

Development of a Lung Vacancy Mouse Model through CRISPR/Cas9-Mediated Deletion of Thyroid Transcription Factor 1 Exon 2

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Cas9 mRNA and sgRNA
2.3. Microinjection
2.4. Genotyping and Sequencing
2.5. Quantitative RT-PCR (qRT-PCR) Analyses
2.6. Hematoxylin and Eosin (H&E) Staining and Immunohistochemistry (IHC) Analysis
2.7. Immunofluorescence (IF) Analysis
2.8. Statistical Analysis
3. Results
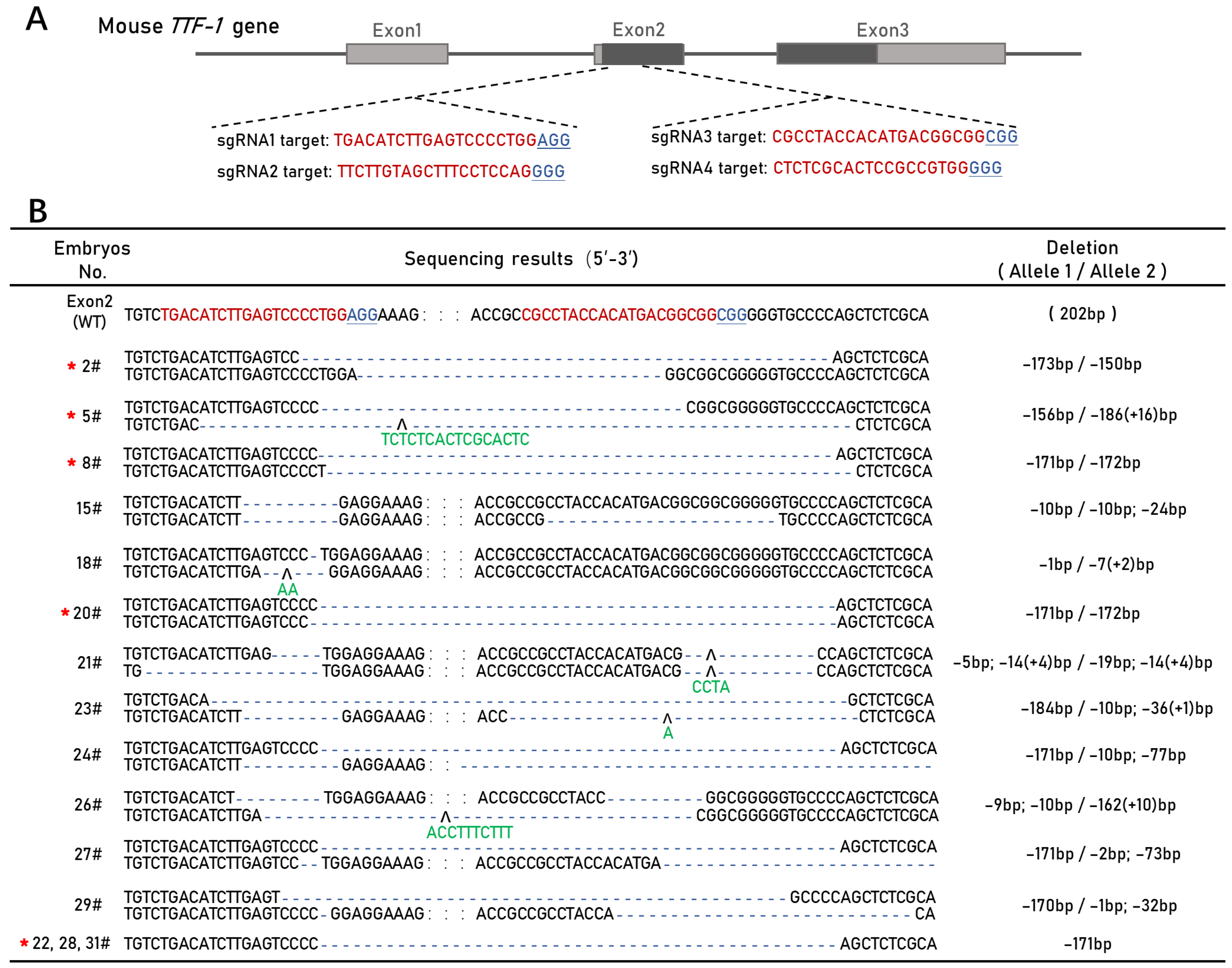
3.1. CRISPR/Cas9-Mediated Mouse TTF-1 Exon 2 Deletion
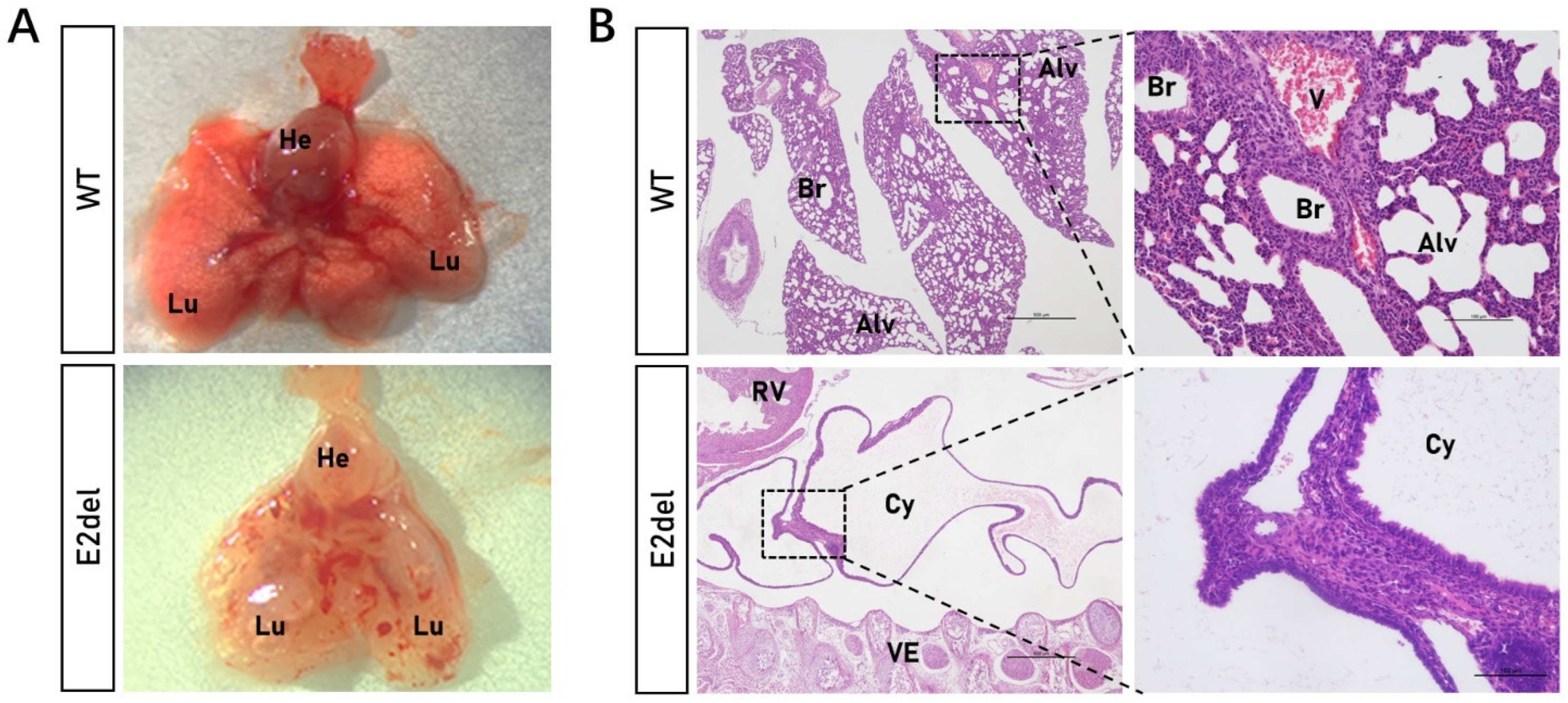
3.2. The E2del Mouse Embryos Displayed Bilateral Sac-like Lungs
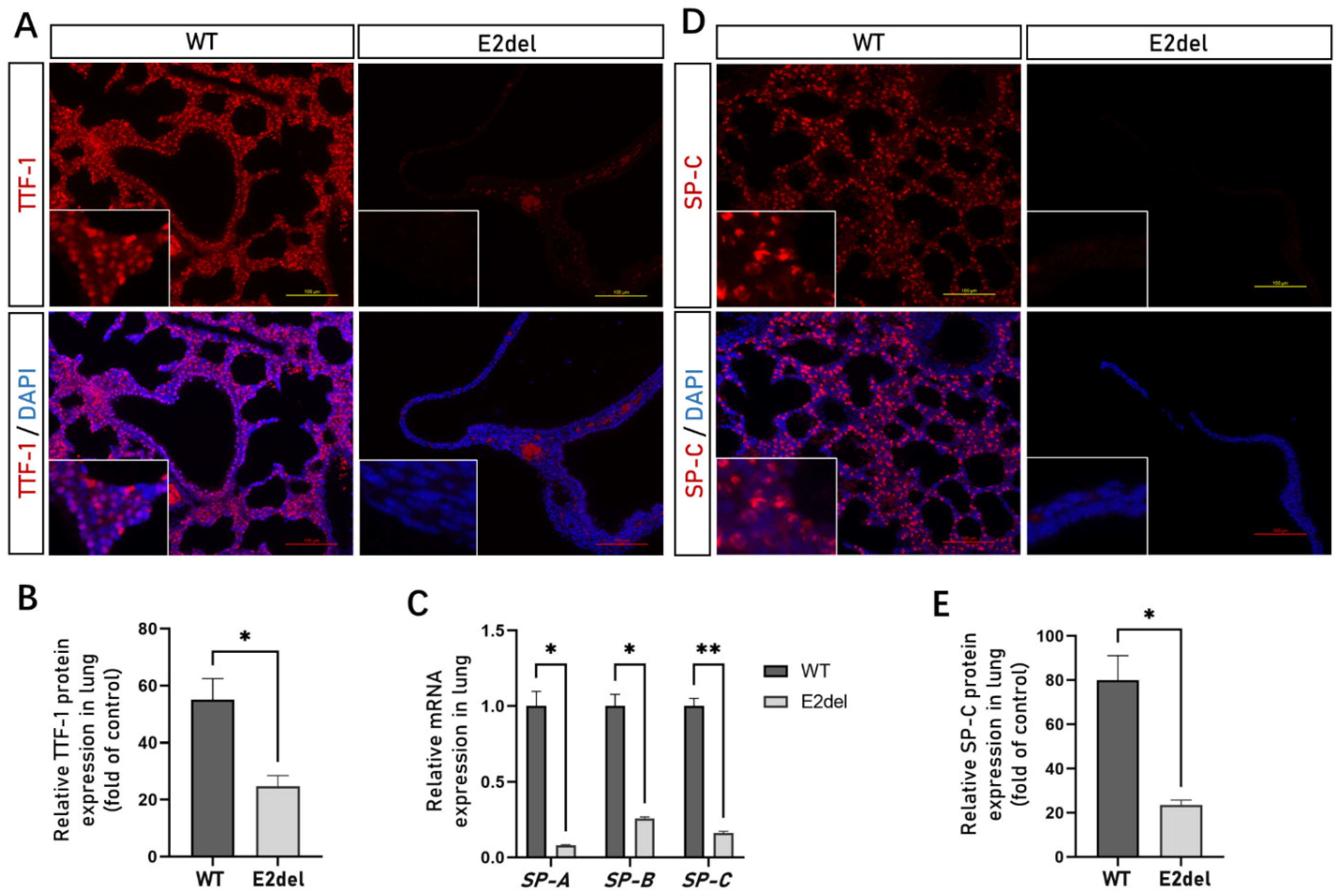
3.3. The E2del Lungs Lacked Alveolar Type II Cells
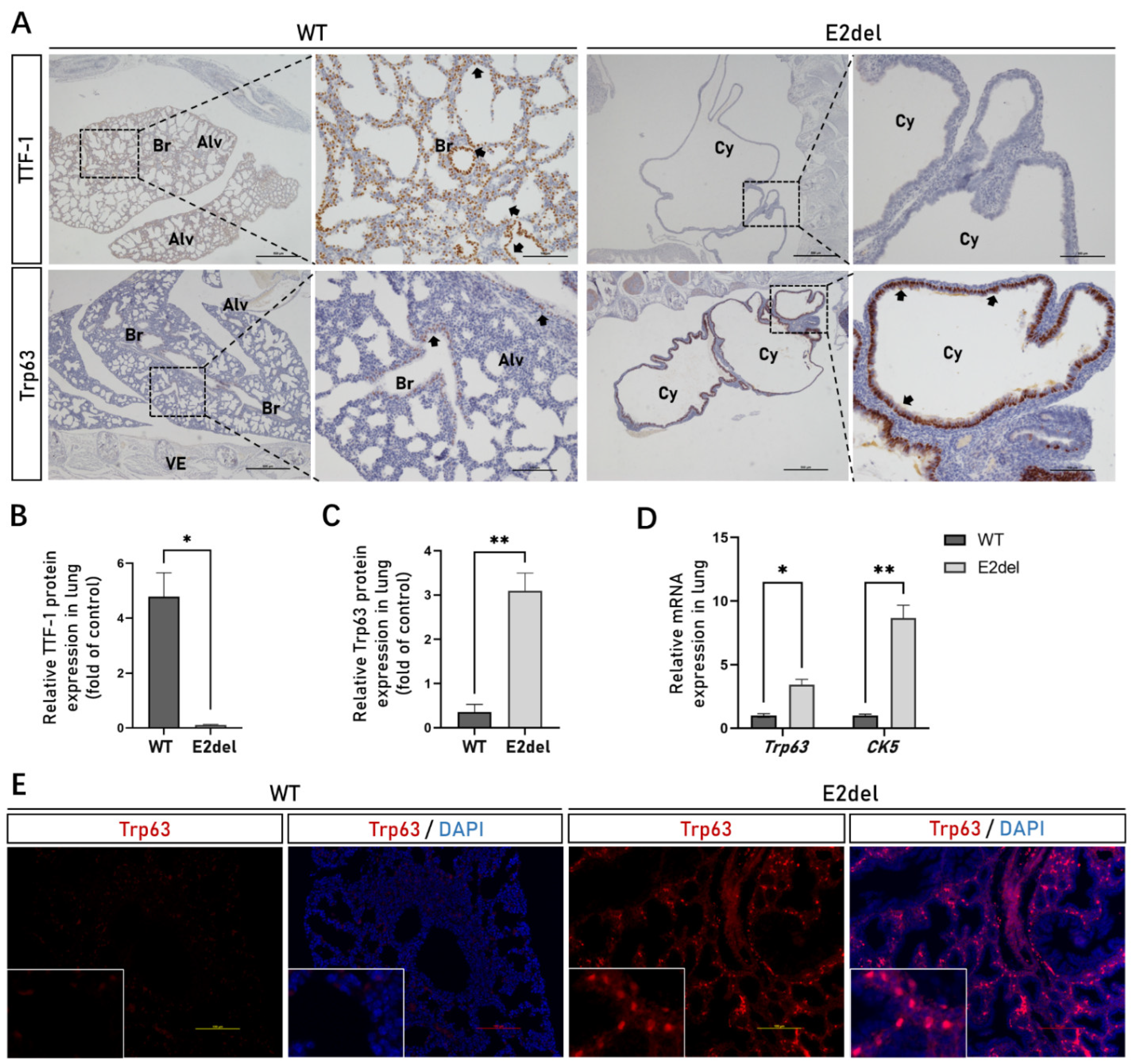
3.4. The E2del Lungs Contained Abnormally Abundant Basal Cells
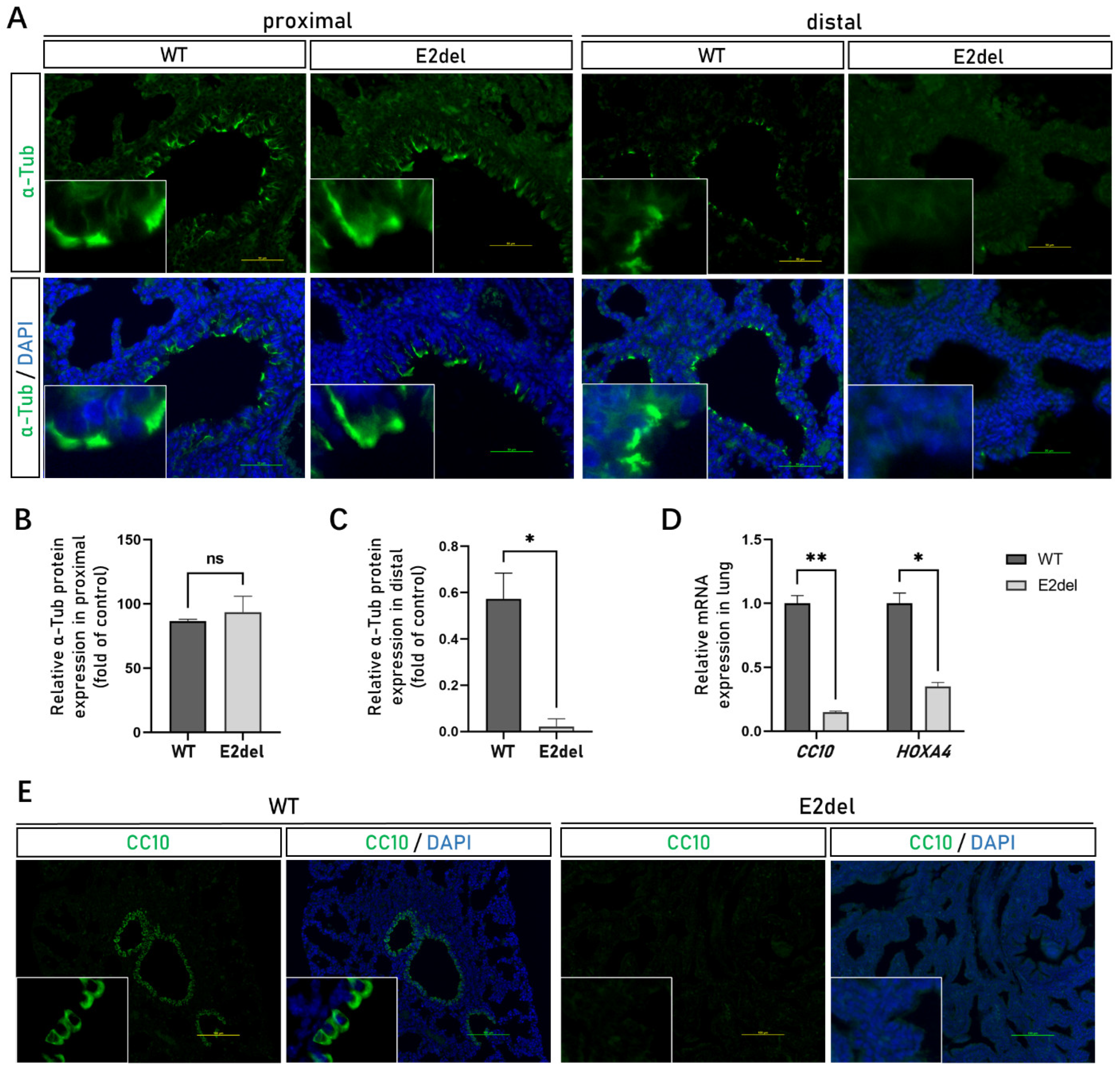
3.5. The Abnormal Development of the Other Functional Cells in the E2del Lungs
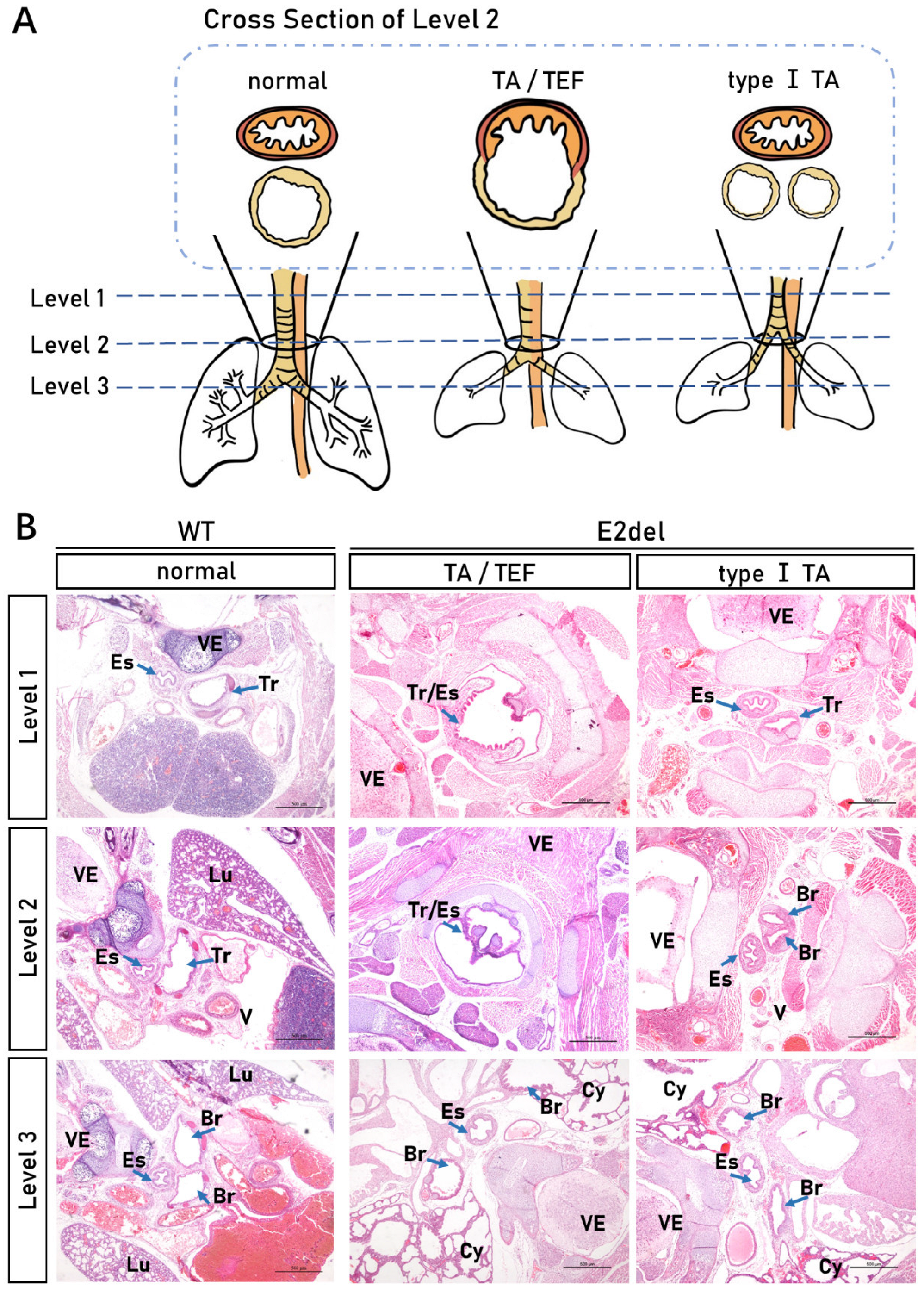
3.6. The E2del Mouse Embryos Showed Malformation Type I TA or TA/TEF
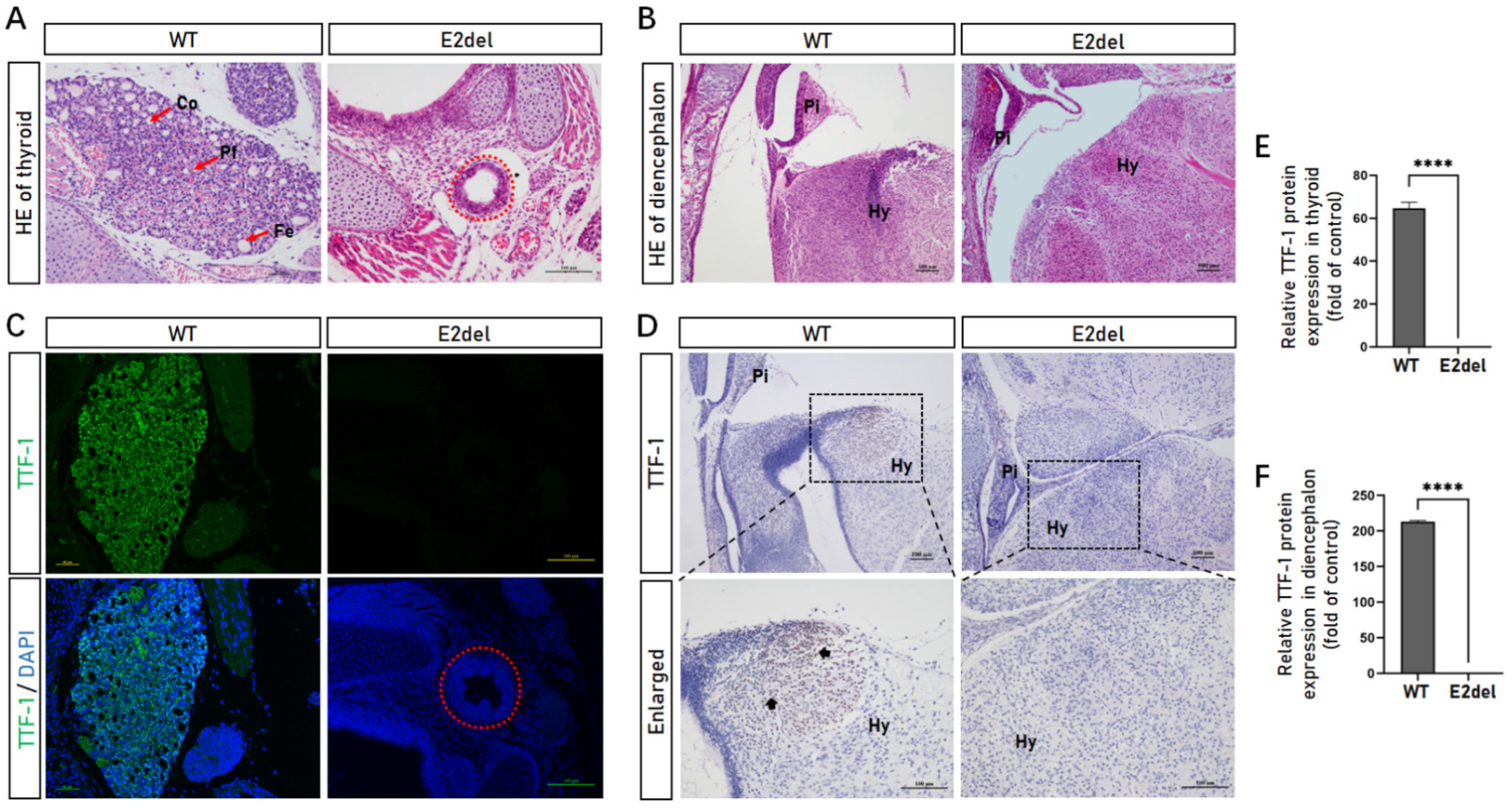
3.7. The E2del Mouse Embryos Had Thyroid Developed and Diencephalon Abnormalities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.; Lansford, R.; Stewart, V.; Young, F.; Alt, F.W. RAG-2-deficient blastocyst complementation: An assay of gene function in lymphocyte development. Proc. Natl. Acad. Sci. USA 1993, 90, 4528–4532. [Google Scholar] [CrossRef] [Green Version]
