

Therapeutic Potential of Mesenchymal Stem Cells in the Treatment of Epilepsy and Their Interaction with Antiseizure Medications

 ,
, 

Abstract
:1. Introduction
2. MSCs
2.1. Umbilical Cord-Derived Mesenchymal Stem Cells (UC-MSCs)
2.2. Bone Marrow Mesenchymal Stem Cells (BMSCs)
2.3. Adipose-Derived Stem Cells (ADSCs)
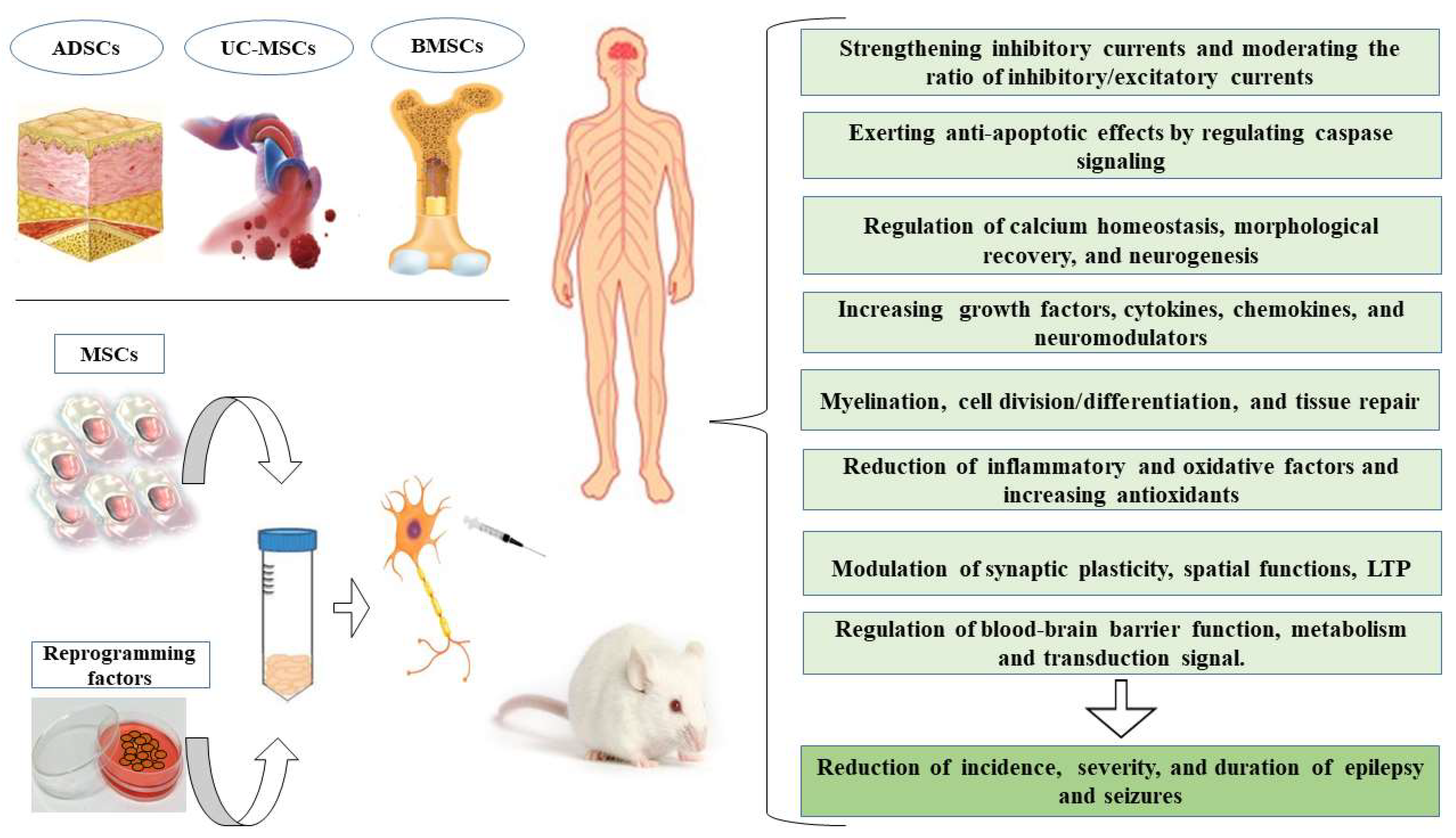
3. Mechanisms for MSCs Enhanced Neuroprotection
4. Possible Mechanisms of Anti-Seizure Effects of MSCs-Based Therapy
5. The Effect of ASMs on MSCs
5.1. The Effect of Valproic Acid (VPA) on MSCs

{kind=link}
| Drugs | The Source of MSCs | Genes and Signaling Pathways Involved | Ref. |
|---|---|---|---|
| Valproic acid | Cryopreserved rat MSCs | Increased cell migration ability | [110] |
| Human umbilical cord-derived MSCs | Enhancement of cell migration and potentiating anti-inflammatory and immunity pathways | [111] | |
| Human bone marrow-derived MSCs | Increased function of stem cells by increasing the expression of CXCR7 | [112] | |
| Cord blood mesenchymal stromal cells | Increased cell migration according to increased activity of SDF1/CXCR4 and SDF-1/CXCR7 signaling pathways | [113] | |
| Human bone marrow-mesenchymal stromal cells | Increased expression of KRIT1 and prevention of the accumulation of intracellular oxidants and reduction of oxidative stress | [114] | |
| Human umbilical cord-derived MSCs | Increased expression of mesenchymal and endodermal genes related to hepatic tissue/strengthening of CXCR4 signaling | [115] | |
| Human MSCs derived from adipose tissue | Increased MSCs migration ability by activating the CXCR4 signaling pathway | [116] | |
| Human bone marrow-derived mesenchymal stromal cells | Activation of CXCR4 signaling pathway and increased MSCs immortality | [117] | |
| Human bone marrow-derived MSCs | Increased expression of liver markers such as ALB, AFP, CK-18, TAT and increasing the differentiation of MSCs into liver cells | [118] | |
| Human umbilical cord-derived MSCs | Regulation of the expression of a group of miRNAs and hepatic differentiation | [119] | |
| Adipose tissue and bone marrow-derived MSCs | Increased expression of osteogenic genes such as RUNX, BMP2, p21WAF1, osterix, and osteopontin | [9] | |
| Tonsil-derived MSCs | Increased bone differentiation with CCN1 protein expression | [120] | |
| MSCs derived from pancreatic islets | Increased expression of genes related to beta cell neogenesis, such as NKX6.1/increase in the number of insulin-positive cells | [121] | |
| Mouse lip derived-MSCs | Increased expression of cardiac structural genes such as Cx43, βMHC, cTnI, and MLC2v and cardiac primary transcription genes such as NKX2.5, HAND2, HAND1, and GATA4 | [122] | |
| Human Wharton’s jelly MSCs | Increased expression of neuronal markers such as Nestin, Neuro-D1 | [123] | |
| Human bone marrow-derived mesenchymal stromal cells | Increased expression of neuronal markers such as GFAP, Musashi, CD133, Nestin-1, MAP-2, and KCNH2/5 | [124] | |
| Human bone marrow-derived MSCs | Increased expression of markers of mature neurons such as Th, VAChT and Htr2a/decreased expression of oligodendrocyte and primary neurons precursors | [125] | |
| Mammary fat tissue and cord blood MSCs | Cell cycle inhibition in G2/M phase/enhancement of p21CIP1/WAF1 signaling pathway activity and cell cycle inhibition | [126] | |
| Mouse MSCs derived from bone marrow | Regulation of the expression of genes involved in energy metabolism such as PGC-1α, Cox6b2, and Atp12a and genes involved in antioxidant defense such as Serpinb1b, Gpx6, and Mt2/increased expression of genes involved in cell stress pathway such as Hsp27, Hox, and Hsp A1l, and anti-apoptotic pathways such as Erc1, Naip1, and Faim2/elevated expression of growth and trophic factors such as FGF-15, FGF-21, NDNF, GDF-1, BMP-3, and NTF3 | [127] | |
| Human bone marrow-derived MSCs | Apoptosis induction and antiproliferative effects in T cells | [128] | |
| Cryopreserved rat MSC | Enhancement of CXCR4 signaling/increased expression of mesenchymal markers such as fibronectin and CD54 | [129] | |
| Human bone marrow-derived MSCs | Increased expression of tumor suppressor genes such as Cx43 and Cx26 and induction of apoptosis | [130] | |
| Phenytoin | Dental pulp-derived-MSCs Mouse lip derived-MSCs | Increased expression of osteoblast-related markers such as osteopontin, RUNX2, and ALP Increased expression of miR-196a-5p and defective cell proliferation | [131,132] |
| Levetiracetam | Adipose and bone marrow derived MSCs Human Wharton’s jelly MSCs | Modulation of the release of inhibitory and excitatory neurotransmitters/regulation of the release of inflammatory factors such as bFGF, TNF-a, IL-6, and BDNF Regulation of the expression of antioxidant genes such as Cu/ZnSOD, signaling proteins such as PEBP1/regulation of the expression of genes related to apoptosis, survival and cell death such as BDNF/GDNF | [133,134] |
| Pregabalin | Bone marrow-derived MSCs | Decreased Notch1/p38-MAPK signaling pathway activity/ Reduction of the level of inflammatory markers such as TNF-α, NF-κB, p65, and IL-6/ Increased level of antioxidants | [135] |
| Gabapentin | Ovine-derived mesenchymal stem cells | Increased speed of mesenchymal cell division | [136] |
| Phenobarbital | TERA2.cl.SP12 stem cells | Induction of necrosis in neurons and reducing their differentiation | [137] |
| Carbamazepine | TERA2.cl.SP12 stem cells | Impairment of cell proliferation and reduced cell viability | [137] |
| Lamotrigine | TERA2.cl.SP12 stem cells | Induction of necrosis and reduction of cell viability | [137] |
5.2. The Effect of Phenytoin on MSCs
5.3. The Effect of Levetiracetam (LEV) on MSCs
5.4. The Effect of Pregabalin on MSCs
5.5. The Effect of Gabapentin on MSCs
6. Human Stem Cell Lines and Risk of Developmental Neurotoxicity with ASMs
7. Limitations of MSCs
8. Ethical Issues
9. Potential Risks and Adverse Effects
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alqurashi, R.S.; Yee, A.S.; Malone, T.; Alrubiaan, S.; Tam, M.W.; Wang, K.; Nandedwalla, R.R.; Field, W.; Alkhelb, D.; Given, K.S. A Warburg-like metabolic program coordinates Wnt, AMPK, and mTOR signaling pathways in epileptogenesis. PLoS ONE 2021, 16, e0252282. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, D.; WHO-CHOICE. Cost-effectiveness of first-line antiepileptic drug treatments in the developing world: A population-level analysis. Epilepsia 2005, 46, 751–759. [Google Scholar] [CrossRef] [PubMed]
- St, L.; Erik, K. Truly “rational” polytherapy: Maximizing efficacy and minimizing drug interactions, drug load, and adverse effects. Curr. Neuropharmacol. 2009, 7, 96–105. [Google Scholar]
- Galanopoulou, A.S.; Kokaia, M.; Loeb, J.A.; Nehlig, A.; Pitkänen, A.; Rogawski, M.A.; Staley, K.J.; Whittemore, V.H.; Edward Dudek, F. Epilepsy therapy development: Technical and methodologic issues in studies with animal models. Epilepsia 2013, 54, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ka, A.; Taher, A.; D’Souza, S.; Barnes, E.H.; Gupta, S.; Troedson, C.; Wade, F.; Teo, O.; Dale, R.C.; Wong, C. Predictors of longitudinal seizure outcomes after epilepsy surgery in childhood. Epilepsy Behav. Rep. 2022, 19, 100561. [Google Scholar] [CrossRef]
- Goodarzi, P.; Aghayan, H.R.; Soleimani, M.; Norouzi-Javidan, A.; Mohamadi-Jahani, F.; Jahangiri, S.; Emami-Razavi, S.H.; Larijani, B.; Arjmand, B. Stem cell therapy for treatment of epilepsy. Acta Med. Iran. 2014, 651–655. [Google Scholar]
