
MEG3 Expression Indicates Lymph Node Metastasis and Presence of Cancer-Associated Fibroblasts in Papillary Thyroid Cancer

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. NYMC Patient Specimens
2.2. lncRNA Annotation
2.3. BRAF Genotyping
2.4. Weighted Gene-Co-Expression Analysis (WGCNA)
2.5. Fusion Detection
2.6. TCGA Data
2.7. Pathway and GO Enrichment Analysis
2.8. Thyroid Differentiation Score and ERK Signature
2.9. Statistical Analysis
2.10. Estimating Tumor Infiltration with CAFs Using TIMER2.0
2.11. Single-Cell Anaplastic Thyroid Cancer Data
3. Results
3.1. Clinical Characteristics of PTC Cohort
3.2. Analysis of Tumor vs. Normal Transcriptomics
3.3. WGCNA Identifies a Gene Co-Expression Module Associated with LNM
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, L.; Welch, H.G. Current Thyroid Cancer Trends in the United States. JAMA Otolaryngol. Head Neck Surg. 2014, 140, 317–322. [Google Scholar] [CrossRef] [Green Version]
- Endo, M.; Nabhan, F.; Porter, K.; Roll, K.; Shirley, L.A.; Azaryan, I.; Tonkovich, D.; Perlick, J.; Ryan, L.E.; Khawaja, R.; et al. Afirma Gene Sequencing Classifier Compared with Gene Expression Classifier in Indeterminate Thyroid Nodules. Thyroid 2019, 29, 1115–1124. [Google Scholar] [CrossRef]
- Patel, K.N.; Angell, T.E.; Babiarz, J.; Barth, N.M.; Blevins, T.; Duh, Q.-Y.; Ghossein, R.A.; Harrell, R.M.; Huang, J.; Kennedy, G.C.; et al. Performance of a Genomic Sequencing Classifier for the Preoperative Diagnosis of Cytologically Indeterminate Thyroid Nodules. JAMA Surg. 2018, 153, 817–824. [Google Scholar] [CrossRef]
- Marti, J.L.; Avadhani, V.; Donatelli, L.A.; Niyogi, S.; Wang, B.; Wong, R.J.; Shaha, A.R.; Ghossein, R.A.; Lin, O.; Morris, L.G.T.; et al. Wide Inter-Institutional Variation in Performance of a Molecular Classifier for Indeterminate Thyroid Nodules. Ann. Surg. Oncol. 2015, 22, 3996–4001. [Google Scholar] [CrossRef]
- Haugen, B.R.; Alexander, E.K.; Bible, K.C.; Doherty, G.M.; Mandel, S.J.; Nikiforov, Y.E.; Pacini, F.; Randolph, G.W.; Sawka, A.M.; Schlumberger, M.; et al. 2015 American Thyroid Association Management Guidelines for Adult Patients with Thyroid Nodules and Differentiated Thyroid Cancer: The American Thyroid Association Guidelines Task Force on Thyroid Nodules and Differentiated Thyroid Cancer. Thyroid 2016, 26, 1–133. [Google Scholar] [CrossRef] [Green Version]
- Mazzaferri, E.L. Management of a Solitary Thyroid Nodule. N. Engl. J. Med. 1993, 328, 553–559. [Google Scholar]
- Mazzaferri, E.L.; Jhiang, S.M. Long-Term Impact of Initial Surgical and Medical Therapy on Papillary and Follicular Thyroid Cancer. Am. J. Med. 1994, 97, 418–428. [Google Scholar] [CrossRef]
- Podnos, Y.D.; Smith, D.; Wagman, L.D.; Ellenhorn, J.D.I. The Implication of Lymph Node Metastasis on Survival in Patients with Well-Differentiated Thyroid Cancer. Am. Surg. 2005, 71, 731–734. [Google Scholar] [CrossRef]
- Ahn, J.E.; Lee, J.H.; Yi, J.S.; Shong, Y.K.; Hong, S.J.; Lee, D.H.; Choi, C.G.; Kim, S.J. Diagnostic Accuracy of CT and Ultrasonography for Evaluating Metastatic Cervical Lymph Nodes in Patients with Thyroid Cancer. World J. Surg. 2008, 32, 1552–1558. [Google Scholar] [CrossRef]
