

Quantitative Targeted Absolute Proteomics for Better Characterization of an In Vitro Human Blood–Brain Barrier Model Derived from Hematopoietic Stem Cells

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. Isolation of Human Brain Microvessels (hBMVs)
2.3. Cell Culture of Human Brain-like Endothelial Cells (hBLECs)
2.4. Preparation of Whole-Cell Lysate and Crude Membrane Fraction
2.5. LC-MS/MS-Based qTAP
3. Results
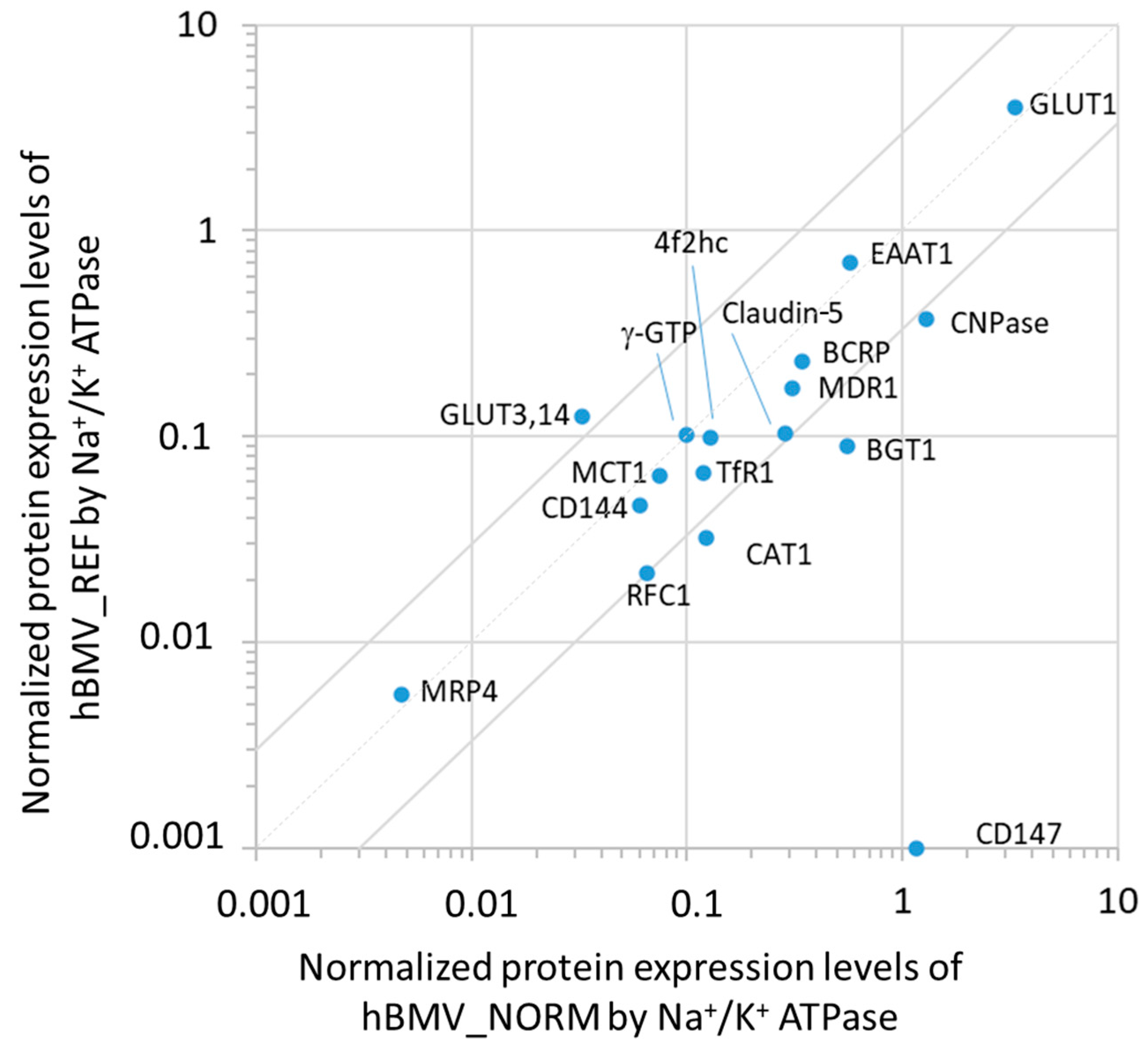
3.1. Analysis of the Protein Expression Levels in hBMVs and Comparison with the Previously Reported Quantification
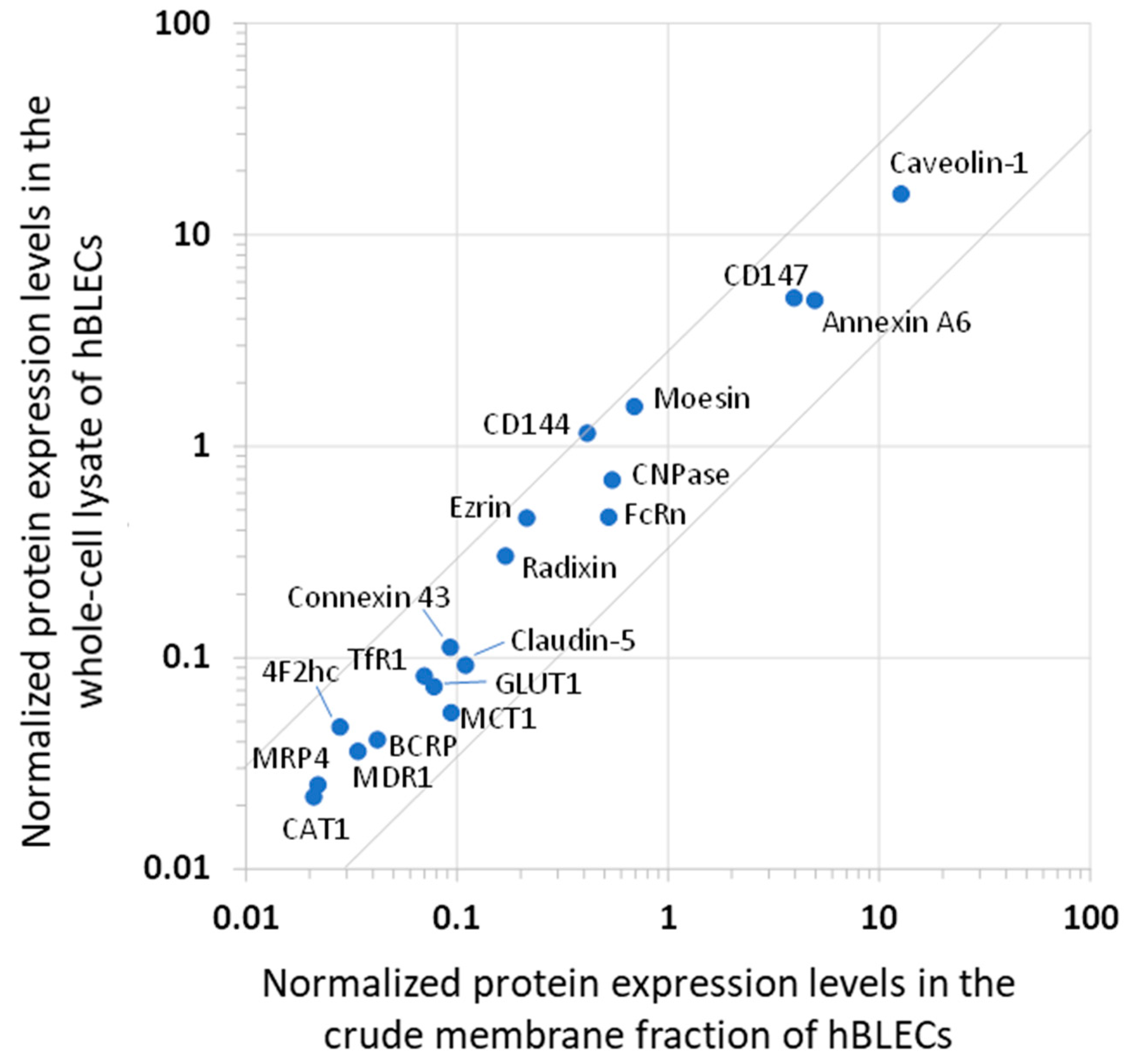
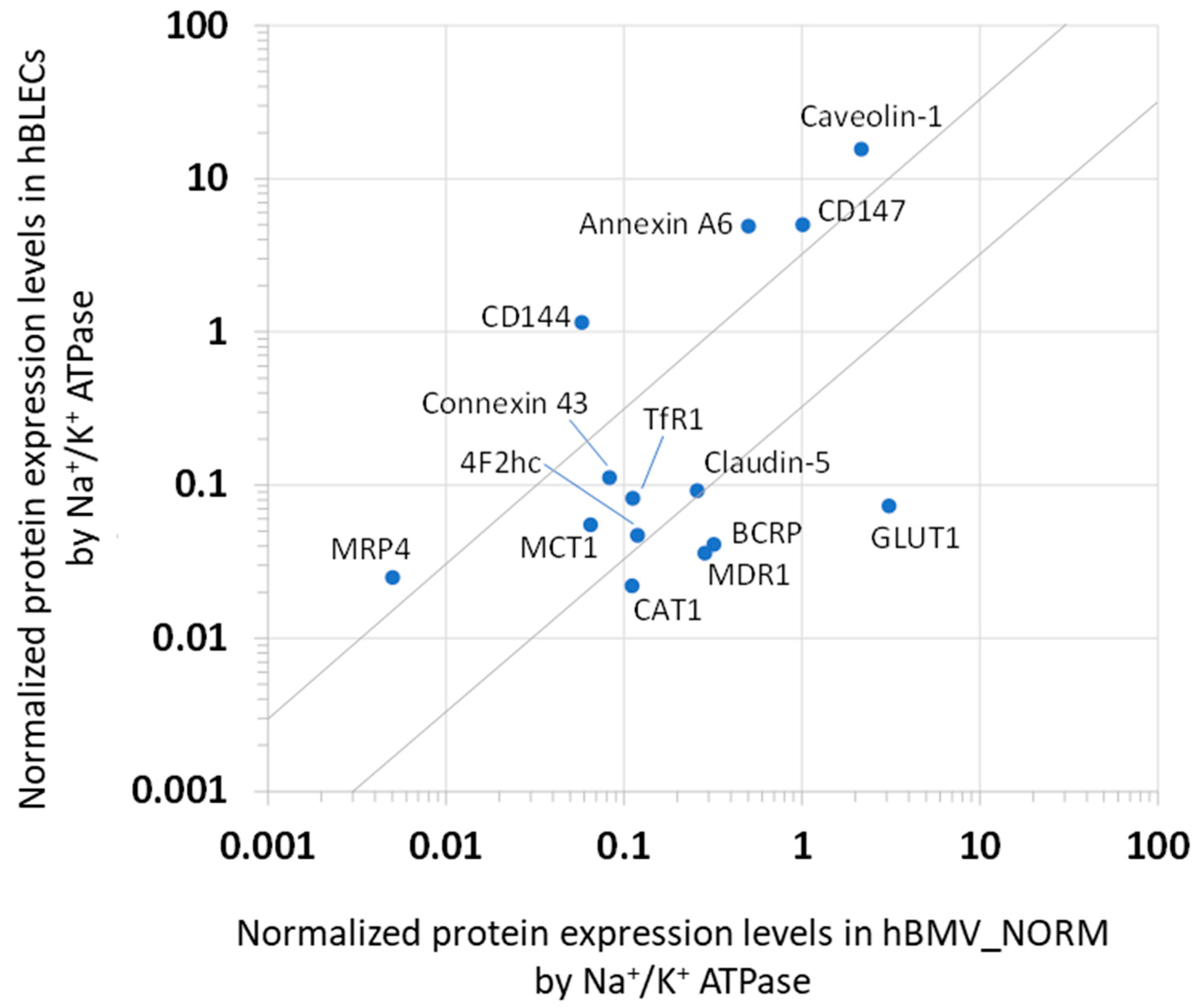
3.2. Analysis of the Protein Expression Levels of hBLECs and Comparison with hBMV_NORM
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tachikawa, M.; Uchida, Y.; Ohtsuki, S.; Terasaki, T. Recent progress in blood–brain barrier and blood–CSF barrier transport research: Pharmaceutical relevance for drug delivery to the brain. AAPS Adv. Pharm. Sci. Ser. 2014, 10, 23–62. [Google Scholar]
- Reese, T.S.; Karnovsky, M.J. Fine structural localization of a blood-brain barrier to exogenous peroxydase. J. Cell Biol. 1967, 34, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Menaceur, C.; Gosselet, F.; Fenart, L.; Saint-Pol, J. The Blood-Brain Barrier, an Evolving Concept Based on Technological Advances and Cell-Cell Communications. Cells 2021, 11, 133. [Google Scholar] [CrossRef] [PubMed]
- Knopp, R.C.; Banks, W.A.; Erickson, M.A. Physical associations of microglia and the vascular blood-brain barrier and their importance in development, health, and disease. Curr. Opin. Neurobiol. 2022, 77, 102648. [Google Scholar] [CrossRef] [PubMed]
- Cecchelli, R.; Berezowski, V.; Lundquist, S.; Culot, M.; Renftel, M.; Dehouck, M.-P.; Fenart, L. Modelling of the blood-brain barrier in drug discovery and development. Nat. Rev. Drug Discov. 2007, 6, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Syvänen, S.; Lindhe, Ö.; Palner, M.; Kornum, B.R.; Rahman, O.; Långström, B.; Knudsen, G.M.; Hammarlund-Udenaes, M. Species differences in blood-brain barrier transport of three positron emission tomography radioligands with emphasis on P-glycoprotein transport. Drug Metab. Dispos. 2009, 37, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Uchida, Y.; Ohtsuki, S.; Katsukura, Y.; Ikeda, C.; Suzuki, T.; Kamiie, J.; Terasaki, T. Quantitative targeted absolute proteomics of human blood-brain barrier transporters and receptors. J. Neurochem. 2011, 117, 333–345. [Google Scholar] [CrossRef]
