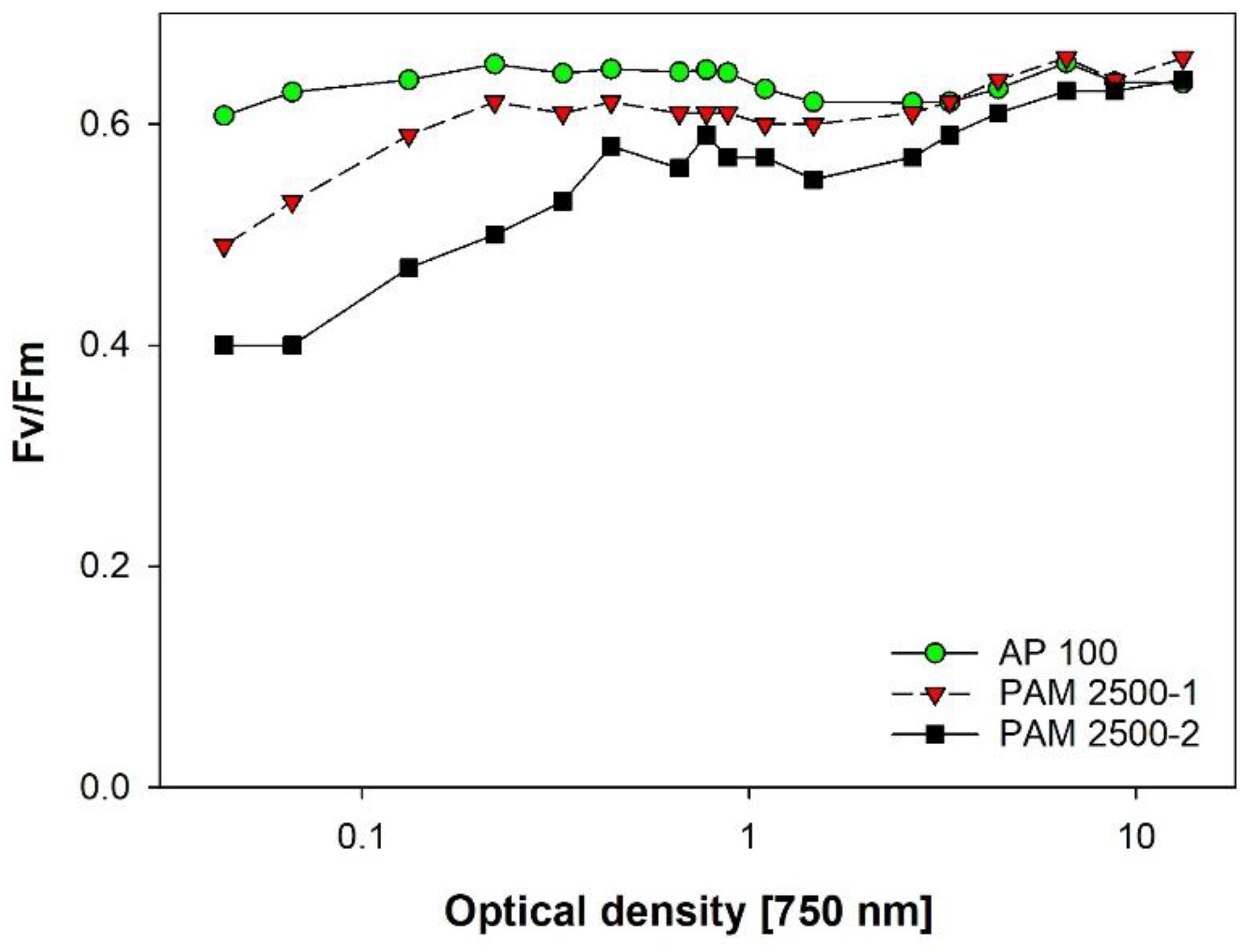
3.8. Vitality of Microalgae via PAM-Fluorescence
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reardon, K.F.; Scheper, T.H. Determination of Cell Concentration and Characterization of Cells. In Biotechnology Set, 2nd ed.; Rehm, H.J., Reed, G., Eds.; Wiley: Hoboken, NJ, USA, 2001; pp. 179–223. [Google Scholar]
- Moheimani, N.R.; Borowitzka, M.A.; Isdepsky, A.; Sing, S.F. Standard Methods for Measuring Growth of Algae and Their Composition. In Algae for Biofuels and Energy; Borowitzka, M.A., Moheimani, N.R., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 265–284. [Google Scholar]
- Bhattacharya, S.; Shivaprakash, M.K. Evaluation of three Spirulina species grown under similar conditions for their growth and biochemicals. J. Sci. Food Agric. 2005, 85, 333–336. [Google Scholar] [CrossRef]
- Sakai, N.; Sakamoto, Y.; Kishimoto, N.; Chihara, M.; Karube, I. Chlorella strains from hot springs tolerant to high temperature and high CO2. Energy Convers. Manag. 1995, 36, 693–696. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Duan, C.; Yi, S.; Gao, Z.; Xiao, C.; Agathos, S.N.; Wang, G.; Li, J. Biotechnological production of astaxanthin from the microalga Haematococcus pluvialis. Biotechnol. Adv. 2020, 43, 107602. [Google Scholar] [CrossRef]
- Lu, Q.; Li, H.; Xiao, Y.; Liu, H. A state-of-the-art review on the synthetic mechanisms, production technologies, and practical application of polyunsaturated fatty acids from microalgae. Algal Res. 2021, 55, 102281. [Google Scholar] [CrossRef]
- Schreiber, U. Detection of rapid induction kinetics with a new type of high-frequency modulated chlorophyll fluorometer. In Current Topics in Photosynthesis: Dedicated to Professor L.N.M. Duysens on the Occasion of His Retirement; Amesz, J., Hoff, A.J., Van Gorkum, H.J., Eds.; Springer: Dordrecht, The Netherlands, 1986; pp. 259–270. [Google Scholar]
- Schreiber, U.; Bilger, W. Rapid assessment of stress effects on plant leaves by chlorophyll fluorescence measurements. In Proceedings of the Plant Response to Stress, held in Sesimbra, Portugal, October 1985; Tenhunen, J.D., Catarino, F.M., Lange, O.L., Oechel, W.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; pp. 27–53. [Google Scholar]
- Schreiber, U.; Endo, T.; Mi, H.; Asada, K. Quenching Analysis of Chlorophyll Fluorescence by the Saturation Pulse Method: Particular Aspects Relating to the Study of Eukaryotic Algae and Cyanobacteria. Plant Cell Physiol. 1995, 36, 873–882. [Google Scholar] [CrossRef]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Annu. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [Green Version]
- Malapascua, J.R.F.; Jerez, C.; Sergejevová, M.; Lopez Figueroa, F.; Masojídek, J. Photosynthesis monitoring to optimize growth of microalgal mass cultures: Application of chlorophyll fluorescence techniques. Aquat. Biol. 2014, 22, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Masojídek, J.; Torzillo, G.; Koblížek, M. Photosynthesis in Microalgae. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 21–36. [Google Scholar]
- Masojídek, J.; Vonshak, A.; Torzillo, G. Chlorophyll Fluorescence Applications in Microalgal Mass Cultures. In Chlorophyll a Fluorescence in Aquatic Sciences: Methods and Applications; Suggett, D.J., Prášil, O., Borowitzka, M.A., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 277–292. [Google Scholar]
