
Application of ALFA-Tagging in the Nematode Model Organisms Caenorhabditis elegans and Pristionchus pacificus

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials & Methods
2.1. Maintenance of Worm Cultures
2.2. Nomenclature of Tagged Genes and Proteins
2.3. CRISPR/Cas9 Mutagenesis
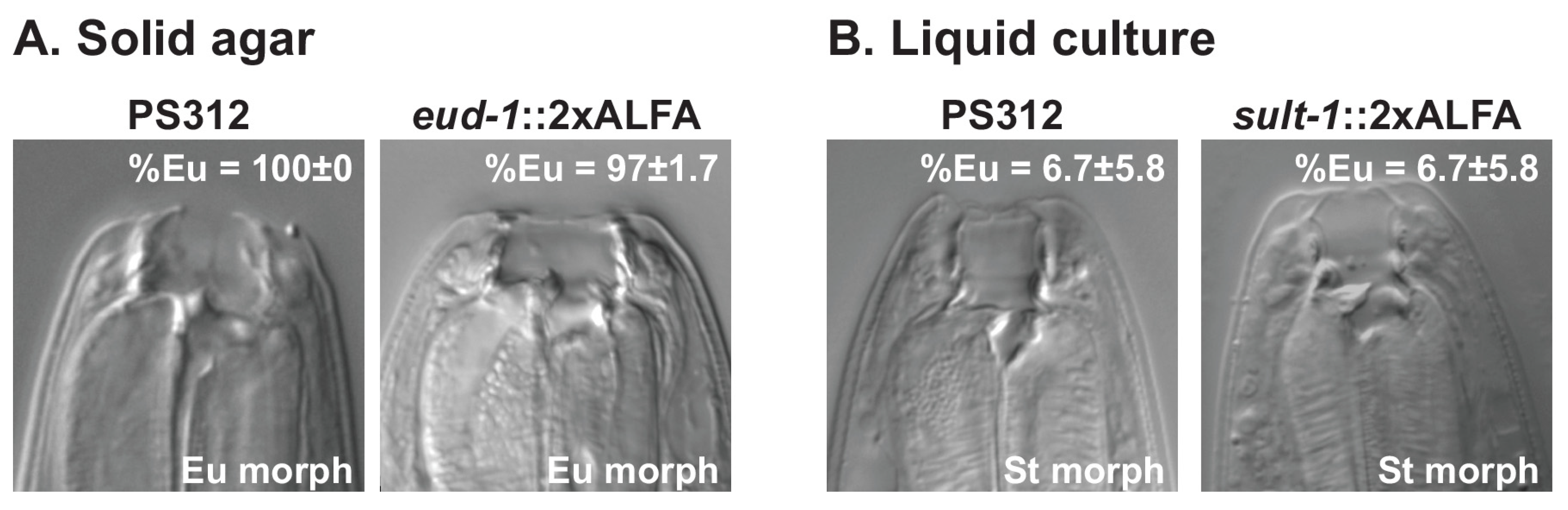
2.4. Mouth-Form Phenotyping in P. pacificus
2.5. ALFA-Tag Immunostaining
2.6. ALFA-Tag Immunoprecipitation
2.7. Western Blot
2.8. Mass Spectrometry Measurements
3. Results and Discussion
3.1. CRISPR-Based ALFA Tagging in C. Elegans and P. Pacificus
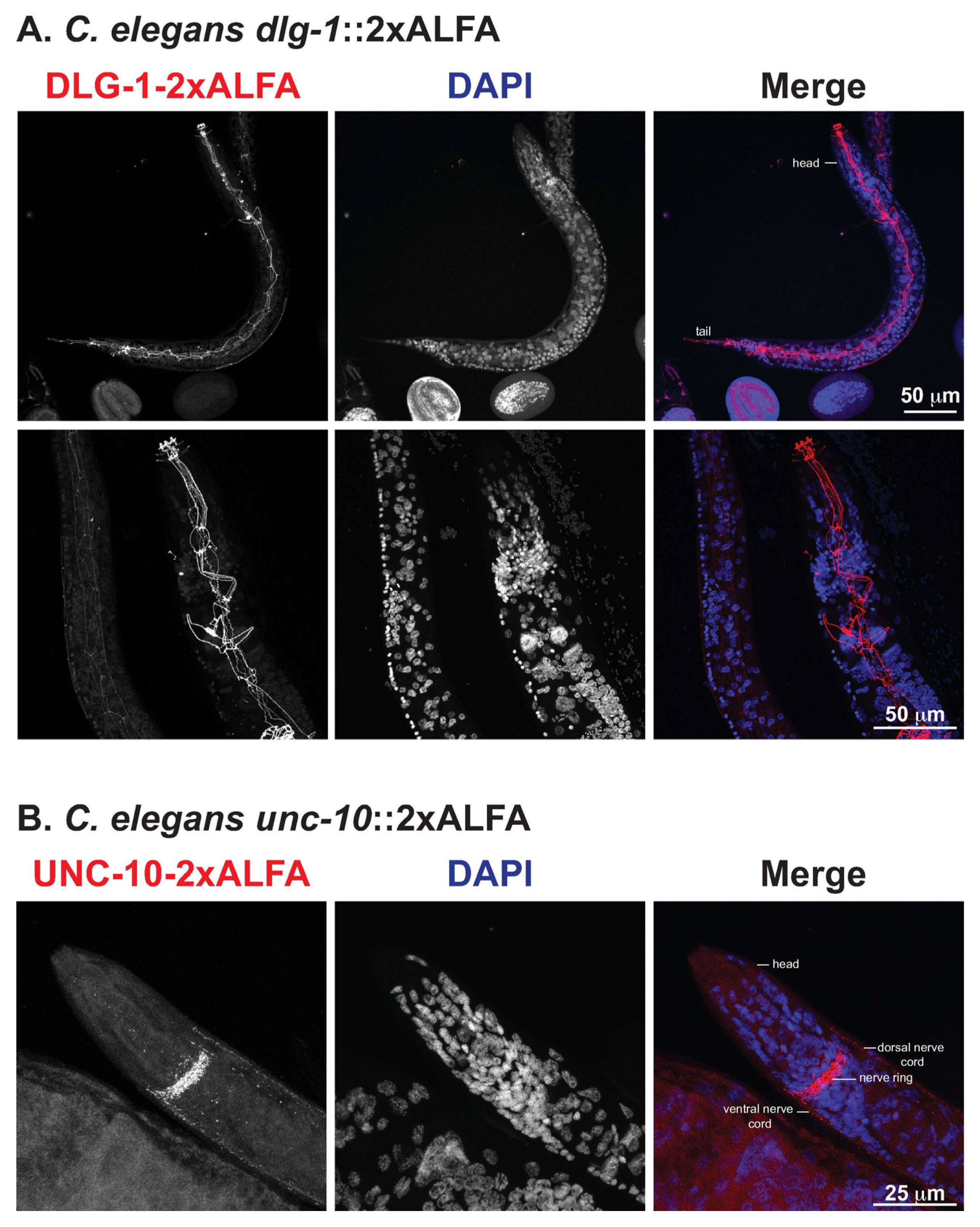
3.2. High-Resolution Immunofluorescence of ALFA-Tagged Proteins in C. Elegans
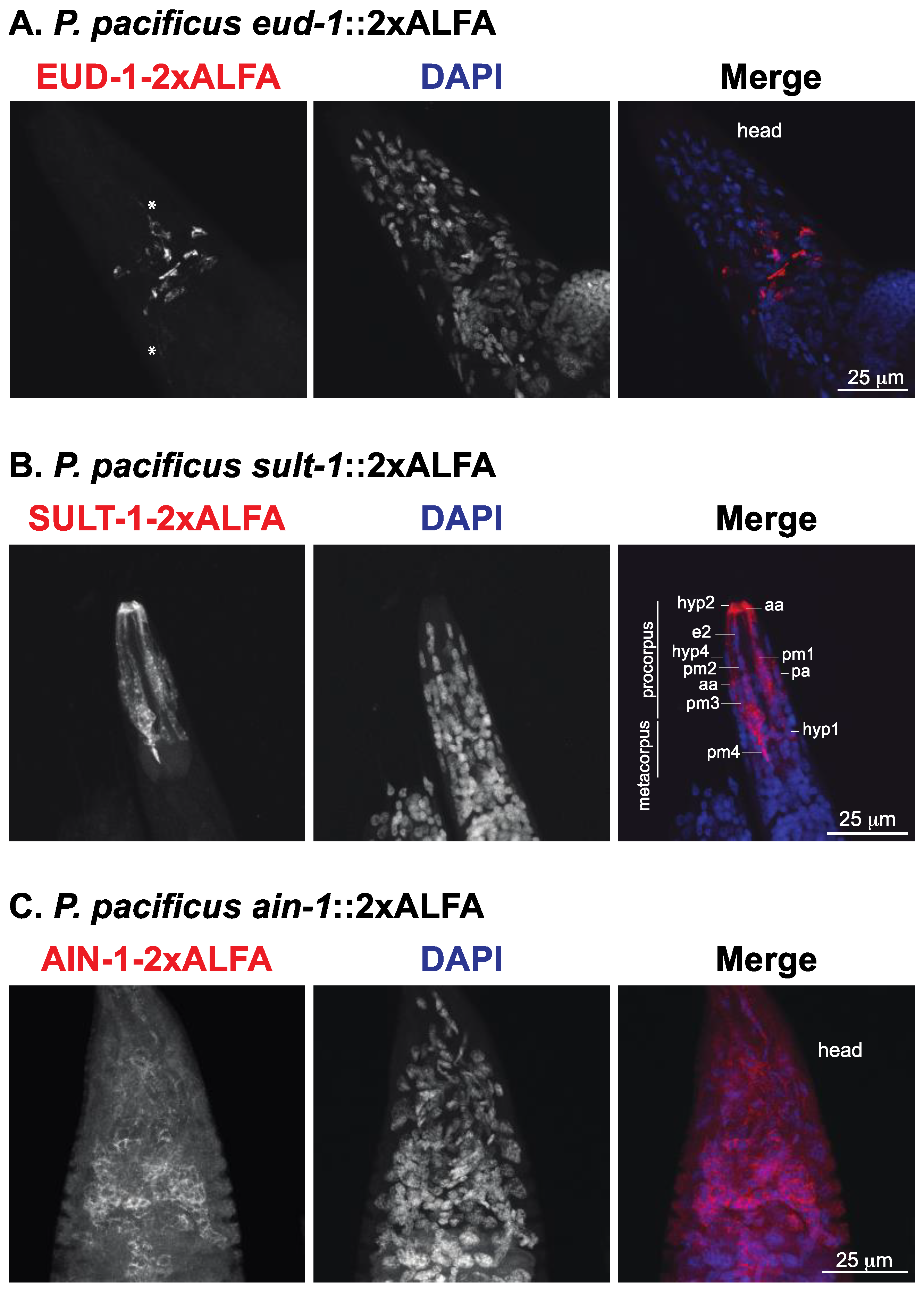
3.3. Visualization of ALFA-Tagged Proteins in P. pacificus Reveals Sub-Localization of Mouth-Form Regulators
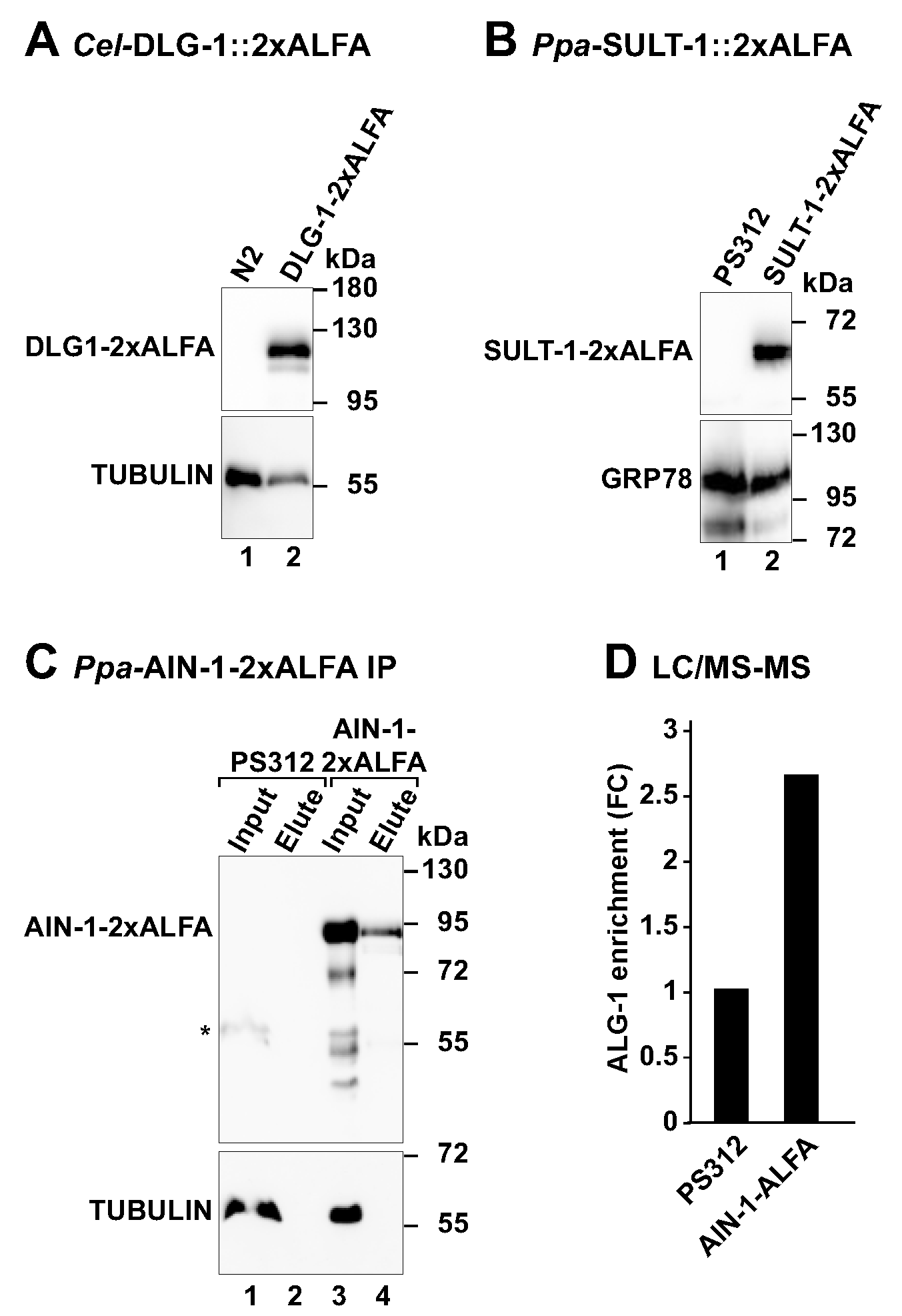
3.4. Detection of ALFA-Tagged Proteins by Western Blot
3.5. Capture of ALFA-Tagged Proteins from Worm Lysates
4. Limitations of the Current Study
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Brizzard, B. Epitope tagging. Biotechniques 2008, 44, 693–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandemoortele, G.; Eyckerman, S.; Gevaert, K. Pick a Tag and Explore the Functions of Your Pet Protein. Trends Biotechnol. 2019, 37, 1078–1090. [Google Scholar] [CrossRef] [PubMed]
- Gotzke, H.; Kilisch, M.; Martinez-Carranza, M.; Sograte-Idrissi, S.; Rajavel, A.; Schlichthaerle, T.; Engels, N.; Jungmann, R.; Stenmark, P.; Opazo, F.; et al. The ALFA-tag is a highly versatile tool for nanobody-based bioscience applications. Nat. Commun. 