- Hopkins, A.M.; DeSimone, E.; Chwalek, K.; Kaplan, D.L. 3D in vitro modeling of the central nervous system. Prog. Neurobiol. 2015, 125, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Weksler, B.B.; Subileau, E.A.; Perrière, N.; Charneau, P.; Holloway, K.; Leveque, M.; Tricoire-Leignel, H.; Nicotra, A.; Bourdoulous, S.; Turowski, P.; et al. Blood-brain barrier-specific properties of a human adult brain endothelial cell line. FASEB J. 2005, 19, 1872–1874. [Google Scholar] [CrossRef] [Green Version]
- Katt, M.E.; Linville, R.M.; Mayo, L.N.; Xu, Z.S.; Searson, P.C. Functional brain-specific microvessels from iPSC-derived human brain microvascular endothelial cells: The role of matrix composition on monolayer formation. Fluids Barriers CNS 2018, 15, 7. [Google Scholar] [CrossRef]
- Lippmann, E.S.; Azarin, S.M.; Kay, J.E.; Nessler, R.A.; Wilson, H.K.; Al-Ahmad, A.; Palecek, S.P.; Shusta, E.V. Derivation of blood-brain barrier endothelial cells from human pluripotent stem cells. Nat. Biotechnol. 2012, 30, 783–791. [Google Scholar] [CrossRef]
- Aday, S.; Cecchelli, R.; Hallier-Vanuxeem, D.; Dehouck, M.P.; Ferreira, L. Stem Cell-Based Human Blood–Brain Barrier Models for Drug Discovery and Delivery. Trends Biotechnol. 2016, 34, 382–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer-Di Ponio, J.; El-Ayoubi, F.; Glacial, F.; Ganeshamoorthy, K.; Driancourt, C.; Godet, M.; Perrière, N.; Guillevic, O.; Olivier Couraud, P.; Uzan, G. Instruction of circulating endothelial progenitors in vitro towards specialized blood-brain barrier and arterial phenotypes. PLoS ONE 2014, 9, e84179. [Google Scholar]
- Cecchelli, R.; Aday, S.; Sevin, E.; Almeida, C.; Culot, M.; Dehouck, L.; Coisne, C.; Engelhardt, B.; Dehouck, M.-P.; Ferreira, L. A stable and reproducible human blood-brain barrier model derived from hematopoietic stem cells. PLoS ONE 2014, 9, e99733. [Google Scholar] [CrossRef] [Green Version]
- Ravid, O.; Elhaik Goldman, S.; Macheto, D.; Bresler, Y.; De Oliveira, R.I.; Liraz-Zaltsman, S.; Gosselet, F.; Dehouck, L.; Beeri, M.S.; Cooper, I. Blood-brain barrier cellular responses toward organophosphates: Natural compensatory processes and exogenous interventions to rescue barrier properties. Front. Cell. Neurosci. 2018, 12, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deligne, C.; Hachani, J.; Duban-Deweer, S.; Meignan, S.; Leblond, P.; Carcaboso, A.M.; Sano, Y.; Shimizu, F.; Kanda, T.; Gosselet, F.; et al. Development of a human in vitro blood-brain tumor barrier model of diffuse intrinsic pontine glioma to better understand the chemoresistance. Fluids Barriers CNS 2020, 17, 37. [Google Scholar] [CrossRef]