- Sukenik, A.; Beardall, J.; Kromkamp, J.C.; Kopecký, J.; Masojídek, J.; Bergeijk, S.A.v.; Gabai, S.; Shaham, E.; Yamshon, A. Photosynthetic performance of outdoor Nannochloropsis mass cultures under a wide range of environmental conditions. Aquat. Microb. Ecol. 2009, 56, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Butterwick, C.; Heaney, S.I.; Talling, J.F. A comparison of eight methods for estimating the biomass and growth of planktonic algae. Br. Phycol. J. 1982, 17, 69–79. [Google Scholar] [CrossRef]
- Bespalov, A.; Steckler, T.; Skolnick, P. Be positive about negatives–recommendations for the publication of negative (or null) results. Eur. Neuropsychopharmacol. 2019, 29, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Fanelli, D. Negative results are disappearing from most disciplines and countries. Scientometrics 2012, 90, 891–904. [Google Scholar] [CrossRef]
- Matosin, N.; Frank, E.; Engel, M.; Lum, J.S.; Newell, K.A. Negativity towards negative results: A discussion of the disconnect between scientific worth and scientific culture. Dis. Model Mech. 2014, 7, 171–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senger, H. Charakterisierung einer Synchronkultur von Scenedesmus obliquus, ihrer potentiellen Photosyntheseleistung und des Photosynthese-Quotienten während des Entwicklungscyclus. Planta 1970, 90, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Stepanov, S.S.; Zolotareva, E.K. Methanol-induced stimulation of growth, intracellular amino acids, and protein content in Chlamydomonas reinhardtii. J. Appl. Phycol. 2015, 27, 1509–1516. [Google Scholar] [CrossRef]
- Wetzel, R.G.; Linkens, G.E. Limnological Analysis; Springer: New York, NY, USA, 1991. [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der quantitativen Phytoplankton Methodik. Int. Ver. Theor. Angew. Limnol. Mitt. 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Rott, E. Some results from phytoplankton counting intercalibrations. Schweiz. Z. Hydrol. 1981, 43, 34–62. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.-D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Humphrey, G.F. New spectrophotometric equations for determining chlorophylls a, b, c1 and c2 in higher plants, algae and natural phytoplankton. Biochem. Physiol. Pflanz. 1975, 167, 191–194. [Google Scholar] [CrossRef]
- Lorenzen, C.J. A method for the continuous measurement of in vivo chlorophyll concentration. Deep. Sea Res. Oceanogr. Abstr. 1966, 13, 223–227. [Google Scholar] [CrossRef]
- Schuurmans, R.M.; van Alphen, P.; Schuurmans, J.M.; Matthijs, H.C.P.; Hellingwerf, K.J. Comparison of the Photosynthetic Yield of Cyanobacteria and Green Algae: Different Methods Give Different Answers. PLoS ONE 2015, 10, e0139061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misumi, M.; Katoh, H.; Tomo, T.; Sonoike, K. Relationship Between Photochemical Quenching and Non-Photochemical Quenching in Six Species of Cyanobacteria Reveals Species Difference in Redox State and Species Commonality in Energy Dissipation. Plant Cell Physiol. 2016, 57, 1510–1517. [Google Scholar] [CrossRef] [PubMed]
