
Static Magnetic Fields Regulate T-Type Calcium Ion Channels and Mediate Mesenchymal Stem Cells Proliferation

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
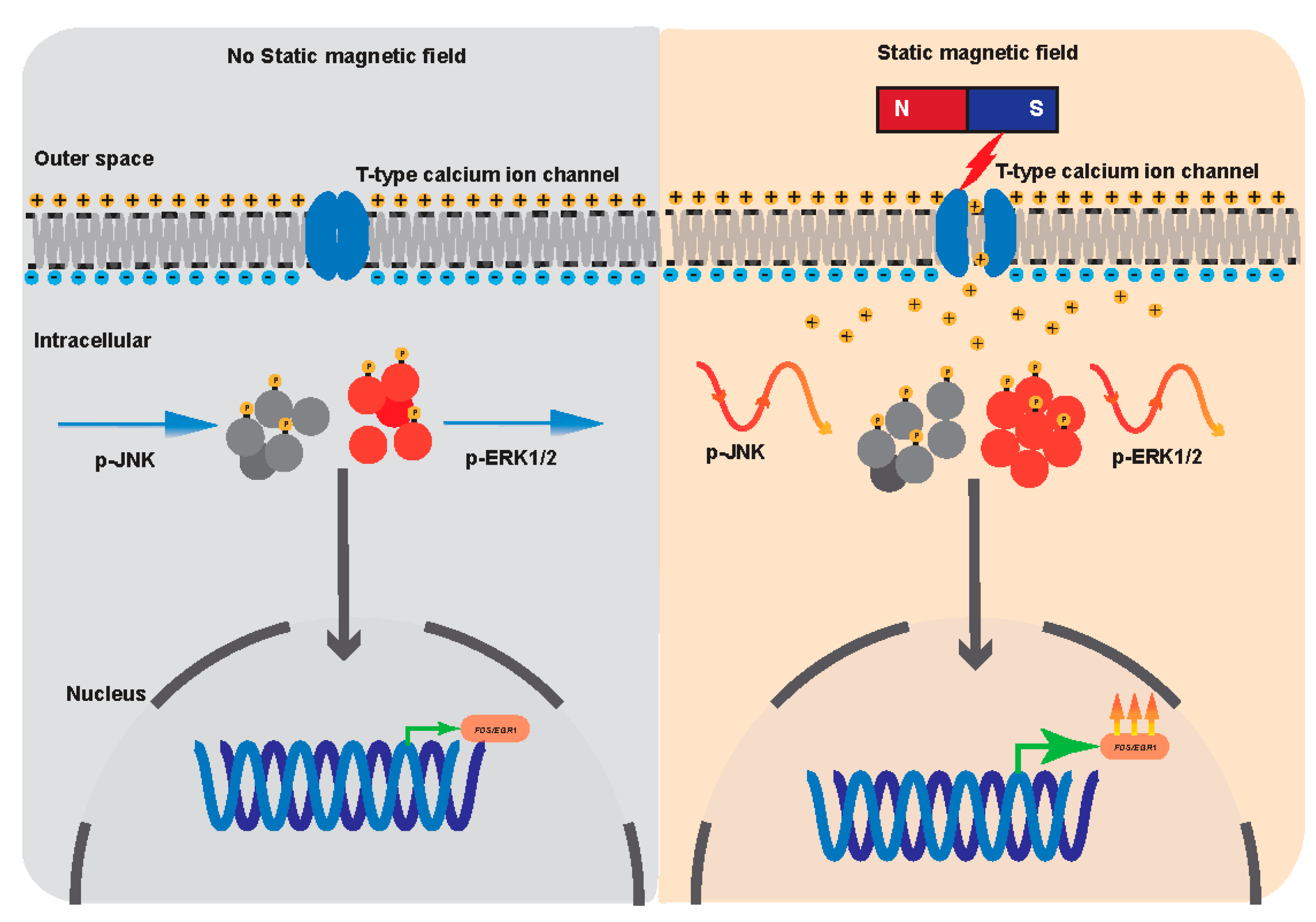
Abstract
:1. Introduction
2. Materials and Methods
2.1. MSC Culture
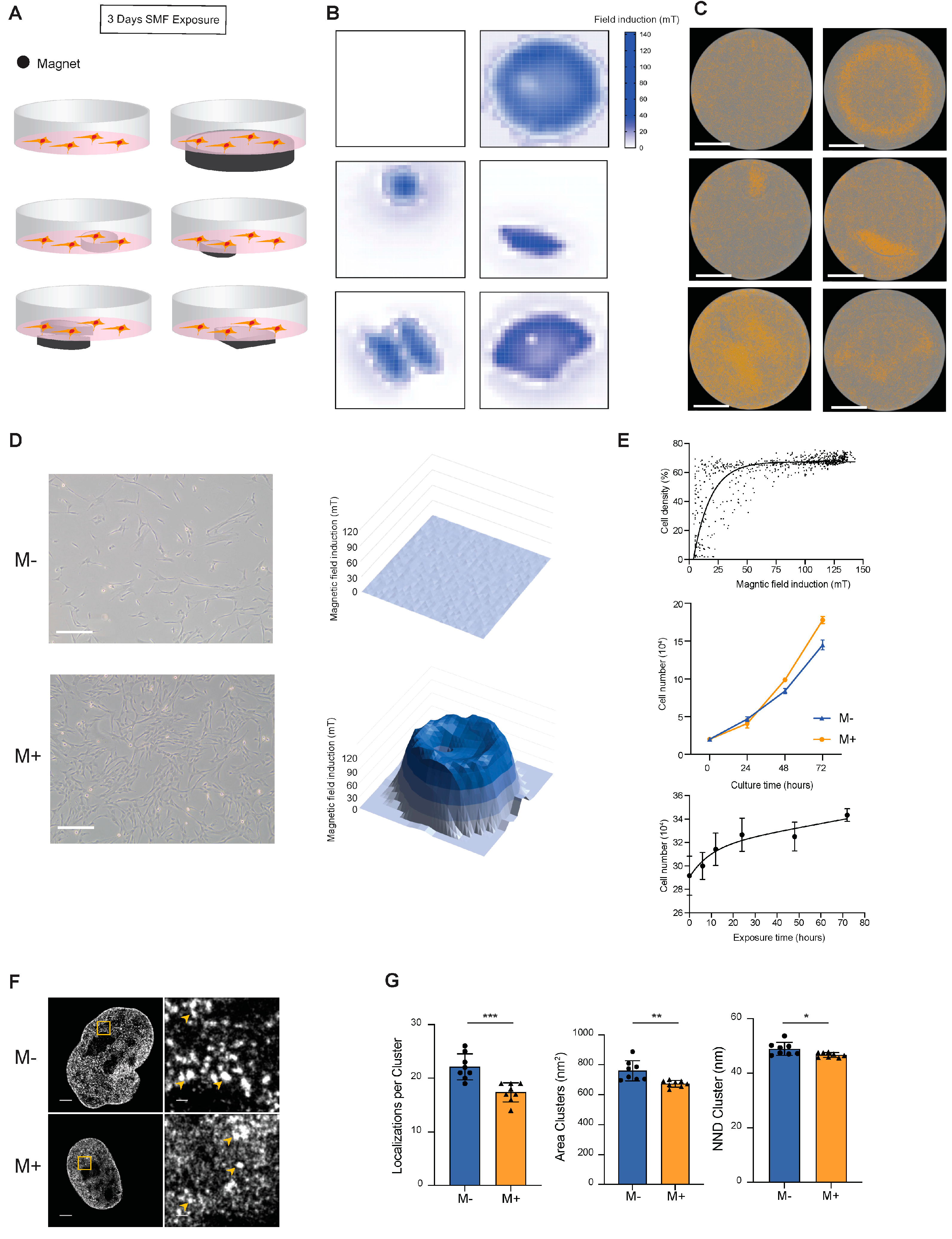
2.2. STORM Images Acquisition and Analysis
2.3. RNA-Seq and Data Analysis
2.4. Quantitative RT-PCR
2.5. Western Blot Analysis
2.6. Flow Cytometry
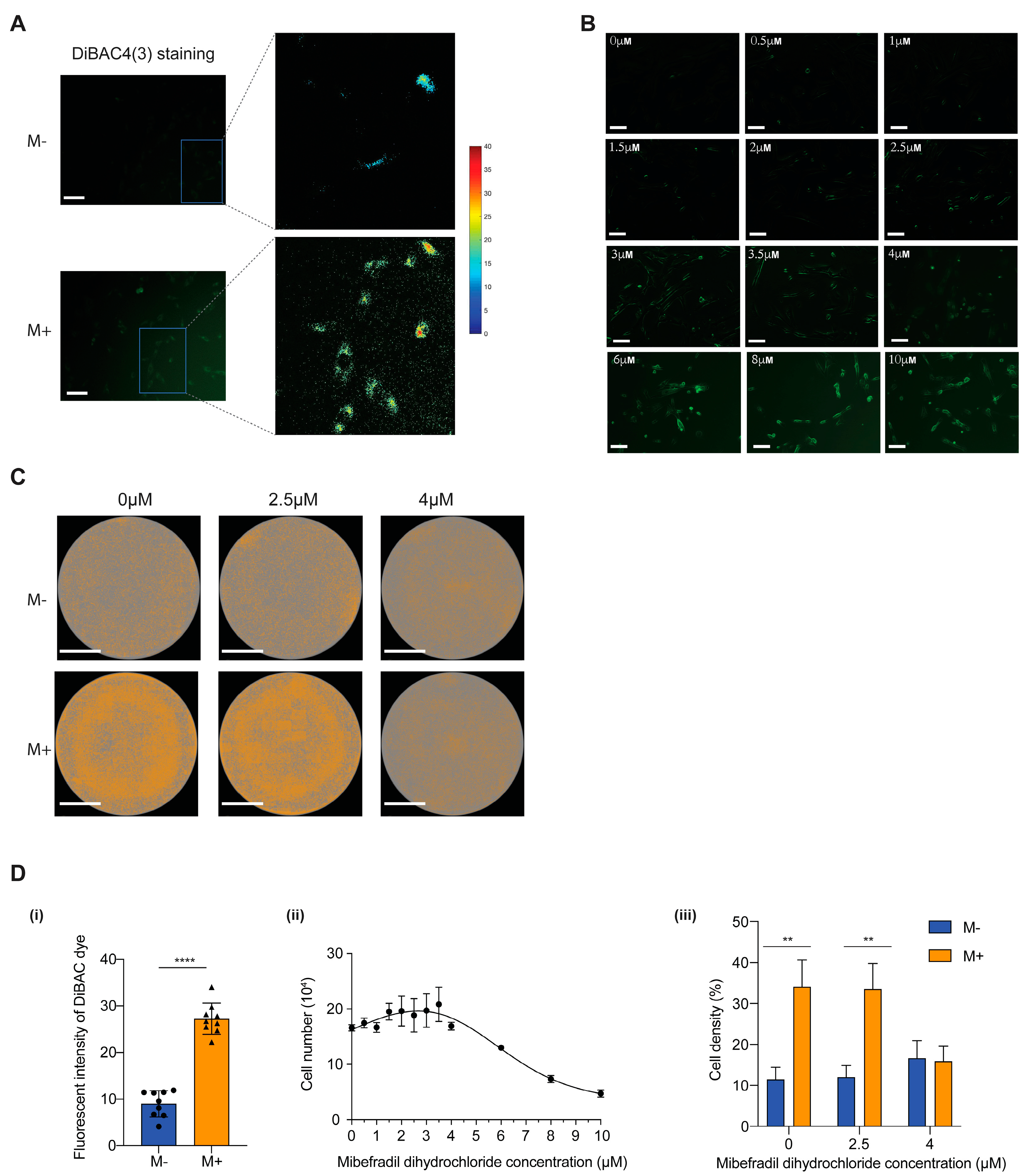
2.7. Measuring Resting Membrane Potential Using the Fluorescent Voltage Reporters DiBAC4(3)
2.8. Statistical Information
3. Results
3.1. Static Magnetic Field Promotes the Proliferation of Mesenchymal Stem Cells
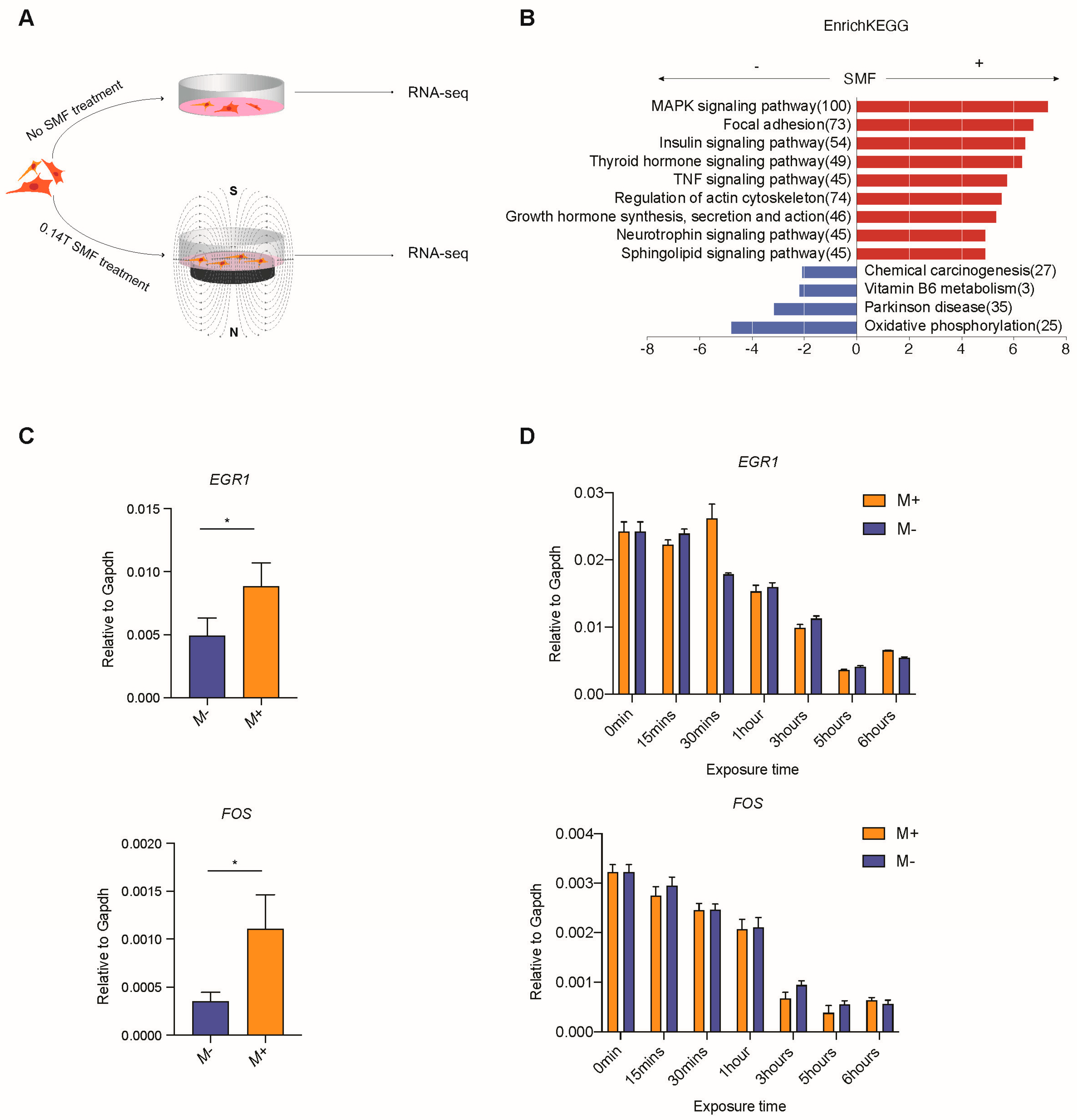
3.2. Exposure to SMF Upregulates Immunoregulatory Factor Genes and MAPK-Signal-Pathway-Related Genes in Mesenchymal Stem Cells
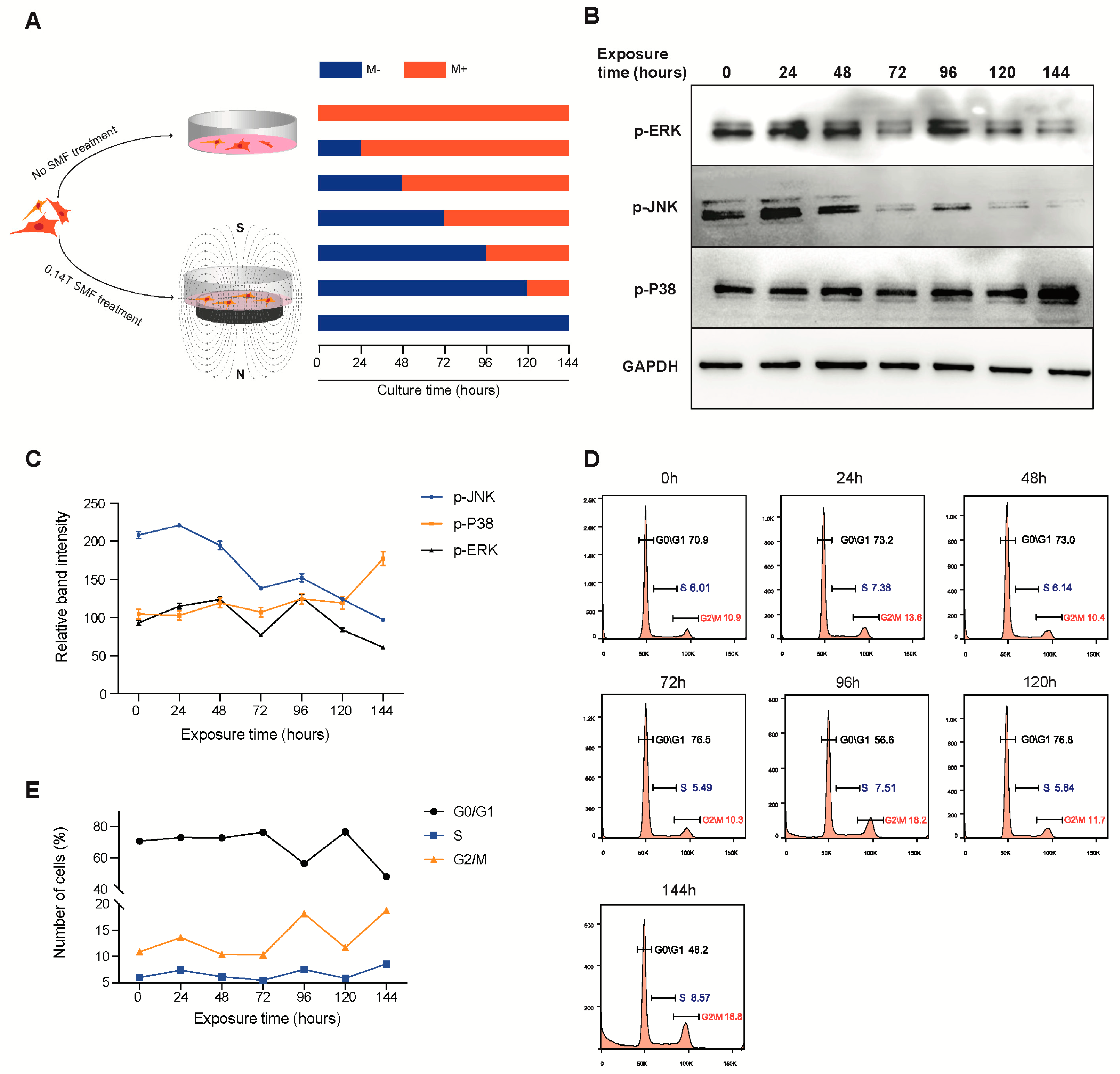
3.3. SMF Exposure Time Modulates the Activation of MAPK Protein in Mesenchymal Stem Cells
3.4. SMF Effecting Transmembrane Depolarization via T-Type Calcium Ion Channel
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marycz, K.; Kornicka, K.; Rocken, M. Static Magnetic Field (SMF) as a Regulator of Stem Cell Fate—New Perspectives in Regenerative Medicine Arising from an Underestimated Tool. Stem Cell Rev. Rep. 2018, 14, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Blumler, P. Magnetic Guiding with Permanent Magnets: Concept, Realization and Applications to Nanoparticles and Cells. Cells 2021, 10, 2708. [Google Scholar] [CrossRef] [PubMed]