- Moya, E.L.J.; Vandenhaute, E.; Rizzi, E.; Boucau, M.C.; Hachani, J.; Maubon, N.; Gosselet, F.; Dehouck, M.P. Miniaturization and Automation of a Human In Vitro Blood-Brain Barrier Model for the High-Throughput Screening of Compounds in the Early Stage of Drug Discovery. Pharmaceutics 2021, 13, 892. [Google Scholar] [CrossRef]
- Versele, R.; Corsi, M.; Fuso, A.; Sevin, E.; Businaro, R.; Gosselet, F.; Fenart, L.; Candela, P. Ketone Bodies Promote Amyloid-β1–40 Clearance in a Human in Vitro Blood–Brain Barrier Model. Int. J. Mol. Sci. 2020, 21, 934. [Google Scholar] [CrossRef] [Green Version]
- Drolez, A.; Vandenhaute, E.; Delannoy, C.P.; Dewald, J.H.; Gosselet, F.; Cecchelli, R.; Julien, S.; Dehouck, M.P.; Delannoy, P.; Mysiorek, C. ST6GALNAC5 expression decreases the interactions between breast cancer cells and the human blood-brain barrier. Int. J. Mol. Sci. 2016, 17, 1309. [Google Scholar] [CrossRef] [Green Version]
- Mossu, A.; Rosito, M.; Khire, T.; Li Chung, H.; Nishihara, H.; Gruber, I.; Luke, E.; Dehouck, L.; Sallusto, F.; Gosselet, F.; et al. A silicon nanomembrane platform for the visualization of immune cell trafficking across the human blood–brain barrier under flow. J. Cereb. Blood Flow Metab. 2019, 39, 395–410. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, Y.; Uchida, Y.; Tachikawa, M.; Inoue, T.; Ohtsuki, S.; Terasaki, T. Quantitative atlas of blood-brain barrier transporters, receptors, and tight junction proteins in rats and common marmoset. J. Pharm. Sci. 2013, 102, 3343–3355. [Google Scholar] [CrossRef] [PubMed]
- Ohtsuki, S.; Ikeda, C.; Uchida, Y.; Sakamoto, Y.; Miller, F.; Glacial, F.; Decleves, X.; Scherrmann, J.-M.; Couraud, P.-O.; Kubo, Y.; et al. Quantitative targeted absolute proteomic analysis of transporters, receptors and junction proteins for validation of human cerebral microvascular endothelial cell line hCMEC/D3 as a human blood-brain barrier model. Mol. Pharm. 2013, 10, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Weksler, B.; Romero, I.A.; Couraud, P.O. The hCMEC/D3 cell line as a model of the human blood brain barrier. Fluids Barriers CNS 2013, 10, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedroso, D.C.S.; Tellechea, A.; Moura, L.; Fidalgo-Carvalho, I.; Duarte, J.; Carvalho, E.; Ferreira, L. Improved survival, vascular differentiation and wound healing potential of stem cells co-cultured with endothelial cells. PLoS ONE 2011, 6, e16114. [Google Scholar] [CrossRef] [Green Version]
- Kamiie, J.; Ohtsuki, S.; Iwase, R.; Ohmine, K.; Katsukura, Y.; Yanai, K.; Sekine, Y.; Uchida, Y.; Ito, S.; Terasaki, T. Quantitative atlas of membrane transporter proteins: Development and application of a highly sensitive simultaneous LC/MS/MS method combined with novel in-silico peptide selection criteria. Pharm. Res. 2008, 25, 1469–1483. [Google Scholar] [CrossRef]
