
Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Subjects of the Study
Assessment of Outcome
2.3. Statistical Analysis
3. Results
3.1. Baseline Characteristics
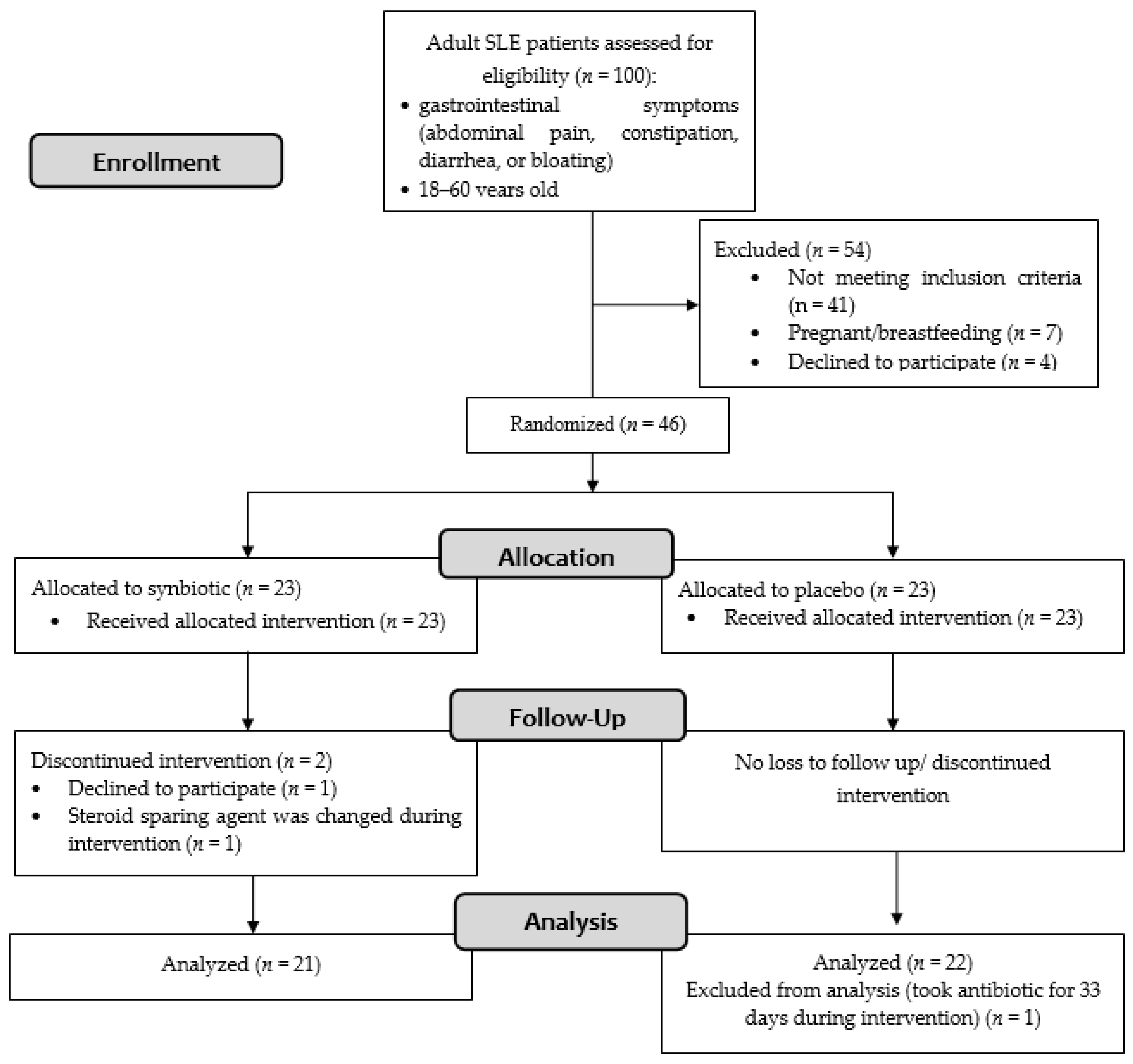