- Matsunari, H.; Nagashima, H.; Watanabe, M.; Umeyama, K.; Nakano, K.; Nagaya, M.; Kobayashi, T.; Yamaguchi, T.; Sumazaki, R.; Herzenberg, L.A.; et al. Blastocyst complementation generates exogenic pancreas in vivo in apancreatic cloned pigs. Proc. Natl. Acad. Sci. USA 2013, 110, 4557–4562. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Yamaguchi, T.; Hamanaka, S.; Kato-Itoh, M.; Yamazaki, Y.; Ibata, M.; Sato, H.; Lee, Y.S.; Usui, J.; Knisely, A.S.; et al. Generation of rat pancreas in mouse by interspecific blastocyst injection of pluripotent stem cells. Cell 2010, 142, 787–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, T.; Sato, H.; Kato-Itoh, M.; Goto, T.; Hara, H.; Sanbo, M.; Mizuno, N.; Kobayashi, T.; Yanagida, A.; Umino, A.; et al. Interspecies organogenesis generates autologous functional islets. Nature 2017, 542, 191–196. [Google Scholar] [CrossRef]
- Matsunari, H.; Watanabe, M.; Hasegawa, K.; Uchikura, A.; Nakano, K.; Umeyama, K.; Masaki, H.; Hamanaka, S.; Yamaguchi, T.; Nagaya, M.; et al. Compensation of disabled organogeneses in genetically modified pig fetuses by blastocyst complementation. Stem Cell Rep. 2020, 14, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Fraidenraich, D.; Stillwell, E.; Romero, E.; Wilkes, D.; Manova, K.; Basson, C.T.; Benezra, R. Rescue of cardiac defects in id knockout embryos by injection