- Sliker, L.; Ciuti, G.; Rentschler, M.; Menciassi, A. Magnetically driven medical devices: A review. Expert Rev. Med. Devices 2015, 12, 737–752. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, F.; Chao, Y.; Yao, L. Bioinspired soft microrobots actuated by magnetic field. Biomed. Microdevices 2021, 23, 52. [Google Scholar] [CrossRef] [PubMed]
- Markov, M.S. Expanding use of pulsed electromagnetic field therapies. Electromagn. Biol. Med. 2007, 26, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Zwolinska, J.; Gąsior, M.; Śnieżek, E.; Kwolek, A. The use of magnetic fields in treatment of patients with rheumatoid arthritis. Review of the literature. Reumatologia 2016, 54, 201–206. [Google Scholar] [CrossRef]
- Rumbaut, R.E.; Mirkovic, D. Magnetic therapy for edema in inflammation: A physiological assessment. Am. J. Physiol. Heart Circ. Physiol. 2008, 294, H19–H20. [Google Scholar] [CrossRef]
- Gmitrov, J.; Ohkubo, C.; Okano, H. Effect of 0.25 T static magnetic field on microcirculation in rabbits. Bioelectromagnetics 2002, 23, 224–229. [Google Scholar] [CrossRef]
- Xu, S.; Okano, H.; Ohkubo, C. Acute effects of whole-body exposure to static magnetic fields and 50-Hz electromagnetic fields on muscle microcirculation in anesthetized mice. Bioelectrochemistry 2001, 53, 127–135. [Google Scholar] [CrossRef]
- Hirose, H.; Nakahara, T.; Miyakoshi, J. Orientation of human glioblastoma cells embedded in type I collagen, caused by exposure to a 10 T static magnetic field. Neurosci. Lett. 2003, 338, 88–90. [Google Scholar] [CrossRef]