2019, 10, 4403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudolph, M.; Carsten, A.; Kulnik, S.; Aepfelbacher, M.; Wolters, M. Live imaging of Yersinia translocon formation and immune recognition in host cells. PLoS Pathog. 2022, 18, e1010251. [Google Scholar] [CrossRef] [PubMed]
- Akhuli, D.; Dhar, A.; Viji, A.S.; Bhojappa, B.; Palani, S. ALIBY: ALFA Nanobody-Based Toolkit for Imaging and Biochemistry in Yeast. mSphere 2022, e0033322. [Google Scholar] [CrossRef]
- van den Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.A.; de Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.S.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef] [Green Version]
- Glazer, I.; Shapiro-Ilan, D.; Sternberg, P.W. Nematodes as Model Organisms; CABI: Wallingford, UK, 2022; p. 376. [Google Scholar]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis Elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Brenner, S. The genetics of Caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Haag, E.S.; Fitch, D.H.A.; Delattre, M. From “the Worm” to “the Worms” and Back Again: The Evolutionary Developmental Biology of Nematodes. Genetics 2018, 210, 397–433. [Google Scholar] [CrossRef] [Green Version]
- Sommer, R.J.; Sternberg, P.W. Apoptosis and change of competence limit the size of the vulva equivalence group in Pristionchus pacificus: A genetic analysis. Curr. Biol. CB 1996, 6, 52–59. [Google Scholar] [CrossRef]
- Schroeder, N.E. Introduction to Pristionchus pacificus anatomy. J. Nematol. 2021, 53, e2021-91. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Giacomelli, M.; Lozano-Fernandez, J.; Edgecombe, G.D.; Fleming, J.F.; Kirstensen, R.M.; Ma, X.; Olesen, J.; Sorensen, M.V.; Thomsen, P.F.; et al. The Ediacaran origin of Ecdysozoa: Integrating fossil and phylogenomic data. J. Geol. Soc. 2022, 179. [Google Scholar] [CrossRef]