of embryonic stem cells. Science 2004, 306, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sturzu, A.C.; Rajarajan, K.; Passer, D.; Plonowska, K.; Riley, A.; Tan, T.C.; Sharma, A.; Xu, A.F.; Engels, M.C.; Feistritzer, R.; et al. Fetal mammalian heart generates a robust compensatory response to cell loss. Circulation 2015, 132, 109–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espejel, S.; Roll, G.R.; McLaughlin, K.J.; Lee, A.Y.; Zhang, J.Y.; Laird, D.J.; Okita, K.; Yamanaka, S.; Willenbring, H. Induced pluripotent stem cell-derived hepatocytes have the functional and proliferative capabilities needed for liver regeneration in mice. J. Clin. Investig 2010, 120, 3120–3126. [Google Scholar] [CrossRef] [Green Version]
- Usui, J.I.; Kobayashi, T.; Yamaguchi, T.; Knisely, A.S.; Nishinakamura, R.; Nakauchi, H. Generation of kidney from pluripotent stem cells via blastocyst complementation. Am. J. Pathol. 2012, 180, 2417–2426. [Google Scholar] [CrossRef]
- Goto, T.; Hara, H.; Sanbo, M.; Masaki, H.; Sato, H.; Yamaguchi, T.; Hochi, S.; Kobayashi, T.; Nakauchi, H.; Hirabayashi, M. Generation of pluripotent stem cell-derived mouse kidneys in Sall1-targeted anephric rats. Nat. Commun. 2019, 10, 451. [Google Scholar] [CrossRef]
- Kitahara, A.; Ran, Q.; Oda, K.; Yasue, A.; Abe, M.; Ye, X.; Sasaoka, T.; Tsuchida, M.; Sakimura, K.; Ajioka, Y.; et al. Generation of lungs by blastocyst complementation in Apneumic Fgf10-deficient mice. Cell Rep. 2020, 31, 107626. [Google Scholar] [CrossRef]