3.2. Follow-up and Adverse Events
3.3. Dietary Assessment
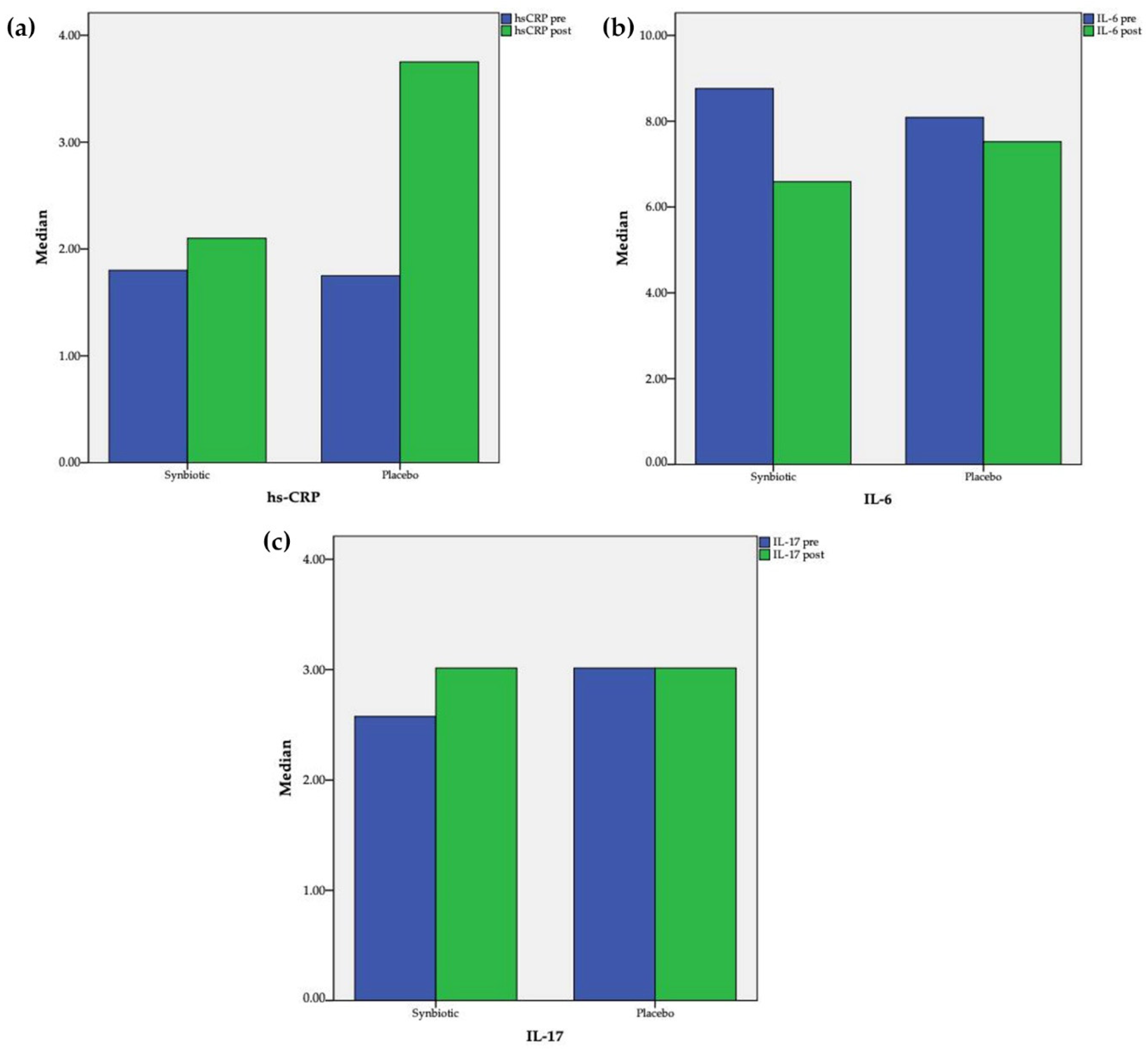
3.4. Changes in Serum Hs-CRP, IL-6, and IL-17
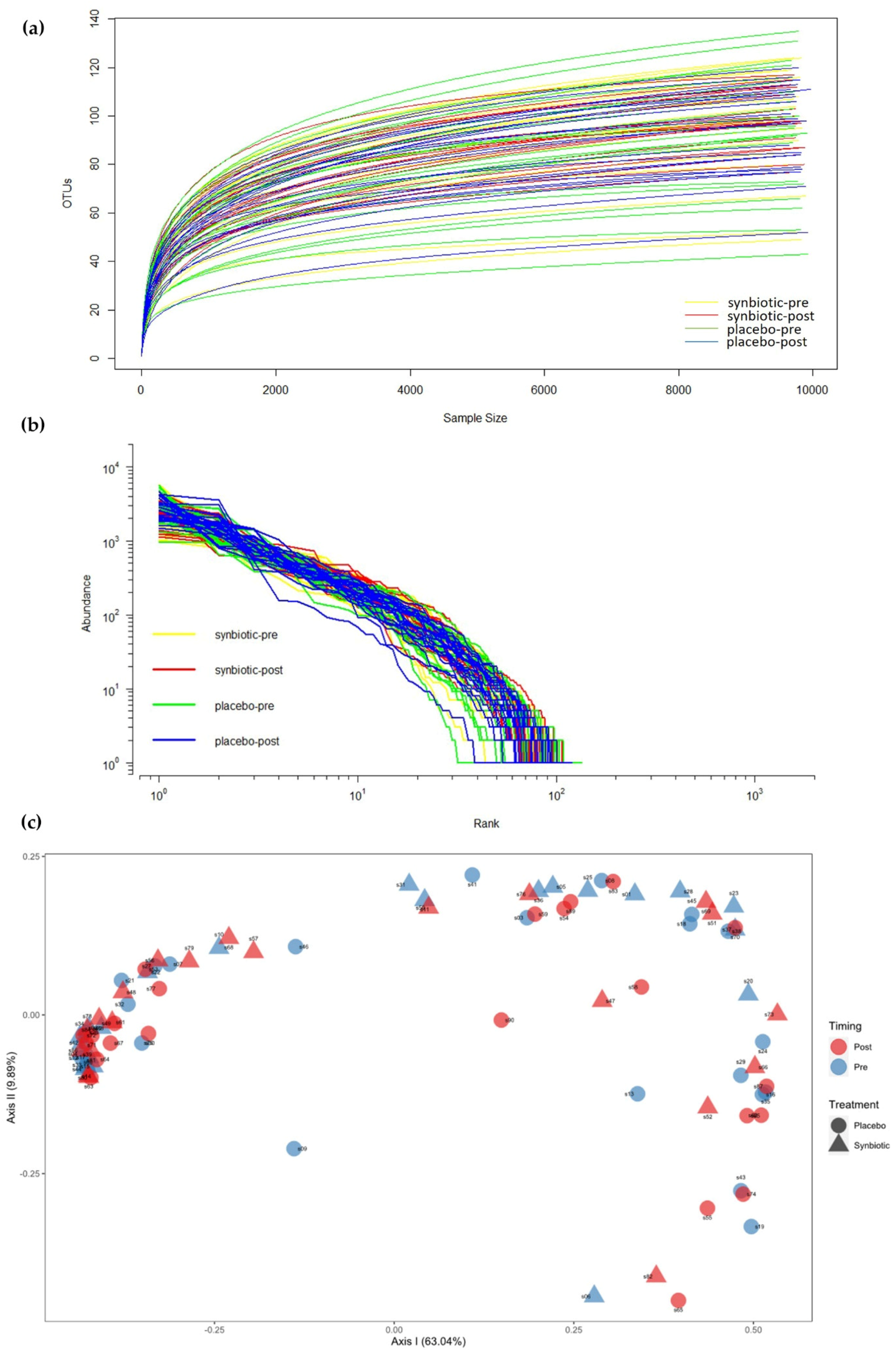