- Wang, J.; Xiang, B.; Deng, J.; Freed, D.H.; Arora, R.C.; Tian, G. Inhibition of Viability, Proliferation, Cytokines Secretion, Surface Antigen Expression, and Adipogenic and Osteogenic Differentiation of Adipose-Derived Stem Cells by Seven-Day Exposure to 0.5 T Static Magnetic Fields. Stem Cells Int. 2016, 2016, 7168175. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, L.; Chen, M.Q.; Zhang, Y.; Li, J.; Feng, X.; Li, W.; Guo, W.; Jia, G.; Wang, H.; et al. Low strength static magnetic field inhibits the proliferation, migration, and adhesion of human vascular smooth muscle cells in a restenosis model through mediating integrins beta1-FAK, Ca2+ signaling pathway. Ann. Biomed. Eng. 2012, 40, 2611–2618. [Google Scholar] [CrossRef] [PubMed]
- Romeo, S.; Sannino, A.; Scarfì, M.R.; Massa, R.; d’Angelo, R.; Zeni, O. Lack of effects on key cellular parameters of MRC-5 human lung fibroblasts exposed to 370 mT static magnetic field. Sci. Rep. 2016, 6, 19398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lew, W.Z.; Huang, Y.-C.; Huang, K.-Y.; Lin, C.-T.; Tsai, M.-T.; Huang, H.-M. Static magnetic fields enhance dental pulp stem cell proliferation by activating the p38 mitogen-activated protein kinase pathway as its putative mechanism. J. Tissue Eng. Regen. Med. 2018, 12, 19–29. [Google Scholar] [CrossRef]
- Zhang, X.; Yarema, K.; Xu, A. Biological Effects of Static Magnetic Fields; Springer: Singapore, 2017. [Google Scholar]
- Yu, B.; Liu, J.; Cheng, J.; Zhang, L.; Song, C.; Tian, X.; Fan, Y.; Lv, Y.; Zhang, X. A Static Magnetic Field Improves Iron Metabolism and Prevents High-Fat-Diet/Streptozocin-Induced Diabetes. Innovation 2021, 2, 100077. [Google Scholar] [CrossRef]
