
Naturally Occurring Bicoumarin Compound Daphnoretin Inhibits Growth and Induces Megakaryocytic Differentiation in Human Chronic Myeloid Leukemia Cells

 , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Preparation of Daphnoretin
2.3. Cell lines and Culture
2.4. Cell Growth and Cell Viability Measurement
2.5. Morphological Examination
2.6. Surface Marker Expression Assessed by Immunofluorescent Analysis
2.7. Surface Marker Expression Assessed by Flow Cytometry Analysis
2.8. Cell Cycle Analysis
2.9. RNA Preparation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
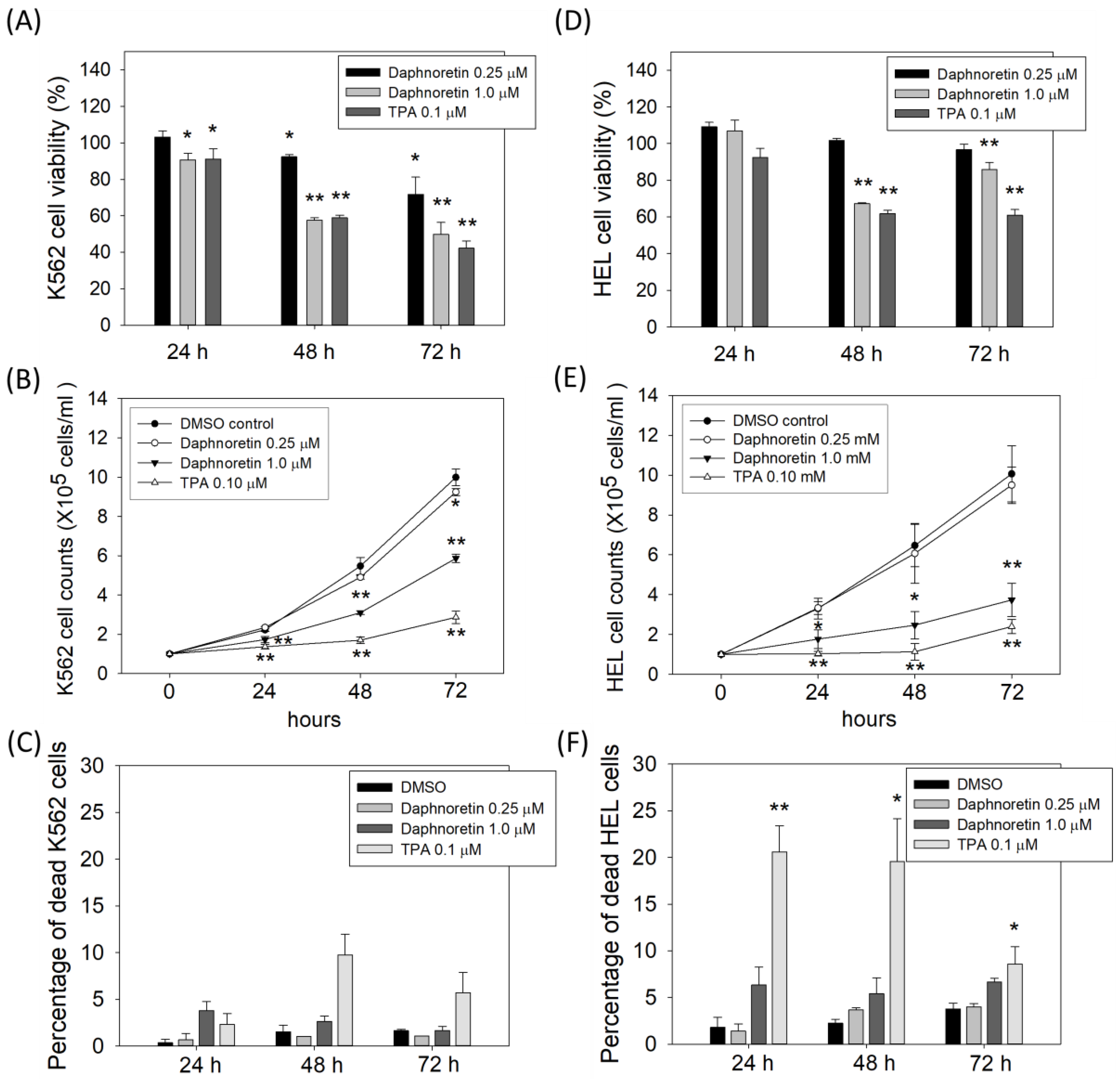
3.1. Daphnoretin Inhibited the Cell Growth
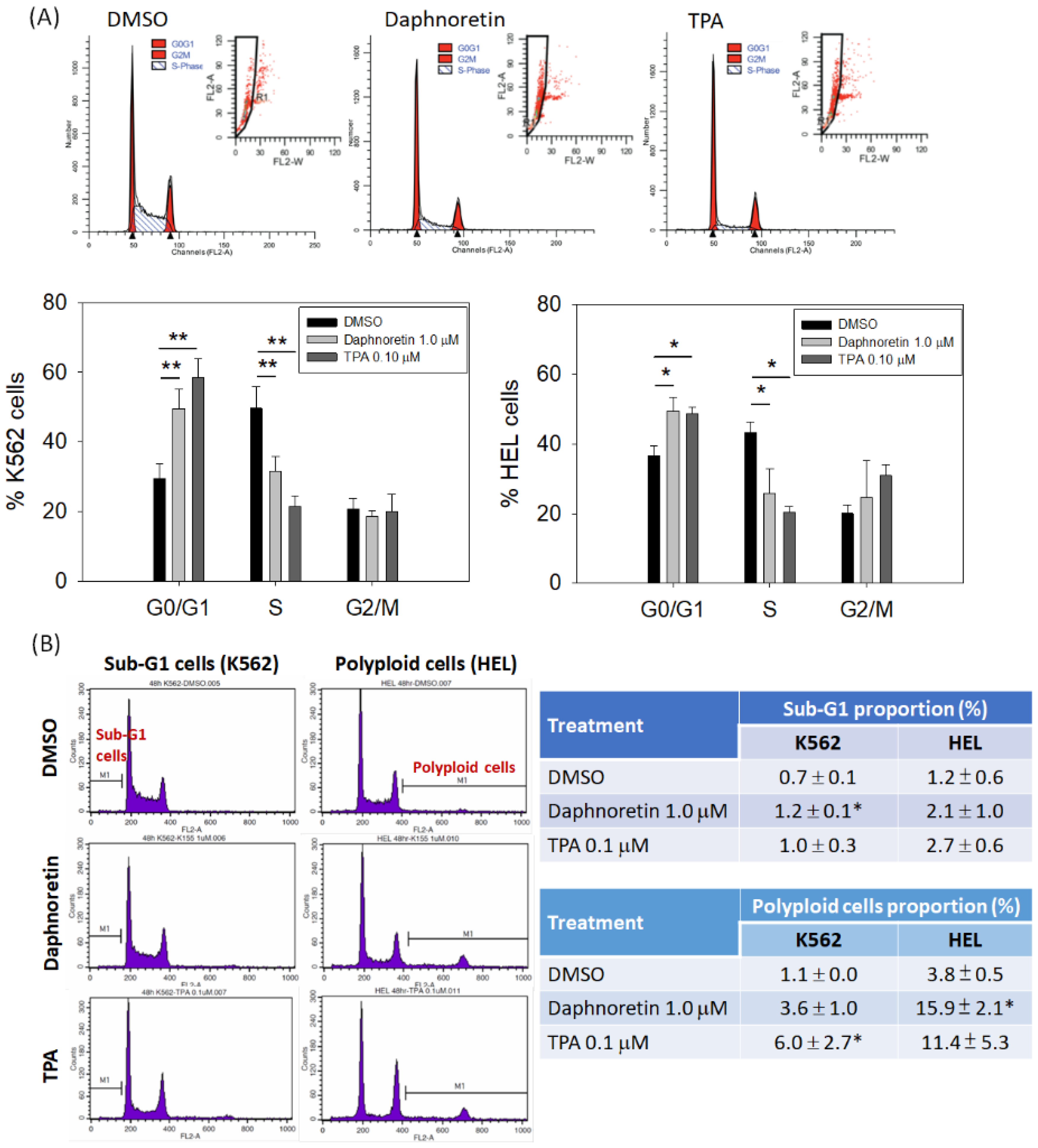
3.2. Effect of Daphnoretin on the Cell Cycle Distribution of K562 and HEL Cells
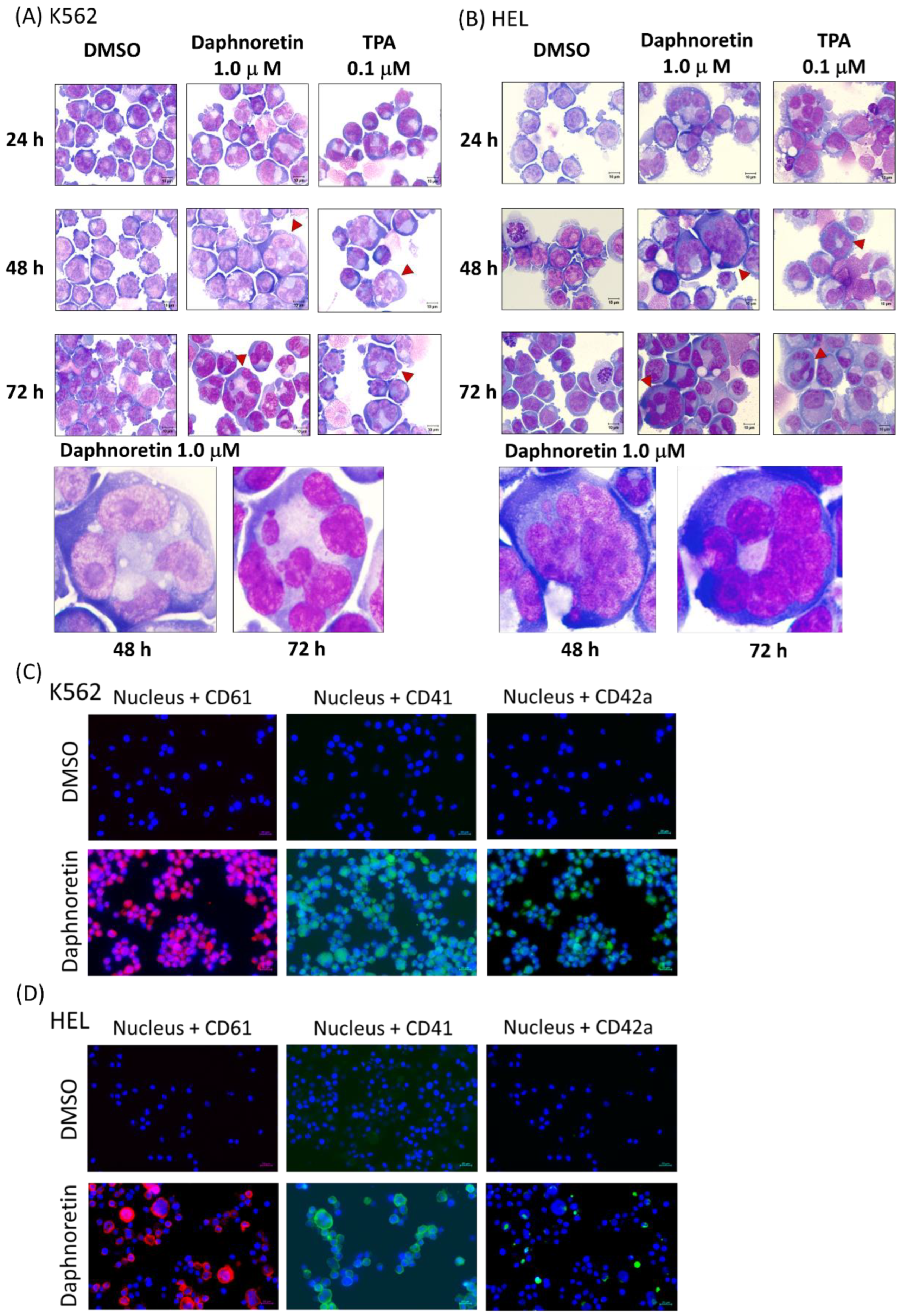
3.3. Daphnoretin-Induced Morphological Changes and Megakaryocyte-Specific Marker Expression
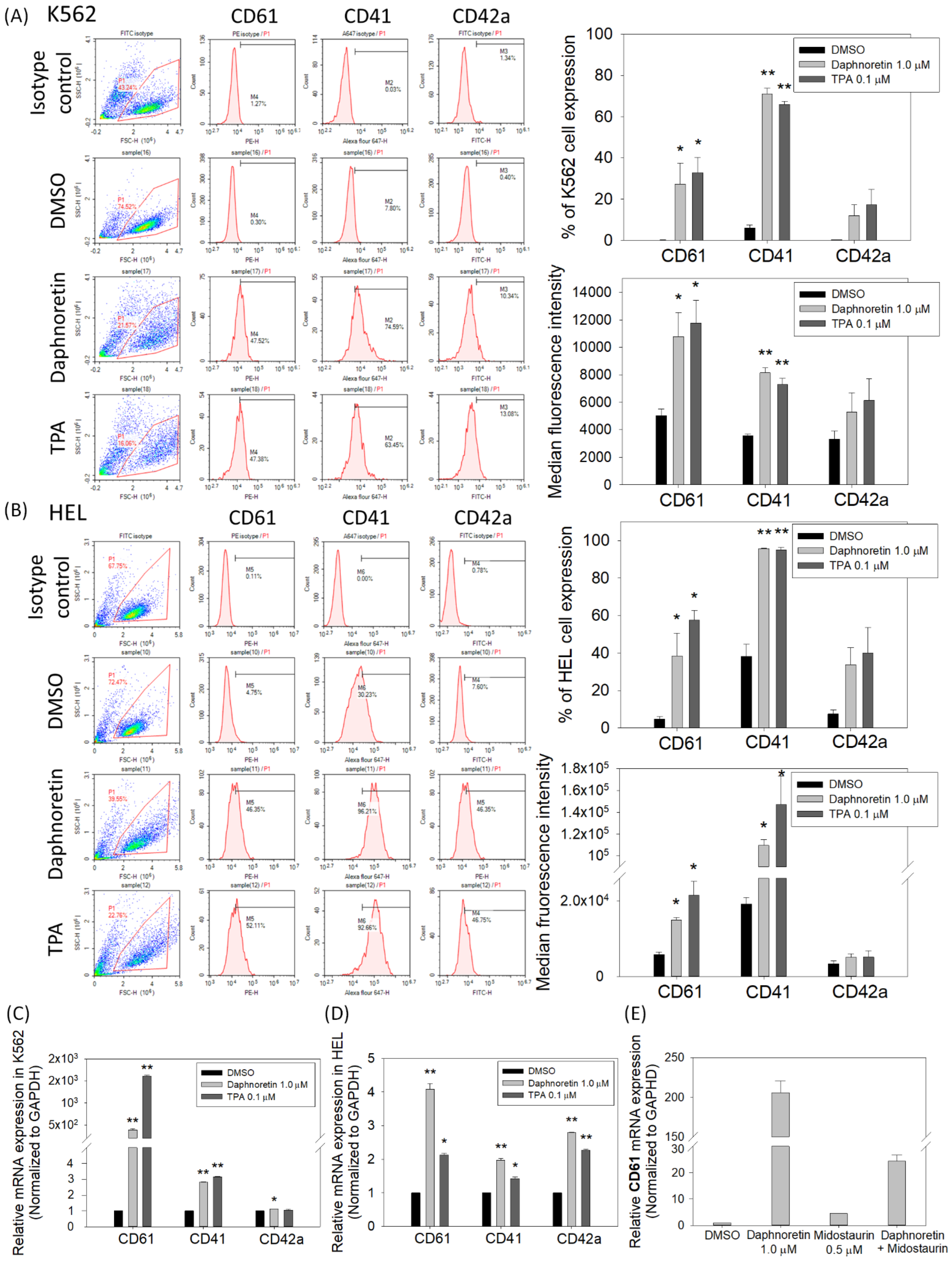
3.4. Daphnoretin Increases the Expression of Megakaryocyte-Specific Markers
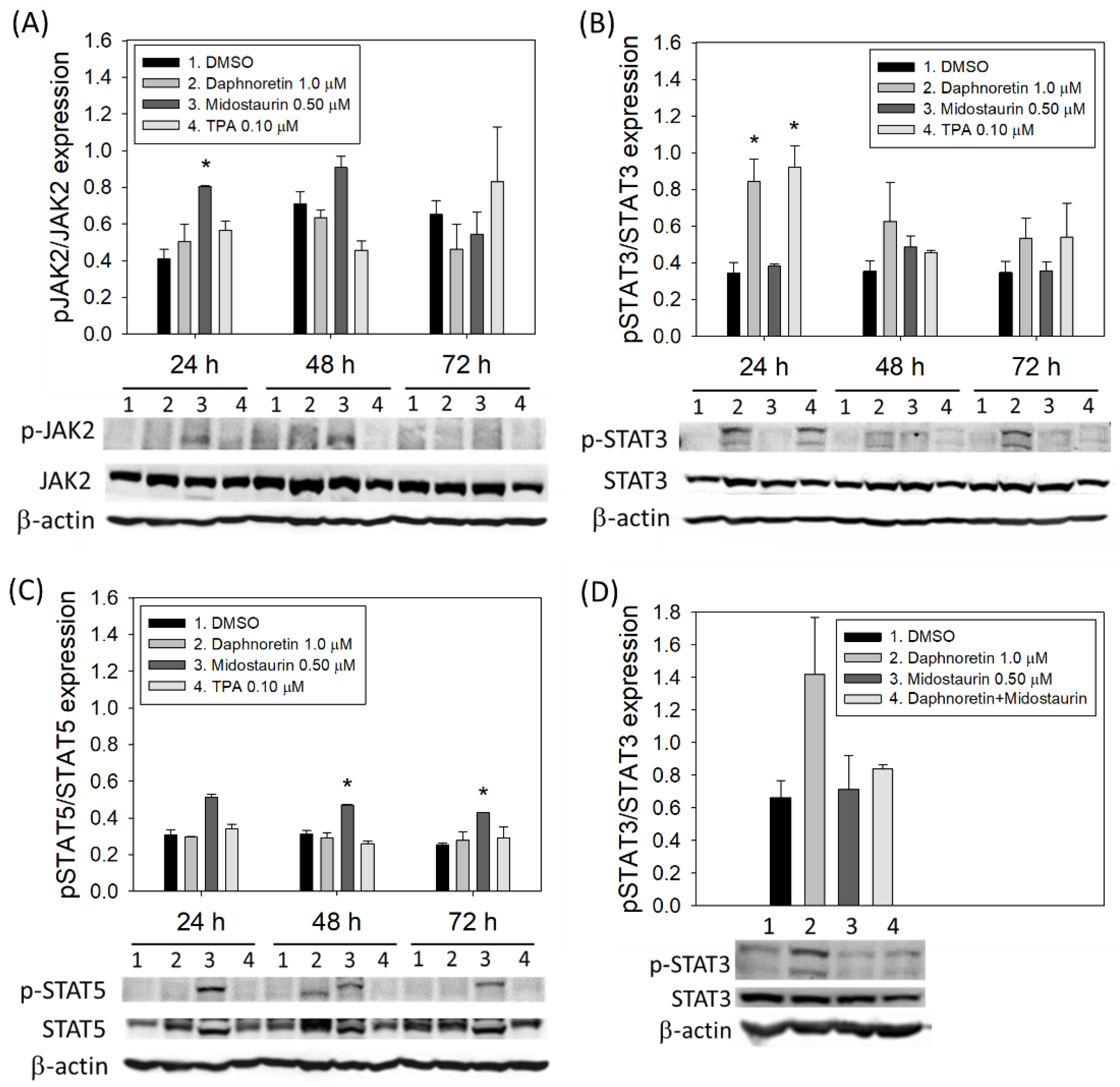
3.5. JAK2 and STAT3/STAT5 Expression in Daphnoretin-Treated Cells
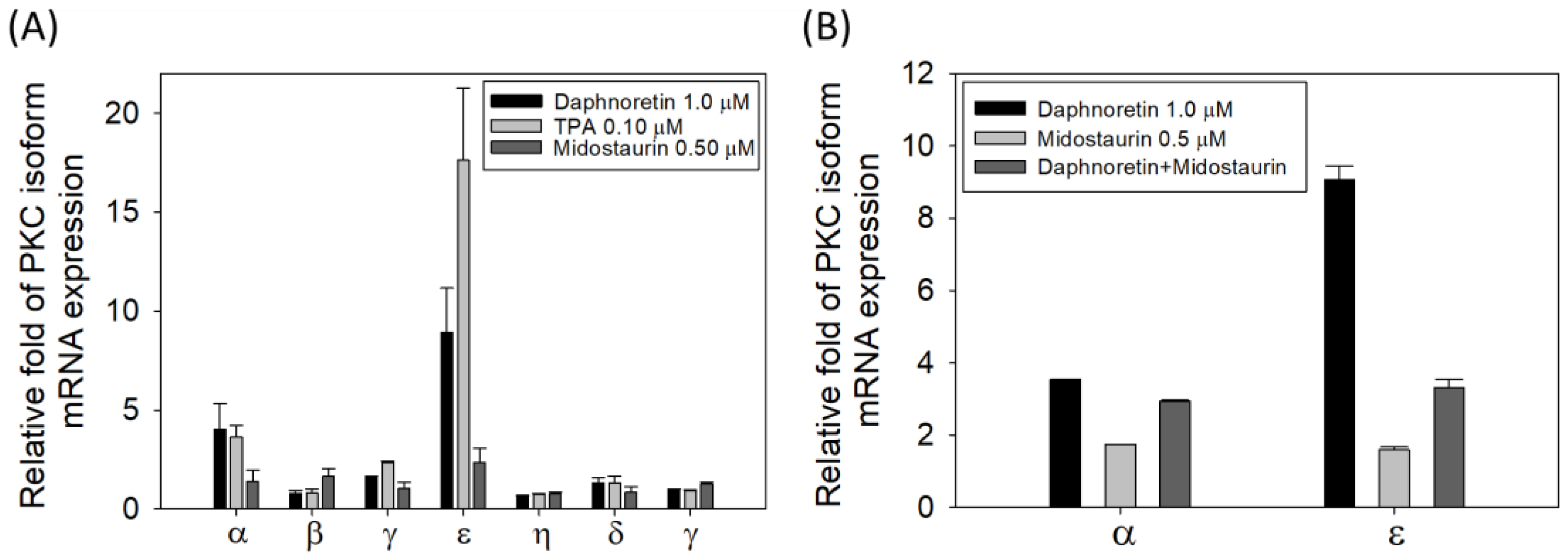
3.6. Relative mRNA Expression of the Different PKC Isozymes under Different Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tefferi, A.; Vainchenker, W. Myeloproliferative neoplasms: Molecular pathophysiology, essential clinical understanding, and treatment strategies. J. Clin. Oncol. 