- Dickinson, D.J.; Goldstein, B. CRISPR-Based Methods for Caenorhabditis elegans Genome Engineering. Genetics 2016, 202, 885–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, K.I.; Ishita, Y.; Chihara, T.; Okumura, M. Screening for CRISPR/Cas9-induced mutations using a co-injection marker in the nematode Pristionchus pacificus. Dev. Genes Evol. 2020, 230, 257–264. [Google Scholar] [CrossRef]
- Han, Z.; Sieriebriennikov, B.; Susoy, V.; Lo, W.S.; Igreja, C.; Dong, C.; Berasategui, A.; Witte, H.; Sommer, R.J. Horizontally Acquired Cellulases Assist the Expansion of Dietary Range in Pristionchus Nematodes. Mol. Biol. Evol. 2022, 39, msab370. [Google Scholar] [CrossRef] [PubMed]
- Werner, M.S.; Sieriebriennikov, B.; Loschko, T.; Namdeo, S.; Lenuzzi, M.; Dardiry, M.; Renahan, T.; Sharma, D.R.; Sommer, R.J. Environmental influence on Pristionchus pacificus mouth form through different culture methods. Sci. Rep. 2017, 7, 7207. [Google Scholar] [CrossRef] [Green Version]
- Stiernagle, T. Maintenance of C. elegans. In Wormbook; Fay, D., Ed.; Oxford University Press: Oxford, UK, 2006. [Google Scholar]
- Han, Z.; Lo, W.S.; Lightfoot, J.W.; Witte, H.; Sun, S.; Sommer, R.J. Improving Transgenesis Efficiency and CRISPR-Associated Tools Through Codon Optimization and Native Intron Addition in Pristionchus Nematodes. Genetics 2020, 216, 947–956. [Google Scholar] [CrossRef]