- Shetty, A.K. Hippocampal injury-induced cognitive and mood dysfunction, altered neurogenesis, and epilepsy: Can early neural stem cell grafting intervention provide protection? Epilepsy Behav. 2014, 38, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Hlebokazov, F.; Dakukina, T.; Potapnev, M.; Kosmacheva, S.; Moroz, L.; Misiuk, N.; Golubeva, T.; Slobina, E.; Krasko, O.; Shakhbazau, A. Clinical benefits of single vs repeated courses of mesenchymal stem cell therapy in epilepsy patients. Clin. Neurol. Neurosurg. 2021, 207, 106736. [Google Scholar] [CrossRef]
- Cho, H.H.; Park, H.T.; Kim, Y.J.; Bae, Y.C.; Suh, K.T.; Jung, J.S. Induction of osteogenic differentiation of human mesenchymal stem cells by histone deacetylase inhibitors. J. Cell. Biochem. 2005, 96, 533–542. [Google Scholar] [CrossRef]
- Odawara, A.; Katoh, H.; Matsuda, N.; Suzuki, I. Physiological maturation and drug responses of human induced pluripotent stem cell-derived cortical neuronal networks in long-term culture. Sci. Rep. 2016, 6, 26181. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kim, C.-H.; Moon, J.-I.; Chung, Y.-G.; Chang, M.-Y.; Han, B.-S.; Ko, S.; Yang, E.; Cha, K.Y.; Lanza, R. Generation of human induced pluripotent stem cells by direct delivery of reprogramming proteins. Cell Stem Cell 2009, 4, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, Y.; Bishop, C.E.; Treff, N.R.; Walker, S.J.; Sandler, V.M.; Becker, S.; Klimanskaya, I.; Wun, W.-S.; Dunn, R.; Hall, R.M. Reprogramming of human somatic cells using human and animal oocytes. Cloning Stem Cells 2009, 11, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Huber, A.; Padrun, V.; Déglon, N.; Aebischer, P.; Möhler, H.; Boison, D. Grafts of adenosine-releasing cells suppress seizures in kindling epilepsy. Proc. Natl. Acad. Sci. USA 2001, 98, 7611–7616. [Google Scholar] [CrossRef] [Green Version]
- Rajabzadeh, N.; Fathi, E.; Farahzadi, R. Stem cell-based regenerative medicine. Stem Cell Investig. 2019, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Najar, M.; Raicevic, G.; Fayyad-Kazan, H.; Bron, D.; Toungouz, M.; Lagneaux, L. Mesenchymal stromal cells and immunomodulation: A gathering of regulatory immune cells. Cytotherapy 2016, 18, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Konala, V.B.R.; Mamidi, M.K.; Bhonde, R.; Das, A.K.; Pochampally, R.; Pal, R. The current landscape of the mesenchymal stromal cell secretome: A new paradigm for cell-free regeneration. Cytotherapy 2016, 18, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Tewary, M.; Shakiba, N.; Zandstra, P.W. Stem cell bioengineering: Building from stem cell biology. Nat. Rev. Genet. 2018, 19, 595–614. [Google Scholar] [CrossRef]
- Lee, K.-H.; Tseng, W.-C.; Yang, C.-Y.; Tarng, D.-C. The anti-inflammatory, anti-oxidative, and anti-apoptotic benefits of stem cells in acute ischemic kidney injury. Int. J. Mol. Sci. 2019, 20, 3529. [Google Scholar] [CrossRef] [Green Version]
- Jager, L.D.; Canda, C.-M.A.; Hall, C.A.; Heilingoetter, C.L.; Huynh, J.; Kwok, S.S.; Kwon, J.H.; Richie, J.R.; Jensen, M.B. Effect of enzymatic and mechanical methods of dissociation on neural progenitor cells derived from induced pluripotent stem cells. Adv. Med. Sci. 2016, 61, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Stucky, E.C.; Schloss, R.S.; Yarmush, M.L.; Shreiber, D.I. Alginate micro-encapsulation of mesenchymal stromal cells enhances modulation of the neuro-inflammatory response. Cytotherapy 2015, 17, 1353–1364. [Google Scholar] [CrossRef] [Green Version]
- Boison, D. Engineered adenosine-releasing cells for epilepsy therapy: Human mesenchymal stem cells and human embryonic stem cells. Neurotherapeutics 2009, 6, 278–283. [Google Scholar] [CrossRef]
- Tesiye, M.R.; Kia, Z.A.; Rajabi-Maham, H. Mesenchymal stem cells and prostate cancer: A concise review of therapeutic potentials and biological aspects. Stem Cell Res. 2022, 63, 102864. [Google Scholar] [CrossRef] [PubMed]
- Aligholi, H.; Safahani, M.; Asadi-Pooya, A.A. Stem cell therapy in patients with epilepsy: A systematic review. Clin. Neurol. Neurosurg. 2021, 200, 106416. [Google Scholar] [CrossRef] [PubMed]
- Lo, B.; Parham, L. Ethical issues in stem cell research. Endocr. Rev. 2009, 30, 204–213. [Google Scholar] [CrossRef]
- Davis, D.S. Embryos created for research purposes. Kennedy Inst. Ethics J. 1995, 5, 343–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherwin, S. No Longer Patient: Feminist Ethics and Health Care; Temple University Press: Philadelphia, PA, USA, 1992. [Google Scholar]
- Baker, C.L.; Pera, M.F. Capturing totipotent stem cells. Cell Stem Cell 2018, 22, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, B.; Xu, J.; Wang, J.; Wu, J.; Shi, C.; Xu, Y.; Dong, J.; Wang, C.; Lai, W. Derivation of pluripotent stem cells with in vivo embryonic and extraembryonic potency. Cell 2017, 169, 243–257. e225. [Google Scholar] [CrossRef] [Green Version]
- Volarevic, V.; Markovic, B.S.; Gazdic, M.; Volarevic, A.; Jovicic, N.; Arsenijevic, N.; Armstrong, L.; Djonov, V.; Lako, M.; Stojkovic, M. Ethical and safety issues of stem cell-based therapy. Int. J. Med. Sci. 2018, 15, 36. [Google Scholar] [CrossRef] [Green Version]
- Moradi, S.; Mahdizadeh, H.; Šarić, T.; Kim, J.; Harati, J.; Shahsavarani, H.; Greber, B.; Moore, J.B. Research and therapy with induced pluripotent stem cells (iPSCs): Social, legal, and ethical considerations. Stem Cell Res. Ther. 2019, 10, 341. [Google Scholar] [CrossRef] [Green Version]
- Liang, J.; Wu, S.; Zhao, H.; Li, S.-l.; Liu, Z.-x.; Wu, J.; Zhou, L. Human umbilical cord mesenchymal stem cells derived from Wharton’s jelly differentiate into cholinergic-like neurons in vitro. Neurosci. Lett. 2013, 532, 59–63. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.-S.; Kim, H.O. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quimby, J.M.; Webb, T.L.; Randall, E.; Marolf, A.; Valdes-Martinez, A.; Dow, S.W. Assessment of intravenous adipose-derived allogeneic mesenchymal stem cells for the treatment of feline chronic kidney disease: A randomized, placebo-controlled clinical trial in eight cats. J. Feline Med. Surg. 2016, 18, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Cai, X.; Wang, L.; Liao, B.; Zhang, H.; Shan, Y.; Chen, Q.; Zhou, T.; Li, X.; Hou, J. Generating a non-integrating human induced pluripotent stem cell bank from urine-derived cells. PLoS ONE 2013, 8, e70573. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qin, H.; Qu, C.; Tuan, R.S.; Shi, S.; Huang, G.T.-J. iPS cells reprogrammed from human mesenchymal-like stem/progenitor cells of dental tissue origin. Stem Cells Dev. 2010, 19, 469–480. [Google Scholar] [CrossRef]
- Staerk, J.; Dawlaty, M.M.; Gao, Q.; Maetzel, D.; Hanna, J.; Sommer, C.A.; Mostoslavsky, G.; Jaenisch, R. Reprogramming of peripheral blood cells to induced pluripotent stem cells. Cell Stem Cell 2010, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Gage, F.H.; Temple, S. Neural stem cells: Generating and regenerating the brain. Neuron 2013, 80, 588–601. [Google Scholar] [CrossRef] [Green Version]
- Grade, S.; Bernardino, L.; Malva, J.O. Oligodendrogenesis from neural stem cells: Perspectives for remyelinating strategies. Int. J. Dev. Neurosci. 2013, 31, 692–700. [Google Scholar] [CrossRef]