2011, 29, 573–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fava, C.; Kantarjian, H.M.; Jabbour, E.; O’Brien, S.; Jain, N.; Rios, M.B.; Garcia-Manero, G.; Ravandi, F.; Verstovsek, S.; Borthakur, G.; et al. Failure to achieve a complete hematologic response at the time of a major cytogenetic response with second-generation tyrosine kinase inhibitors is associated with a poor prognosis among patients with chronic myeloid leukemia in accelerated or blast phase. Blood 2009, 113, 5058–5063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifacio, M.; Stagno, F.; Scaffidi, L.; Krampera, M.; Di Raimondo, F. Management of Chronic Myeloid Leukemia in Advanced Phase. Front. Oncol. 2019, 9, 1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Thé, H.; Chen, Z. Acute promyelocytic leukaemia: Novel insights into the mechanisms of cure. Nat. Rev. Cancer 2010, 10, 775–783. [Google Scholar] [CrossRef] [PubMed]
- De Thé, H. Differentiation therapy revisited. Nat. Rev. Cancer 2018, 18, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Yen, J.H.; Lin, C.Y.; Chuang, C.H.; Chin, H.K.; Wu, M.J.; Chen, P.Y. Nobiletin Promotes Megakaryocytic Differentiation through the MAPK/ERK-Dependent EGR1 Expression and Exerts Anti-Leukemic Effects in Human Chronic Myeloid Leukemia (CML) K562 Cells. Cells 2020, 9, 877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reina-Campos, M.; Diaz-Meco, M.T.; Moscat, J. The Dual Roles of the Atypical Protein Kinase Cs in Cancer. Cancer Cell 2019, 36, 218–235. [Google Scholar] [CrossRef]
- Klein, E.; Ben-Bassat, H.; Neumann, H.; Ralph, P.; Zeuthen, J.; Polliack, A.; Vánky, F. Properties of the K562 cell line, derived from a patient with chronic myeloid leukemia. Int. J. Cancer 1976, 18, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Papayannopoulou, T. HEL cells: A new human erythroleukemia cell line with spontaneous and induced globin expression. Science 1982, 216, 1233–1235. [Google Scholar] [CrossRef]