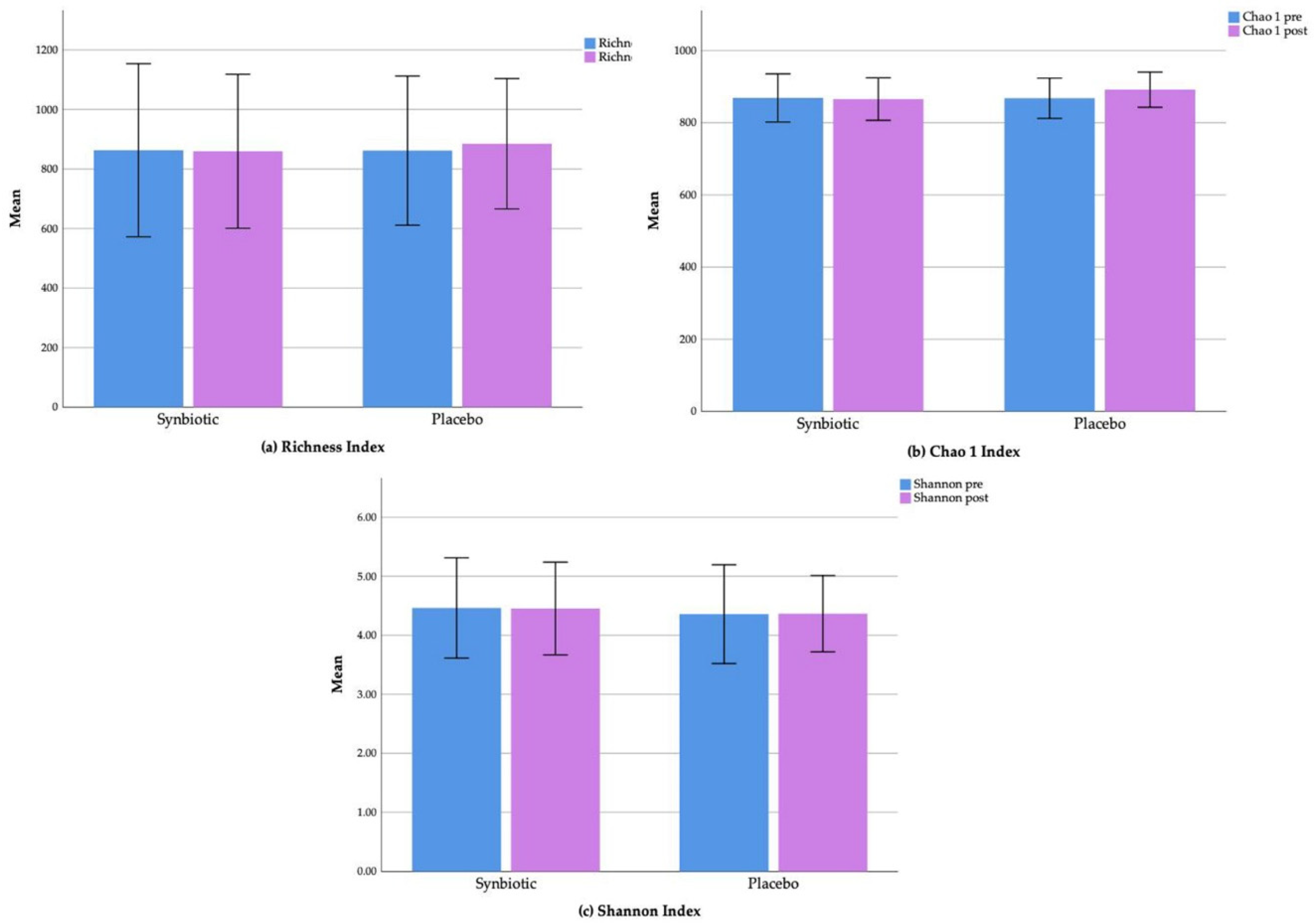
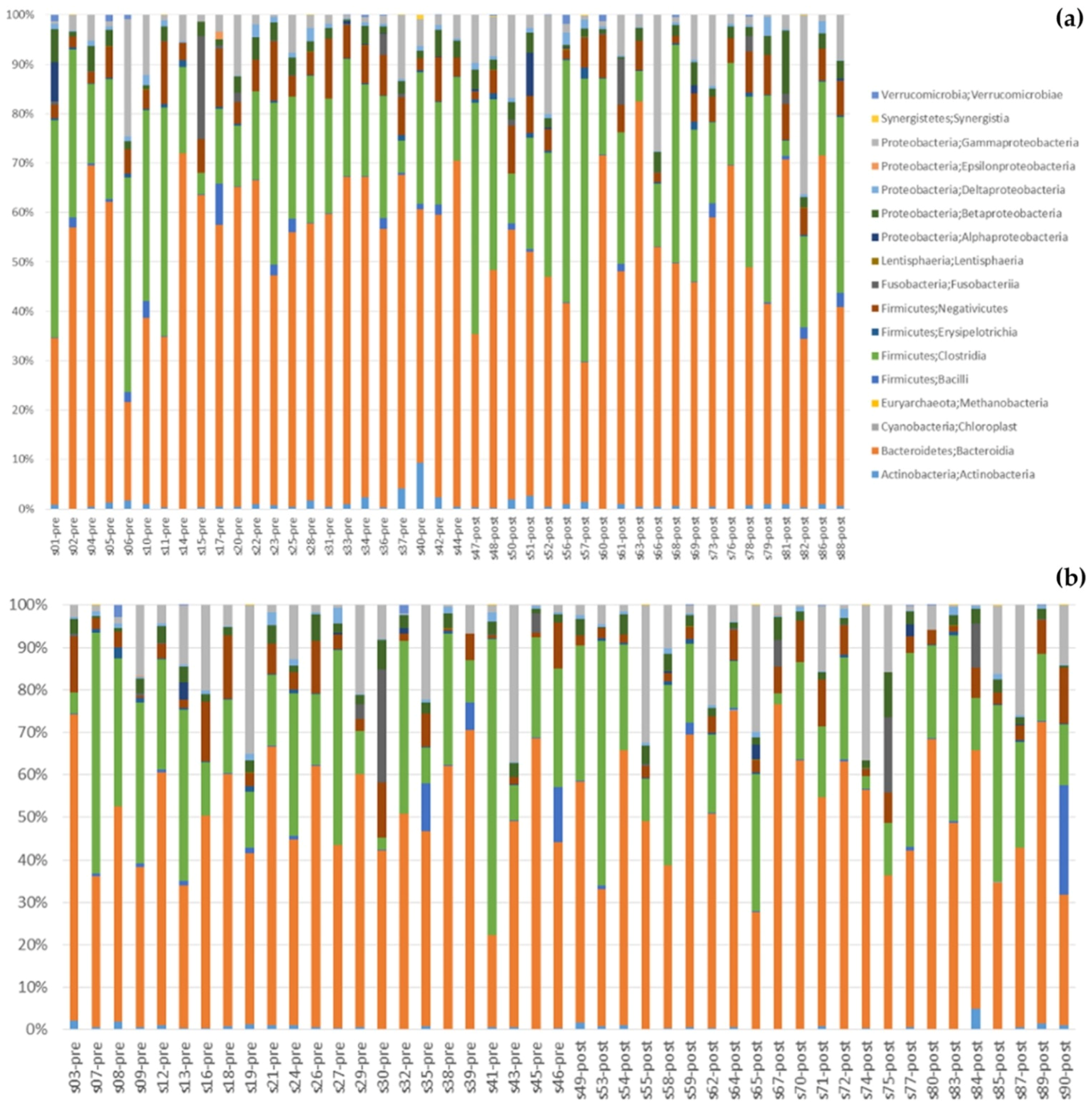
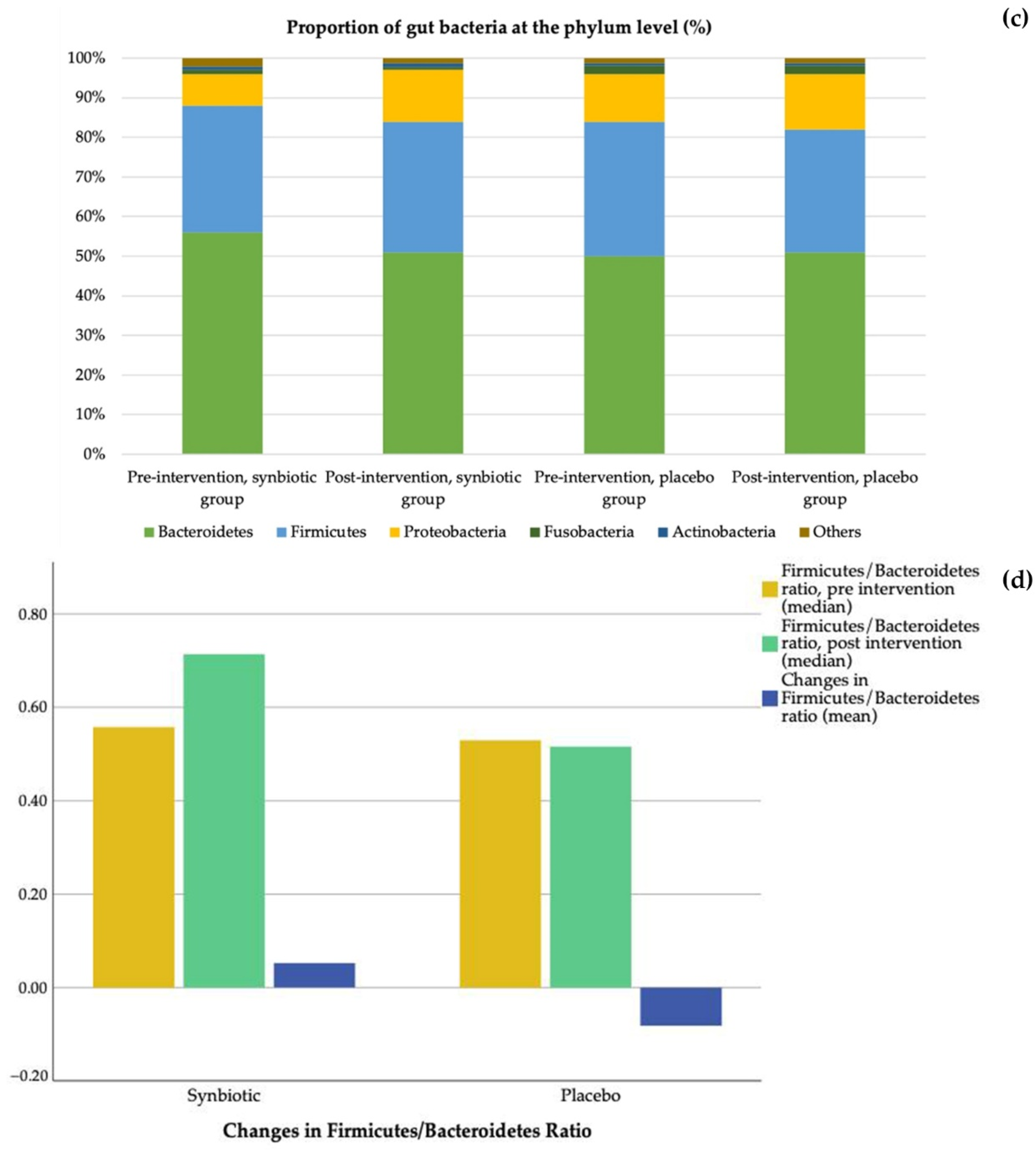
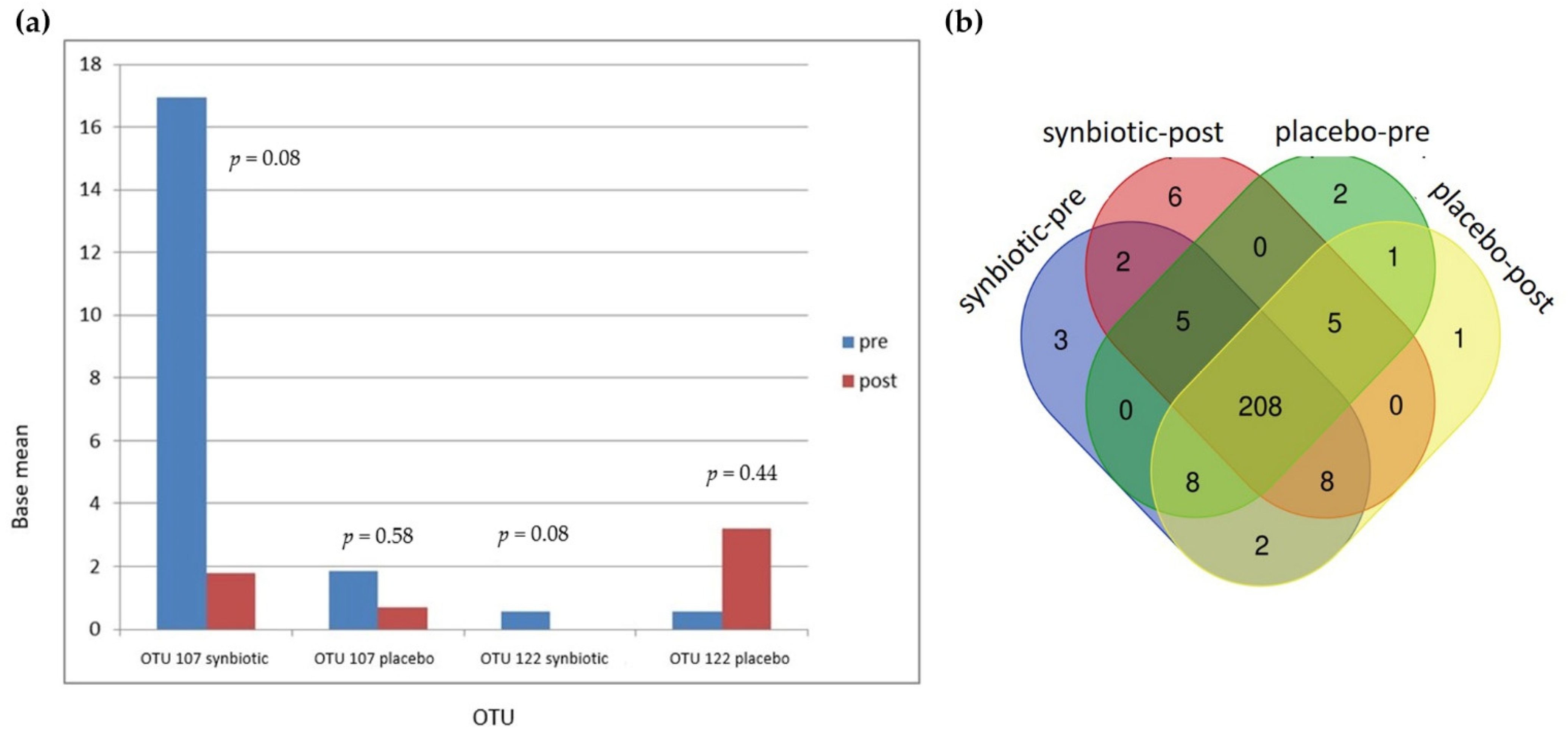
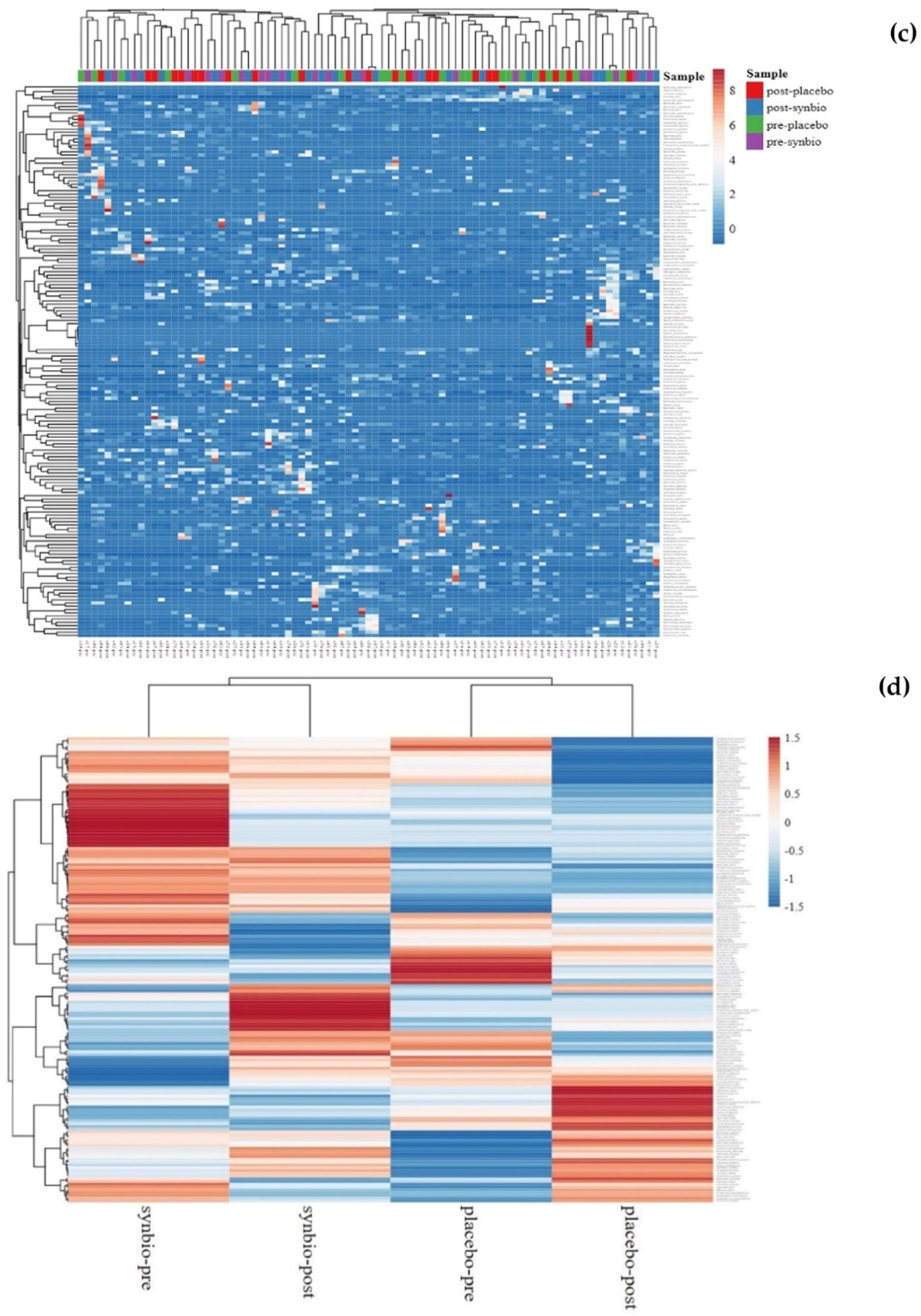
