

Hydrogen Sulfide Regulates SERCA2a Ubiquitylation via Muscle RING Finger-1 S-Sulfhydration to Affect Cardiac Contractility in db/db Mice
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model and Treatment Protocols
2.2. Echocardiography Analysis
2.3. LC-MS/MS Analysis
2.4. Glucose Tolerance Test Analysis
2.5. Immunoprecipitation
2.6. Immunoblot Analysis
2.7. Measurement of SERCA2a Activity
2.8. S-Sulfhydration Assay
2.9. Neonatal Rat Cardiomyocytes Culture
2.10. Cell Experiment
2.11. Point Mutation
2.12. H2S Level Detection with 7-Azido-4-Methylcoumarin
2.13. Measurement of Intracellular Levels of Polysulfide
2.14. [Ca2+]i Assay
2.15. Caspase3 Activity Assays
2.16. Statistical Analysis
3. Results
3.1. Alterations in Body Weight and Glucose Levels in db/db Mice
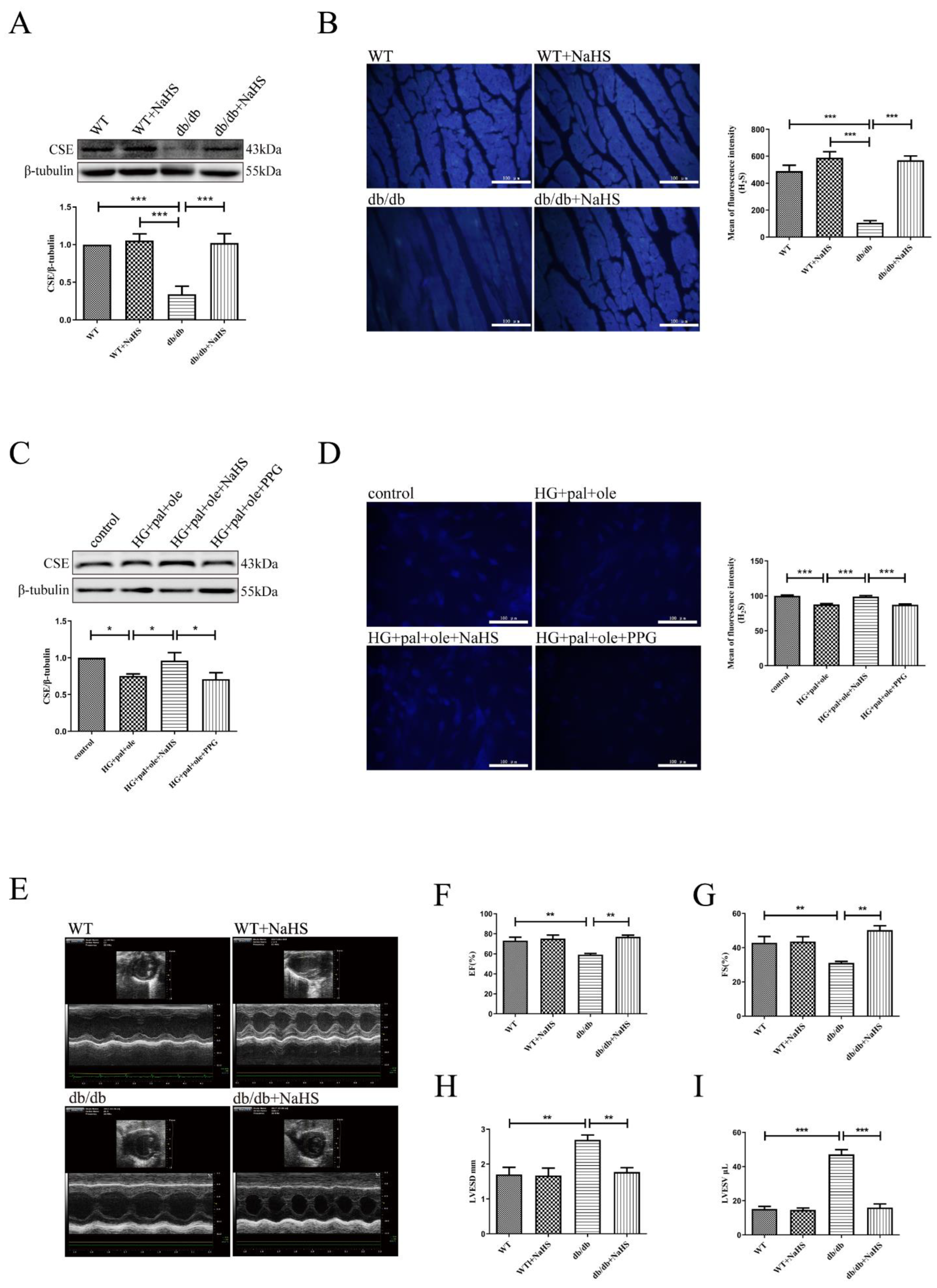
3.2. Decreased CSE Expression and the Effects of H2S on Cardiac Function and Structure in Mice with T2DM
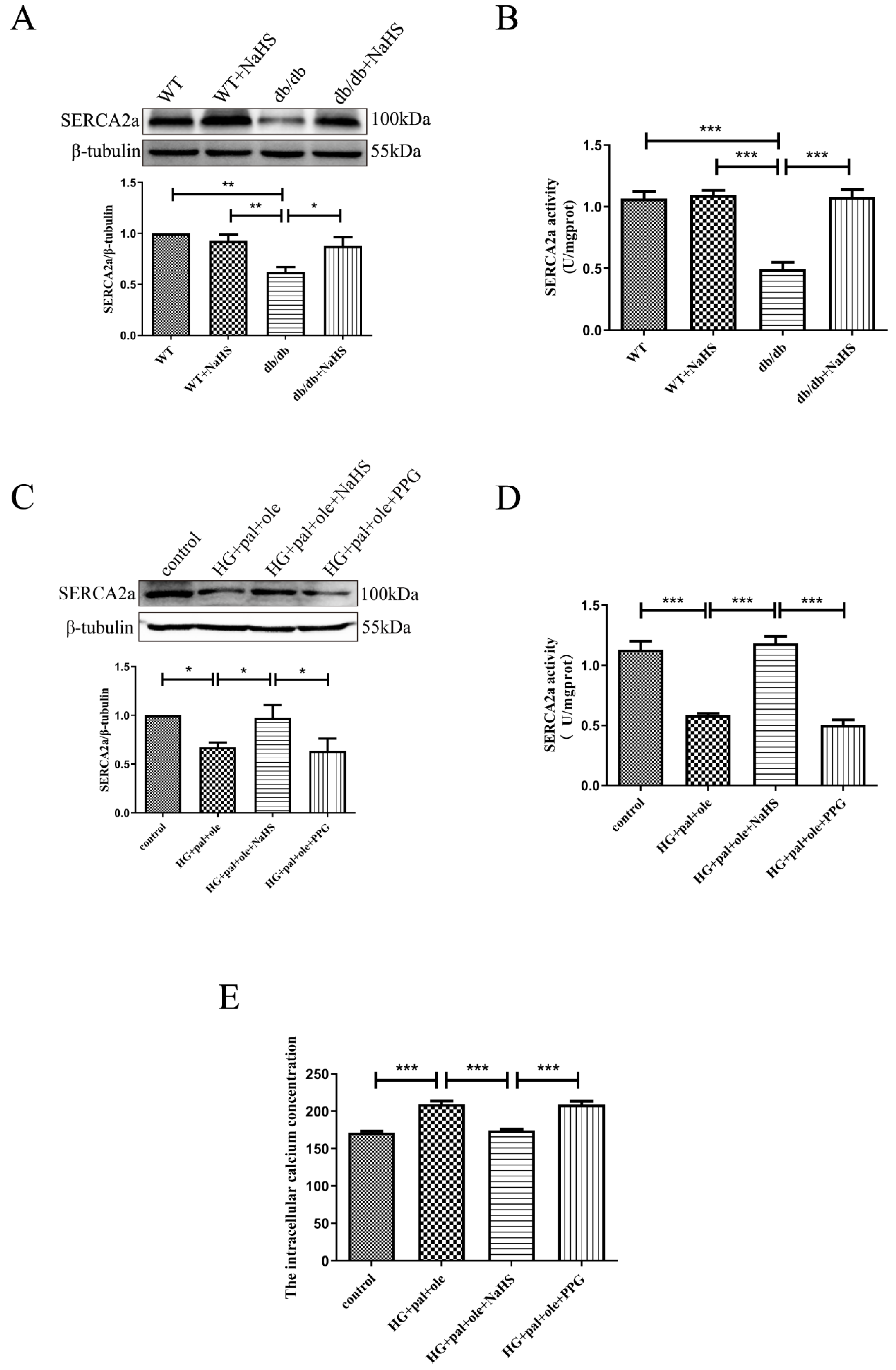
3.3. Exogenous H2S Restored SERCA2a Protein Levels and Activity in db/db Mice
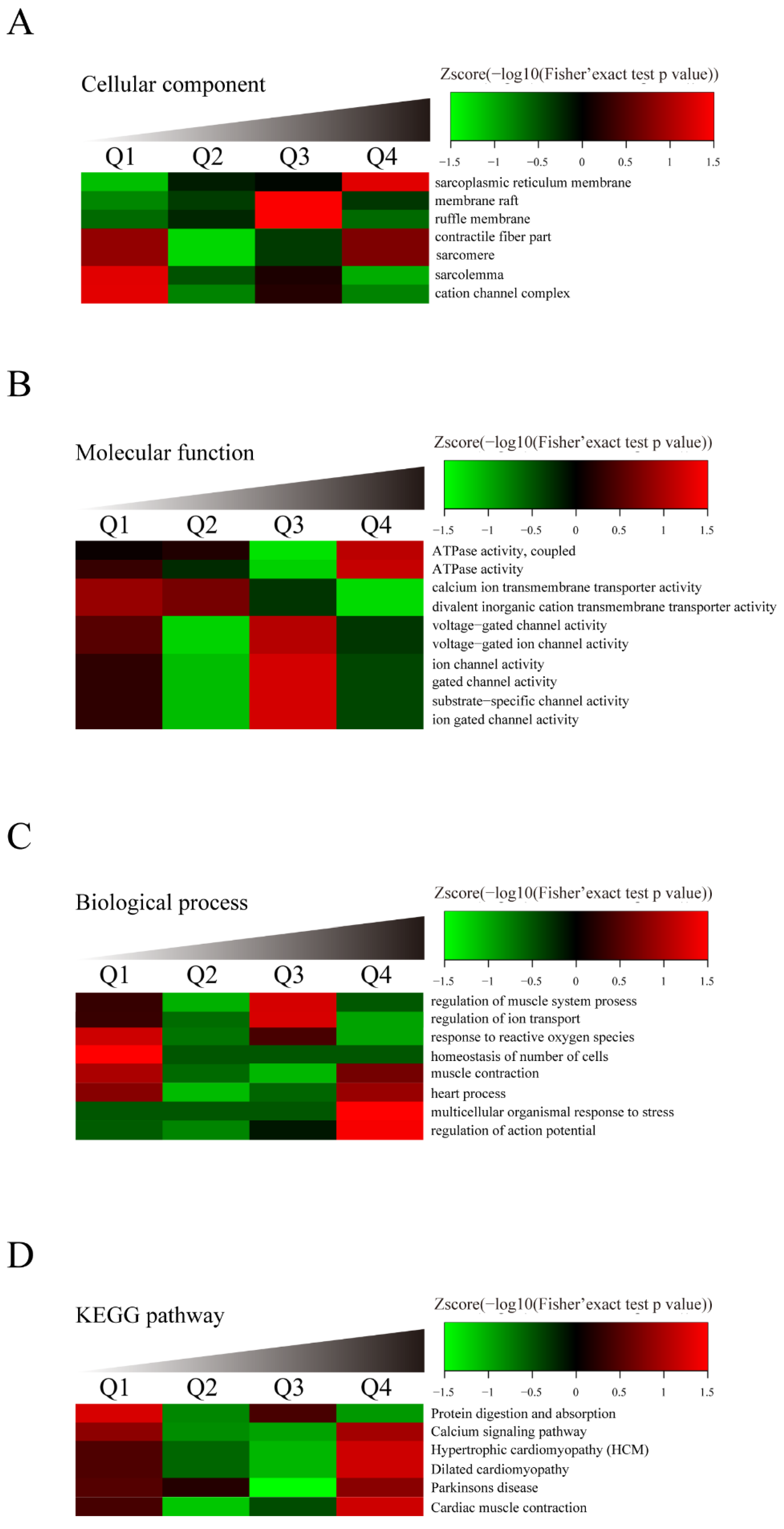
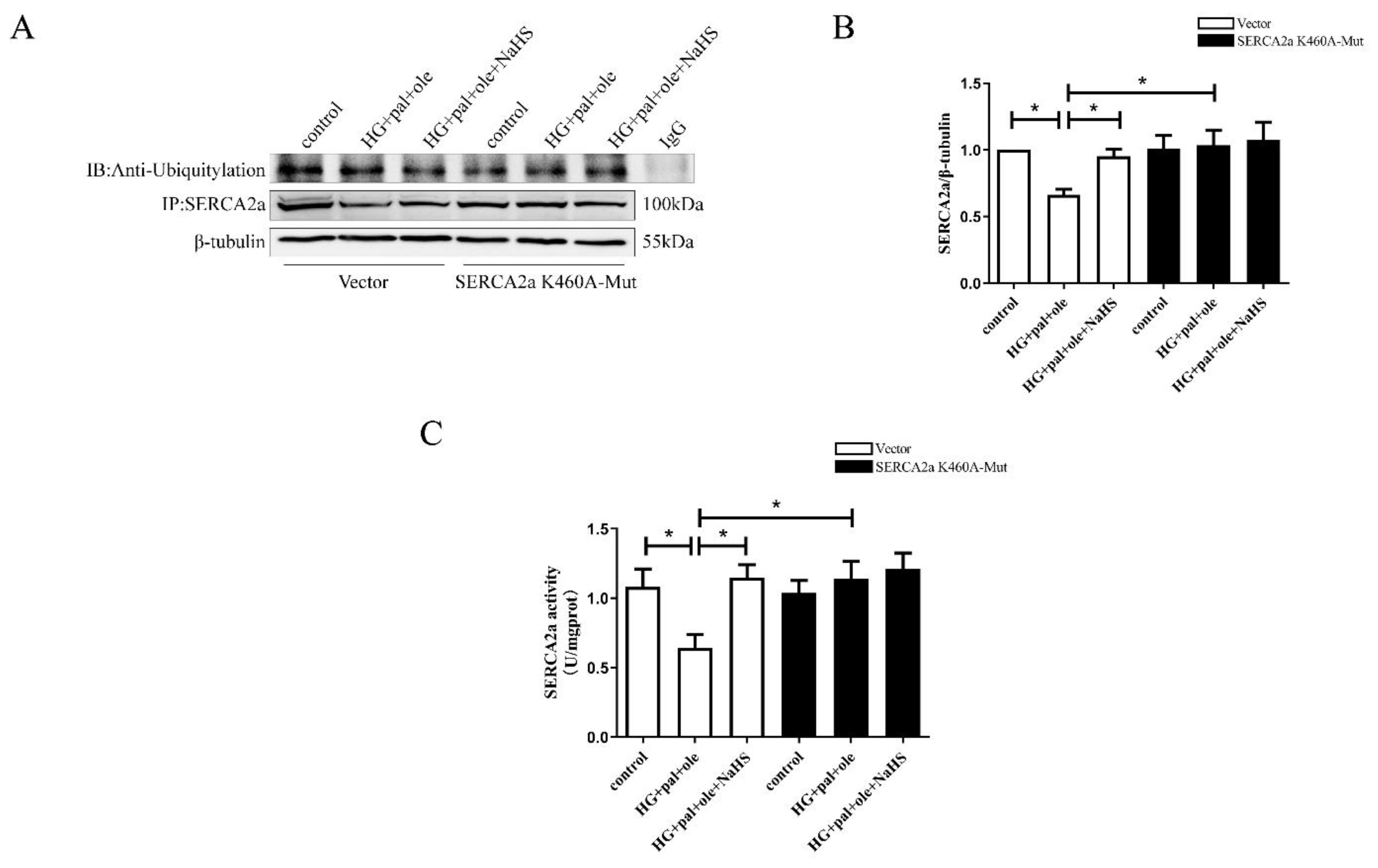
3.4. Identification of SERCA2a Ubiquitylation by LC–MS/MS Analysis
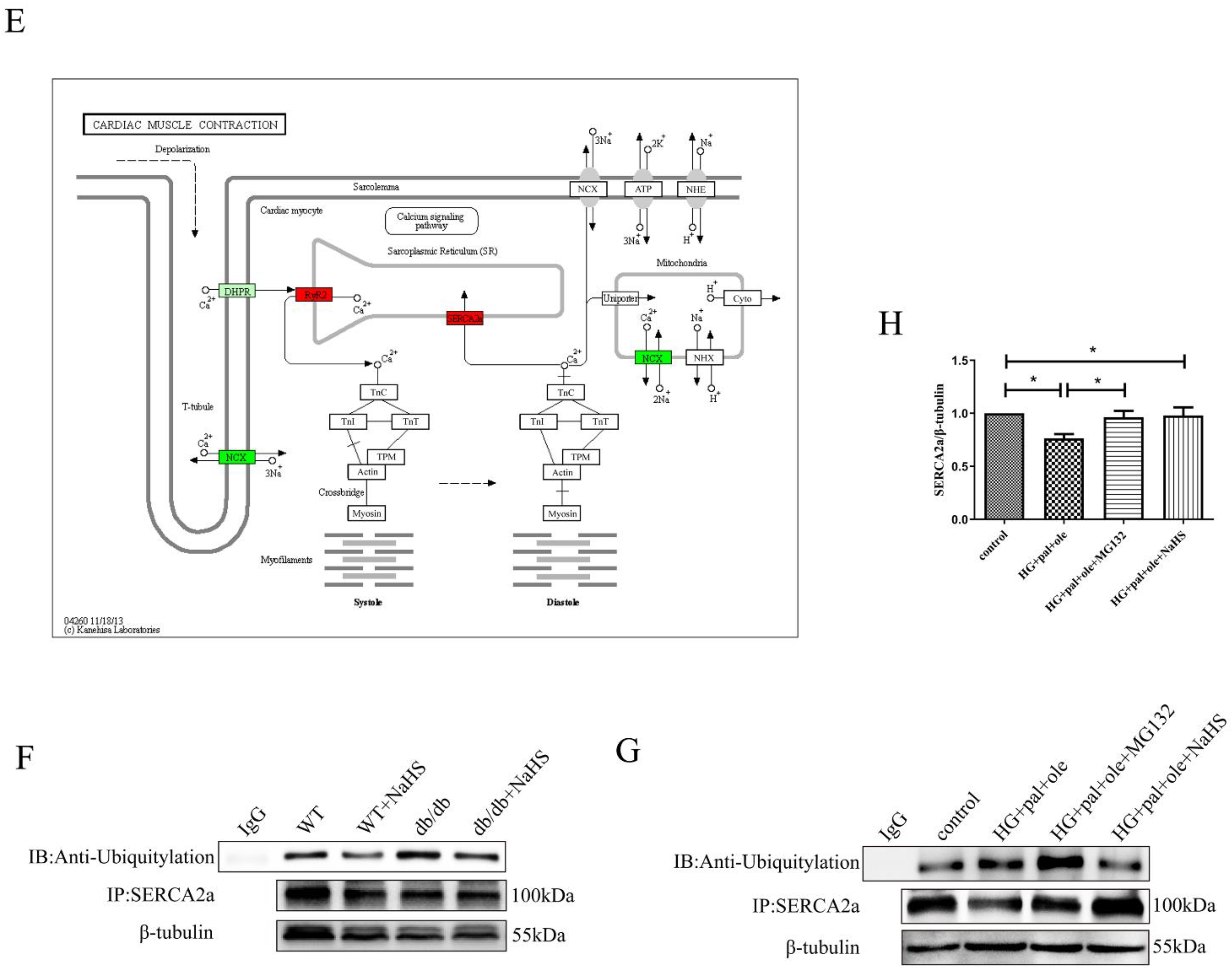
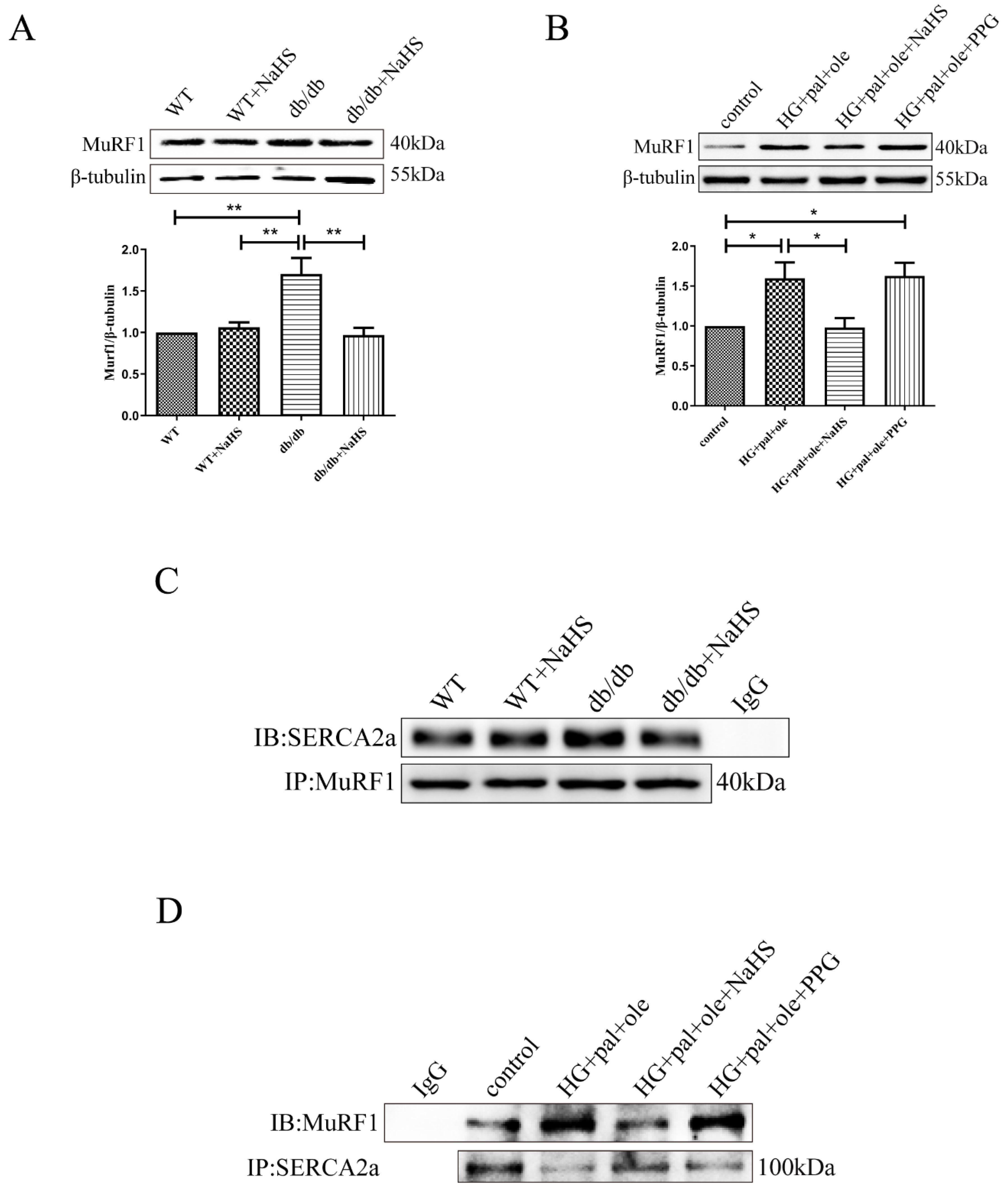