- Jeong, H.-S.; Baek, C.-H.; Son, Y.-I.; Choi, J.-Y.; Kim, H.-J.; Ko, Y.-H.; Chung, J.-H.; Baek, H.-J. Integrated 18F-FDG PET/CT for the Initial Evaluation of Cervical Node Level of Patients with Papillary Thyroid Carcinoma: Comparison with Ultrasound and Contrast-Enhanced CT. Clin. Endocrinol. 2006, 65, 402–407. [Google Scholar] [CrossRef]
- Xia, S.; Wang, C.; Ni, X.; Ni, Z.; Dong, Y.; Zhan, W. NONHSAT076754 Aids Ultrasonography in Predicting Lymph Node Metastasis and Promotes Migration and Invasion of Papillary Thyroid Cancer Cells. Oncotarget 2017, 8, 2293–2306. [Google Scholar] [CrossRef]
- Han, P.A.; Kim, H.-S.; Cho, S.; Fazeli, R.; Najafian, A.; Khawaja, H.; McAlexander, M.; Dy, B.; Sorensen, M.; Aronova, A.; et al. Association of BRAF V600E Mutation and MicroRNA Expression with Central Lymph Node Metastases in Papillary Thyroid Cancer: A Prospective Study from Four Endocrine Surgery Centers. Thyroid 2016, 26, 532–542. [Google Scholar]
- Dutenhefner, S.E.; Marui, S.; Santos, A.B.O.; de Lima, E.U.; Inoue, M.; Neto, J.S.B.; Shiang, C.; Fukushima, J.T.; Cernea, C.R.; Friguglietti, C.U.M. BRAF: A Tool in the Decision to Perform Elective Neck Dissection? Thyroid 2013, 23, 1541–1546. [Google Scholar] [CrossRef]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long Non-Coding RNAs: Insights into Functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and Functions of Long Noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.T. Epigenetic Regulation by Long Noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [Green Version]
- Huarte, M. The Emerging Role of lncRNAs in Cancer. Nat. Med. 2015, 21, 1253–1261. [Google Scholar] [CrossRef]
- Tripathi, V.; Ellis, J.D.; Shen, Z.; Song, D.Y.; Pan, Q.; Watt, A.T.; Freier, S.M.; Bennett, C.F.; Sharma, A.; Bubulya, P.A.; et al. The Nuclear-Retained Noncoding RNA MALAT1 Regulates Alternative Splicing by Modulating SR Splicing Factor Phosphorylation. Mol. Cell 2010, 39, 925–938. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yu, S.; Jiang, L.; Wang, X.; Song, X. HOTAIR Is a Promising Novel Biomarker in Patients with Thyroid Cancer. Exp. Ther. Med. 2017, 13, 2274–2278. [Google Scholar] [CrossRef] [Green Version]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional Demarcation of Active and Silent Chromatin Domains in Human HOX Loci by Noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar] [CrossRef] [Green Version]
- Ji, P.; Diederichs, S.; Wang, W.; Böing, S.; Metzger, R.; Schneider, P.M.; Tidow, N.; Brandt, B.; Buerger, H.; Bulk, E.; et al. MALAT-1, a Novel Noncoding RNA, and Thymosin beta4 Predict Metastasis and Survival in Early-Stage Non-Small Cell Lung Cancer. Oncogene 2003, 22, 8031–8041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedaghati, M.; Kebebew, E. Long Noncoding RNAs in Thyroid Cancer. Curr. Opin. Endocrinol. Diabetes Obes. 2019, 26, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Chang, Y.; Du, W.; Liu, S.; Qi, J.; Luo, R.; Luo, S. Upregulation of lncRNA-ATB by Transforming Growth Factor β1 (TGF-β1) Promotes Migration and Invasion of Papillary Thyroid Carcinoma Cells. Med. Sci. Monit. 2018, 24, 5152–5158. [Google Scholar] [CrossRef] [PubMed]