- Campbell, D.; Hurry, V.; Clarke, A.K.; Gustafsson, P.; Oquist, G. Chlorophyll fluorescence analysis of cyanobacterial photosynthesis and acclimation. Microbiol. Mol. Biol. Rev. 1998, 62, 667–683. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Misumi, M.; Sonoike, K. Estimation of photosynthesis in cyanobacteria by pulse-amplitude modulation chlorophyll fluorescence: Problems and solutions. Photosynth. Res. 2017, 133, 63–73. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic Assignments, Strain Histories and Properties of Pure Cultures of Cyanobacteria. Microbiology 1979, 111, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Zarrouk, C. Contribution a l’etude d’une Cyanophycee. Influence de Divers’ Facteurs Physiques et Chimiques Sur La Croissance et La Photosynthese de Spirulina maxima (Setch et Gardner) Geitler. Ph.D. Thesis, Universit´e De Paris, Paris, France, 1966. [Google Scholar]
- Nalewajko, C. Dry Weight, Ash, and Volume Data for Some Freshwater Planktonic Algae. J. Fish. Res. Board Can. 1966, 23, 1285–1288. [Google Scholar] [CrossRef]
- Weil, J.; Trudel, M.; Tucker, S.; Brodeur, R.D.; Juanes, F. Percent ash-free dry weight as a robust method to estimate energy density across taxa. Ecol. Evol. 2019, 9, 13244–13254. [Google Scholar] [CrossRef] [PubMed]
- Apha. Standard Methods for the Examination of Water, 22nd ed.; American Public Health Association: Albany, NY, USA, 2012. [Google Scholar]
- Schagerl, M.; Angel, R.; Donabaum, U.; Gschwandner, A.; Woebken, D. Limnospira fusiformis harbors dinitrogenase reductase (nifH)-like genes, but does not show N2 fixation activity. Algal Res. 2022, 66, 102771. [Google Scholar] [CrossRef]
- Gosselain, V.; Hamilton, P.B.; Descy, J.-P. Estimating phytoplankton carbon from microscopic counts: An application for riverine systems. Hydrobiologia 2000, 438, 75–90. [Google Scholar] [CrossRef]
- Montagnes, D.J.S.; Berges, J.A.; Harrison, P.J.; Taylor, F.J.R. Estimating carbon, nitrogen, protein, and chlorophyll a from volume in marine phytoplankton. Limnol. Oceanogr. 1994, 39, 1044–1060. [Google Scholar] [CrossRef] [Green Version]
- Rocha, O.; Duncan, A. The relationship between cell carbon and cell volume in freshwater algal species used in zooplanktonic studies. J. Plankton Res. 1985, 7, 279–294. [Google Scholar] [CrossRef]
- Menden-Deuer, S.; Lessard, E.J. Carbon to volume relationships for dinoflagellates, diatoms, and other protist plankton. Limnol. Oceanogr. 2000, 45, 569–579. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.P.; Monteiro, C.M.; Malcata, F.X. Simultaneous effect of irradiance and temperature on biochemical composition of the microalga Pavlova lutheri. J. Appl. Phycol. 2009, 21, 543–552. [Google Scholar] [CrossRef]
- da Silva Ferreira, V.; Sant’Anna, C. Impact of culture conditions on the chlorophyll content of microalgae for biotechnological applications. World J. Microbiol. Biotechnol. 2016, 33, 20. [Google Scholar] [CrossRef] [PubMed]
- Canfield, D.E., Jr.; Linda, S.B.; Hodgson, L.M. Chlorophyll-biomass-nutrient relationships for natural assemblages of Florida phytoplankton. JAWRA J. Am. Water Resour. Assoc. 1985, 21, 381–391. [Google Scholar] [CrossRef]
- Kasprzak, P.; Padisák, J.; Koschel, R.; Krienitz, L.; Gervais, F. Chlorophyll a concentration across a trophic gradient of lakes: An estimator of phytoplankton biomass? Limnologica 2008, 38, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Schagerl, M.; Künzl, G. Chlorophyll a extraction from freshwater algae—A reevaluation. Biologia 2007, 62, 270–275. [Google Scholar] [CrossRef]