- Kornicka, K.; Marycz, K.; Marędziak, M.; Tomaszewski, K.A.; Nicpoń, J. The effects of the DNA methyltranfserases inhibitor 5-Azacitidine on ageing, oxidative stress and DNA methylation of adipose derived stem cells. J. Cell Mol. Med. 2017, 21, 387–401. [Google Scholar] [CrossRef]
- Kornicka, K.; Marycz, K.; Tomaszewski, K.A.; Marędziak, M.; Śmieszek, A. The Effect of Age on Osteogenic and Adipogenic Differentiation Potential of Human Adipose Derived Stromal Stem Cells (hASCs) and the Impact of Stress Factors in the Course of the Differentiation Process. Oxid. Med. Cell Longev. 2015, 2015, 309169. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Ghaderi, A.; Abtahi, S. Mesenchymal Stem Cells: Miraculous Healers or Dormant Killers? Stem Cell Rev. Rep. 2018, 14, 722–733. [Google Scholar] [CrossRef]
- Hielscher, D.; Kaebisch, C.; Braun, B.J.V.; Gray, K.; Tobiasch, E. Stem Cell Sources and Graft Material for Vascular Tissue Engineering. Stem Cell Rev. Rep. 2018, 14, 642–667. [Google Scholar] [CrossRef]
- Cislo-Pakuluk, A.; Marycz, K. A Promising Tool in Retina Regeneration: Current Perspectives and Challenges When Using Mesenchymal Progenitor Stem Cells in Veterinary and Human Ophthalmological Applications. Stem Cell Rev. Rep. 2017, 13, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zablotskii, V.; Dejneka, A.; Kubinová, Š.; Le-Roy, D.; Dumas-Bouchiat, F.; Givord, D.; Dempsey, N.M.; Syková, E. Life on magnets: Stem cell networking on micro-magnet arrays. PLoS ONE 2013, 8, e70416. [Google Scholar]