- Goedert, L.; Plaça, J.R.; Fuziwara, C.S.; Machado, M.C.R.; Plaça, D.R.; Almeida, P.P.; Sanches, T.P.; Dos Santos, J.F.; Corveloni, A.C.; Pereira, I.E.G.; et al. Identification of Long Noncoding RNAs Deregulated in Papillary Thyroid Cancer and Correlated with BRAF V600E Mutation by Bioinformatics Integrative Analysis. Sci. Rep. 2017, 7, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, P.; Zhou, X.-Y.; Du, X. Circulating Long Non-Coding RNAs in Cancer: Current Status and Future Perspectives. Mol. Cancer 2016, 15, 39. [Google Scholar] [CrossRef] [Green Version]
- Lan, X.; Zhang, H.; Wang, Z.; Dong, W.; Sun, W.; Shao, L.; Zhang, T.; Zhang, D. Genome-Wide Analysis of Long Noncoding RNA Expression Profile in Papillary Thyroid Carcinoma. Gene 2015, 569, 109–117. [Google Scholar] [CrossRef]
- Yang, M.; Tian, J.; Guo, X.; Yang, Y.; Guan, R.; Qiu, M.; Li, Y.; Sun, X.; Zhen, Y.; Zhang, Y.; et al. Long Noncoding RNA Are Aberrantly Expressed in Human Papillary Thyroid Carcinoma. Oncol. Lett. 2016, 12, 544–552. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Shao, Q.; Xie, K.; Zhang, Y.; Dong, T.; Xia, Y.; Tang, W. The Long Non-Coding RNA ENST00000537266 and ENST00000426615 Influence Papillary Thyroid Cancer Cell Proliferation and Motility. Cell. Physiol. Biochem. 2016, 38, 368–378. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.-D.; Ye, C.-X.; Shen, Z.-L.; Yang, Y.; Wang, B.; Guo, P.; Gao, Z.-D.; Ye, Y.-J.; Jiang, K.-W.; et al. Long Noncoding RNA HIT000218960 Promotes Papillary Thyroid Cancer Oncogenesis and Tumor Progression by Upregulating the Expression of High Mobility Group AT-Hook 2 (HMGA2) Gene. Cell Cycle 2017, 16, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Vickers, K.C.; Roteta, L.A.; Hucheson-Dilks, H.; Han, L.; Guo, Y. Mining Diverse Small RNA Species in the Deep Transcriptome. Trends Biochem. Sci. 2015, 40, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Han, L.; Vickers, K.C.; Samuels, D.C.; Guo, Y. Alternative Applications for Distinct RNA Sequencing Strategies. Brief. Bioinform. 2015, 16, 629–639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liyanarachchi, S.; Li, W.; Yan, P.; Bundschuh, R.; Brock, P.; Senter, L.; Ringel, M.D.; de la Chapelle, A.; He, H. Genome-Wide Expression Screening Discloses Long Noncoding RNAs Involved in Thyroid Carcinogenesis. J. Clin. Endocrinol. Metab. 2016, 101, 4005–4013. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, H.; Wu, L.; Yao, J.; Meng, X.; Jiang, H.; Xiao, C.; Wu, F. Identification of Specific Long Non-Coding RNA Expression: Profile and Analysis of Association with Clinicopathologic Characteristics and BRAF Mutation in Papillary Thyroid Cancer. Thyroid 2016, 26, 1719–1732. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Liao, T.; Wen, D.; Dong, C.; Zhou, L.; Yang, S.; Wang, Y.; Ji, Q. Corrigendum: Long Intergenic Non-Coding RNA 271 Is Predictive of a Poorer Prognosis of Papillary Thyroid Cancer. Sci. Rep. 2017, 7, 42321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.