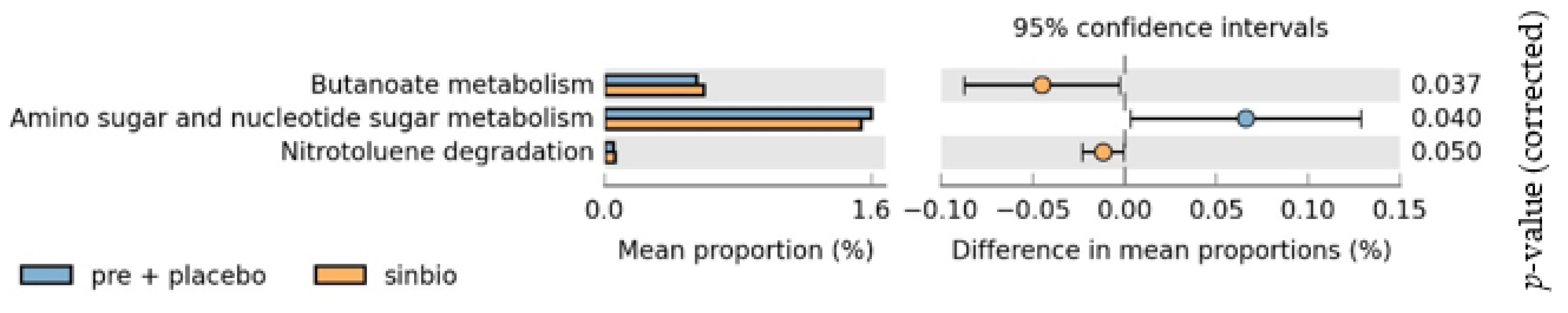
3.5. Gut Microbiota
3.6. SLE Disease Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bertsias, G.; Cervera, R.; Boumpas, D.T. Systemic Lupus Erythematosus: Pathogenesis and Clinical Features. 2012. Available online: https://www.eular.org/myuploaddata/files/sample%20chapter20_mod%2017.pdf (accessed on 1 October 2022).