- Sadri, M.; Abdolmaleki, P.; Abrun, S.; Beiki, B.; Samani, F.S. Static Magnetic Field Effect on Cell Alignment, Growth, and Differentiation in Human Cord-Derived Mesenchymal Stem Cells. Cell Mol. Bioeng. 2017, 10, 249–262. [Google Scholar] [CrossRef]
- Bates, M.; Huang, B.; Dempsey, G.T.; Zhuang, X. Multicolor super-resolution imaging with photo-switchable fluorescent probes. Science 2007, 317, 1749–1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricci, M.A.; Manzo, C.; García-Parajo, M.F.; Lakadamyali, M.; Cosma, M.P. Chromatin fibers are formed by heterogeneous groups of nucleosomes in vivo. Cell 2015, 160, 1145–1158. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.S.; Levin, M. General principles for measuring resting membrane potential and ion concentration using fluorescent bioelectricity reporters. Cold Spring Harb. Protoc. 2012, 2012, 385–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, D.S.; Levin, M. Measuring resting membrane potential using the fluorescent voltage reporters DiBAC4(3) and CC2-DMPE. Cold Spring Harb. Protoc. 2012, 2012, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Otterstrom, J.; Castells-Garcia, A.; Vicario, C.; Gomez-Garcia, P.A.; Cosma, M.P.; Lakadamyali, M. Super-resolution microscopy reveals how histone tail acetylation affects DNA compaction within nucleosomes in vivo. Nucleic Acids Res. 2019, 47, 8470–8484. [Google Scholar] [CrossRef] [Green Version]
- Castells-Garcia, A.; Ed-Daoui, I.; González-Almela, E.; Vicario, C.; Ottestrom, J.; Lakadamyali, M.; Neguembor, M.V.; Cosma, M.P. Super resolution microscopy reveals how elongating RNA polymerase II and nascent RNA interact with nucleosome clutches. Nucleic Acids Res. 2022, 50, 175–190. [Google Scholar] [CrossRef]