-H.; Liang, L.; He, R.-Q.; Wen, D.-Y.; Deng, G.-F.; Yang, H.; He, Y.; Ma, W.; Cai, X.-Y.; Chen, J.-Q.; et al. RNA-Sequencing Investigation Identifies an Effective Risk Score Generated by Three Novel lncRNAs for the Survival of Papillary Thyroid Cancer Patients. Oncotarget 2017, 8, 74139–74158. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; He, X.; Hoadley, K.A.; Parker, J.S.; Hayes, D.N.; Perou, C.M. Comparison of RNA-Seq by Poly (A) Capture, Ribosomal RNA Depletion, and DNA Microarray for Expression Profiling. BMC Genom. 2014, 15, 419. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Zhao, S.; Sheng, Q.; Guo, M.; Lehmann, B.; Pietenpol, J.; Samuels, D.C.; Shyr, Y. RNAseq by Total RNA Library Identifies Additional RNAs Compared to Poly(A) RNA Library. BioMed Res. Int. 2015, 2015, 862130. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, T.J.; Dadafarin, S.; Jones, M.; Rodríguez, T.; Gupta, A.; Shin, E.; Moscatello, A.; Iacob, C.; Islam, H.; Tiwari, R.K.; et al. Androgen Activity Is Associated With PD-L1 Downregulation in Thyroid Cancer. Front. Cell Dev. Biol. 2021, 9, 663130. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, X.; Klibanski, A. MEG3 Noncoding RNA: A Tumor Suppressor. J. Mol. Endocrinol. 2012, 48, R45–R53. [Google Scholar] [CrossRef]
- Ying, L.; Huang, Y.; Chen, H.; Wang, Y.; Xia, L.; Chen, Y.; Liu, Y.; Qiu, F. Downregulated MEG3 Activates Autophagy and Increases Cell Proliferation in Bladder Cancer. Mol. Biosyst. 2013, 9, 407–411. [Google Scholar] [CrossRef]
- Harrow, J.; Frankish, A.; Gonzalez, J.M.; Tapanari, E.; Diekhans, M.; Kokocinski, F.; Aken, B.L.; Barrell, D.; Zadissa, A.; Searle, S.; et al. GENCODE: The Reference Human Genome Annotation for The ENCODE Project. Genome Res. 2012, 22, 1760–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benlloch, S.; Payá, A.; Alenda, C.; Bessa, X.; Andreu, M.; Jover, R.; Castells, A.; Llor, X.; Aranda, F.I.; Massutí, B. Detection of BRAF V600E Mutation in Colorectal Cancer: Comparison of Automatic Sequencing and Real-Time Chemistry Methodology. J. Mol. Diagn. 2006, 8, 540–543. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Horvath, S. A General Framework for Weighted Gene Co-Expression Network Analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 1–45. [Google Scholar] [CrossRef]
- Liberzon, A.; Birger, C.; Thorvaldsdóttir, H.; Ghandi, M.; Mesirov, J.P.; Tamayo, P. The Molecular Signatures Database (MSigDB) Hallmark Gene Set Collection. Cell Syst. 2015, 1, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Goldman, M.; Craft, B.; Brooks, A.; Zhu, J.; Haussler, D. The UCSC Xena Platform for Cancer Genomics Data Visualization and Interpretation. BioRxiv 2018, 326470. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas Research Network. Integrated Genomic Characterization of Papillary Thyroid Carcinoma. Cell 2014, 159, 676–690. [Google Scholar] [CrossRef] [Green Version]