- Mori, M.; Furuhashi, K.; Danielsson, J.A.; Hirata, Y.; Kakiuchi, M.; Lin, C.S.; Ohta, M.; Riccio, P.; Takahashi, Y.; Xu, X.; et al. Generation of functional lungs via conditional blastocyst complementation using pluripotent stem cells. Nat. Med. 2019, 25, 1691–1698. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Li, E.; Ustiyan, V.; Wang, G.; Guo, M.; Na, C.L.; Kalin, G.T.; Galvan, V.; Xu, Y.; Weaver, T.E.; et al. In Vivo generation of lung and thyroid tissues from embryonic stem cells using blastocyst complementation. Am. J. Respir. Crit. Care. Med. 2020, 203, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Sekine, K.; Ohuchi, H.; Fujiwara, M.; Yamasaki, M.; Yoshizawa, T.; Sato, T.; Yagishita, N.; Matsui, D.; Koga, Y.; Itoh, N.; et al. Fgf10 is essential for limb and lung formation. Nat. Genet. 1999, 21, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Ustiyan, V.; Wen, B.; Kalin, G.T.; Whitsett, J.A.; Kalin, T.V.; Kalinichenko, V.V. Blastocyst complementation reveals that NKX2-1 establishes the proximal-peripheral boundary of the airway epithelium. Dev. Dyn. 2021, 250, 1001–1020. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Hara, Y.; Pineau, T.; Fernandez-Salguero, P.; Fox, C.H.; Ward, J.M.; Gonzalez, F.J. The T/ebp null mouse: Thyroid-specific enhancer-binding protein is essential for the organogenesis