- Sun, S.; Theska, T.; Witte, H.; Ragsdale, E.J.; Sommer, R.J. The oscillating Mucin-type protein DPY-6 has a conserved role in nematode mouth and cuticle formation. Genetics 2022, 220, iyab233. [Google Scholar] [CrossRef]
- Sangaletti, R.; Bianchi, L. A method for culturing embryonic C. elegans cells. J. Vis. Exp. 2013, 79, e50649. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Bossinger, O.; Klebes, A.; Segbert, C.; Theres, C.; Knust, E. Zonula adherens formation in Caenorhabditis elegans requires dlg-1, the homologue of the Drosophila gene discs large. Dev. Biol. 2001, 230, 29–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMahon, L.; Legouis, R.; Vonesch, J.L.; Labouesse, M. Assembly of C. elegans apical junctions involves positioning and compaction by LET-413 and protein aggregation by the MAGUK protein DLG-1. J. Cell Sci. 2001, 114 Pt 12, 2265–2277. [Google Scholar] [CrossRef] [PubMed]
- Koushika, S.P.; Richmond, J.E.; Hadwiger, G.; Weimer, R.M.; Jorgensen, E.M.; Nonet, M.L. A post-docking role for active zone protein Rim. Nat. Neurosci. 2001, 4, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Kushibiki, Y.; Suzuki, T.; Jin, Y.; Taru, H. RIMB-1/RIM-Binding Protein and UNC-10/RIM Redundantly Regulate Presynaptic Localization of the Voltage-Gated Calcium Channel in Caenorhabditis elegans. J. Neurosci. 2019, 39, 8617–8631. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.J.; Barry, N.C.; Wassie, A.T.; Sinha, A.; Bhattacharya, A.; Asano, S.; Zhang, C.; Chen, F.; Hobert, O.; Goodman, M.B.; et al. Expansion microscopy of C. elegans. Elife 2020, 9, e46249. [Google Scholar] [CrossRef] [PubMed]