- Ding, L.; Morrison, S.J. Haematopoietic stem cells and early lymphoid progenitors occupy distinct bone marrow niches. Nature 2013, 495, 231–235. [Google Scholar] [CrossRef] [Green Version]
- Peters, A.; Burridge, P.W.; Pryzhkova, M.V.; Levine, M.A.; Park, T.-S.; Roxbury, C.; Yuan, X.; Péault, B.; Zambidis, E.T. Challenges and strategies for generating therapeutic patient-specific hemangioblasts and hematopoietic stem cells from human pluripotent stem cells. Int. J. Dev. Biol. 2010, 54, 965. [Google Scholar] [CrossRef] [Green Version]
- Anasetti, C.; Logan, B.R.; Lee, S.J.; Waller, E.K.; Weisdorf, D.J.; Wingard, J.R.; Cutler, C.S.; Westervelt, P.; Woolfrey, A.; Couban, S. Peripheral-blood stem cells versus bone marrow from unrelated donors. New Engl. J. Med. 2012, 367, 1487–1496. [Google Scholar] [CrossRef] [Green Version]
- Briquet, A.; Dubois, S.; Bekaert, S.; Dolhet, M.; Beguin, Y.; Gothot, A. Prolonged ex vivo culture of human bone marrow mesenchymal stem cells influences their supportive activity toward NOD/SCID-repopulating cells and committed progenitor cells of B lymphoid and myeloid lineages. Haematologica 2010, 95, 47. [Google Scholar] [CrossRef] [PubMed]
- Vicinanza, C.; Aquila, I.; Scalise, M.; Cristiano, F.; Marino, F.; Cianflone, E.; Mancuso, T.; Marotta, P.; Sacco, W.; Lewis, F.C. Adult cardiac stem cells are multipotent and robustly myogenic: C-kit expression is necessary but not sufficient for their identification. Cell Death Differ. 2017, 24, 2101–2116. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Wang, A.; Yuan, F.; Yan, Z.; Liu, B.; Chu, J.S.; Helms, J.A.; Li, S. Differentiation of multipotent vascular stem cells contributes to vascular diseases. Nat. Commun. 2012, 3, 875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nitta, M.; Tamaki, T.; Tono, K.; Okada, Y.; Masuda, M.; Akatsuka, A.; Hoshi, A.; Usui, Y.; Terachi, T. Reconstitution of experimental neurogenic bladder dysfunction using skeletal muscle-derived multipotent stem cells. Transplantation 2010, 89, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef]
- Pelagalli, A.; Nardelli, A.; Lucarelli, E.; Zannetti, A.; Brunetti, A. Autocrine signals increase ovine mesenchymal stem cells migration through Aquaporin-1 and CXCR4 overexpression. J. Cell. Physiol. 2018, 233, 6241–6249. [Google Scholar] [CrossRef]
- Sobacchi, C.; Palagano, E.; Villa, A.; Menale, C. Soluble factors on stage to direct mesenchymal stem cells fate. Front. Bioeng. Biotechnol. 2017, 5, 32. [Google Scholar] [CrossRef]
- Baez-Jurado, E.; Hidalgo-Lanussa, O.; Barrera-Bailón, B.; Sahebkar, A.; Ashraf, G.M.; Echeverria, V.; Barreto, G.E. Secretome of mesenchymal stem cells and its potential protective effects on brain pathologies. Mol. Neurobiol. 2019, 56, 6902–6927. [Google Scholar] [CrossRef]
- Phinney, D.G.; Hill, K.; Michelson, C.; DuTreil, M.; Hughes, C.; Humphries, S.; Wilkinson, R.; Baddoo, M.; Bayly, E. Biological activities encoded by the murine mesenchymal stem cell transcriptome provide a basis for their developmental potential and broad therapeutic efficacy. Stem Cells 2006, 24, 186–198. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Voulgari-Kokota, A.; Fairless, R.; Karamita, M.; Kyrargyri, V.; Tseveleki, V.; Evangelidou, M.; Delorme, B.; Charbord, P.; Diem, R.; Probert, L. Mesenchymal stem cells protect CNS neurons against glutamate excitotoxicity by inhibiting glutamate receptor expression and function. Exp. Neurol. 2012, 236, 161–170. [Google Scholar] [CrossRef] [PubMed]
- De la Rosa, M.B.; Sharma, A.D.; Mallapragada, S.K.; Sakaguchi, D.S. Transdifferentiation of brain-derived neurotrophic factor (BDNF)-secreting mesenchymal stem cells significantly enhance BDNF secretion and Schwann cell marker proteins. J. Biosci. Bioeng. 2017, 124, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Zha, K.; Yang, Y.; Tian, G.; Sun, Z.; Yang, Z.; Li, X.; Sui, X.; Liu, S.; Zhao, J.; Guo, Q. Nerve growth factor (NGF) and NGF receptors in mesenchymal stem/stromal cells: Impact on potential therapies. Stem Cells Transl. Med. 2021, 10, 1008–1020. [Google Scholar] [CrossRef]
- Gu, J.; Huang, L.; Zhang, C.; Wang, Y.; Zhang, R.; Tu, Z.; Wang, H.; Zhou, X.; Xiao, Z.; Liu, Z. Therapeutic evidence of umbilical cord-derived mesenchymal stem cell transplantation for cerebral palsy: A randomized, controlled trial. Stem Cell Res. Ther. 2020, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Xiang, E.; Han, B.; Zhang, Q.; Rao, W.; Wang, Z.; Chang, C.; Zhang, Y.; Tu, C.; Li, C.; Wu, D. Human umbilical cord-derived mesenchymal stem cells prevent the progression of early diabetic nephropathy through inhibiting inflammation and fibrosis. Stem Cell Res. Ther. 2020, 11, 336. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Cheng, H.; Dai, G.; Wang, X.; Hua, R.; Liu, X.; Wang, P.; Chen, G.; Yue, W.; An, Y. Umbilical cord mesenchymal stem cell transplantation significantly improves neurological function in patients with sequelae of traumatic brain injury. Brain Res. 2013, 1532, 76–84. [Google Scholar] [CrossRef]
- Huang, P.-Y.; Shih, Y.-H.; Tseng, Y.-j.; Ko, T.-L.; Fu, Y.-S.; Lin, Y.-Y. Xenograft of human umbilical mesenchymal stem cells from Wharton’s jelly as a potential therapy for rat pilocarpine-induced epilepsy. Brain Behav. Immun. 2016, 54, 45–58. [Google Scholar] [CrossRef] [Green Version]
- Costa-Ferro, Z.S.M.; de Borba Cunha, F.; de Freitas Souza, B.S.; Leal, M.M.T.; da Silva, A.A.; de Bellis Kühn, T.I.B.; Forte, A.; Sekiya, E.J.; Soares, M.B.P.; Dos Santos, R.R. Antiepileptic and neuroprotective effects of human umbilical cord blood mononuclear cells in a pilocarpine-induced epilepsy model. Cytotechnology 2014, 66, 193–199. [Google Scholar] [CrossRef] [Green Version]
- Mohammed, A.S.; Ewais, M.M.; Tawfik, M.K.; Essawy, S.S. Effects of intravenous human umbilical cord blood mesenchymal stem cell therapy versus gabapentin in pentylenetetrazole-induced chronic epilepsy in rats. Pharmacology 2014, 94, 41–50. [Google Scholar] [CrossRef]
- Park, G.Y.; Lee, E.M.; Seo, M.-S.; Seo, Y.-J.; Oh, J.S.; Son, W.-C.; Kim, K.S.; Kim, J.S.; Kang, J.K.; Kang, K.-S. Preserved hippocampal glucose metabolism on 18F-FDG PET after transplantation of human umbilical cord blood-derived mesenchymal stem cells in chronic epileptic rats. J. Korean Med. Sci. 2015, 30, 1232–1240. [Google Scholar] [CrossRef] [Green Version]
- Luo, Q.; Xian, P.; Wang, T.; Wu, S.; Sun, T.; Wang, W.; Wang, B.; Yang, H.; Yang, Y.; Wang, H. Antioxidant activity of mesenchymal stem cell-derived extracellular vesicles restores hippocampal neurons following seizure damage. Theranostics 2021, 11, 5986. [Google Scholar] [CrossRef] [PubMed]
- Hlebokazov, F.; Dakukina, T.; Ihnatsenko, S.; Kosmacheva, S.; Potapnev, M.; Shakhbazau, A.; Goncharova, N.; Makhrov, M.; Korolevich, P.; Misyuk, N. Treatment of refractory epilepsy patients with autologous mesenchymal stem cells reduces seizure frequency: An open label study. Adv. Med. Sci. 2017, 62, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Fukumura, S.; Sasaki, M.; Kataoka-Sasaki, Y.; Oka, S.; Nakazaki, M.; Nagahama, H.; Morita, T.; Sakai, T.; Tsutsumi, H.; Kocsis, J.D. Intravenous infusion of mesenchymal stem cells reduces epileptogenesis in a rat model of status epilepticus. Epilepsy Res. 2018, 141, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Ali, I.; Aertgeerts, S.; Le Blon, D.; Bertoglio, D.; Hoornaert, C.; Ponsaerts, P.; Dedeurwaerdere, S. Intracerebral delivery of the M2 polarizing cytokine interleukin 13 using mesenchymal stem cell implants in a model of temporal lobe epilepsy in mice. Epilepsia 2017, 58, 1063–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huicong, K.; Zheng, X.; Furong, W.; Zhouping, T.; Feng, X.; Qi, H.; Xiaoyan, L.; Xiaojiang, H.; Na, Z.; Ke, X. The imbalanced expression of adenosine receptors in an epilepsy model corrected using targeted mesenchymal stem cell transplantation. Mol. Neurobiol. 2013, 48, 921–930. [Google Scholar] [CrossRef]