- Pratilas, C.A.; Taylor, B.S.; Ye, Q.; Viale, A.; Sander, C.; Solit, D.B.; Rosen, N. V600EBRAF Is Associated with Disabled Feedback Inhibition of RAF–MEK Signaling and Elevated Transcriptional Output of the Pathway. Proc. Natl. Acad. Sci. USA 2009, 106, 4519–4524. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Fu, J.; Zeng, Z.; Cohen, D.; Li, J.; Chen, Q.; Li, B.; Liu, X.S. TIMER2.0 for Analysis of Tumor-Infiltrating Immune Cells. Nucleic Acids Res. 2020, 48, W509–W514. [Google Scholar] [CrossRef]
- Gao, R.; Bai, S.; Henderson, Y.C.; Lin, Y.; Schalck, A.; Yan, Y.; Kumar, T.; Hu, M.; Sei, E.; Davis, A.; et al. Delineating Copy Number and Clonal Substructure in Human Tumors from Single-Cell Transcriptomes. Nat. Biotechnol. 2021, 39, 599–608. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, Y.; Shang, Y.; Mai, J.; Shi, S.; Lu, M.; Bu, C.; Zhang, Z.; Zhang, Z.; Li, Y.; et al. CancerSCEM: A Database of Single-Cell Expression Map across Various Human Cancers. Nucleic Acids Res. 2022, 50, D1147–D1155. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, M.N.; Kimura, E.T.; Gandhi, M.; Biddinger, P.W.; Knauf, J.A.; Basolo, F.; Zhu, Z.; Giannini, R.; Salvatore, G.; Fusco, A.; et al. BRAF Mutations in Thyroid Tumors Are Restricted to Papillary Carcinomas and Anaplastic or Poorly Differentiated Carcinomas Arising from Papillary Carcinomas. J. Clin. Endocrinol. Metab. 2003, 88, 5399–5404. [Google Scholar] [CrossRef] [PubMed]
- Xing, M.; Alzahrani, A.S.; Carson, K.A.; Viola, D.; Elisei, R.; Bendlova, B.; Yip, L.; Mian, C.; Vianello, F.; Tuttle, R.M.; et al. Association between BRAF V600E Mutation and Mortality in Patients with Papillary Thyroid Cancer. JAMA 2013, 309, 1493–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.-K.; Lee, S.; Kim, S.-J.; Jee, H.-G.; Kim, B.-A.; Cho, H.; Song, Y.S.; Cho, S.W.; Won, J.-K.; Shin, J.-Y.; et al. Comprehensive Analysis of the Transcriptional and Mutational Landscape of Follicular and Papillary Thyroid Cancers. PLoS Genet. 2016, 12, e1006239. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.S.; Won, J.-K.; Yoo, S.-K.; Jung, K.C.; Kim, M.J.; Kim, S.-J.; Cho, S.W.; Lee, K.E.; Yi, K.H.; Seo, J.-S.; et al. Comprehensive Transcriptomic and Genomic Profiling of Subtypes of Follicular Variant of Papillary Thyroid Carcinoma. Thyroid 2018, 28, 1468–1478. [Google Scholar] [CrossRef]
- Hamatani, K.; Eguchi, H.; Koyama, K.; Mukai, M.; Nakachi, K.; Kusunoki, Y. A Novel RET Rearrangement (ACBD5/RET) by Pericentric Inversion, Inv (10)(p12. 1; q11. 2), in Papillary Thyroid Cancer from an Atomic Bomb Survivor Exposed to High-Dose Radiation. Oncol. Rep. 2014, 32, 1809–1814. [Google Scholar] [CrossRef] [Green Version]
- Greco, A.; Miranda, C.; Pierotti, M.A. Rearrangements of NTRK1 Gene in Papillary Thyroid Carcinoma. Mol. Cell. Endocrinol. 2010, 321, 44–49. [Google Scholar] [CrossRef]
- Tsai, M.-C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long Noncoding RNA as Modular Scaffold of Histone Modification Complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Nikiforov, Y.E.; Carty, S.E.; Chiosea, S.I.; Coyne, C.; Duvvuri, U.; Ferris, R.L.; Gooding, W.E.; LeBeau, S.O.; Ohori, N.P.; Seethala, R.R.; et al. Impact of the Multi-Gene ThyroSeq Next-Generation Sequencing Assay on Cancer Diagnosis in Thyroid Nodules with Atypia of Undetermined Significance/Follicular Lesion of Undetermined Significance Cytology. Thyroid 2015, 25, 1217–1223. [Google Scholar] [CrossRef]