- Papayannopoulou, T.; Nakamoto, B.; Yokochi, T.; Chait, A.; Kannagi, R. Human erythroleukemia cell line (HEL) undergoes a drastic macrophage-like shift with TPA. Blood 1983, 62, 832–845. [Google Scholar] [CrossRef] [PubMed]
- Alitalo, R. Induced differentiation of K562 leukemia cells: A model for studies of gene expression in early megakaryoblasts. Leuk. Res. 1990, 14, 501–514. [Google Scholar] [CrossRef]
- Long, M.W.; Heffner, C.H.; Williams, J.L.; Peters, C.; Prochownik, E.V. Regulation of megakaryocyte phenotype in human erythroleukemia cells. J. Clin. Investig. 1990, 85, 1072–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutsch, V.R.; Tomer, A. Megakaryocyte development and platelet production. Br. J. Haematol. 2006, 134, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Séverin, S.; Ghevaert, C.; Mazharian, A. The mitogen-activated protein kinase signaling pathways: Role in megakaryocyte differentiation. J. Thromb. Haemost. 2010, 8, 17–26. [Google Scholar] [CrossRef]
- Kaushansky, K. Thrombopoietin and the hematopoietic stem cell. Ann. N. Y. Acad. Sci. 2005, 1044, 139–141. [Google Scholar] [CrossRef] [PubMed]
- Kaushansky, K. Thrombopoietin: The primary regulator of megakaryocyte and platelet production. Thromb. Haemost. 1995, 74, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Hashimoto, Y.; Kidokoro, M. (+)-Nortrachelogenin, a new pharmacologically active lignan from Wikstroemia indica. J. Nat. Prod. 1979, 42, 159–162. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.M.; Zhu, L.; Jiang, J.G.; Yang, L.; Wang, D.Y. Bioactive components and pharmacological action of Wikstroemia indica (L.) C. A. Mey and its clinical application. Curr. Pharm. Biotechnol. 2009, 10, 743–752. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.H.; Tagahara, K.; Suzuki, H.; Wu, R.Y.; Haruna, M.; Hall, I.H.; Huang, H.C.; Ito, K.; Iida, T.; Lai, J.S. Antitumor agents. 49 tricin, kaempferol-3-O-beta-D-glucopyranoside and (+)-nortrachelogenin, antileukemic principles from Wikstroemia indica. J. Nat. Prod. 1981, 44, 530–535. [Google Scholar] [CrossRef]
- Chen, H.C.; Chou, C.K.; Kuo, Y.H.; Yeh, S.F. Identification of a protein kinase C (PKC) activator, daphnoretin, that suppresses hepatitis B virus gene expression in human hepatoma cells. Biochem. Pharmacol. 1996, 52, 1025–1032. [Google Scholar] [CrossRef]
- Ko, F.N.; Chang, Y.L.; Kuo, Y.H.; Lin, Y.L.; Teng, C.M. Daphnoretin, a new protein kinase C activator isolated from Wikstroemia indica C.A. Mey. Biochem. J. 1993, 295 Pt 1, 321–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.A.; Liu, C.K.; Hsu, M.L.; Chi, C.W.; Ko, C.C.; Chen, J.S.; Lai, C.T.; Chang, H.H.; Lee, T.Y.; Lai, Y.L.; et al. Daphnoretin modulates differentiation and maturation of human dendritic cells through down-regulation of c-Jun N-terminal kinase. Int. Immunopharmacol. 2017, 51, 25–30. [Google Scholar] [CrossRef]
- Hall, I.H.; Tagahara, K.; Lee, K.H. Antitumor agents LIII: The effects of daphnoretin on nucleic acid and protein synthesis of Ehrlich ascites tumor cells. J. Pharm. Sci. 1982, 71, 741–744. [Google Scholar] [CrossRef] [PubMed]