- Ragsdale, E.J.; Muller, M.R.; Rodelsperger, C.; Sommer, R.J. A developmental switch coupled to the evolution of plasticity acts through a sulfatase. Cell 2013, 155, 922–933. [Google Scholar] [CrossRef] [Green Version]
- Bui, L.T.; Ivers, N.A.; Ragsdale, E.J. A sulfotransferase dosage-dependently regulates mouthpart polyphenism in the nematode Pristionchus pacificus. Nat. Commun. 2018, 9, 4119. [Google Scholar] [CrossRef] [Green Version]
- Namdeo, S.; Moreno, E.; Rodelsperger, C.; Baskaran, P.; Witte, H.; Sommer, R.J. Two independent sulfation processes regulate mouth-form plasticity in the nematode Pristionchus pacificus. Development 2018, 145, dev166272. [Google Scholar] [CrossRef] [Green Version]
- Igreja, C.; Sommer, R.J. The Role of Sulfation in Nematode Development and Phenotypic Plasticity. Front. Mol. Biosci. 2022, 9, 838148. [Google Scholar] [CrossRef]
- Ding, L.; Spencer, A.; Morita, K.; Han, M. The developmental timing regulator AIN-1 interacts with miRISCs and may target the argonaute protein ALG-1 to cytoplasmic P bodies in C. elegans. Mol. Cell 2005, 19, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Jonas, S.; Izaurralde, E. The role of disordered protein regions in the assembly of decapping complexes and RNP granules. Genes Dev. 2013, 27, 2628–2641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieriebriennikov, B.; Prabh, N.; Dardiry, M.; Witte, H.; Roseler, W.; Kieninger, M.R.; Rodelsperger, C.; Sommer, R.J. A Developmental Switch Generating Phenotypic Plasticity Is Part of a Conserved Multi-gene Locus. Cell Rep. 2018, 23, 2835–2843 e4. [Google Scholar] [CrossRef] [PubMed]