- Güttinger, M.; Fedele, D.; Koch, P.; Padrun, V.; Pralong, W.F.; Brüstle, O.; Boison, D. Suppression of kindled seizures by paracrine adenosine release from stem cell–derived brain implants. Epilepsia 2005, 46, 1162–1169. [Google Scholar] [CrossRef]
- Li, T.; Ren, G.; Kaplan, D.L.; Boison, D. Human mesenchymal stem cell grafts engineered to release adenosine reduce chronic seizures in a mouse model of CA3-selective epileptogenesis. Epilepsy Res. 2009, 84, 238–241. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Li, T.; Lan, J.Q.; Wilz, A.; Simon, R.P.; Boison, D. Lentiviral RNAi-induced downregulation of adenosine kinase in human mesenchymal stem cell grafts: A novel perspective for seizure control. Exp. Neurol. 2007, 208, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Abdanipour, A.; Tiraihi, T.; Mirnajafi-Zadeh, J. Improvement of the pilocarpine epilepsy model in rat using bone marrow stromal cell therapy. Neurol. Res. 2011, 33, 625–632. [Google Scholar] [CrossRef]
- Costa-Ferro, Z.S.; Vitola, A.S.; Pedroso, M.F.; Cunha, F.B.; Xavier, L.L.; Machado, D.C.; Soares, M.B.; Ribeiro-dos-Santos, R. Prevention of seizures and reorganization of hippocampal functions by transplantation of bone marrow cells in the acute phase of experimental epilepsy. Seizure 2010, 19, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Costa-Ferro, Z.S.; Souza, B.S.; Leal, M.M.; Kaneto, C.M.; Azevedo, C.M.; da Silva, I.C.; Soares, M.B.; Ribeiro-dos-Santos, R.; DaCosta, J.C. Transplantation of bone marrow mononuclear cells decreases seizure incidence, mitigates neuronal loss and modulates pro-inflammatory cytokine production in epileptic rats. Neurobiol. Dis. 2012, 46, 302–313. [Google Scholar] [CrossRef]
- Long, Q.; Qiu, B.; Wang, K.; Yang, J.; Jia, C.; Xin, W.; Wang, P.; Han, R.; Fei, Z.; Liu, W. Genetically engineered bone marrow mesenchymal stem cells improve functional outcome in a rat model of epilepsy. Brain Res. 2013, 1532, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Long, Q.; Qiu, B.; Liu, W.; Fei, Z.; Feng, G.; Wang, P.; Zhong, J.; Yi, X.; Liu, Y.; Zhang, Y. Functional recovery and neuronal regeneration of a rat model of epilepsy by transplantation of Hes1-down regulated bone marrow stromal cells. Neuroscience 2012, 212, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Zanirati, G.; Azevedo, P.N.; Marinowic, D.R.; Rodrigues, F.; de Oliveira Dias, A.C.; Venturin, G.T.; Greggio, S.; Simão, F.; DaCosta, J.C. Transplantation of bone marrow mononuclear cells modulates hippocampal expression of growth factors in chronically epileptic animals. CNS Neurosci. Ther. 2015, 21, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Kassab, R.B.; Salem, F.E.H.; Lokman, M.S. The anticonvulsant activity of bone marrow-derived mesenchymal stem cells in ptz-induced epileptic seizures. Int. J. Pharm. Biol. Sci. 2015, 6, 444–455. [Google Scholar]
- Salem, N.A.; El-Shamarka, M.; Khadrawy, Y.; El-Shebiney, S. New prospects of mesenchymal stem cells for ameliorating temporal lobe epilepsy. Inflammopharmacology 2018, 26, 963–972. [Google Scholar] [CrossRef]
- Leal, M.M.T.; Costa-Ferro, Z.S.M.; de Freitas Souza, B.S.; Azevedo, C.M.; Carvalho, T.M.; Kaneto, C.M.; Carvalho, R.H.; Dos Santos, R.R.; Soares, M.B.P. Early transplantation of bone marrow mononuclear cells promotes neuroprotection and modulation of inflammation after status epilepticus in mice by paracrine mechanisms. Neurochem. Res. 2014, 39, 259–268. [Google Scholar] [CrossRef]
- Salah, R.S.; Ahmed, H.H.; Abd-Allah, S.H.; Hassan, R.E.; Khalil, W.K.; Abd-Rabou, A.A.; Sabry, G.M. The anti-epileptic efficiency of mesenchymal stem cells against pilocarpine model of acute epilepsy. Res. J. Pharm. Technol. 2021, 14, 1255–1266. [Google Scholar] [CrossRef]
- DaCosta, J.C.; Portuguez, M.W.; Marinowic, D.R.; Schilling, L.P.; Torres, C.M.; DaCosta, D.I.; Carrion, M.J.M.; Raupp, E.F.; Machado, D.C.; Soder, R.B. Safety and seizure control in patients with mesial temporal lobe epilepsy treated with regional superselective intra-arterial injection of autologous bone marrow mononuclear cells. J. Tissue Eng. Regen. Med. 2018, 12, e648–e656. [Google Scholar] [CrossRef]
- Venturin, G.T.; Greggio, S.; Marinowic, D.R.; Zanirati, G.; Cammarota, M.; Machado, D.C.; DaCosta, J.C. Bone marrow mononuclear cells reduce seizure frequency and improve cognitive outcome in chronic epileptic rats. Life Sci. 2011, 89, 229–234. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhao, Y.; Pan, X.; Zhang, Y.; Lin, L.; Wu, Y.; Huang, Y.; He, H. Adipose-derived stem cell transplantation improves learning and memory via releasing neurotrophins in rat model of temporal lobe epilepsy. Brain Res. 2021, 1750, 147121. [Google Scholar] [CrossRef] [PubMed]
- Jeon, D.; Chu, K.; Lee, S.T.; Jung, K.H.; Kang, K.M.; Ban, J.J.; Kim, S.; Seo, J.S.; Won, C.H.; Kim, M. A cell-free extract from human adipose stem cells protects mice against epilepsy. Epilepsia 2011, 52, 1617–1626. [Google Scholar] [CrossRef] [PubMed]
- Tamura, B.; Almeida, D.; Felizardo, R.; Olanda, G.; Bocca, L.; Pinhal, N.; Alves-de-Moraes, L.; Covolan, L.; Cãmara, N.; Longo, B. Convulsive seizure protection after hippocampal transplantation of mesenchymal cells from adipose tissue in mice. J. Stem Cell Res. Ther. 2014, 4, 2. [Google Scholar]
- Zhao, H.; Li, S.; He, L.; Tang, F.; Han, X.; Deng, W.; Lin, Z.; Huang, R.; Li, Z. Ameliorating Effect of Umbilical Cord Mesenchymal Stem Cells in a Human Induced Pluripotent Stem Cell Model of Dravet Syndrome. Mol. Neurobiol. 2021, 59, 749–761. [Google Scholar] [CrossRef] [PubMed]
- Milczarek, O.; Jarocha, D.; Starowicz–Filip, A.; Kwiatkowski, S.; Badyra, B.; Majka, M. Multiple autologous bone marrow-derived CD271+ mesenchymal stem cell transplantation overcomes drug-resistant epilepsy in children. Stem Cells Transl. Med. 2018, 7, 20–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepanik, E.; Mierzewska, H.; Antczak-Marach, D.; Figiel-Dabrowska, A.; Terczynska, I.; Tryfon, J.; Krzesniak, N.; Noszczyk, B.H.; Sawicka, E.; Domanska-Janik, K. Intrathecal infusion of autologous adipose-derived regenerative cells in autoimmune refractory epilepsy: Evaluation of safety and efficacy. Stem Cells Int. 2020, 2020, 7104243. [Google Scholar] [CrossRef] [Green Version]
- Hunt, R.F.; Girskis, K.M.; Rubenstein, J.L.; Alvarez-Buylla, A.; Baraban, S.C. GABA progenitors grafted into the adult epileptic brain control seizures and abnormal behavior. Nat. Neurosci. 2013, 16, 692–697. [Google Scholar] [CrossRef] [Green Version]
- Mazini, L.; Rochette, L.; Admou, B.; Amal, S.; Malka, G. Hopes and limits of adipose-derived stem cells (ADSCs) and mesenchymal stem cells (MSCs) in wound healing. Int. J. Mol. Sci. 2020, 21, 1306. [Google Scholar] [CrossRef] [Green Version]