- Uchida, Y.; Zhang, Z.; Tachikawa, M.; Terasaki, T. Quantitative targeted absolute proteomics of rat blood-cerebrospinal fluid barrier transporters: Comparison with a human specimen. J. Neurochem. 2015, 134, 1104–1115. [Google Scholar] [CrossRef]
- Uchida, Y.; Yagi, Y.; Takao, M.; Takao, M.; Tano, M.; Umetsu, M.; Hirano, S.; Usui, T.; Tachikawa, M.; Terasaki, T. Comparison of Absolute Protein Abundances of Transporters and Receptors among Blood-Brain Barriers at Different Cerebral Regions and the Blood-Spinal Cord Barrier in Humans and Rats. Mol. Pharm. 2020, 17, 2006–2020. [Google Scholar] [CrossRef]
- Cornford, E.M.; Hyman, S. Localization of brain endothelial luminal and abluminal transporters with immunogold electron microscopy. NeuroRx 2005, 2, 27–43. [Google Scholar] [CrossRef]
- Nguyen, Y.T.K.; Ha, H.T.T.; Nguyen, T.H.; Nguyen, L.N. The role of SLC transporters for brain health and disease. Cell. Mol. Life Sci. 2021, 79, 20. [Google Scholar] [CrossRef]
- Wang, H.; Lv, J.J.; Zhao, Y.; Wei, H.L.; Zhang, T.J.; Yang, H.J.; Chen, Z.N.; Jiang, J.L. Endothelial genetic deletion of CD147 induces changes in the dual function of the blood-brain barrier and is implicated in Alzheimer’s disease. CNS Neurosci. Ther. 2021, 27, 1048–1063. [Google Scholar] [CrossRef]
- Orlowski, M.; Sessa, G.; Green, J.P. γ-Glutamyl transpeptidase in brain capillaries: Possible site of a blood-brain barrier for amino acids. Science 1974, 184, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Risau, W.; Dingler, A.; Albrecht, U.; Dehouck, M.-P.; Cecchelli, R. Blood–Brain Barrier Pericytes Are the Main Source of γ-Glutamyltranspeptidase Activity in Brain Capillaries. J. Neurochem. 1992, 58, 667–672. [Google Scholar] [CrossRef] [PubMed]
- Vandenhaute, E.; Dehouck, L.; Boucau, M.-C.; Sevin, E.; Uzbekov, R.; Tardivel, M.; Gosselet, F.; Fenart, L.; Cecchelli, R.; Dehouck, M.-P. Modelling the neurovascular unit and the blood-brain barrier with the unique function of pericytes. Curr. Neurovasc. Res. 2011, 4, 258–269. [Google Scholar] [CrossRef]
- Kickinger, S.; Hellsberg, E.; Frølund, B.; Schousboe, A.; Ecker, G.F.; Wellendorph, P. Structural and molecular aspects of betaine-GABA transporter 1 (BGT1) and its relation to brain function. Neuropharmacology 2019, 161, 107644. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Gravel, M.; Zhang, R.; Thibault, P.; Braun, P.E. Process outgrowth in oligodendrocytes is mediated by CNP, a novel microtubule assembly myelin protein. J. Cell Biol. 2005, 170, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Parkin, G.M.; Udawela, M.; Gibbons, A.; Dean, B. Glutamate transporters, EAAT1 and EAAT2, are potentially important in the pathophysiology and treatment of schizophrenia and affective disorders. World J. Psychiatry 2018, 8, 51–63. [Google Scholar] [CrossRef]