- Pinckney, J.; Papa, R.; Zingmark, R. Comparison of high-performance liquid chromatographic, spectrophotometric, and fluorometric methods for determining chlorophyll a concentrations in estaurine sediments. J. Microbiol. Methods 1994, 19, 59–66. [Google Scholar] [CrossRef]
- Simon, D.; Helliwell, S. Extraction and quantification of chlorophyll a from freshwater green algae. Water Res. 1998, 32, 2220–2223. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. 4.1—Determination of Chlorophylls and Total Carotenoids: Spectrophotometric Method. In A Manual of Chemical & Biological Methods for Seawater Analysis; Parsons, T.R., Maita, Y., Lalli, C.M., Eds.; Pergamon: Amsterdam, The Netherlands, 1984; pp. 101–104. [Google Scholar]
- Poikāne, S.; Alves, M.H.; Argillier, C.; van den Berg, M.; Buzzi, F.; Hoehn, E.; de Hoyos, C.; Karottki, I.; Laplace-Treyture, C.; Solheim, A.L.; et al. Defining Chlorophyll-a Reference Conditions in European Lakes. Environ. Manag. 2010, 45, 1286–1298. [Google Scholar] [CrossRef] [Green Version]
- Mountourakis, F.; Papazi, A.; Kotzabasis, K. The Microalga Chlorella vulgaris as a Natural Bioenergetic System for Effective CO2 Mitigation—New Perspectives against Global Warming. Symmetry 2021, 13, 997. [Google Scholar] [CrossRef]
- Takeda, H.; Hirokawa, T. Cell-Wall Chemistry, Structure and Components. In Cell Components; Linskens, H.-F., Jackson, J.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1985; pp. 31–53. [Google Scholar]
- Tsuzuki, M.; Ohnuma, E.; Sato, N.; Takaku, T.; Kawaguchi, A. Effects of CO(2) Concentration during Growth on Fatty Acid Composition in Microalgae. Plant Physiol. 1990, 93, 851–856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahnke, J.; Mahlmann, D.M.; Jacobs, P.; Priefer, U.B. The influence of growth conditions on the cell dry weight per unit biovolume of Klebsormidium flaccidum (Charophyta), a typical ubiquitous soil alga. J. Appl. Phycol. 2011, 23, 655–664. [Google Scholar] [CrossRef]
- Jahnke, J.; Mahlmann, D.M. Differences in the cellular dry weight per unit biovolume of Phormidium autumnale (Cyanobacteria) dependent on growth conditions. J. Appl. Phycol. 2010, 22, 117–122. [Google Scholar] [CrossRef]
- Vonshak, A. Microalgae: Laboratory growth techniques and the biotechnology of biomass production. In Photosynthesis and Production in a Changing Environment: A Field and Laboratory Manual; Hall, D.O., Scurlock, J.M.O., Bolhàr-Nordenkampf, H.R., Leegood, R.C., Long, S.P., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 337–355. [Google Scholar]
- Hyka, P.; Lickova, S.; Přibyl, P.; Melzoch, K.; Kovar, K. Flow cytometry for the development of biotechnological processes with microalgae. Biotechnol. Adv. 2013, 31, 2–16. [Google Scholar] [CrossRef] [Green Version]
- Chioccioli, M.; Hankamer, B.; Ross, I.L. Flow Cytometry Pulse Width Data Enables Rapid and Sensitive Estimation of Biomass Dry Weight in the Microalgae Chlamydomonas reinhardtii and Chlorella vulgaris. PLoS ONE 2014, 9, e97269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, T. Routine Management of Microalgae Using Autofluorescence from Chlorophyll. Molecules 2019, 24, 4441. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T.L. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef]
- Santos-Ballardo, D.U.; Rossi, S.; Hernández, V.; Gómez, R.V.; del Carmen Rendón-Unceta, M.; Caro-Corrales, J.; Valdez-Ortiz, A. A simple spectrophotometric method for biomass measurement of important microalgae species in aquaculture. Aquaculture 2015, 448, 87–92. [Google Scholar] [CrossRef]