3.5. MuRF1 Is Required for SERCA2a Degradation
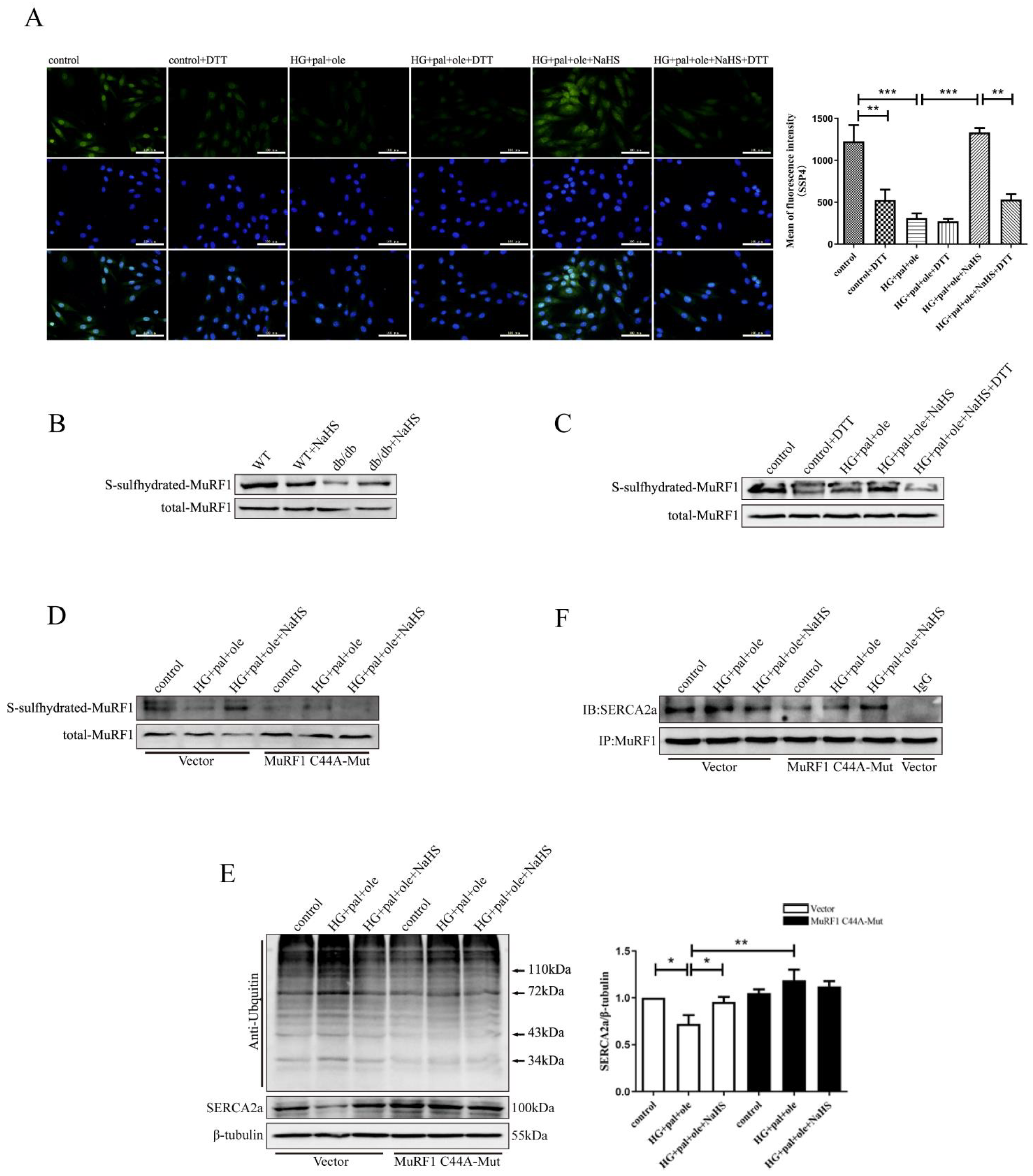
3.6. H2S Promoted MuRF1 S-Sulfhydration to Affect Its Activity
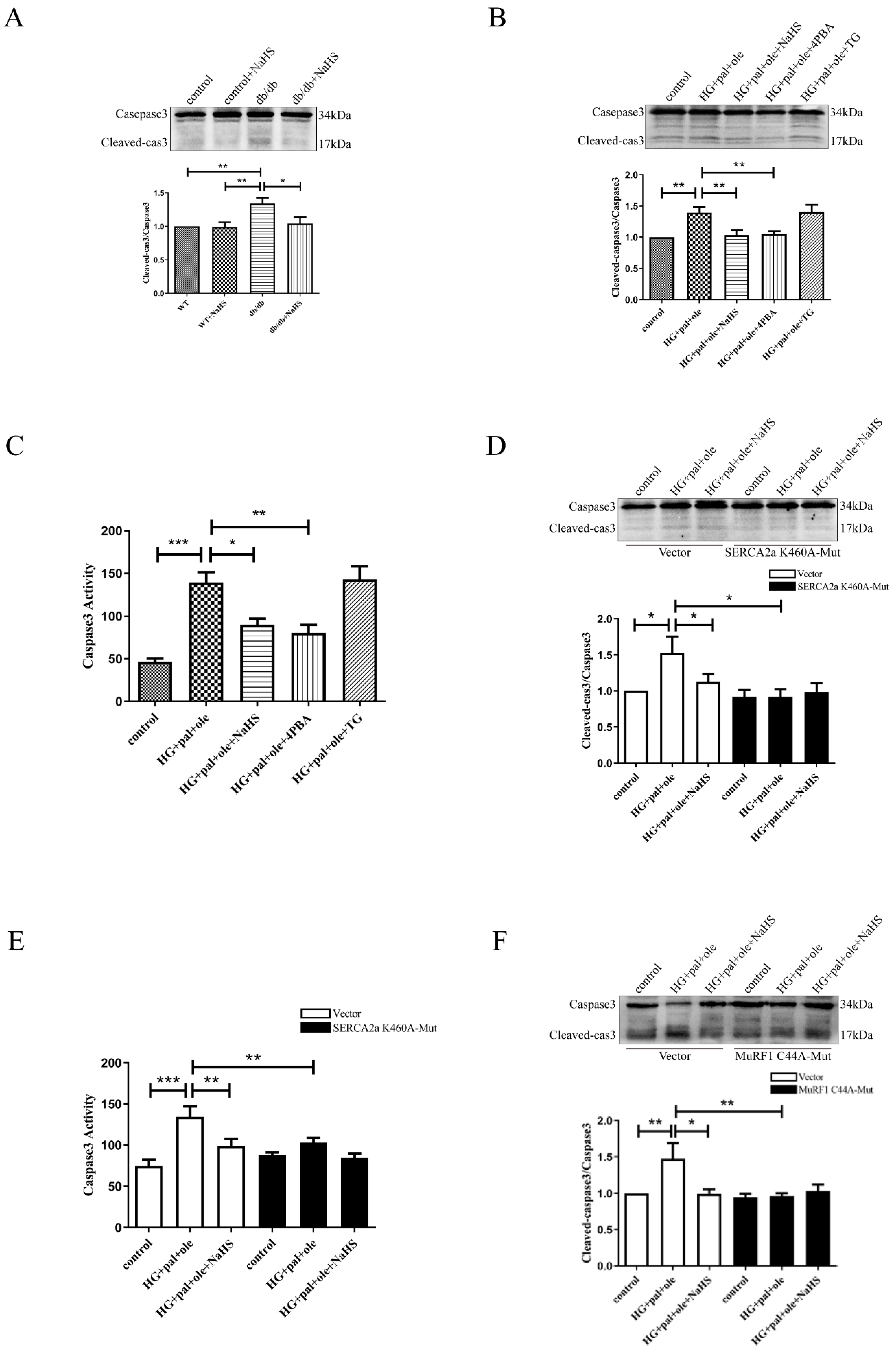
3.7. Exogenous H2S Regulates ER Stress and Apoptosis via SERCA2a Ubiquitylation
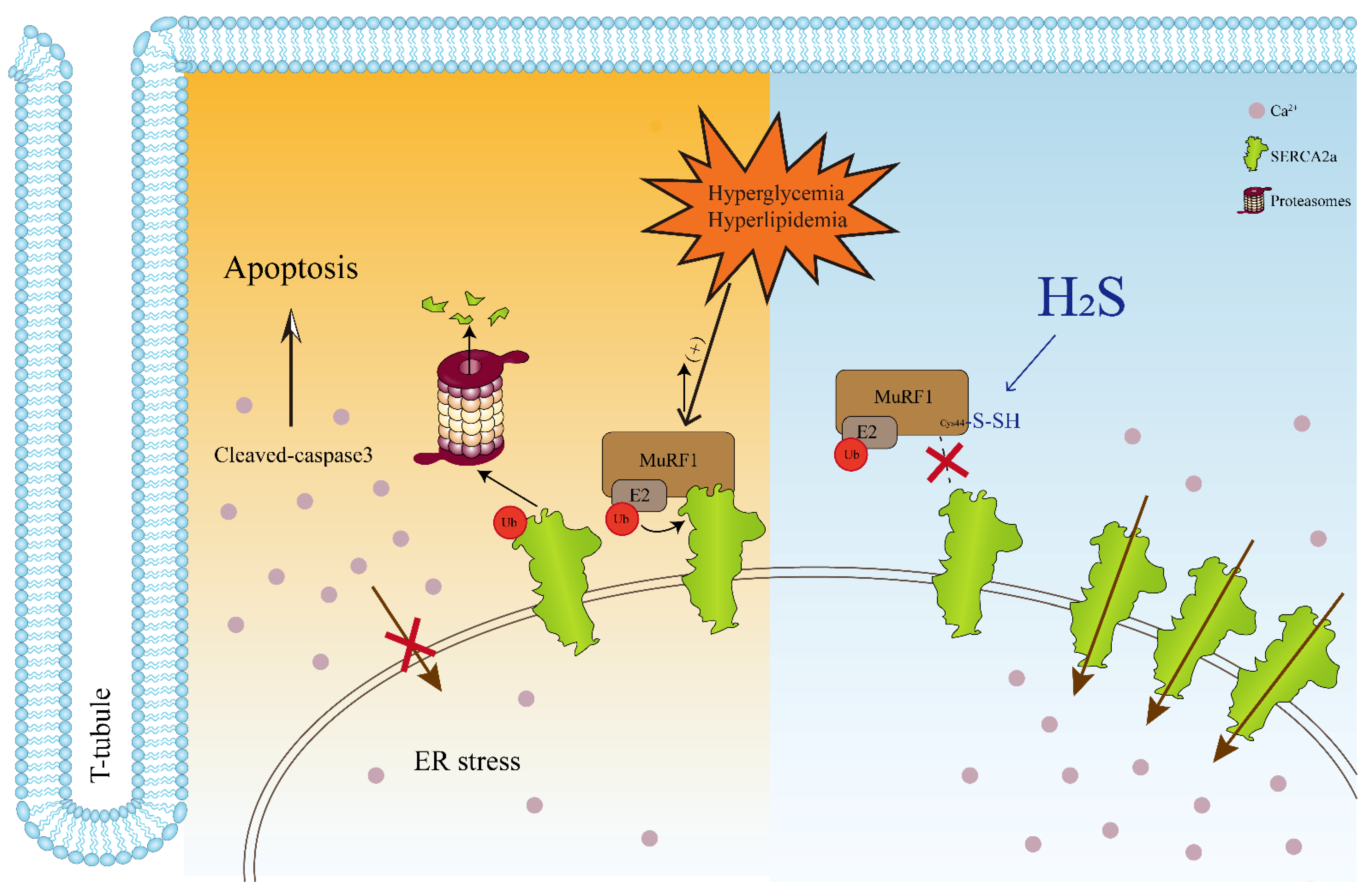
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miki, T.; Yuda, S.; Kouzu, H.; Miura, T. Diabetic cardiomyopathy: Pathophysiology and clinical features. Heart Fail. Rev. 2012, 18, 149–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danaei, G.; Finucane, M.M.; Lu, Y.; Singh, G.M.; Cowan, M.J.; Paciorek, C.J.; Lin, J.K.; Farzadfar, F.; Khang, Y.-H.; Stevens, G.A.; et al. National, regional, and global trends in fasting plasma glucose and diabetes prevalence since 1980: Systematic analysis of health examination surveys and epidemiological studies with 370 country-years and 2·7 million participants. Lancet 2011, 378, 31–40. [Google Scholar] [CrossRef]
- Dillmann, W.H. Diabetic Cardiomyopathy. Circ. Res. 2019, 124, 1160–1162. [Google Scholar] [CrossRef]
- From, A.M.; Leibson, C.L.; Bursi, F.; Redfield, M.M.; Weston, S.A.; Jacobsen, S.J.; Rodeheffer, R.J.; Roger, V.L. Diabetes in heart failure: Prevalence and impact on outcome in the population. Am. J. Med. 2006, 119, 591–599. [Google Scholar] [CrossRef] [PubMed]