- Katz-Agranov, N. The microbiome and systemic lupus erythematosus. Immunol. Res. 2017, 65, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fasano, A. Zonulin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, J.; Kanazawa, A.; Azuma, K.; Ikeda, F.; Goto, H.; Komiya, K.; Kanno, R.; Tamura, Y.; Asahara, T.; Takahashi, T.; et al. Probiotic reduces bacterial translocation in type 2 diabetes mellitus: A randomized controlled study. Sci. Rep. 2017, 7, 12115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lin, J.; Xiao, L.; Zhang, X.; Zhao, L.; Wang, M.; Li, L. Gut microbiota in systemic lupus erythematosus: A fuse and a solution. J. Autoimmun. 2022, 132, 102867. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Meng, X.; Chen, B.; Zhao, L.; Zhang, X. Gut microbiota in lupus: A butterfly effect? Curr. Rheumatol. Rep. 2021, 23, 27. [Google Scholar] [CrossRef]
- Zhang, H.; Liao, X.; Sparks, J.B.; Luo, X.M. Dynamics of gut microbiota in autoimmune lupus. Appl. Environ. Microbiol. 2014, 80, 7551–7560. [Google Scholar] [CrossRef] [Green Version]
- Gerges, M.A.; Esmaeel, N.E.; Makram, W.K.; Sharaf, D.M.; Gebriel, M.G. Altered profile of fecal microbiota in newly diagnosed systemic lupus erythematosus Egyptian patients. Int. J. Microbiol. 2021, 2021, 9934533. [Google Scholar] [CrossRef]
- Hevia, A.; Milani, C.; López, P.; Cuervo, A.; Arboleya, S.; Duranti, S.; Turroni, F.; González, S.; Suárez, A.; Gueimonde, M.; et al. Intestinal dysbiosis associated with systemic lupus erythematosus. MBio 2014, 5, e1548. [Google Scholar] [CrossRef] [Green Version]
- Rojo, D.; Hevia, A.; Bargiela, R.; López, P.; Cuervo, A.; González, S.; Suárez, A.; Sánchez, B.; Martinez-Martinez, M.; Milani, C.; et al. Ranking the impact of human health disorders on gut metabolism: Systemic lupus erythematosus and obesity as study cases. Sci. Rep. 2015, 5, 8310. [Google Scholar] [CrossRef] [Green Version]
- Azzouz, D.; Omarbekova, A.; Heguy, A.; Schwudke, D.; Gisch, N.; Rovin, B.H.; Caricchio, R.; Buyon, J.; Alekseyenko, A.; Silverman, G.J. Lupus nephritis is linked to disease-activity associated expansions and immunity to a gut commensal. Ann. Rheum. Dis. 2019, 78, 947–956. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Shao, T.; Li, H.; Xie, Z.; Wen, C. Alterations of the gut microbiome in Chinese patients with systemic lupus erythematosus. Gut Pathog. 2016, 8, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Qing, P.; Yang, H.; Wu, Y.; Liu, Y.; Luo, Y. Gut Microbiome and Metabolites in Systemic Lupus Erythematosus: Link, Mechanisms and Intervention. Front. Immunol. 2021, 12, 686501. [Google Scholar] [CrossRef]
- Markowiak, P.; Slizewska, K. Effect of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef] [PubMed]
- Mu, Q.; Zhang, H.; Liao, X.; Lin, K.; Liu, H.; Edwards, M.R.; Ahmed, S.A.; Yuan, R.; Li, L.; Cecere, T.E.; et al. Control of lupus nephritis by changes of gut microbiota. Microbiome 2017, 5, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemi, A.; Soltani, S.; Ghorabi, S.; Keshtkar, A.; Daneshzad, E.; Nasri, F.; Mazloomi, S.M. Effect of probiotic and synbiotic supplementation on inflammatory markers in health and disease status: A systematic review and meta-analysis of clinical trials. Clin. Nutr. 2020, 39, 789–819. [Google Scholar] [CrossRef] [PubMed]
- Lukiw, W.J. Bacteroides fragilis lipopolysaccharide and inflammatory signaling in Alzheimer’s diseases. Front. Microbiol. 2016, 7, 1544. [Google Scholar] [CrossRef] [Green Version]
- Rezaieyazdi, Z.; Sahebari, M.; Hatef, M.B.; Abbasi, B.; Rafatpanah, H.; Afshari, J.T.; Esmaily, H. Is there any correlation between high sensitive CRP and disease activity in systemic lupus erythematosus? Lupus 2011, 20, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Barnes, E.V.; Narain, S.; Naranjo, A.; Shuster, J.; Segal, M.S.; Sobel, E.S.; E Armstrong, A.; E Santiago, B.; Reeves, W.H.; Richards, H.B. High sensitivity C-reactive protein in systemic lupus erythematosus: Relation to disease activity, clinical presentation and implications for cardiovascular risk. Lupus 2005, 14, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Gheita, T.A.; El-Gazzar, I.I.; Azkalany, G.; El-Fishawy, H.S.; El-Faramawy, A. High-sensitivity C-reactive protein (hs-CRP) in systemic lupus erythematosus patients without cardiac involvement; relation to disease activity, damage and intima-media thickness. Egypt Rheumatol. 2012, 34, 147–152. [Google Scholar] [CrossRef]
- Enocsson, H.; Sjöwall, C.; Kastbom, A.; Skogh, T.; Eloranta, M.-L.; Rönnblom, L.; Wetterö, J. Association of serum c-reactive protein levels with lupus disease activity in the absence of measurable interferon- and a c-reactive protein gene variant. Arthritis Reumatol. 2014, 66, 1568–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazidi, M.; Rezaie, P.; Ferns, G.A.; Vatanparast, H. Impact of probiotic administration on serum C-reactive protein concentrations: Systematic review and meta-analysis of randomized control trials. Nutrients 2017, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M. From CRP to IL-6 to IL-1: Moving upstream to identify novel targets for atheroprotection. Circ. Res. 2016, 118, 145–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vong, L.; Lorentz, R.J.; Assa, A.; Glogauer, M.; Sherman, P.M. Probiotic Lactobacillus rhamnosus inhibits the formation of neutrophil extracellular traps. J. Immunol. 2014, 192, 1870–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, Y.; Chen, X.; Xiao, J.; Zhou, D.B.; Lu, X.X.; Li, W.; Xie, B.; Kuang, X.; Chen, Q. Neutrophil extracellular traps promote lipopolysaccharide-induced airway inflammation and mucus hypersecretion in mice. Oncotarget 2018, 9, 13276–13286. [Google Scholar] [CrossRef] [Green Version]
- Schuster, S.; Hurrell, B.; Tacchini-Cottier, F. Crosstalk between neutrophils and dendritic cells: A context-dependent process. J. Leuk. Biol. 2013, 95, 671–675. [Google Scholar] [CrossRef]