- Chen, S.X.; Zhao, F.; Huang, X.J. MAPK signaling pathway and erectile dysfunction. Zhonghua Nan Ke Xue 2018, 24, 442–446. [Google Scholar]
- Zablotskii, V.; Polyakova, T.; Dejneka, A. Cells in the Non-Uniform Magnetic World: How Cells Respond to High-Gradient Magnetic Fields. Bioessays 2018, 40, e1800017. [Google Scholar] [CrossRef] [PubMed]
- Zablotskii, V.; Polyakova, T.; Lunov, O.; Dejneka, A. How a High-Gradient Magnetic Field Could Affect Cell Life. Sci. Rep. 2016, 6, 37407. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.R.; Yang, J.W.; Lee, Y.T.; Tsai, L.Y. Static magnetic field expose enhances neurotransmission in crayfish nervous system. Int. J. Radiat. Biol. 2008, 84, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Sundelacruz, S.; Levin, M.; Kaplan, D.L. Role of membrane potential in the regulation of cell proliferation and differentiation. Stem Cell Rev. Rep. 2009, 5, 231–246. [Google Scholar] [CrossRef] [PubMed]
- Catterall, W.A. Voltage-gated calcium channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef] [PubMed]
- Jia, C.; Zhou, Z.; Liu, R.; Chen, S.; Xia, R. EGF receptor clustering is induced by a 0.4 mT power frequency magnetic field and blocked by the EGF receptor tyrosine kinase inhibitor PD153035. Bioelectromagnetics 2007, 28, 197–207. [Google Scholar] [CrossRef]
- Simko, M. Cell type specific redox status is responsible for diverse electromagnetic field effects. Curr. Med. Chem. 2007, 14, 1141–1152. [Google Scholar] [CrossRef]