- Friedrich, R.E.; Holstein, A.-F.; Middendorff, R.; Davidoff, M.S. Vascular wall cells contribute to tumourigenesis in cutaneous neurofibromas of patients with neurofibromatosis type 1. A comparative histological, ultrastructural and immunohistochemical study. Anticancer Res. 2012, 5, 2139–2158. [Google Scholar]
- Mahringer, A.; Fricker, G. ABC transporters at the blood-brain barrier. Expert Opin. Drug Metab. Toxicol. 2016, 12, 499–508. [Google Scholar] [CrossRef]
- Bendayan, R.; Ronaldson, P.T.; Gingras, D.; Bendayan, M. In situ localization of P-glycoprotein (ABCB1) in human and rat brain. J. Histochem. Cytochem. 2006, 54, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Van Vliet, E.A.; Iyer, A.M.; Mesarosova, L.; Çolakoglu, H.; Anink, J.J.; Van Tellingen, O.; Maragakis, N.J.; Shefner, J.; Bunt, T.; Aronica, E. Expression and Cellular Distribution of P-Glycoprotein and Breast Cancer Resistance Protein in Amyotrophic Lateral Sclerosis Patients. J. Neuropathol. Exp. Neurol. 2020, 79, 266–276. [Google Scholar] [CrossRef] [Green Version]
- Kuntz, M.; Candela, P.; Saint-Pol, J.; Lamartinière, Y.; Boucau, M.C.; Sevin, E.; Fenart, L.; Gosselet, F. Bexarotene Promotes Cholesterol Efflux and Restricts Apical-to-Basolateral Transport of Amyloid-β Peptides in an in Vitro Model of the Human Blood-Brain Barrier. J. Alzheimer’s Dis. 2015, 48, 849–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pragallapati, S.; Manyam, R. Glucose transporter 1 in health and disease. J. Oral Maxillofac. Pathol. 2019, 23, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, A.K. Glucose transport in brain and retina: Implications in the management and complications of diabetes. Diabetes Metab. Res. Rev. 1999, 15, 261–273. [Google Scholar] [CrossRef]
- Zhao, Y.; Xin, Y.; He, Z.; Hu, W. Function of connexins in the interaction between glial and vascular cells in the central nervous system and related neurological diseases. Neural Plast. 2018, 2018, 6323901. [Google Scholar] [CrossRef]
- Loureiro, J.A.; Andrade, S.; Duarte, A.; Neves, A.R.; Queiroz, J.F.; Nunes, C.; Sevin, E.; Fenart, L.; Gosselet, F.; Coelho, M.A.N.; et al. Resveratrol and grape extract-loaded solid lipid nanoparticles for the treatment of Alzheimer’s disease. Molecules 2017, 22, 277. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, M.J.; Sevin, E.; Gosselet, F.; Lima, J.; Coelho, M.A.N.; Loureiro, J.A.; Pereira, M.C. Receptor-mediated PLGA nanoparticles for glioblastoma multiforme treatment. Int. J. Pharm. 2018, 545, 84–92. [Google Scholar] [CrossRef]