of the thyroid, lung, ventral forebrain, and pituitary. Genes Dev. 1996, 10, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Minoo, P.; Su, G.; Drum, H.; Bringas, P.; Kimura, S. Defects in tracheoesophageal and lung morphogenesis in Nkx2.1(-/-) mouse embryos. Dev. Biol. 1999, 209, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Smith, T.H.; Battle, S.L.; Ferrell, S.; Hawkins, R.D. Superovulation alters global DNA methylation in early mouse embryo development. Epigenetics 2019, 14, 780–790. [Google Scholar] [CrossRef]
- Boggaram, V. Thyroid transcription factor-1 (TTF-1/Nkx2.1/TITF1) gene regulation in the lung. Clin. Sci. 2009, 116, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Rock, J.R.; Onaitis, M.W.; Rawlins, E.L.; Lu, Y.; Clark, C.P.; Xue, Y.; Randell, S.H.; Hogan, B.L. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc. Natl. Acad. Sci. USA 2009, 106, 12771–12775. [Google Scholar] [CrossRef]
- Mehta, A.; Dobersch, S.; Dammann, R.H.; Bellusci, S.; Ilinskaya, O.N.; Braun, T.; Barreto, G. Validation of Tuba1a as appropriate internal control for normalization of gene expression analysis during mouse lung development. Int. J. Mol. Sci. 2015, 16, 4492–4511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkauskas, C.E.; Chung, M.I.; Fioret, B.; Gao, X.; Katsura, H.; Hogan, B.L. Lung organoids: Current uses and future promise. Development 2017, 144, 986–997. [Google Scholar] [CrossRef] [Green Version]