- Al Kury, L.T. Calcium Homeostasis in Ventricular Myocytes of Diabetic Cardiomyopathy. J. Diabetes Res. 2020, 2020, 1942086. [Google Scholar] [CrossRef] [PubMed]
- Bidasee, K.R.; Zhang, Y.; Shao, C.H.; Wang, M.; Patel, K.P.; Dincer, U.D.; Besch, H.R. Diabetes increases formation of advanced glycation end products on Sarco(endo)plasmic reticulum Ca2+-ATPase. Diabetes 2004, 53, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Hornbeck, P.V.; Zhang, B.; Murray, B.; Kornhauser, J.M.; Latham, V.; Skrzypek, E. PhosphoSitePlus, 2014: Mutations, PTMs and recalibrations. Nucleic Acids Res. 2014, 43, D512–D520. [Google Scholar] [CrossRef] [Green Version]
- Sitsel, A.; De Raeymaecker, J.; Drachmann, N.D.; Derua, R.; Smaardijk, S.; Andersen, J.L.; Vandecaetsbeek, I.; Chen, J.; De Maeyer, M.; Waelkens, E.; et al. Structures of the heart specific SERCA2a Ca(2+)-ATPase. EMBO J. 2019, 38, e100020. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [Green Version]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef]
- Paul, B.D.; Snyder, S.H. Modes of physiologic H2S signaling in the brain and peripheral tissues. Antioxidants Redox Signal. 