- Sheykhhasan, M.; Wong, J.K.; Seifalian, A.M. Human adipose-derived stem cells with great therapeutic potential. Curr. Stem Cell Res. Ther. 2019, 14, 532–548. [Google Scholar] [CrossRef]
- Mead, B.; Logan, A.; Berry, M.; Leadbeater, W.; Scheven, B.A. Paracrine-mediated neuroprotection and neuritogenesis of axotomised retinal ganglion cells by human dental pulp stem cells: Comparison with human bone marrow and adipose-derived mesenchymal stem cells. PLoS ONE 2014, 9, e109305. [Google Scholar] [CrossRef]
- Lunn, J.S.; Sakowski, S.A.; McGinley, L.M.; Pacut, C.; Hazel, T.G.; Johe, K.; Feldman, E.L. Autocrine production of IGF-I increases stem cell-mediated neuroprotection. Stem Cells 2015, 33, 1480–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laroni, A.; Rosbo, N.K.d.; Uccelli, A. Mesenchymal stem cells for the treatment of neurological diseases: Immunoregulation beyond neuroprotection. Immunol. Lett. 2015, 168, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Fresnedo, A.; Perez-Vega, C.; Domingo, R.A.; Lee, S.J.; Perkerson, R.B.; Zubair, A.C.; Takahisa, K.; Tatum, W.; Quinones-Hinojosa, A.; Middlebrooks, E.H. Mesenchymal stem cell therapy for focal epilepsy: A systematic review of preclinical models and clinical studies. Epilepsia 2022, 63, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Waldau, B.; Hattiangady, B.; Kuruba, R.; Shetty, A.K. Medial ganglionic eminence-derived neural stem cell grafts ease spontaneous seizures and restore GDNF expression in a rat model of chronic temporal lobe epilepsy. Stem Cells 2010, 28, 1153–1164. [Google Scholar] [CrossRef]
- Casalia, M.L.; Howard, M.A.; Baraban, S.C. Persistent seizure control in epileptic mice transplanted with gamma-aminobutyric acid progenitors. Ann. Neurol. 2017, 82, 530–542. [Google Scholar] [CrossRef]
- Shetty, A.K.; Upadhya, D. GABA-ergic cell therapy for epilepsy: Advances, limitations and challenges. Neurosci. Biobehav. Rev. 2016, 62, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Henderson, K.W.; Gupta, J.; Tagliatela, S.; Litvina, E.; Zheng, X.; Van Zandt, M.A.; Woods, N.; Grund, E.; Lin, D.; Royston, S. Long-term seizure suppression and optogenetic analyses of synaptic connectivity in epileptic mice with hippocampal grafts of GABAergic interneurons. J. Neurosci. 2014, 34, 13492–13504. [Google Scholar] [CrossRef] [Green Version]
- Upadhya, D.; Hattiangady, B.; Castro, O.W.; Shuai, B.; Kodali, M.; Attaluri, S.; Bates, A.; Dong, Y.; Zhang, S.-C.; Prockop, D.J. Human induced pluripotent stem cell-derived MGE cell grafting after status epilepticus attenuates chronic epilepsy and comorbidities via synaptic integration. Proc. Natl. Acad. Sci. USA 2019, 116, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Gurvich, N.; Klein, P.S. Lithium and valproic acid: Parallels and contrasts in diverse signaling contexts. Pharm. Ther. 2002, 96, 45–66. [Google Scholar] [CrossRef]
- Johannessen, C.U. Mechanisms of action of valproate: A commentatory. Neurochem. Int. 2000, 37, 103–110. [Google Scholar] [CrossRef]
- Kim, H.N.; Shin, J.Y.; Kim, D.Y.; Lee, J.E.; Lee, P.H. Priming mesenchymal stem cells with uric acid enhances neuroprotective properties in parkinsonian models. J. Tissue Eng. 2021, 12, 20417314211004816. [Google Scholar] [CrossRef]
- Kim, D.S.; Jang, I.K.; Lee, M.W.; Ko, Y.J.; Lee, D.-H.; Lee, J.W.; Sung, K.W.; Koo, H.H.; Yoo, K.H. Enhanced immunosuppressive properties of human mesenchymal stem cells primed by interferon-γ. EBioMedicine 2018, 28, 261–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamora, D.; Lim, S.H.; Pan, A.; Tan, L.; Tan, E.K. Valproate-induced Parkinsonism in epilepsy patients. Mov. Disord. 2007, 22, 130–133. [Google Scholar] [CrossRef] [PubMed]
- Easterford, K.; Clough, P.; Kellett, M.; Fallon, K.; Duncan, S. Reversible parkinsonism with normal β-CIT-SPECT in patients exposed to sodium valproate. Neurology 2004, 62, 1435–1437. [Google Scholar] [CrossRef] [PubMed]
- Ristić, A.J.; Vojvodić, N.; Janković, S.; Sindelić, A.; Sokić, D. The frequency of reversible parkinsonism and cognitive decline associated with valproate treatment: A study of 364 patients with different types of epilepsy. Epilepsia 2006, 47, 2183–2185. [Google Scholar] [CrossRef]
- Meador, K.J.; Baker, G.A.; Browning, N.; Clayton-Smith, J.; Combs-Cantrell, D.T.; Cohen, M.; Kalayjian, L.A.; Kanner, A.; Liporace, J.D.; Pennell, P.B. Cognitive function at 3 years of age after fetal exposure to antiepileptic drugs. N. Engl. J. Med. 2009, 360, 1597–1605. [Google Scholar] [CrossRef] [Green Version]
- Meador, K.J.; Penovich, P.; Baker, G.A.; Pennell, P.B.; Bromfield, E.; Pack, A.; Liporace, J.D.; Sam, M.; Kalayjian, L.A.; Thurman, D.J. Antiepileptic drug use in women of childbearing age. Epilepsy Behav. 2009, 15, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Ornoy, A. Valproic acid in pregnancy: How much are we endangering the embryo and fetus? Reprod. Toxicol. 2009, 28, 1–10. [Google Scholar] [CrossRef]
- Levy, O.; Rothhammer, V.; Mascanfroni, I.; Tong, Z.; Kuai, R.; De Biasio, M.; Wang, Q.; Majid, T.; Perrault, C.; Yeste, A. A cell-based drug delivery platform for treating central nervous system inflammation. J. Mol. Med. 2021, 99, 663–671. [Google Scholar] [CrossRef]
- Tsai, L.-K.; Leng, Y.; Wang, Z.; Leeds, P.; Chuang, D.-M. The mood stabilizers valproic acid and lithium enhance mesenchymal stem cell migration via distinct mechanisms. Neuropsychopharmacology 2010, 35, 2225–2237. [Google Scholar] [CrossRef] [Green Version]
- Bidkhori, H.R.; Ahmadiankia, N.; Matin, M.M.; Heirani-Tabasi, A.; Farshchian, M.; Naderi-Meshkin, H.; Shahriyari, M.; Dastpak, M.; Bahrami, A.R. Chemically primed bone-marrow derived mesenchymal stem cells show enhanced expression of chemokine receptors contributed to their migration capability. Iran. J. Basic Med. Sci. 2016, 19, 14. [Google Scholar] [PubMed]
- Marquez-Curtis, L.A.; Qiu, Y.; Xu, A.; Janowska-Wieczorek, A. Migration, proliferation, and differentiation of cord blood mesenchymal stromal cells treated with histone deacetylase inhibitor valproic acid. Stem Cells Int. 2014, 2014, 610495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, K.H.; Han, D.M.R.; Jeong, S.-G.; Choi, M.R.; Chai, Y.G.; Cho, G.-W. Proteomic analysis reveals KRIT1 as a modulator for the antioxidant effects of valproic acid in human bone-marrow mesenchymal stromal cells. Drug Chem. Toxicol. 2015, 38, 286–292. [Google Scholar] [CrossRef]
- An, S.Y.; Han, J.; Lim, H.J.; Park, S.Y.; Kim, J.H.; Do, B.R.; Kim, J.H. Valproic acid promotes differentiation of hepatocyte-like cells from whole human umbilical cord-derived mesenchymal stem cells. Tissue Cell 2014, 46, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Heirani-Tabasi, A.; Naderi-Meshkin, H.; Matin, M.M.; Mirahmadi, M.; Shahriyari, M.; Ahmadiankia, N.; Sanjar Moussavi, N.; Bidkhori, H.R.; Raeesolmohaddeseen, M.; Bahrami, A.R. Augmented migration of mesenchymal stem cells correlates with the subsidiary CXCR4 variant. Cell Adhes. Migr. 2018, 12, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Piñeiro-Ramil, M.; Castro-Viñuelas, R.; Sanjurjo-Rodríguez, C.; Rodríguez-Fernández, S.; Hermida-Gómez, T.; Blanco-García, F.J.; Fuentes-Boquete, I.; Díaz-Prado, S. Immortalizing mesenchymal stromal cells from aged donors while keeping their essential features. Stem Cells Int. 2020, 2020, 5726947. [Google Scholar] [CrossRef]