- Kim, T.H.; Park, Y.J.; Lim, J.A.; Ahn, H.Y.; Lee, E.K.; Lee, Y.J.; Kim, K.W.; Hahn, S.K.; Youn, Y.K.; Kim, K.H.; et al. The Association of the BRAFV600E Mutation with Prognostic Factors and Poor Clinical Outcome in Papillary Thyroid Cancer: A Meta-Analysis. Cancer 2012, 118, 1764–1773. [Google Scholar] [CrossRef]
- Xu, T.-P.; Ma, P.; Wang, W.-Y.; Shuai, Y.; Wang, Y.-F.; Yu, T.; Xia, R.; Shu, Y.-Q. KLF5 and MYC Modulated LINC00346 Contributes to Gastric Cancer Progression through Acting as a Competing Endogeous RNA and Indicates Poor Outcome. Cell Death Differ. 2019, 26, 2179–2193. [Google Scholar] [CrossRef] [PubMed]
- Lessard, L.; Liu, M.; Marzese, D.M.; Wang, H.; Chong, K.; Kawas, N.; Donovan, N.C.; Kiyohara, E.; Hsu, S.; Nelson, N.; et al. The CASC15 Long Intergenic Noncoding RNA Locus Is Involved in Melanoma Progression and Phenotype Switching. J. Investig. Dermatol. 2015, 135, 2464–2474. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Bao, Y.; Wu, Z.; Zhao, T.; Wang, D.; Shi, J.; Liu, B.; Sun, S.; Yang, F.; Wang, L.; et al. Long Noncoding RNA EGFR-AS1 Promotes Cell Growth and Metastasis via Affecting HuR Mediated mRNA Stability of EGFR in Renal Cancer. Cell Death Dis. 2019, 10, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, K.; Jin, S.; Du, Y.; Yu, J.; Feng, H.; Fan, Q.; Ma, W. Long Noncoding RNA DIO3OS Interacts with miR-122 to Promote Proliferation and Invasion of Pancreatic Cancer Cells through Upregulating ALDOA. Cancer Cell Int. 2019, 19, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, G.K.; Mitra, S.; Subhash, S.; Hertwig, F.; Kanduri, M.; Mishra, K.; Fransson, S.; Ganeshram, A.; Mondal, T.; Bandaru, S.; et al. The Risk-Associated Long Noncoding RNA NBAT-1 Controls Neuroblastoma Progression by Regulating Cell Proliferation and Neuronal Differentiation. Cancer Cell 2014, 26, 722–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yan, G.; Zhang, Y.; Jia, X.; Bu, P. Long Non-Coding RNA MEG3 Suppresses Migration and Invasion of Thyroid Carcinoma by Targeting of Rac1. Neoplasma 2015, 62, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Joyce, J.A. Therapeutic Targeting of the Tumor Microenvironment. Cancer Cell 2005, 7, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Keller, E.T.; Garfield, D.H.; Shen, K.; Wang, J. Stromal Cells in Tumor Microenvironment and Breast Cancer. Cancer Metastasis Rev. 2013, 32, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Molinaro, E.; Romei, C.; Biagini, A.; Sabini, E.; Agate, L.; Mazzeo, S.; Materazzi, G.; Sellari-Franceschini, S.; Ribechini, A.; Torregrossa, L.; et al. Anaplastic Thyroid Carcinoma: From Clinicopathology to Genetics and Advanced Therapies. Nat. Rev. Endocrinol. 2017, 13, 644–660. [Google Scholar] [CrossRef]