- Li, W.; Chen, Z.; Chin, I.; Chen, Z.; Dai, H. The Role of VE-cadherin in Blood-brain Barrier Integrity Under Central Nervous System Pathological Conditions. Curr. Neuropharmacol. 2018, 16, 1375–1384. [Google Scholar] [CrossRef]
- Kawaguchi, K.; Yoshida, S.; Hatano, R.; Asano, S. Pathophysiological roles of ezrin/radixin/moesin proteins. Biol. Pharm. Bull. 2017, 40, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Pottiez, G.; Duban-Deweer, S.; Deracinois, B.; Gosselet, F.; Camoin, L.; Hachani, J.; Couraud, P.-O.; Cecchelli, R.; Dehouck, M.-P.; Fenart, L.; et al. A differential proteomic approach identifies structural and functional components that contribute to the differentiation of brain capillary endothelial cells. J. Proteom. 2011, 75, 628–641. [Google Scholar] [CrossRef]
- Kirk, P.; Wilson, M.C.; Heddle, C.; Brown, M.H.; Barclay, A.N.; Halestrap, A.P. CD147 is tightly associated with lactate transporters MCT1 and MCT4 and facilitates their cell surface expression. EMBO J. 2000, 19, 3896–3904. [Google Scholar] [CrossRef]
- Cubells, L.; Vilà De Muga, S.; Tebar, F.; Wood, P.; Evans, R.; Ingelmo-Torres, M.; Calvo, M.; Gaus, K.; Pol, A.; Grewal, T.; et al. Annexin a6-Induced alterations in cholesterol transport and caveolin export from the Golgi complex. Traffic 2007, 8, 1568–1589. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, Y.; Uchida, Y.; Tachikawa, M.; Ohtsuki, S.; Couraud, P.O.; Suzuki, T.; Terasaki, T. Oxidative stress-induced activation of Abl and Src kinases rapidly induces P-glycoprotein internalization via phosphorylation of caveolin-1 on tyrosine-14, decreasing cortisol efflux at the blood–brain barrier. J. Cereb. Blood Flow Metab. 2020, 40, 420–436. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, Y.; Uchida, Y.; Kuroda, T.; Tachikawa, M.; Couraud, P.O.; Suzuki, T.; Terasaki, T. Distinct roles of ezrin, radixin and moesin in maintaining the plasma membrane localizations and functions of human blood–brain barrier transporters. J. Cereb. Blood Flow Metab. 2020, 40, 1533–1545. [Google Scholar] [CrossRef] [PubMed]
- Omori, K.; Tachikawa, M.; Hirose, S.; Taii, A.; Akanuma, S.-i.; Hosoya, K.-i.; Terasaki, T. Developmental changes in transporter and receptor protein expression levels at the rat blood-brain barrier based on quantitative targeted absolute proteomics. Drug Metab. Pharmacokinet. 2020, 35, 117–123. [Google Scholar] [CrossRef]
- Schlachetzki, F.; Zhu, C.; Pardridge, W.M. Expression of the neonatal Fc receptor (FcRn) at the blood-brain barrier. J. Neurochem. 2002, 81, 203–206. [Google Scholar] [CrossRef]