- Sarno, F.; Pepe, G.; Termolino, P.; Carafa, V.; Massaro, C.; Merciai, F.; Campiglia, P.; Nebbioso, A.; Altucci, L. Trifolium Repens Blocks Proliferation in Chronic Myelogenous Leukemia via the BCR-ABL/STAT5 Pathway. Cells 2020, 9, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.Y.; Kan, J.T.; Cheng, Z.Y.; Wang, X.L.; Zhu, Y.Z.; Guo, W. Daphnoretin-induced apoptosis in HeLa cells: A possible mitochondria-dependent pathway. Cytotechnology 2014, 66, 51–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.F.; Wu, Z.; Bai, X.; Zhang, Y.; He, P. Effect of daphnoretin on the proliferation and apoptosis of A549 lung cancer cells in vitro. Oncol. Lett. 2014, 8, 1139–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Guo, H.; Gao, X.; Li, M.; Bian, H. Daphnoretin: An invasion inhibitor and apoptosis accelerator for colon cancer cells by regulating the Akt signal pathway. Biomed. Pharmacother. 2019, 111, 1013–1021. [Google Scholar] [CrossRef]
- Huang, Y.C.; Chao, D.K.; Clifford Chao, K.S.; Chen, Y.J. Oral small-molecule tyrosine kinase inhibitor midostaurin (PKC412) inhibits growth and induces megakaryocytic differentiation in human leukemia cells. Toxicol. Vitr. 2009, 23, 979–985. [Google Scholar] [CrossRef]
- Huang, T.Y.; Peng, S.F.; Huang, Y.P.; Tsai, C.H.; Tsai, F.J.; Huang, C.Y.; Tang, C.H.; Yang, J.S.; Hsu, Y.M.; Yin, M.C.; et al. Combinational treatment of all-trans retinoic acid (ATRA) and bisdemethoxycurcumin (BDMC)-induced apoptosis in liver cancer Hep3B cells. J. Food Biochem. 2020, 44, e13122. [Google Scholar] [CrossRef]