- Yacoub, R.; Jacob, A.; Wlaschin, J.; McGregor, M.; Quigg, R.J.; Alexander, J.J. Lupus: The microbiome angle. Immunobiology 2018, 223, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Chakraborti, C.K. New-found link between microbiota and obesity. World J. Gastrointest Pathophysiol. 2015, 6, 110–119. [Google Scholar] [CrossRef]
- Song, Y.L.; Liu, C.X.; McTeague, M.; Finegold, S.M. Bacteroides nordii sp. nov. and Bacteroides salyersae sp. nov. isolated from clinical specimens of human intestinal origin. J. Clin. Microbiol. 2004, 42, 5565–5570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, J.G.; Kim, H.; Lee, Y.; Kim, M.; Yong, D.; Jeong, S.H.; Lee, K. Bacteroides nordii and Bacteroides salyersiae isolated from post-operative peritonitis patients. Lab. Med. Online 2016, 6, 111–115. [Google Scholar] [CrossRef]
- Schirmer, M.; Smeekens, S.P.; Vlamakis, H.; Jaeger, M.; Oosting, M.; Franzosa, E.A.; Ter Horst, R.; Jansen, T.; Jacobs, L.; Bonder, M.J.; et al. Linking the human gut microbiome to inflammatory cytokine production capacity. Cell 2016, 167, 1125–1136. [Google Scholar] [CrossRef] [Green Version]
- Downes, J.; Munson, M.; Wade, W.G. Dialister invisus sp. nov., isolated from the human oral cavity. Int. J. Syst. Evol. Microbiol. 2003, 53, 1937–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorkildsen, L.T.; Nwosu, F.C.; Avershina, E.; Ricanek, P.; Perminow, G.; Brackmann, S.; Vatn, M.H.; Rudi, K. Dominant fecal microbiota in newly diagnosed untreated inflammatory bowel disease patients. Gastroenterol. Res. Pract. 2013, 636785, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tito, R.Y.; Cypers, H.; Joossens, M.; Varkas, G.; Van Praet, L.; Glorieus, E.; Bosch, F.V.D.; De Vos, M.; Raes, J.; Elewaut, D. Brief report: Dialister as a microbial marker of disease activity in spondyloarthritis. Arthritis Rheumatol. 2017, 69, 114–121. [Google Scholar] [CrossRef]
- Lopetuso, L.R.; Scaldaferri, F.; Petito, V.; Gasbarrini, A. Commensal Clostridia: Leading players in the maintenance of gut homeostasis. Gut Pathog. 2013, 5, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilinskaya, O.N.; Ulyanova, V.V.; Yarullina, D.R.; Gataullin, I.G. Secretome of intestinal Bacilli: A natural guard against pathologies. Front. Microbiol. 2017, 8, 1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Wang, D.; Zhou, Q.; Xu, J. Bacteremia due to vancomycin-resistant Leuconostoc lactis in a patient with pneumonia and abdominal infection. Am. J. Med. Sci. 2015, 349, 282–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchandin, H.; Jumas-Bilak, E. The Family Veillonellaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 433–453. [Google Scholar]
- Jensen, A.; Hoshino, T.; Kilian, M. Taxonomy of the Anginosus group of the genus Streptococcus and description of Streptococcus anginosus subsp. whileyi subsp. nov. and Streptococcus constellatus subsp. viborgensis subsp. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 2506–2519. [Google Scholar] [CrossRef] [PubMed]
- Dewhirst, F.E.; Paster, B.J.; Tzellas, N.; Coleman, B.; Downes, J.; A Spratt, D.; Wade, W. Characterization of novel human oral isolates and cloned 16S rDNA sequences that fall in the family Coriobacteriaceae: Description of Olsenella gen. nov., reclassification of Lactobacillus uli as Olsenella uli comb. nov. and description of Olsenella profusa sp. nov. Int. J. Syst. Evol. Microbiol. 2001, 51, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Chery, J.; Dvoskin, D.; Morato, F.P.; Fahoum, B. Lactobacillus fermentum, a pathogen in documented cholecystitis. Int. J. Surg. Case Rep. 2013, 4, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Malik, I.; Ghosh, S.; Nutt, C.; Macdonald, A.; Bal, A.M.; Collier, A. Gemella haemolysans bacteraemia in a patient with solitary liver abscess. J. Microbiol. Immunol. Infect. 2010, 43, 438–441. [Google Scholar] [CrossRef] [Green Version]
- Hadano, Y.; Kinugasa, Y.; Ohkusu, K.; Ishibashi, K.; Isoda, M. Gemella haemolysans bacteremia in a patient with secondary peritonitis due to a duodenal ulcer perforation: A case report. IDCases 2018, 12, 133–135. [Google Scholar] [CrossRef]
- Uzal, F.A.; Navarro, M.A.; Li, J.; Freedman, J.C.; Shrestha, A.; McClane, B.A. Comparative pathogenesis of enteric clostridial infections in humans and animals. Anaerobe 2018, 53, 11–20. [Google Scholar] [CrossRef]
- Munson, E.; Carroll, K.C. What’s in a Name? New Bacterial Species and Changes to Taxonomic Status from 2012 through 2015. J. Clin. Microbiol. 2016, 55, 24–42. [Google Scholar] [CrossRef] [Green Version]
- Ino, K.; Nakase, K.; Suzuki, K.; Nakamura, A.; Fujieda, A.; Katayama, N. Bacteremia due to Leuconostoc pseudomesenteroides in a patient with acute lymphoblastic leukemia: Case report and review of the literature. Case Rep. Hematol. 2016, 2016, 7648628. [Google Scholar] [CrossRef] [Green Version]
- Sizova, M.V.; Muller, P.A.; Stancyk, D.; Panikov, N.S.; Mandalakis, M.; Hazen, A.; Hohmann, T.; Doerfert, S.N.; Fowle, W.; Earl, A.M.; et al. Oribacterium parvum sp. nov. and Oribacterium asaccharolyticum sp. nov., obligately anaerobic bacteria from the human oral cavity, and emended description of the genus Oribacterium. Int. J. Syst. Evol. Microbiol. 2014, 64, 2642–2649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, V.N.; Dantas, M.J.; Andrade, P.; Pinto-de-Sousa, J. Secondary peritonitis by Actinomyces odontolyticus. Porto Biomed. J. 2017, 2, 174–175. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.J.; Liu, J. Human Microbiota and Ophthalmic Disease. Yale J. Biol. Med. 2016, 89, 325–330. [Google Scholar] [PubMed]
- Tandon, D.; Haque, M.M.; Saravanan, R.; Shaikh, S.; Dubey, A.K.; Mande, S.S. A snapshot of gut microbiota of an adult urban population from Western region of India. PLoS ONE 2018, 13, e0195643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Y.W. Fusobacterium nucleatum: A commensal-turned pathogen. Curr. Opin. Microbiol. 2015, 23, 141–147. [Google Scholar] [CrossRef]