- Boers, J.E.; Ambergen, A.W.; Thunnissen, F.B. Number and proliferation of Clara cells in normal human airway epithelium. Am. J. Respir. Crit. Care. Med. 1999, 159, 1585–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrisey, E.E.; Hogan, B.L. Preparing for the first breath: Genetic and cellular mechanisms in lung development. Dev. Cell 2010, 18, 8–23. [Google Scholar] [CrossRef] [Green Version]
- Little, D.R.; Lynch, A.M.; Yan, Y.; Akiyama, H.; Kimura, S.; Chen, J. Differential chromatin binding of the lung lineage transcription factor NKX2-1 resolves opposing murine alveolar cell fates in vivo. Nat. Commun. 2021, 12, 2509. [Google Scholar] [CrossRef] [PubMed]
- Stuart, W.D.; Fink-Baldauf, I.M.; Tomoshige, K.; Guo, M.; Maeda, Y. CRISPRi-mediated functional analysis of NKX2-1-binding sites in the lung. Commun. Biol. 2021, 4, 568. [Google Scholar] [CrossRef] [PubMed]
- Billmyre, K.K.; Hutson, M.; Klingensmith, J. One shall become two: Separation of the esophagus and trachea from the common foregut tube. Dev. Dyn. 2015, 244, 277–288. [Google Scholar] [CrossRef] [Green Version]
- Perl, A.-K.T.; Wert, S.E.; Nagy, A.; Lobe, C.G.; Whitsett, J.A. Early restriction of peripheral and proximal cell lineages during formation of the lung. Proc. Natl. Acad. Sci. USA 2002, 99, 10482–10487. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.-Y.; Slack, J.M.W.; Tosh, D. Conversion of columnar to stratified squamous epithelium in the developing mouse oesophagus. Dev. Biol. 2005, 284, 157–170. [Google Scholar] [CrossRef]