- Rashid, S.; Qazi, R.-e.-M.; Malick, T.S.; Salim, A.; Khan, I.; Ilyas, A.; Haneef, K. Effect of valproic acid on the hepatic differentiation of mesenchymal stem cells in 2D and 3D microenvironments. Mol. Cell. Biochem. 2021, 476, 909–919. [Google Scholar] [CrossRef]
- Raut, A.; Khanna, A. Enhanced expression of hepatocyte-specific microRNAs in valproic acid mediated hepatic trans-differentiation of human umbilical cord derived mesenchymal stem cells. Exp. Cell Res. 2016, 343, 237–247. [Google Scholar] [CrossRef]
- Yu, Y.; Oh, S.-Y.; Kim, H.Y.; Choi, J.-Y.; Jo, S.A.; Jo, I. Valproic Acid-Induced CCN1 Promotes Osteogenic Differentiation by Increasing CCN1 Protein Stability through HDAC1 Inhibition in Tonsil-Derived Mesenchymal Stem Cells. Cells 2022, 11, 534. [Google Scholar] [CrossRef]
- Coskun, E.; Ercin, M.; Gezginci-Oktayoglu, S. The role of epigenetic regulation and pluripotency-related microRNAs in differentiation of pancreatic stem cells to beta cells. J. Cell. Biochem. 2018, 119, 455–467. [Google Scholar] [CrossRef]
- Najafipour, H.; Bagheri-Hosseinabadi, Z.; Eslaminejad, T.; Mollaei, H.R. The effect of sodium valproate on differentiation of human adipose-derived stem cells into cardiomyocyte-like cells in two-dimensional culture and fibrin scaffold conditions. Cell Tissue Res. 2019, 378, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, S.; Tiraihi, T.; Soleimani, M. Differentiation of mesenchymal stem cells into neuron-like cells using composite 3D scaffold combined with valproic acid induction. J. Biomater. Appl. 2018, 32, 702–715. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-G.; Ohn, T.; Kim, S.H.; Cho, G.-W. Valproic acid promotes neuronal differentiation by induction of neuroprogenitors in human bone-marrow mesenchymal stromal cells. Neurosci. Lett. 2013, 554, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Fila-Danilow, A.; Borkowska, P.; Paul-Samojedny, M.; Kowalczyk, M.; Kowalski, J. The influence of TSA and VPA on the in vitro differentiation of bone marrow mesenchymal stem cells into neuronal lineage cells: Gene expression studies. Adv. Hyg. Exp. Med./Postep. Hig. I Med. Dosw. 2017, 71, 573897. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Park, J.R.; Seo, M.S.; Roh, K.H.; Park, S.B.; Hwang, J.W.; Sun, B.; Seo, K.; Lee, Y.S.; Kang, S.K.; et al. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif. 2009, 42, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Linares, G.R.; Chiu, C.-T.; Scheuing, L.; Leng, Y.; Liao, H.-M.; Maric, D.; Chuang, D.-M. Preconditioning mesenchymal stem cells with the mood stabilizers lithium and valproic acid enhances therapeutic efficacy in a mouse model of Huntington’s disease. Exp. Neurol. 2016, 281, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Killer, M.C.; Nold, P.; Henkenius, K.; Fritz, L.; Riedlinger, T.; Barckhausen, C.; Frech, M.; Hackstein, H.; Neubauer, A.; Brendel, C. Immunosuppressive capacity of mesenchymal stem cells correlates with metabolic activity and can be enhanced by valproic acid. Stem Cell Res. Ther. 2017, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, L.-K.; Wang, Z.; Munasinghe, J.; Leng, Y.; Leeds, P.; Chuang, D.-M. Mesenchymal stem cells primed with valproate and lithium robustly migrate to infarcted regions and facilitate recovery in a stroke model. Stroke 2011, 42, 2932–2939. [Google Scholar] [CrossRef]
- Ryu, C.H.; Park, K.Y.; Kim, S.M.; Jeong, C.H.; Woo, J.S.; Hou, Y.; Jeun, S.-S. Valproic acid enhances anti-tumor effect of mesenchymal stem cell mediated HSV-TK gene therapy in intracranial glioma. Biochem. Biophys. Res. Commun. 2012, 421, 585–590. [Google Scholar] [CrossRef]
- Asgharian-Rezaee, M.; Alipour-Farmad, R.; Tayarani-Najaran, Z. Comparison of osteogenic potential of phenytoin with dexamethasone in cultured dental pulp stem cells. Rep. Biochem. Mol. Biol. 2020, 9, 331. [Google Scholar] [CrossRef]
- Yoshioka, H.; Ramakrishnan, S.S.; Suzuki, A.; Iwata, J. Phenytoin inhibits cell proliferation through microRNA-196a-5p in mouse lip mesenchymal cells. Int. J. Mol. Sci. 2021, 22, 1746. [Google Scholar] [CrossRef] [PubMed]
- Hegazy, M.I.; Asaad, A.M.; Rashed, L.A.; Ahmed, H.H. Combined Treatment of Levetiracetam and Mesenchymal Stem Cells Reverses the Biochemical Aberrations in the Acute Phase of Epilepsy Induced by Pilocarpine in Rats. Biomed. Pharmacol. J. 2022, 15, 91–115. [Google Scholar] [CrossRef]
- Abd El Motteleb, D.M.; Hussein, S.; Hasan, M.M.; Mosaad, H. Comparison between the effect of human Wharton’s jelly–derived mesenchymal stem cells and levetiracetam on brain infarcts in rats. J. Cell. Biochem. 2018, 119, 9790–9800. [Google Scholar] [CrossRef] [PubMed]
- Al-Massri, K.F.; Ahmed, L.A.; El-Abhar, H.S. Mesenchymal stem cells therapy enhances the efficacy of pregabalin and prevents its motor impairment in paclitaxel-induced neuropathy in rats: Role of Notch1 receptor and JAK/STAT signaling pathway. Behav. Brain Res. 2019, 360, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Sauerbier, S.; Gutwald, R.; Wiedmann-Al-Ahmad, M.; Haberstroh, J.; Obermeyer, J.; Kuenz, A.; Betz, H.; Wolter, F.; Duttenhöfer, F.; Schmelzeisen, R. Effect of Gabapentin-Lactam and Gamma-Aminobutyric Acid/Lactam Analogs on Proliferation and Phenotype of Ovine Mesenchymal Stem Cells. Int. J. Oral Maxillofac. Implant. 2013, 28, e230–e238. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.S.; Livesey, J.C.; Halliwell, R.F. An evaluation of a human stem cell line to identify risk of developmental neurotoxicity with antiepileptic drugs. Toxicology In Vitro 2015, 29, 592–599. [Google Scholar] [CrossRef] [PubMed]
- Cammalleri, M.; Martini, D.; Ristori, C.; Timperio, A.M.; Bagnoli, P. Vascular endothelial growth factor up-regulation in the mouse hippocampus and its role in the control of epileptiform activity. Eur. J. Neurosci. 2011, 33, 482–498. [Google Scholar] [CrossRef] [PubMed]
- Carpentino, J.E.; Hartman, N.W.; Grabel, L.B.; Naegele, J.R. Region-specific differentiation of embryonic stem cell-derived neural progenitor transplants into the adult mouse hippocampus following seizures. J. Neurosci. Res. 2008, 86, 512–524. [Google Scholar] [CrossRef]
- Rüschenschmidt, C.; Koch, P.G.; Brüstle, O.; Beck, H. Functional Properties of ES Cell–Derived Neurons Engrafted into the Hippocampus of Adult Normal and Chronically Epileptic Rats. Epilepsia 2005, 46, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Maisano, X.; Litvina, E.; Tagliatela, S.; Aaron, G.B.; Grabel, L.B.; Naegele, J.R. Differentiation and functional incorporation of embryonic stem cell-derived GABAergic interneurons in the dentate gyrus of mice with temporal lobe epilepsy. J. Neurosci. 2012, 32, 46–61. [Google Scholar] [CrossRef] [Green Version]
- Assen, L.S.; Jongsma, K.R.; Isasi, R.; Tryfonidou, M.A.; Bredenoord, A.L. Recognizing the ethical implications of stem cell research: A call for broadening the scope. Stem Cell Rep. 2021, 16, 1656–1661. [Google Scholar] [CrossRef] [PubMed]
- Zarzeczny, A.; Scott, C.; Hyun, I.; Bennett, J.; Chandler, J.; Chargé, S.; Heine, H.; Isasi, R.; Kato, K.; Lovell-Badge, R. iPS cells: Mapping the policy issues. Cell 2009, 139, 1032–1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugan, V. Stem cell issue: Embryonic Stem Cell Research: A Decade of Debate from Bush to Obama. Yale J. Biol. Med. 2009, 82, 101. [Google Scholar] [PubMed]