- Sun, W.; Shen, X.; Lu, D.; Lu, D.; Chiang, H. Superposition of an incoherent magnetic field inhibited EGF receptor clustering and phosphorylation induced by a 1.8 GHz pulse-modulated radiofrequency radiation. Int. J. Radiat. Biol. 2013, 89, 378–383. [Google Scholar] [CrossRef]
- Prina-Mello, A.; Farrell, E.; Prendergast, P.J.; Campbell, V.; Coey, J.M.D. Influence of strong static magnetic fields on primary cortical neurons. Bioelectromagnetics 2006, 27, 35–42. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Minami, I.; Kadotani, H.; Todo, T.; Nishida, E. Circadian clock-controlled diurnal oscillation of Ras/ERK signaling in mouse liver. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2013, 89, 59–65. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.; Choi, Y.; Lee, K.; Krishnan, V.; Pelled, G.; Gilad, A.A.; Choi, J. Regulation of Electromagnetic Perceptive Gene Using Ferromagnetic Particles for the External Control of Calcium Ion Transport. Biomolecules 2020, 10, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baureus Koch, C.L.; Sommarin, M.; Persson, B.R.R.; Salford, L.G.; Eberhardt, J.L. Interaction between weak low frequency magnetic fields and cell membranes. Bioelectromagnetics 2003, 24, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Dong, L.; Zhang, B.; Qi, N. Effects of extremely low-frequency magnetic field on growth and differentiation of human mesenchymal stem cells. Electromagn. Biol. Med. 2010, 29, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Kinouchi, Y.; Tanimoto, S.; Ushita, T.; Sato, K.; Yamaguchi, H.; Miyamoto, H. Effects of static magnetic fields on diffusion