2015, 22, 411–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Tian, Z.; Liu, N.; Zhang, L.; Gao, Z.; Sun, X.; Yu, M.; Wu, J.; Yang, F.; Zhao, Y.; et al. Exogenous H2S switches cardiac energy substrate metabolism by regulating SIRT3 expression in db/db mice. J. Mol. Med. 2018, 96, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Du, H.; Wang, B.; Chen, J.; Lu, F.; Peng, S.; Sun, Y.; Liu, N.; Sun, X.; Shiyun, D.; et al. Exogenous H2S Induces Hrd1 S-sulfhydration and Prevents CD36 Translocation via VAMP3 Ubiquitylation in Diabetic Hearts. Aging Dis. 2020, 11, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Yu, X.; Li, T.; Wu, J.; Zhao, Y.; Liu, J.; Sun, A.; Dong, S.; Wu, J.; Zhong, X.; et al. Exogenous H2S regulates endoplasmic reticulum-mitochondria cross-talk to inhibit apoptotic pathways in STZ-induced type I diabetes. Am. J. Physiol. Endocrinol. Metab. 2017, 312, E190–E203. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jiao, L.; Sun, L.; Li, Y.; Gao, Y.; Xu, C.; Shao, Y.; Li, M.; Li, C.; Lu, Y.; et al. LncRNA ZFAS1 as a SERCA2a Inhibitor to Cause Intracellular Ca 2+ Overload and Contractile Dysfunction in a Mouse Model of Myocardial Infarction. Circ. Res. 2018, 122, 1354–1368. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y. KEGG Mapper for inferring cellular functions from protein sequences. Protein Sci. 2020, 29, 28–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2022, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef]