- Kieninger, M.R.; Ivers, N.A.; Rodelsperger, C.; Markov, G.V.; Sommer, R.J.; Ragsdale, E.J. The Nuclear Hormone Receptor NHR-40 Acts Downstream of the Sulfatase EUD-1 as Part of a Developmental Plasticity Switch in Pristionchus. Curr. Biol. CB 2016, 26, 2174–2179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serobyan, V.; Xiao, H.; Namdeo, S.; Rodelsperger, C.; Sieriebriennikov, B.; Witte, H.; Roseler, W.; Sommer, R.J. Chromatin remodelling and antisense-mediated up-regulation of the developmental switch gene eud-1 control predatory feeding plasticity. Nat. Commun. 2016, 7, 12337. [Google Scholar] [CrossRef]
- Schlager, B.; Wang, X.; Braach, G.; Sommer, R.J. Molecular cloning of a dominant roller mutant and establishment of DNA-mediated transformation in the nematode Pristionchus pacificus. Genesis 2009, 47, 300–304. [Google Scholar] [CrossRef]
- Witte, H.; Moreno, E.; Rodelsperger, C.; Kim, J.; Kim, J.S.; Streit, A.; Sommer, R.J. Gene inactivation using the CRISPR/Cas9 system in the nematode Pristionchus pacificus. Dev. Genes Evol. 2015, 225, 55–62. [Google Scholar] [CrossRef]
- Harry, C.J.; Messar, S.M.; Ragsdale, E.J. Comparative reconstruction of the predatory feeding structures of the polyphenic nematode Pristionchus pacificus. Evol. Dev. 2022, 24, 16–36. [Google Scholar] [CrossRef]
- de Beer, M.A.; Giepmans, B.N.G. Nanobody-Based Probes for Subcellular Protein Identification and Visualization. Front. Cell Neurosci. 2020, 14, 573278. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Source | Modified Gene/ID | Comment |
|---|---|---|---|
| Caenorhabditis elegans | |||
| N2 (wild type) | C. elegans Genetics Center | -- | -- |
| RS4102 | Sommer Lab | unc-10(tu1781[unc-10::2xALFA]) | C-terminally tagged Ce UNC-10 |