- Wu, M.H.; Lee, T.H.; Lee, H.P.; Li, T.M.; Lee, I.T.; Shieh, P.C.; Tang, C.H. Kuei-Lu-Er-Xian-Jiao extract enhances BMP-2 production in osteoblasts. Biomedicine 2017, 7, 2. [Google Scholar] [CrossRef]
- Huang, Y.C.; Liao, W.L.; Lin, J.M.; Chen, C.C.; Liu, S.P.; Chen, S.Y.; Lin, Y.N.; Lei, Y.J.; Liu, H.T.; Chen, Y.J.; et al. High levels of circulating endothelial progenitor cells in patients with diabetic retinopathy are positively associated with ARHGAP22 expression. Oncotarget 2018, 9, 17858–17866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.L.; Lin, J.M.; Liu, S.P.; Chen, S.Y.; Lin, H.J.; Wang, Y.H.; Lei, Y.J.; Huang, Y.C.; Tsai, F.J. Loss of Response Gene to Complement 32 (RGC-32) in Diabetic Mouse Retina Is Involved in Retinopathy Development. Int. J. Mol. Sci. 2018, 19, 3629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, Y.; Martin, J.F.; Vainchenker, W.; Erusalimsky, J.D. Inhibition of protein kinase C suppresses megakaryocytic differentiation and stimulates erythroid differentiation in HEL cells. Blood 1996, 87, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirito, K.; Osawa, M.; Morita, H.; Shimizu, R.; Yamamoto, M.; Oda, A.; Fujita, H.; Tanaka, M.; Nakajima, K.; Miura, Y.; et al. A functional role of Stat3 in in vivo megakaryopoiesis. Blood 2002, 99, 3220–3227. [Google Scholar] [CrossRef] [Green Version]
- Miyakawa, Y.; Oda, A.; Druker, B.J.; Miyazaki, H.; Handa, M.; Ohashi, H.; Ikeda, Y. Thrombopoietin induces tyrosine phosphorylation of Stat3 and Stat5 in human blood platelets. Blood 1996, 87, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Columbyova, L.; Loda, M.; Scadden, D.T. Thrombopoietin receptor expression in human cancer cell lines and primary tissues. Cancer Res. 1995, 55, 3509–3512. [Google Scholar]