- Stammers, A.N.; Susser, S.E.; Hamm, N.C.; Hlynsky, M.W.; Kimber, D.E.; Kehler, D.S.; Duhamel, T.A. The regulation of sarco(endo)plasmic reticulum calcium-ATPases (SERCA). Can. J. Physiol. Pharmacol. 2015, 93, 843–854. [Google Scholar] [CrossRef]
- Møller, J.V.; Olesen, C.; Winther, A.-M.L.; Nissen, P. The sarcoplasmic Ca2+-ATPase: Design of a perfect chemi-osmotic pump. Q. Rev. Biophys. 2010, 43, 501–566. [Google Scholar] [CrossRef]
- Centner, T.; Yano, J.; Kimura, E.; McElhinny, A.S.; Pelin, K.; Witt, C.C.; Bang, M.-L.; Trombitas, K.; Granzier, H.; Gregorio, C.C.; et al. Identification of muscle specific ring finger proteins as potential regulators of the titin kinase domain. J. Mol. Biol. 2001, 306, 717–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazaki, M.; Esser, K.A. Cellular mechanisms regulating protein synthesis and skeletal muscle hypertrophy in animals. J. Appl. Physiol. 2009, 106, 1367–1373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitch, W.E.; Goldberg, A.L. Mechanisms of muscle wasting. The role of the ubiquitin-proteasome pathway. N. Engl. J. Med. 1996, 335, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Bodine, S.C.; Baehr, L.M. Skeletal muscle atrophy and the E3 ubiquitin ligases MuRF1 and MAFbx/atrogin-1. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E469–E484. [Google Scholar] [CrossRef] [Green Version]
- Golebiowski, F.; Matic, I.; Tatham, M.H.; Cole, C.; Yin, Y.; Nakamura, A.; Cox, J.; Barton, G.J.; Mann, M.; Hay, R.T. System-wide changes to SUMO modifications in response to heat shock. Sci. Signal. 2009, 2, ra24. [Google Scholar] [CrossRef] [Green Version]
- Linden, D.R. Hydrogen Sulfide Signaling in the Gastrointestinal Tract. Antioxidants Redox Signal. 2014, 20, 818–830. [Google Scholar] [CrossRef]
- Sun, X.; Zhao, D.; Lu, F.; Peng, S.; Yu, M.; Liu, N.; Sun, Y.; Du, H.; Wang, B.; Chen, J.; et al. Hydrogen sulfide regulates muscle RING finger-1 protein S-sulfhydration at Cys 44 to prevent cardiac structural damage in diabetic cardiomyopathy. Br. J. Pharmacol. 2019, 177, 836–856. [Google Scholar] [CrossRef]
- Fu, S.; Yang, L.; Li, P.; Hofmann, O.; Dicker, L.; Hide, W.; Lin, X.; Watkins, S.M.; Ivanov, A.R.; Hotamisligil, G.S. Aberrant lipid metabolism disrupts calcium homeostasis causing liver endoplasmic reticulum stress in obesity. Nature 2011, 473, 528–531. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Tikunova, S.B.; Kline, K.P.; Siddiqui, J.K.; Davis, J.P. Disease-related cardiac troponins alter thin filament Ca2+ association and dissociation rates. PLoS ONE 2012, 7, e38259. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, C.; Iwasawa, S.; Ogawa, H.; Hirata, A.; Tsueda, J.; Inesi, G. Crystal structures of the calcium pump and sarcolipin in the Mg2+-bound E1 state. Nature 2013, 495, 260–264. [Google Scholar] [CrossRef]
- Winther, A.-M.L.; Bublitz, M.; Karlsen, J.L.; Møller, J.V.; Hansen, J.B.; Nissen, P.; Buch-Pedersen, M.J. The sarcolipin-bound calcium pump stabilizes calcium sites exposed to the cytoplasm. Nature 2013, 495, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Budhidarmo, R.; Nakatani, Y.; Day, C.L. RINGs hold the key to ubiquitin transfer. Trends Biochem. Sci. 2012, 37, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Baehr, L.M.; Hughes, D.C.; Lynch, S.A.; Van Haver, D.; Maia, T.M.; Marshall, A.G.; Radoshevich, L.; Impens, F.; Waddell, D.S.; Bodine, S.C. Identification of the MuRF1 Skeletal Muscle Ubiquitylome Through Quantitative Proteomics. Function 2021, 2, zqab029. [Google Scholar] [CrossRef] [PubMed]