- Renesme, L.; Cobey, K.D.; Le, M.; Lalu, M.M.; Thebaud, B. Establishment of a consensus definition for mesenchymal stromal cells (MSC) and reporting guidelines for clinical trials of MSC therapy: A modified Delphi study protocol. BMJ Open 2021, 11, e054740. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Musiał-Wysocka, A.; Kot, M.; Majka, M. The pros and cons of mesenchymal stem cell-based therapies. Cell Transplant. 2019, 28, 801–812. [Google Scholar] [CrossRef] [Green Version]
- Molendijk, I.; Bonsing, B.A.; Roelofs, H.; Peeters, K.C.; Wasser, M.N.; Dijkstra, G.; van der Woude, C.J.; Duijvestein, M.; Veenendaal, R.A.; Zwaginga, J.-J. Allogeneic bone marrow–derived mesenchymal stromal cells promote healing of refractory perianal fistulas in patients with Crohn’s disease. Gastroenterology 2015, 149, 918–927. e916. [Google Scholar] [CrossRef] [Green Version]
- Shi, M.; Zhang, Z.; Xu, R.; Lin, H.; Fu, J.; Zou, Z.; Zhang, A.; Shi, J.; Chen, L.; Lv, S. Human mesenchymal stem cell transfusion is safe and improves liver function in acute-on-chronic liver failure patients. Stem Cells Transl. Med. 2012, 1, 725–731. [Google Scholar] [CrossRef]
- Volarevic, V.; Ljujic, B.; Stojkovic, P.; Lukic, A.; Arsenijevic, N.; Stojkovic, M. Human stem cell research and regenerative medicinepresent and future. Br. Med. Bull. 2011. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Eckert, M.A.; Riazifar, H.; Kang, D.-K.; Agalliu, D.; Zhao, W. From blood to the brain: Can systemically transplanted mesenchymal stem cells cross the blood-brain barrier? Stem Cells Int. 2013, 2013, 435093. [Google Scholar] [CrossRef]
- Liu, X.; Ye, R.; Yan, T.; Yu, S.P.; Wei, L.; Xu, G.; Fan, X.; Jiang, Y.; Stetler, R.A.; Liu, G. Cell based therapies for ischemic stroke: From basic science to bedside. Prog. Neurobiol. 2014, 115, 92–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walczak, P.; Zhang, J.; Gilad, A.A.; Kedziorek, D.A.; Ruiz-Cabello, J.; Young, R.G.; Pittenger, M.F.; Van Zijl, P.C.; Huang, J.; Bulte, J.W. Dual-modality monitoring of targeted intraarterial delivery of mesenchymal stem cells after transient ischemia. Stroke 2008, 39, 1569–1574. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, B.A. Stem Cell Therapies as an Emerging Paradigm in Stroke (STEPS): Bridging basic and clinical science for cellular and neurogenic factor therapy in treating stroke. Stroke 2009, 40, 510–515. [Google Scholar]
- Kean, T.J.; Lin, P.; Caplan, A.I.; Dennis, J.E. MSCs: Delivery routes and engraftment, cell-targeting strategies, and immune modulation. Stem Cells Int. 2013, 2013, 732742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Peng, Q.; Wang, H.; Pan, J.; Zhou, Y.; Pan, K.; Li, J.; Wu, Y.; Wang, Y. Intrathecal injection of allogenic bone marrow-derived mesenchymal stromal cells in treatment of patients with severe ischemic stroke: Study protocol for a randomized controlled observer-blinded trial. Transl. Stroke Res. 2019, 10, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Cacione, D.G.; do Carmo Novaes, F.; Moreno, D.H. Stem cell therapy for treatment of thromboangiitis obliterans (Buerger’s disease). Cochrane Database Syst. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Kim, S.-n.; Lee, H.-J.; Kim, J.; Cho, Y.-K.; Shin, D.-H.; Tak, S.-J.; Moon, S.-H.; Kang, J.-E.; Ji, I.-M. Manufacture of clinical-grade human clonal mesenchymal stem cell products from single colony forming unit-derived colonies based on the subfractionation culturing method. Tissue Eng. Part C Methods 2015, 21, 1251–1262. [Google Scholar] [CrossRef]

| Stem Cells | 1-Allogenic adult stem cells, derived from another person’s specialized tissue; have ethical concern 2-Autologous adult stem cells, derived from the individual’s own specialized tissue; without ethical concern | Embryonic stem cells | Origin | Characteristic | Types | Ethical Issues | Ref |
| Derived from an embryo in the first four blastomeric cleavage phases | Totipotent | - |
| [27] | |||
| Derived from an embryo after the fourth blastomeric cleavage phase | Pluripotent | Embryonic stem cell | [28] | ||||
| Adult stem cells | Mesenchymal stem cells (MSCs) | Multipotent | Umbilical cord-derived mesenchymal stem cells, bone marrow, and adipose-derived mesenchymal stem cells | Since the use of these cells does not require the destruction of the embryo, it brings fewer ethical problems. However, as allogenic stem cells contain donor DNA, this may be associated with safety, legal and ethical challenges associated with the privacy of individuals. Autologous stem cells do not pose a particular ethical problem and do not interfere with the recipient’s immune system [29,30] | [31,32,33] | ||
| Induced pluripotent stem cells | Pluripotent | Fibroblasts, dental pulp, deciduous teeth, renal epithelial cells, and urine | [34,35,36] | ||||
| Neural stem cells. Derived out of the periventricular sub-ependymallayer and sub-granular zone of the dentate gyrus | Multipotent | Glia (oligodendrocyte precursor cells) and neurons | [37,38] | ||||
| Hematopoietic stem cells | Multipotent | Lymphoid progenitor cells, hemangioblasts, peripheral blood stem cells, myeloid progenitor cells | [39,40,41,42] | ||||
| Myoblasts | Multipotent | Cardiac, skeletal, smooth muscle cells | [43,44,45] |
| Model | Type of Stem Cell | Injection Method | Volume Concentration | Measured Parameters | Findings | Ref |
|---|---|---|---|---|---|---|
| Pilocarpine induced SE rats | hUC-MSCs | Bilateral intra hippocampus | 105 cells | Hippocampal morphology/inhibitory transmission/epileptic properties | Recovery of hippocampal and GABAergic neurons/Reduced duration and incidence of epilepsy/Maintenance of neuronal circuits’ integrity/Attenuation of MFS and glutamate toxicity/Expression of various cytokines | [58] |
| Pilocarpine-lithium induced SE rats | hUCBC | Tail vein | 1 × 106 cells/rat | Frequency and duration of SRS/hippocampal neuronal densities | Both frequency and duration of SRS were decreased/elevated neuronal densities in the hippocampus | [59] |
| PTZ-induced epileptic rats | hUCB-MSCs | Tail vein | 106 MSCs/rat | Cognitive and motor function/seizure activity/oxidant and antioxidant measurements/GABA level determination | Decreased oxidative stress impairments and cognitive and motor dysfunction/enhanced GABAergic circuits | [60] |
| Lithium- pilocarpine induced SE rats | hUCB-MSCs | Intra right hippocampus | 5 × 105/2 µL | Hippocampal volume/inflammatory changes/hippocampal glucose metabolism | Elevated hippocampal glucose metabolism/bilaterally migration of cell in hippocampi | [61] |
| Pilocarpine induced SE mice | MSC-EVs | In vitro/In vivo; Intravenous | 50 μg MSC-EVs diluted in 150 μL sterile PBS | Anti-oxidative function/SE activity/neural function and morphology | Antioxidant activity of MSC-EVs by Nrf2 signaling/restore structural alterations and neuronal dysfunction/learning, memory and SE improvement | [62] |
| Human model of drug-resistant epilepsy | Autologous BMSCs Neuro-induced MSCs | Intravenous | 2–5 × 106 cells/mL 2.7–8 × 106 cells/mL | Epileptic activity/cognitive function | Cognitive improvements/immunoregulatory effects | [63] |
| Lithium- pilocarpine induced SE rat | Autologous BMSCs | Intravenous | 1.0 × 106 cells/mL | Seizure frequency/ cognitive function | Reduced neuronal cell death/Inhibition of aberrant MFS/Cognitive function improvement/Attenuation of epileptogenesis | [64] |