- Wang, H.; Hu, H.; Zhang, Q.; Yang, Y.; Li, Y.; Hu, Y.; Ruan, X.; Yang, Y.; Zhang, Z.; Shu, C.; et al. Dynamic transcriptomes of human myeloid leukemia cells. Genomics 2013, 102, 250–256. [Google Scholar] [CrossRef]
- Zhang, X.; Ren, R. Bcr-Abl efficiently induces a myeloproliferative disease and production of excess interleukin-3 and granulocyte-macrophage colony-stimulating factor in mice: A novel model for chronic myelogenous leukemia. Blood 1998, 92, 3829–3840. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers |
|---|---|
| CD41 | 5′ -TGCTGCTCACCATCCTGGTC 5′ -AACCCAAAGCTTGGAGGCAAC |
| CD42a | 5′ -TCTGTATCAGAAGCCCTGTCTTCAC 5′ -GCATCGGGAGCTTTGTCTTG |
| CD61 | 5′ -TGTATGGGACTCAAGATTGGA 5‘ -AGGGATGGCTATTAGGTTCA |
| PKC α | 5′ -CAAGGTTCATGCAGCCCAAC 5′ -ACTGTGTCCCTGGCAAAACA |
| PKC β | 5′ -GACCAAACACCCAGGCAAAC 5′ -GATGGCGGGTGAAAAATCGG |
| PKC γ | 5′ -GAGATCCCGCCTCCTTTCAG 5′ -CTGGGGTGCAGGATATGACG |
| PKC ε | 5′ -CGGCGAGGAAATACATGCAC 5′ -GGGCAGGAATGAAGAACCGA |
| PKC η | 5′ -GGTGCTGAAGAAGGACGTGA 5′ -AAAACAGACGATCGGGGGTC |
| PKC δ | 5′ -TGGTTGGTGCGTTGTAGCAG 5′ -TAGGAGTTGAAGGCGATGCG |
| PKC θ | 5′ -AAACCTCAAGGCCGAATGC 5′ -AGAAGGTGGCAGTGAACTCG |
| GAPDH | 5′ -ATGAGAAGTATGACAACAGCCT 5′ -AGTCCTTCCACGATACCAAAGT |
| Multiple Nuclei Cell (%) | ||||
|---|---|---|---|---|
| DMSO | Daphnoretin (1.0 μM) | TPA (0.1 μM) | ||
| 24 h | K562 | 1.5 ± 1.5 | 44.5 ± 3.3 * | 51.4 ± 4.8 * |