- Roopenian, D.C.; Akilesh, S. FcRn: The neonatal Fc receptor comes of age. Nat. Rev. Immunol. 2007, 7, 715–725. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Protein Expression Levels | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Protein Name/Gene Name | hBLEC Crude Membrane Fraction | hBLEC Whole-Cell Lysate | hBMV_NORM Whole-Cell Lysate | hBMV_PATHO Whole-Cell Lysate | hBMV_REF Whole-Cell Lysate | |||||
| Value a (fmol/μg Protein) | Normalized b | Quadratic Mean c (fmol/μg Protein) | Normalized b | Quadratic Mean d (fmol/μg Protein) | Normalized b | Quadratic Mean d (fmol/μg Protein) | Normalized b | Value e (fmol/μg Protein) | Normalized b | |
| Transporters or Receptors | ||||||||||
| MDR1/ABCB1 | 0.536 ± 0.024 | 0.034 | 0.221 ± 0.065 | 0.036 | 7.25 ± 2.57 | 0.285 | 5.44 ± 2.23 | 0.380 | 6.06 ± 1.69 | 0.173 |
| MRP4/ABCC4 | 0.324 ± 0.015 | 0.022 | 0.152 ± 0.037 | 0.025 | 0.118 ± 0.001 | 0.005 | 0.072 ± 0.087 | 0.005 | 0.195 ± 0.069 | 0.006 |
| BCRP/ABCG2 | 0.659 ± 0.057 | 0.042 | 0.251 ± 0.014 | 0.041 | 8.15 ± 2.45 | 0.320 | 5.44 ± 2.41 | 0.379 | 8.14 ± 2.26 | 0.232 |
| EAAT1/SLC1A3 | U.L.Q. (<0.173) | - | U.L.Q. (<0.114) | - | 14.2 ± 1.1 | 0.559 | 10.4 ± 2.2 | 0.728 | 24.5 ± 12.5 | 0.698 |
| GLUT1/SLC2A1 | 1.22 ± 0.11 | 0.078 | 0.443 ± 0.068 | 0.073 | 78.6 ± 23.5 | 3.09 | 55.2 ± 21.2 | 3.85 | 139 ± 46 | 3.960 |
| BGT1/SLC6A12 | U.L.Q. (<1.320) | - | U.L.Q. (< 0.989) | - | 13.3 ± 2.3 | 0.523 | 6.73 ± 1.42 | 0.470 | 3.16 ± 0.94 | 0.090 |
| GLUT3,14/SLC2A3,14 | 0.234 * | 0.015 | N.D. | - | 0.800 ± 0.048 | 0.031 | 1.06 ± 0.49 | 0.074 | 4.40 ± 1.00 | 0.125 |
| CAT1/SLC7A1 | 0.329 * | 0.021 | 0.136 ± 0.041 | 0.022 | 2.84 ± 1.73 | 0.111 | 3.44 ± 2.23 | 0.240 | 1.13 ± 0.18 | 0.032 |
| MCT1/SLC16A1 | 1.46 ± 0.15 | 0.094 | 0.335 ± 0.043 | 0.055 | 1.65 ± 1.61 | 0.065 | 3.77 ± 1.66 | 0.263 | 2.27 ± 0.85 | 0.065 |
| RFC1/SLC19A1 | 0.248 ** | 0.016 | N.D. | - | 1.61 ± 0.12 | 0.063 | 0.515 ± 0.25 | 0.036 | 0.763 ± 0.041 | 0.022 |
| TfR1/TFRC | 1.17 ± 0.09 | 0.075 | 0.500 ± 0.146 | 0.082 | 2.86 ± 0.85 | 0.112 | 2.05 ± 0.35 | 0.143 | 2.34 ± 0.76 | 0.067 |
| FcRn/FCGRT | 8.16 ± 0.47 | 0.523 | 2.82 ± 0.35 | 0.463 | N.D. | - | N.D. | - | N.D. | - |
| Others | ||||||||||
| Claudin-5/CLDN5 | 1.72 ± 0.14 | 0.110 | 0.562 ± 0.043 | 0.092 | 6.59 ± 3.49 | 0.258 | 4.05 ± 1.09 | 0.283 | 3.62 ± 0.96 | 0.103 |
| CD144/CDH5 | 6.47 ± 0.20 | 0.415 | 7.02 ± 1.00 | 1.15 | 1.48 ± 0.17 | 0.058 | 1.25 ± 0.58 | 0.087 | 1.61 ± 0.43 | 0.046 |
| 4F2hc/SLC3A2 | 0.438 ± 0.020 | 0.028 | 0.288 ± 0.040 | 0.047 | 3.0 ± 0.89 | 0.119 | 2.01 ± 1.24 | 0.140 | 3.47 ± 0.83 | 0.099 |
| CD147/BSG | 61.8 ± 2.2 | 3.96 | 30.6 ± 2.5 | 5.02 | 25.6 ± 22.8 | 1.01 | 66.4 ± 11.5 | 4.64 | U.L.Q. (<0.083) | - |