in solutions. Bioelectromagnetics 1988, 9, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Polyakova, T.; Zablotskii, V.; Dejneka, A. Cell Membrane Pore Formation and Change in Ion Channel Activity in High-gradient Magnetic Fields. IEEE Magn. Lett. 2017, 8, 1–5. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.; Li, C.; Masood, M.; Zhang, Z.; González-Almela, E.; Castells-Garcia, A.; Zou, G.; Xu, X.; Wang, L.; Zhao, G.; et al. Static Magnetic Fields Regulate T-Type Calcium Ion Channels and Mediate Mesenchymal Stem Cells Proliferation. Cells 2022, 11, 2460. https://doi.org/10.3390/cells11152460
Wu H, Li C, Masood M, Zhang Z, González-Almela E, Castells-Garcia A, Zou G, Xu X, Wang L, Zhao G, et al. Static Magnetic Fields Regulate T-Type Calcium Ion Channels and Mediate Mesenchymal Stem Cells Proliferation. Cells. 2022; 11(15):2460. https://doi.org/10.3390/cells11152460
Chicago/Turabian StyleWu, Haokaifeng, Chuang Li, Muqaddas Masood, Zhen Zhang, Esther González-Almela, Alvaro Castells-Garcia, Gaoyang Zou, Xiaoduo Xu, Luqin Wang, Guoqing Zhao, and et al. 2022. "Static Magnetic Fields Regulate T-Type Calcium Ion Channels and Mediate Mesenchymal Stem Cells Proliferation" Cells 11, no. 15: 2460. https://doi.org/10.3390/cells11152460
APA StyleWu, H., Li, C., Masood, M., Zhang, Z., González-Almela, E., Castells-Garcia, A., Zou, G., Xu, X., Wang, L., Zhao, G., Yu, S., Zhu, P., Wang, B., Qin, D., & Liu, J. (2022). Static Magnetic Fields Regulate T-Type Calcium Ion Channels and Mediate Mesenchymal Stem Cells Proliferation. Cells, 11(15), 2460. https://doi.org/10.3390/cells11152460