| Kainic acid induced SE rat | BMSCs | Bilateral intraventral hippocampus | 20,000 cell in 1.5 μL PBS | Epileptic activity | MSC transplant did not change the duration and severity of seizure | [65] |
| Kindling epilepsy rats | Autologous BMSCs | Unilateral intrahippocampal | 2 μL | Adenosine receptors expression | Bidirectional change in adenosine receptors/Maintenance of the balance of adenosine receptors | [66] |
| Kindling epilepsy rats | ESCs | Lateral brain ventricles | - | Seizure activity | Prevention of seizure activity | [67] |
| Kainic acid induced epileptic mouse | hMSCs | Infrahippocampal | 2.5 μL | Seizure activity | hMSCs worked as a vehicle to deliver adenosine/ Improvement of seizure activity | [68] |
| Kainic acid induced epileptic rats | hMSCs | Left hippocampus | 5 μL | Brain injury/ hippocampal morphology/epileptic activity | Reduced seizure duration and hippocampal neural loss/Amelioration of seizures/Reduced apoptosis | [69] |
| Pilocarpine induced epileptic rats | BMSCs | Intravenous | 500 mL | Histological and morphological analysis/ | Functional and structural improvements of hippocampus | [70] |
| Lithium-induced SE rats | BMCs | Intravenous | 1 × 107 cells/mL | Epileptic activity/morphological change/LTP | Lower duration and frequency/ suppressed seizure/Decreased neural lose/Prevention of spontaneous seizures/Reduction of cell loss and hippocampal change/Improvement of LTP formation | [71] |
| Pilocarpine induced epileptic rats | BMCs | Intravenous | 1 × 107 cells/mL in a volume of 100 μL | Seizure activity/ inflammatory and anti-inflammatory cytokines/ | Seizure duration and frequency reduction/Decreased pro-inflammatory cytokines/Elevation of anti-inflammatory cytokine | [72] |
| Lithium chloride pilocarpine induced epileptic rats | BMSCs | Right lateral ventricle | 5 × 106 cells | GABAergic transmission/epileptic activity | Hes1 silencing caused BMSCs to differentiate into GABAergic neuron | [73] |
| Pilocarpine induced epileptic rats | BMSCs | lateral ventricle | 5 × 106 cells | Epileptic activity/cell migration and differentiation | Rate of mortality reduction/SRS frequency and epileptic activity decreased/GABAergic neurons increased | [74] |
| Pilocarpine induced SRS rats | BMMCs | Tail vein | 1 × 107 cells | Expression levels of GDNF, NGF, BDNF, TGF-β1, and VEGF, and their receptors | Upregulation of trophic and growth factors/Neuroprotective effects | [75] |
| PTZ-induced epileptic seizures rats | BMSCs | Intravenous | 3 × 106 cells/rat | The level of excitatory and inhibitory neurotransmitters/oxidative and anti-oxidative factors/HPA axis hormones | Reduction in epileptic activity by balancing between inhibitory and excitatory neurotransmitters/ Anti-oxidant activity/Modulation of sex hormonal profile and inflammatory response | [76] |
| Pilocarpine induced epileptic rats | BMSCs | Intravenous | 2 × 107 cells | Excitatory and inhibitory neurotransmitters concentration/oxidation and anti-oxidation activity/immunomodulatory factors | Reduction in oxidative functions and lipid peroxidation/Downregulation of inflammatory cytokines/Upregulation of anti-inflammatory cytokines and IGF-R signaling/Reduced excitation in hippocampus | [77] |
| Pilocarpine induced epileptic rats | BMMCs | Intravenous | 1 × 107 cells | Cytokine production/epileptic activity | Reduced inflammatory cytokines/Increased anti-inflammatory cytokines | [78] |
| Pilocarpine induced epileptic rats | BM-MSCs AD-MSCs | Tail vein | 100 µL | HSP-70, S100β, and caspase-8 levels/TLR-4/BBB integrity | Decline in the level of S100β, HSP-70, and caspase-8, and TLR-4 gene expression/Neuroprotection/Reduced neural loss | [79] |
| Temporal lobe epilepsy patients | BMMC | Intra-arterial | 15 mL 1.52 × 108 to 10 × 108 cells | MRI/EEG/verbal and nonverbal memory | Better memory performance/Decreased epileptic spikes | [80] |
| Lithium–pilocarpine induced-SE rats | BMMCs | Intravenous | 1×107 cells in 200 μL | SRS activity/cognitive function | Decline in frequency of seizures/Improved long-term spatial memory and learning | [81] |
| Kainic acid induced epileptic rat | ADSC | Intra left hippocampus | 50,000 cells | Neuronal cell markers/learning and memory/neurotrophins/apoptotic and anti-apoptotic factors | Elevated anti-apoptotic and neuronal cell marker expression/Learning and memory improvement | [82] |
| Pilocarpine induced epileptic mice | ADSC | Intraperitoneal | 40 mg/kg | BBB leakage/SRS/cognitive functions | Attenuation of seizure spikes/Reduced BBB leakage | [83] |
| Convulsive seizure induction by maximum electroshock, mice model of seizure | MCAT | Intrahippocampal | 1 × 105 cells (50,000 per hemisphere) | Seizure duration and activity/mortality rate/inflammatory and anti-inflammatory markers | Anticonvulsant effects/Downregulation of inflammatory markers | [84] |
| Model | Type of Stem Cell | Injection Method | Volume Concentration | Measured Parameters | Findings | Ref |
|---|---|---|---|---|---|---|
| DS patient- derived induced pluripotent stem cells | hUC-MSCs | In vitro | - | Oxidative stress markers/Ca 2+ levels/Inflammation | Increased anti-oxidative enzymes: GSH, SOD1/2, and GPX and reduced oxidative markers: MDA, Ca 2+ and ROS/Elevated anti-inflammatory factors: TGF-β and IL-10/6 and lowered interleukin-1 and TNF-α | [85] |
| Drug-resistant epilepsy patients | Autologous BMSCs | Intravenous injection followed by intrathecal injection | 1.0–1.5 × 106 cells/kg/f 0.1 × 106 cells/kg, respectively | Epileptic activity/Seizure frequency/Depression and anxiety | Decreased seizure frequency and paroxysmal activity/Reduced depression and anxiety score | [8] |
| Drug-resistant epilepsy patients | BMNCs | Intrathecal BMNCs: 0.5 × 109; intravenous: 0.38 × 109–1.72 × 109 BMMSCs: 18.5 × 106–40 × 106 | - | Cognitive function/ Epileptic seizure | Cognitive and neurological improvement | [86] |
| Autoimmune refractory epilepsy patients | Autologous ADRCs | Intrathecal 3 times every 3 months | 4 mL | Cognitive function/Epileptic effects/Inflammatory and anti-inflammatory markers | Elevated level of anti-inflammatory cytokines/Reduced epileptic activity/Cognitive function improvement | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesiye, M.R.; Gol, M.; Fadardi, M.R.; Kani, S.N.M.; Costa, A.-M.; Ghasemi-Kasman, M.; Biagini, G. Therapeutic Potential of Mesenchymal Stem Cells in the Treatment of Epilepsy and Their Interaction with Antiseizure Medications. Cells 2022, 11, 4129. https://doi.org/10.3390/cells11244129
Tesiye MR, Gol M, Fadardi MR, Kani SNM, Costa A-M, Ghasemi-Kasman M, Biagini G. Therapeutic Potential of Mesenchymal Stem Cells in the Treatment of Epilepsy and Their Interaction with Antiseizure Medications. Cells. 2022; 11(24):4129. https://doi.org/10.3390/cells11244129
Chicago/Turabian StyleTesiye, Maryam Rahimi, Mohammad Gol, Mohammad Rajabi Fadardi, Seyede Nasim Mousavi Kani, Anna-Maria Costa, Maryam Ghasemi-Kasman, and Giuseppe Biagini. 2022. "Therapeutic Potential of Mesenchymal Stem Cells in the Treatment of Epilepsy and Their Interaction with Antiseizure Medications" Cells 11, no. 24: 4129. https://doi.org/10.3390/cells11244129
APA StyleTesiye, M. R., Gol, M., Fadardi, M. R., Kani, S. N. M., Costa, A. -M., Ghasemi-Kasman, M., & Biagini, G. (2022). Therapeutic Potential of Mesenchymal Stem Cells in the Treatment of Epilepsy and Their Interaction with Antiseizure Medications. Cells, 11(24), 4129. https://doi.org/10.3390/cells11244129