| Caveolin-1/CAV1 | 199 ± 2 | 12.7 | 94.8 ± 8.7 | 15.6 | 55.0 ± 20.4 | 2.16 | 40.1 ± 7.8 | 2.79 | N.D. | - |
| Connexin 43/CX43 | 1.46 ** | 0.093 | 0.682 ±0.099 | 0.112 | 2.12 ± 0.67 | 0.083 | 1.74 ± 0.15 | 0.121 | N.D. | - |
| Annexin A6/ANXA6 | 77.4 ± 1.9 | 4.96 | 29.8 ± 4.57 | 4.90 | 12.8 ± 5.4 | 0.502 | 6.34 ± 0.99 | 0.442 | N.D. | - |
| Ezrin/EZR | 3.34 ± 0.21 | 0.214 | 2.79 ± 0.65 | 0.458 | N.D. | - | N.D. | - | N.D. | - |
| Radixin/RDX | 2.65 ± 0.73 | 0.170 | 1.84 ± 0.53 | 0.302 | N.D. | - | N.D. | - | N.D. | - |
| Moesin/MSN | 10.8 ± 0.3 | 0.692 | 9.36 ± 1.87 | 1.54 | N.D. | - | N.D. | - | N.D. | - |
| Markers proteins | ||||||||||
| γ-GTP/GGT | U.L.Q. (<0.127) | - | U.L.Q. (<0.098) | - | 2.37 ± 0.79 | 0.093 | 1.80 ± 0.19 | 0.1126 | 3.57 ± 0.71 | 0.102- |
| GFAP/GFAP | U.L.Q. (<0.095) | - | U.L.Q. (<0.080) | - | 5.38 ± 2.72 | 0.211 | 11.2 ± 0.2 | 0.781 | N.D. | - |
| CNPase/CNP | 8.49 ± 0.46 | 0.544 | 4.22 ± 0.59 | 0.693 | 36.9 ± 41.1 | 1.45 | 18.1 ± 1.9 | 1.26 | 12.96 ± 4.12 | 0.369 |
| Na+/K+ ATPase | 15.6 ± 1.2 | 1 | 6.09 ± 0.83 | 1 | 25.5 ± 6.4 | 1 | 14.3 ± 14.8 | 1 | 35.1 ± 12.6 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dehouck, M.-P.; Tachikawa, M.; Hoshi, Y.; Omori, K.; Maurage, C.-A.; Strecker, G.; Dehouck, L.; Boucau, M.-C.; Uchida, Y.; Gosselet, F.; et al. Quantitative Targeted Absolute Proteomics for Better Characterization of an In Vitro Human Blood–Brain Barrier Model Derived from Hematopoietic Stem Cells. Cells 2022, 11, 3963. https://doi.org/10.3390/cells11243963
Dehouck M-P, Tachikawa M, Hoshi Y, Omori K, Maurage C-A, Strecker G, Dehouck L, Boucau M-C, Uchida Y, Gosselet F, et al. Quantitative Targeted Absolute Proteomics for Better Characterization of an In Vitro Human Blood–Brain Barrier Model Derived from Hematopoietic Stem Cells. Cells. 2022; 11(24):3963. https://doi.org/10.3390/cells11243963
Chicago/Turabian StyleDehouck, Marie-Pierre, Masanori Tachikawa, Yutaro Hoshi, Kotaro Omori, Claude-Alain Maurage, Guillaume Strecker, Lucie Dehouck, Marie-Christine Boucau, Yasuo Uchida, Fabien Gosselet, and et al. 2022. "Quantitative Targeted Absolute Proteomics for Better Characterization of an In Vitro Human Blood–Brain Barrier Model Derived from Hematopoietic Stem Cells" Cells 11, no. 24: 3963. https://doi.org/10.3390/cells11243963
APA StyleDehouck, M. -P., Tachikawa, M., Hoshi, Y., Omori, K., Maurage, C. -A., Strecker, G., Dehouck, L., Boucau, M. -C., Uchida, Y., Gosselet, F., Terasaki, T., & Karamanos, Y. (2022). Quantitative Targeted Absolute Proteomics for Better Characterization of an In Vitro Human Blood–Brain Barrier Model Derived from Hematopoietic Stem Cells. Cells, 11(24), 3963. https://doi.org/10.3390/cells11243963