- Iciek, M.; Kowalczyk-Pachel, D.; Bilska-Wilkosz, A.; Kwiecień, I.; Górny, M.; Włodek, L. S-sulfhydration as a cellular redox regulation. Biosci. Rep. 2015, 36, e00304. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, S.; Zhao, D.; Li, Q.; Wang, M.; Zhang, S.; Pang, K.; Huang, J.; Lu, F.; Chen, H.; Zhang, W. Hydrogen Sulfide Regulates SERCA2a Ubiquitylation via Muscle RING Finger-1 S-Sulfhydration to Affect Cardiac Contractility in db/db Mice. Cells 2022, 11, 3465. https://doi.org/10.3390/cells11213465
Peng S, Zhao D, Li Q, Wang M, Zhang S, Pang K, Huang J, Lu F, Chen H, Zhang W. Hydrogen Sulfide Regulates SERCA2a Ubiquitylation via Muscle RING Finger-1 S-Sulfhydration to Affect Cardiac Contractility in db/db Mice. Cells. 2022; 11(21):3465. https://doi.org/10.3390/cells11213465
Chicago/Turabian StylePeng, Shuo, Dechao Zhao, Qianzhu Li, Mengyi Wang, Shiwu Zhang, Kemiao Pang, Jiayi Huang, Fanghao Lu, He Chen, and Weihua Zhang. 2022. "Hydrogen Sulfide Regulates SERCA2a Ubiquitylation via Muscle RING Finger-1 S-Sulfhydration to Affect Cardiac Contractility in db/db Mice" Cells 11, no. 21: 3465. https://doi.org/10.3390/cells11213465
APA StylePeng, S., Zhao, D., Li, Q., Wang, M., Zhang, S., Pang, K., Huang, J., Lu, F., Chen, H., & Zhang, W. (2022). Hydrogen Sulfide Regulates SERCA2a Ubiquitylation via Muscle RING Finger-1 S-Sulfhydration to Affect Cardiac Contractility in db/db Mice. Cells, 11(21), 3465. https://doi.org/10.3390/cells11213465