- Lander, T.A.; Schauer, G.; Bendel-Stenzel, E.; Sidman, J.D. Tracheal agenesis in newborns. Laryngoscope 2004, 114, 1633–1636. [Google Scholar] [CrossRef]
- Kusakabe, T.; Kawaguchi, A.; Hoshi, N.; Kawaguchi, R.; Hoshi, S.; Kimura, S. Thyroid-specific enhancer-binding protein/NKX2.1 is required for the maintenance of ordered architecture and function of the differentiated thyroid. Mol. Endocrinol. 2006, 20, 1796–1809. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence (5′-3′) | Length (bp) | |
|---|---|---|---|
| SP-A | Forward | TTCCACCAATGGGCAGTCAG | 192 |
| Reverse | GAAGCCCCATCCAGGTAGTG | ||
| SP-B | Forward | GGCCTCACACTCAGGACTTC | 110 |
| Reverse | CAGGCACTTGGGGATCACG | ||
| SP-C | Forward | ATACTGAGATGGTCCTTGAGATG | 135 |
| Reverse | GCCGCTGGTAGTCATACAC | ||
| PDPN | Forward | AGAGAACACGAGAGTACAACCA | 99 |
| Reverse | CGTTTCATCCCCTGCATTATCT | ||
| LAMP3 | Forward | CATCACCAGCCAAGATCGGA | 155 |
| Reverse | AGATGCATGGGTTAGGCTGG | ||
| ABCA3 | Forward | GGGTGATGGACCCAACGAAT | 155 |
| Reverse | TGCCACCATCTTCCATTCCC | ||
| CC10 | Forward | GAGGCCCTCCTCATGGAATC | 130 |
| Reverse | TCCTGGTCTCTTGTGGGAGG | ||
| HOXA4 | Forward | GCTCTCGAACCGCCTATACC | 192 |
| Reverse | TCGCATCTTGGTGTTGGGAA | ||
| Trp63 | Forward | TGCGTCGGAGGAATGAAC | 160 |
| Reverse | ATACTTGCTGCTTTCTGATGC | ||