- Srinivas, S.P.; Mutharasan, R. Inner filter effects and their interferences in the interpretation of culture fluorescence. Biotechnol. Bioeng. 1987, 30, 769–774. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, M.-S.; Han, B.; Xing, X.-H. Correcting for the inner filter effect in measurements of fluorescent proteins in high-cell-density cultures. Anal. Biochem. 2009, 390, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Panigrahi, S.K.; Mishra, A.K. Study on the dependence of fluorescence intensity on optical density of solutions: The use of fluorescence observation field for inner filter effect corrections. Photochem. Photobiol. Sci. 2019, 18, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Falkowski, P.; Kiefer, D.A. Chlorophyll a fluorescence in phytoplankton: Relationship to photosynthesis and biomass. J. Plankton Res. 1985, 7, 715–731. [Google Scholar] [CrossRef]
- Catherine, A.; Escoffier, N.; Belhocine, A.; Nasri, A.B.; Hamlaoui, S.; Yéprémian, C.; Bernard, C.; Troussellier, M. On the use of the FluoroProbe®, a phytoplankton quantification method based on fluorescence excitation spectra for large-scale surveys of lakes and reservoirs. Water Res. 2012, 46, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Kruskopf, M.; Flynn, K.J. Chlorophyll content and fluorescence responses cannot be used to gauge reliably phytoplankton biomass, nutrient status or growth rate. New Phytol. 2006, 169, 525–536. [Google Scholar] [CrossRef]
- Rolton, A.; McCullough, A.; Tuckey, N.P.L.; Finnie, B.; Cooper, I.; Packer, M.A.; Vignier, J. Early biomarker indicators of health in two commercially produced microalgal species important for aquaculture. Aquaculture 2020, 521, 735053. [Google Scholar] [CrossRef]
- Spilling, K.; Seppälä, J. Measurement of Fluorescence for Monitoring Algal Growth and Health. In Biofuels from Algae: Methods and Protocols; Spilling, K., Ed.; Springer: New York, NY, USA, 2020; pp. 41–45. [Google Scholar]
- Bemejo-Padilla, E.; Kinsou, H.; Filali, R.; Perez-Bibbins, B.; Taidi, B. Rapid indicators for monitoring the health of Chlamydomonas nivalis biomass during preservation. J. Appl. Phycol. 2021, 33, 2723–2732. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Causally Linked to Biomass—Absolute Units | Dry Mass |
|---|---|
| Derived—absolute units | Biovolume Packed cell volume Cell number Carbon content In vitro Chl-a |
| Derived—relative units | Optical density In vivo fluorescence |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schagerl, M.; Siedler, R.; Konopáčová, E.; Ali, S.S. Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements. Cells 2022, 11, 2455. https://doi.org/10.3390/cells11152455
Schagerl M, Siedler R, Konopáčová E, Ali SS. Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements. Cells. 2022; 11(15):2455. https://doi.org/10.3390/cells11152455
Chicago/Turabian StyleSchagerl, Michael, Rainer Siedler, Eliška Konopáčová, and Sameh Samir Ali. 2022. "Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements" Cells 11, no. 15: 2455. https://doi.org/10.3390/cells11152455
APA StyleSchagerl, M., Siedler, R., Konopáčová, E., & Ali, S. S. (2022). Estimating Biomass and Vitality of Microalgae for Monitoring Cultures: A Roadmap for Reliable Measurements. Cells, 11(15), 2455. https://doi.org/10.3390/cells11152455