- Mondal, T.; Subhash, S.; Vaid, R.; Enroth, S.; Uday, S.; Reinius, B.; Mitra, S.; Mohammed, A.; James, A.R.; Hoberg, E.; et al. MEG3 Long Noncoding RNA Regulates the TGF-β Pathway Genes through Formation of RNA-DNA Triplex Structures. Nat. Commun. 2015, 6, 7743. [Google Scholar] [CrossRef] [Green Version]
- Kameyama, K. Expression of MMP-1 in the Capsule of Thyroid Cancer—Relationship with Invasiveness. Pathol. Res. Pract. 1996, 192, 20–26. [Google Scholar] [CrossRef]
- Ren, L.; Xu, Y.; Liu, C.; Wang, S.; Qin, G. IL-17RB Enhances Thyroid Cancer Cell Invasion and Metastasis via ERK1/2 Pathway-Mediated MMP-9 Expression. Mol. Immunol. 2017, 90, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Wang, X.; Yu, X.; Zhang, Y.; Qin, L. MMP16 Promotes Tumor Metastasis and Indicates Poor Prognosis in Hepatocellular Carcinoma. Oncotarget 2017, 8, 72197–72204. [Google Scholar] [CrossRef] [Green Version]
- Hwang, H.S.; Orloff, L.A. Efficacy of Preoperative Neck Ultrasound in the Detection of Cervical Lymph Node Metastasis from Thyroid Cancer. Laryngoscope 2011, 121, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Stulak, J.M.; Grant, C.S.; Farley, D.R.; Thompson, G.B.; van Heerden, J.A.; Hay, I.D.; Reading, C.C.; Charboneau, J.W. Value of Preoperative Ultrasonography in the Surgical Management of Initial and Reoperative Papillary Thyroid Cancer. Arch. Surg. 2006, 141, 489–494, discussion 494–496. [Google Scholar] [CrossRef]
- Prensner, J.R.; Iyer, M.K.; Sahu, A.; Asangani, I.A.; Cao, Q.; Patel, L.; Vergara, I.A.; Davicioni, E.; Erho, N.; Ghadessi, M.; et al. The Long Noncoding RNA SChLAP1 Promotes Aggressive Prostate Cancer and Antagonizes the SWI/SNF Complex. Nat. Genet. 2013, 45, 1392–1398. [Google Scholar] [CrossRef] [Green Version]
- Gutschner, T.; Diederichs, S. The Hallmarks of Cancer: A Long Non-Coding RNA Point of View. RNA Biol. 2012, 9, 703–719. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Guo, W. Long Non-coding RNA MEG3 Suppresses the Growth of Glioma Cells by Regulating the miR-96-5p/MTSS1 Signaling Pathway. Mol. Med. Rep. 2019, 20, 4215–4225. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Chen, J.Y.; Zhong, Y.; Xie, L.; Li, J.S. lncRNA MEG3 Inhibits the Growth of Hepatocellular Carcinoma Cells by Sponging miR-9-5p to Upregulate SOX11. Braz. J. Med. Biol. Res. 2019, 52, e8631. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Diao, H.; Zhong, W.; Wang, T.; Wen, P.; Wu, C. A Cancer Cell Cluster Marked by LincRNA MEG3 Leads Pancreatic Ductal Adenocarcinoma Metastasis. Front. Oncol. 2021, 11, 656564. [Google Scholar] [CrossRef]
- Miyazono, K. Positive and Negative Regulation of TGF-Beta Signaling. J. Cell Sci. 2000, 113 Pt 7, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Terashima, M.; Tange, S.; Ishimura, A.; Suzuki, T. MEG3 Long Noncoding RNA Contributes to the Epigenetic Regulation of Epithelial-Mesenchymal Transition in Lung Cancer Cell Lines. J. Biol. Chem. 2017, 292, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Gokey, J.J.; Snowball, J.; Sridharan, A.; Speth, J.P.; Black, K.E.; Hariri, L.P.; Perl, A.-K.T.; Xu, Y.; Whitsett, J.A. MEG3 Is Increased in Idiopathic Pulmonary Fibrosis and Regulates Epithelial Cell Differentiation. JCI Insight 2018, 3, 122490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive Analysis of Normal Adjacent to Tumor Transcriptomes. Nat. Commun. 2017, 8, 1077. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