| CK5 | Forward | TCCAGTGTGTCCTTCCGAAGT | 223 |
| Reverse | TGCCTCCGCCAGAACTGTA | ||
| Gapdh | Forward | GCCTTCCGTGTTCCTACC | 101 |
| Reverse | GCCTGCTTCACCACCTTC | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, L.; Li, M.; Yin, Z.; Lv, L.; Zhou, M.; Wang, Y.; Zhang, M.; Guo, T.; Guo, X.; Liu, H.; et al. Development of a Lung Vacancy Mouse Model through CRISPR/Cas9-Mediated Deletion of Thyroid Transcription Factor 1 Exon 2. Cells 2022, 11, 3874. https://doi.org/10.3390/cells11233874
Zhao L, Li M, Yin Z, Lv L, Zhou M, Wang Y, Zhang M, Guo T, Guo X, Liu H, et al. Development of a Lung Vacancy Mouse Model through CRISPR/Cas9-Mediated Deletion of Thyroid Transcription Factor 1 Exon 2. Cells. 2022; 11(23):3874. https://doi.org/10.3390/cells11233874
Chicago/Turabian StyleZhao, Lihua, Meishuang Li, Zhibao Yin, Limin Lv, Meng Zhou, Yixi Wang, Manling Zhang, Tianxu Guo, Xiyun Guo, Han Liu, and et al. 2022. "Development of a Lung Vacancy Mouse Model through CRISPR/Cas9-Mediated Deletion of Thyroid Transcription Factor 1 Exon 2" Cells 11, no. 23: 3874. https://doi.org/10.3390/cells11233874
APA StyleZhao, L., Li, M., Yin, Z., Lv, L., Zhou, M., Wang, Y., Zhang, M., Guo, T., Guo, X., Liu, H., Cheng, L., Liang, X., Duo, S., & Li, R. (2022). Development of a Lung Vacancy Mouse Model through CRISPR/Cas9-Mediated Deletion of Thyroid Transcription Factor 1 Exon 2. Cells, 11(23), 3874. https://doi.org/10.3390/cells11233874