- Ulger-Toprak, N.; Lawson, P.A.; Summanen, P.; O’Neal, L.; Finegold, S.M. Peptoniphilus duerdenii sp. nov. and Peptoniphilus koenoeneniae sp. nov., isolated from human clinical specimens. Int. J. Syst. Evol. Microbiol. 2012, 62, 2336–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christopherson, M.R.; A Dawson, J.; Stevenson, D.M.; Cunningham, A.C.; Bramhacharya, S.; Weimer, P.J.; Kendziorski, C.; Suen, G. Unique aspects of fiber degradation by the ruminal ethanologen Ruminococcus albus 7 revealed by physiological and transcriptomic analysis. Genomics 2014, 15, 1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selma, M.V.; Tomas-Barberan, F.A.; Beltran, D.; Garcia-Villalba, R.; Espin, J.C. Gordonibacter urolithinfaciens sp. nov., a urolithin-producing bacterium isolated from the human gut. Int. J. Syst. Evol. Microbiol. 2014, 64, 2346–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijer, K.; de Vos, P.; Priebe, M.G. Butyrate and other short-chain fatty acids as modulators of immunity: What relevance for health? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 715–721. [Google Scholar] [CrossRef] [PubMed]
- Tygat, H.L.P.; Lebeer, S. The Sweet tooth of bacteria: Common themes in bacterial glycoconjugates. Microbiol. Mol. Biol. Rev. 2014, 78, 372–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawada-Matsuo, M.; Mazda, Y.; Oogai, Y.; Kajiya, M.; Kawai, T.; Yamada, S.; Miyawaki, S.; Oho, T.; Komatsuzawa, H. GlmS and NagB regulate amino sugar metabolism in opposing directions and affect Streptococcus mutans virulence. PLoS ONE 2012, 7, e33382. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptive Characteristics | Synbiotic Treatment (n = 23) | Placebo (n = 23) | p |
|---|---|---|---|
| Age (y) | 35 (24, 40) | 27 (22, 40) | 0.41 |
| SLEDAI-2K score | 14 (9, 18) | 9 (8, 18) | 0.32 |
| Physician general assessment score | 2 (1, 2) | 1 (1, 2) | |
| Organ involvement (%) | |||
| Lupus nephritis | 10 (43) | 10 (43) | 1 |
| Neuropsychiatric lupus | 2 (9) | 2 (9) | 1 |
| Hematology | 2 (9) | 7 (30) | 0.14 |
| Mucocutaneus | 19 (56) | 15 (44) | 0.18 |
| Musculosceletal | 21 (91) | 21 (91) | 1 |
| Antiphospholipid syndrome (%) | 4 (17) | 4 (17) | |
| Steroid dose per day (mg prednisone) | 5 (5, 10) | 5 (2.5, 7.5) | 0.72 |
| Immunosuppressant used (%) | |||
| Hydroxychloroquine | 3 (13) | 2 (9) | 1 |
| Mycophenolate sodium | 18 (78) | 15 (65) | 0.33 |
| Azathioprine | 2 (9) | 5 (22) | 0.41 |
| Cyclosporine | 0 (0) | 2 (9) | 0.5 |
| >1 steroid sparing agent | 2 (8.7) | 1 (4.3) | |
| Vitamin D supplementation (%) | 6 (26) | 2 (9 | 0.24 |
| Other medications (%) | |||
| Proton pump inhibitors | 10 (43) | 11(48) | 1 |
| Statins | 2 (9) | 1(4) | 0.77 |
| BMI (kg/m2) | 23.41 ± 6.19 | 22.09 ± 4.17 | 0.40 |
| hs-CRP (mg/L) | 2.4 (1, 5.9) | 1.9 (0.4, 5.2) | 0.48 |
| Nutrient | Synbiotic Treatment (n = 21) | Placebo (n = 22) | p | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Pre | Post | p | Change Post-pre | Pre | Post | p | Change Post-pre | ||
| Energy (kcal/day) | 1318.54 ± 448.21 | 1024.8 ± 261.83 | 0.003 | −294.27 ± 397.84 | 1352.37 ± 363.06 | 1081.40 ± 392.63 | 0.007 | −270.97 ± 423.70 | 0.85 |
| Fiber (g) | 9.59 (6.70; 11.46) | 5.57 (4.28; 8.50) | 0.002 | −3.85 (−5.03; −0.11) | 8.58 (6.2; 13.88) | 5.96 (4.33; 9.12) | 0.005 | −3.43 (−6.57; 0.43) | 0.81 |
| PUFA (g) | 7.04 (4.43; 10.18) | 4.21 (3.14; 6.41) | 0.02 | −2.35 ± 3.82 | 6.37 (5.01; 10.87) | 5.43 (3.81; 6.60) | 0.06 | −2.48 ± 5.45 | 0.93 |
| Vitamin A (μg) | 835.98 (476.67; 1272.98) | 576.96 (350.04; 1320.07) | 0.46 | −4.05 (−513.18; 177.95) | 705.41 (516.82; 1001.18) | 680.37 (399.91; 1120.00) | 0.76 | −30.14 (−358.16; 268.78) | 0.81 |
| Vitamin E (mg) | 3.6 (2.42; 5.31) | 2.4 (1.54; 2.93) | 0.01 | −0.62 (−2.62; 0.09) | 3.21 (2.07; 4.14) | 1.96 (1.37; 3.07) | 0.01 | −0.74 (−1.98; 0.01) | 0.78 |
| Vitamin C (mg) | 67.06 (33.84; 103.43) | 28.2 (21.49; 41.14) | 0.001 | −20.72 (−47.37; −4.09) | 37.69 (16.46; 65.76) | 27.63 (16.70; 41.03) | 0.006 | −8.47 (−31.38; 0.72) | 0.25 |
| Zinc (mg) | 4.87 ± 1.77 | 3.81 ± 1.51 | 0.02 | −1.06 ± 1.90 | 5.26 ± 2.27 | 4.02 ± 1.56 | 0.01 | −1.25 ± 2.18 | 0.76 |
| Disease Activity | Lupus Nephritis | |||||
|---|---|---|---|---|---|---|
| SLEDAI-2K <6 (Mild Disease Activity) | SLEDAI-2K ≥6 (Moderate Disease Activity) | p | (+) | (−) | p | |
| Bacteroidetes | 43.19 ± 11.45 | 54.22 ± 13.81 | 0.04 | 51.91 ± 14.86 | 52.88 ± 13.74 | 0.87 |
| Firmicutes | 38.59 ± 13.88 | 30.48 ± 12.08 | 0.1 | 51.91 ± 14.86 | 52.88 ± 13.74 | 0.54 |
| Proteobacteria | 10.30 (4.54; 20.52) | 7.05 (5.20; 12.78) | 0.83 | 7.44 (4.92; 10.78) | 6.89 (5.00; 17.89) | 0.42 |
| Actinobacteria | 0.60 (0.37; 0.72) | 0.52 (0.31;1.14) | 0.79 | 0.53 (0.24; 1.05) | 0.59 (0.38; 1.20) | 0.68 |
| Firmicutes/Bacteroidetes ratio | 1.04 (0.56; 1.37) | 0.52 (0.34; 0.72) | 0.06 | 0.57 (0.39; 0.88) | 0.54 (0.30; 1.04) | 0.46 |
| Campylobacter (OTU 149) | 0 (0; 0) | 0 (0; 0.01) | 0.32 | 0 (0; 0.02) | 0 (0; 0) | 0.04 |
| Streptococcus (OTU 062) | 0.62 (0.24; 1.53) | 0.14 (0.06; 0.97) | 0.12 | 0.24 (0.08; 1.46) | 0.13 (0.05; 0.77) | 0.42 |
| Streptococcus anginosus (OTU 241) | 0 (0; 0) | 0.92 | 0 (0; 0) | 0.25 | ||
| Veillonella dispar (OTU 059) | 0.01 (0; 0.14) | 0.01 (0; 0.11) | 0.74 | 0.02 (0; 0.24) | 0.01 (0; 0.05) | 0.24 |
| Ruminococcus gnavus (OTU 068) | 0 (0; 0.01) | 0.02 (0; 0.18) | 0.13 | 0.04 (0; 0.28) | 0.01 (0; 0.12) | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widhani, A.; Djauzi, S.; Suyatna, F.D.; Dewi, B.E. Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial. Cells 2022, 11, 3419. https://doi.org/10.3390/cells11213419
Widhani A, Djauzi S, Suyatna FD, Dewi BE. Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial. Cells. 2022; 11(21):3419. https://doi.org/10.3390/cells11213419
Chicago/Turabian StyleWidhani, Alvina, Samsuridjal Djauzi, Franciscus Dhyanagiri Suyatna, and Beti Ernawati Dewi. 2022. "Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial" Cells 11, no. 21: 3419. https://doi.org/10.3390/cells11213419
APA StyleWidhani, A., Djauzi, S., Suyatna, F. D., & Dewi, B. E. (2022). Changes in Gut Microbiota and Systemic Inflammation after Synbiotic Supplementation in Patients with Systemic Lupus Erythematosus: A Randomized, Double-Blind, Placebo-Controlled Trial. Cells, 11(21), 3419. https://doi.org/10.3390/cells11213419