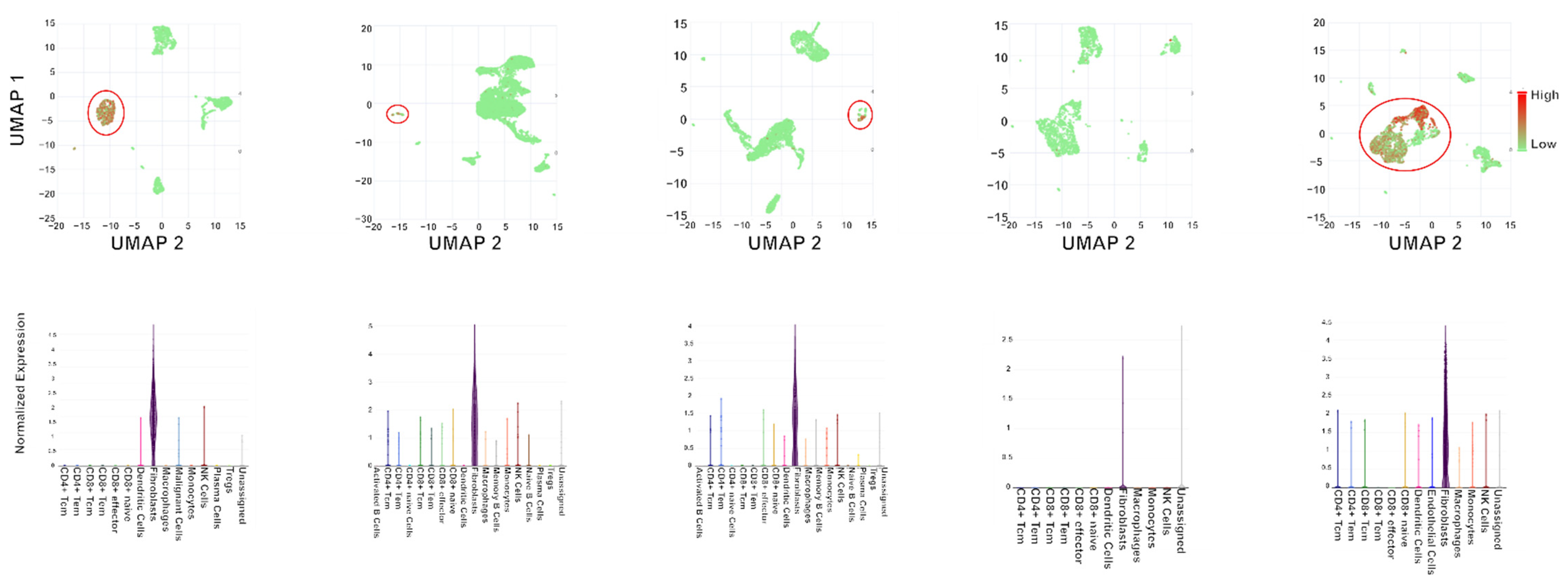{kind=link}
| BRAFV600E | BRAFWT | Total | p-Value | |
|---|---|---|---|---|
| N | 36 (80%) | 9 (20%) | 44 | |
| Age | 47.8 (20–76) | 49 (34–60) | 48 (20–76) | 0.41 |
| Female sex | 26 (72%) | 8 (89%) | 34 (76%) | 0.65 |
| Thyroiditis | 17 (47%) | 5 (56%) | 22 (49%) | 0.72 |
| Tumor size | 1.9 cm (0.45–4.3 cm) | 1.7 cm (0.9–3.2 cm) | 2.8 cm (0.45–4.3 cm) | 0.54 |
| Lymph node metastasis | 15 (42%) | 4 (44%) | 19 (42%) | 1 |
| Invasion | 27 (75%) | 7 (77%) | 34 (76%) | 1 |
| T stage | 1 | |||
| T1–T2 | 9 (25%) | 2 (22%) | 11 (24%) | |
| T3–T4 | 27 (75%) | 7 (77%) | 34 (76%) |
| Gene Name | GS.LN | p*.GS.LN | MM.black | p.MM.black |
|---|---|---|---|---|
| LINC00346 | 0.395204 | 0.009589 | 0.623782 | 1.02 × 10−05 |
| RP1-79C4.4 | 0.383766 | 0.01211 | 0.760822 | 4.99 × 10−09 |
| RP5-1071N3.1 | 0.381124 | 0.012767 | 0.306905 | 0.048048 |
| AC159540.2 | 0.31683 | 0.040921 | 0.290224 | 0.062257 |
| MEG3 | 0.316771 | 0.040961 | 0.91168 | 4.85 × 10−17 |
| NBAT1 | 0.308945 | 0.046506 | 0.682119 | 6.54 × 10−07 |
| RP11-124N14.3 | 0.30859 | 0.046772 | 0.502946 | 0.000687 |
| ITPK1-AS1 | 0.29417 | 0.058626 | 0.362727 | 0.018234 |
| CASC15 | 0.287496 | 0.06487 | 0.68056 | 7.09 × 10−07 |
| EGFR-AS1 | 0.267027 | 0.087358 | 0.213671 | 0.174238 |
| DIO3OS | 0.262486 | 0.093083 | 0.700355 | 2.43 × 10−07 |
| DNM3OS | 0.26201 | 0.093699 | 0.588983 | 4.07 × 10−05 |
| LINC00702 | 0.257338 | 0.099919 | 0.66197 | 1.81 × 10−06 |
| TEX26-AS1 | 0.254926 | 0.10325 | 0.690438 | 4.20 × 10−07 |
| RP11-273B20.1 | 0.25004 | 0.11026 | 0.585077 | 4.71 × 10−05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dadafarin, S.; Rodríguez, T.C.; Carnazza, M.A.; Tiwari, R.K.; Moscatello, A.; Geliebter, J. MEG3 Expression Indicates Lymph Node Metastasis and Presence of Cancer-Associated Fibroblasts in Papillary Thyroid Cancer. Cells 2022, 11, 3181. https://doi.org/10.3390/cells11193181
Dadafarin S, Rodríguez TC, Carnazza MA, Tiwari RK, Moscatello A, Geliebter J. MEG3 Expression Indicates Lymph Node Metastasis and Presence of Cancer-Associated Fibroblasts in Papillary Thyroid Cancer. Cells. 2022; 11(19):3181. https://doi.org/10.3390/cells11193181
Chicago/Turabian StyleDadafarin, Sina, Tomás C. Rodríguez, Michelle A. Carnazza, Raj K. Tiwari, Augustine Moscatello, and Jan Geliebter. 2022. "MEG3 Expression Indicates Lymph Node Metastasis and Presence of Cancer-Associated Fibroblasts in Papillary Thyroid Cancer" Cells 11, no. 19: 3181. https://doi.org/10.3390/cells11193181
APA StyleDadafarin, S., Rodríguez, T. C., Carnazza, M. A., Tiwari, R. K., Moscatello, A., & Geliebter, J. (2022). MEG3 Expression Indicates Lymph Node Metastasis and Presence of Cancer-Associated Fibroblasts in Papillary Thyroid Cancer. Cells, 11(19), 3181. https://doi.org/10.3390/cells11193181