| HEL | 3.8 ± 0.3 | 25.8 ± 5.3 * | 22.5 ± 6.7 * | |
| 48 h | K562 | 7.0 ± 1.5 | 44.7 ± 10.9 * | 54.4 ± 13.8 * |
| HEL | 3.1 ± 1.4 | 34.1 ± 4.3 ** | 39.4 ± 4.2 ** | |
| 72 h | K562 | 6.2 ± 0.3 | 43.3 ± 2.1 * | 67.4 ± 4.4 * |
| HEL | 6.0 ± 1.2 | 34.8 ± 5.0 ** | 39.6 ± 5.8 ** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.-C.; Huang, C.-P.; Lin, C.-P.; Yang, K.-C.; Lei, Y.-J.; Wang, H.-P.; Kuo, Y.-H.; Chen, Y.-J. Naturally Occurring Bicoumarin Compound Daphnoretin Inhibits Growth and Induces Megakaryocytic Differentiation in Human Chronic Myeloid Leukemia Cells. Cells 2022, 11, 3252. https://doi.org/10.3390/cells11203252
Huang Y-C, Huang C-P, Lin C-P, Yang K-C, Lei Y-J, Wang H-P, Kuo Y-H, Chen Y-J. Naturally Occurring Bicoumarin Compound Daphnoretin Inhibits Growth and Induces Megakaryocytic Differentiation in Human Chronic Myeloid Leukemia Cells. Cells. 2022; 11(20):3252. https://doi.org/10.3390/cells11203252
Chicago/Turabian StyleHuang, Yu-Chuen, Chun-Ping Huang, Chin-Ping Lin, Kai-Chien Yang, Yu-Jie Lei, Hao-Pei Wang, Yueh-Hsiung Kuo, and Yu-Jen Chen. 2022. "Naturally Occurring Bicoumarin Compound Daphnoretin Inhibits Growth and Induces Megakaryocytic Differentiation in Human Chronic Myeloid Leukemia Cells" Cells 11, no. 20: 3252. https://doi.org/10.3390/cells11203252
APA StyleHuang, Y. -C., Huang, C. -P., Lin, C. -P., Yang, K. -C., Lei, Y. -J., Wang, H. -P., Kuo, Y. -H., & Chen, Y. -J. (2022). Naturally Occurring Bicoumarin Compound Daphnoretin Inhibits Growth and Induces Megakaryocytic Differentiation in Human Chronic Myeloid Leukemia Cells. Cells, 11(20), 3252. https://doi.org/10.3390/cells11203252