| RS4103 | dlg-1(tu1782[dlg-1::2xALFA]) | C-terminally tagged Ce DLG-1 | |
| Pristionchus pacificus | |||
| 312 (wild type) | Sommer Lab | -- | -- |
| RSC011 (wild type) | -- | -- | |
| RS4031 | eud-1(tu1729[eud-1::2xALFA]) | C-terminally tagged Ppa EUD-1 | |
| RS4068 | ain-1(tu1753[ain-1::2xALFA]) | C-terminally tagged Ppa AIN-1 | |
| RS4085 | sult-1(tu1760[sult-1::2xALFA]) | C-terminally tagged Ppa SULT-1 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Igreja, C.; Loschko, T.; Schäfer, A.; Sharma, R.; Quiobe, S.P.; Aloshy, E.; Witte, H.; Sommer, R.J. Application of ALFA-Tagging in the Nematode Model Organisms Caenorhabditis elegans and Pristionchus pacificus. Cells 2022, 11, 3875. https://doi.org/10.3390/cells11233875
Igreja C, Loschko T, Schäfer A, Sharma R, Quiobe SP, Aloshy E, Witte H, Sommer RJ. Application of ALFA-Tagging in the Nematode Model Organisms Caenorhabditis elegans and Pristionchus pacificus. Cells. 2022; 11(23):3875. https://doi.org/10.3390/cells11233875
Chicago/Turabian StyleIgreja, Catia, Tobias Loschko, Alejandra Schäfer, Radhika Sharma, Shiela Pearl Quiobe, Elbin Aloshy, Hanh Witte, and Ralf J. Sommer. 2022. "Application of ALFA-Tagging in the Nematode Model Organisms Caenorhabditis elegans and Pristionchus pacificus" Cells 11, no. 23: 3875. https://doi.org/10.3390/cells11233875
APA StyleIgreja, C., Loschko, T., Schäfer, A., Sharma, R., Quiobe, S. P., Aloshy, E., Witte, H., & Sommer, R. J. (2022). Application of ALFA-Tagging in the Nematode Model Organisms Caenorhabditis elegans and Pristionchus pacificus. Cells, 11(23), 3875. https://doi.org/10.3390/cells11233875