

Bone Tissue and the Nervous System: What Do They Have in Common?

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Search Strategy
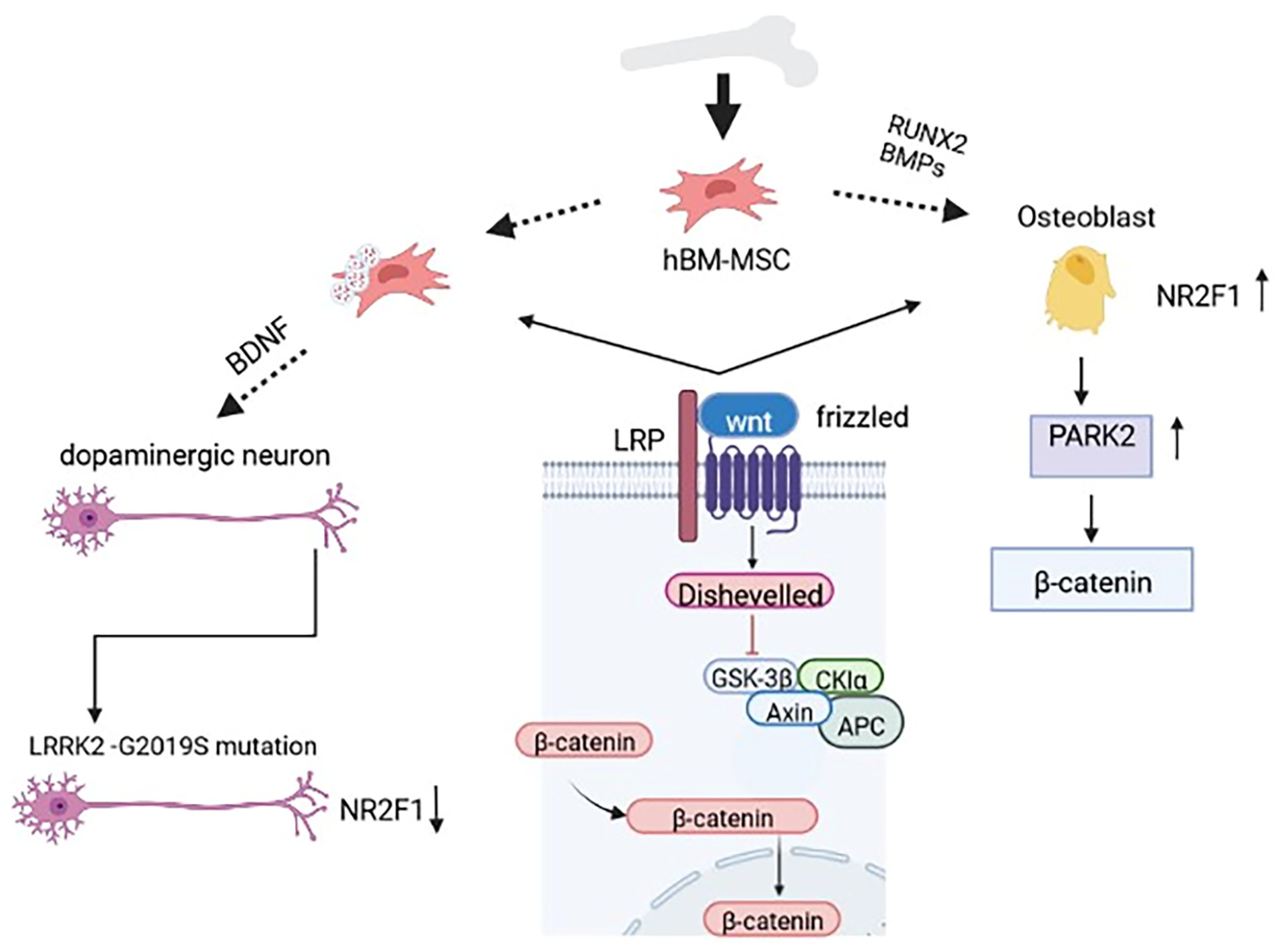
3. Bone Marrow Mesenchymal Stem Cells (BM-MSCs)
4. Crosstalk between Bone and Neural Cells
5. Experimental Models for the Study of Skeletal and Neuronal Cells
5.1. In Vitro Models
5.2. In Vivo Models

{kind=link}
| 2D In Vitro Models | 3D In Vitro Models | In Vivo Models |
|---|---|---|
| CELL CULTURE Cell culture is effective for functional and gene expression studies, as well as for protein analysis. Main cells used: SH-SY5Y, PBMS, MSCs, MLO-Y4, MLC3T3-E1, MNC and RAW264 [95,125]. | Three-dimensional cell culture is useful for detailed research of stem cell behavior, drug development, disease modeling and genetic screening. | Zebrafish and mammals share many skeletal development and biological phenomena. |
6. Bone and Neuronal Cells in Degenerative Diseases
7. Bone-Marrow-Derived Cells Influence Neurogenesis
8. Conclusions and New Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Bruner, E. Geometric morphometrics and paleoneurology: Brain shape evolution in the genus Homo. J. Hum. Evol. 2004, 47, 279–303. [Google Scholar] [CrossRef] [PubMed]
- Gunz, P.; Neubauer, S.; Maureille, B.; Hublin, J.-J. Brain development after birth differs between Neanderthals and modern humans. Curr. Biol. 2010, 20, R921–R922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boeckx, C.A.; Benítez-Burraco, A. The shape of the human language-ready brain. Front. Psychol. 2014, 5, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valenti, M.T.; Dalle Carbonare, L.; Mottes, M. Osteogenic differentiation in healthy and pathological conditions. Int. J. Mol. Sci. 2016, 18, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adugna, D.G.; Aragie, H.; Kibret, A.A.; Belay, D.G. Therapeutic Application of Stem Cells in the Repair of Traumatic Brain Injury. Stem Cells Cloning Adv. Appl. 2022, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhou, J.; Liang, C.; Xiao, Q.; Chen, Y.; Yu, B. Effects of highly selective sensory/motor nerve injury on bone metabolism and bone remodeling in rats. J. Musculoskelet. Neuronal Interact. 2022, 22, 524–535. [Google Scholar]
- Eastell, R.; O’Neill, T.W.; Hofbauer, L.C.; Langdahl, B.; Reid, I.R.; Gold, D.T.; Cummings, S.R. Postmenopausal osteoporosis. Nat. Rev. Dis. Prim. 2016, 2, 16069. [Google Scholar] [CrossRef]
- Catalano, A.; Martino, G.; Bellone, F.; Gaudio, A.; Lasco, C.; Langher, V.; Lasco, A.; Morabito, N. Anxiety levels predict fracture risk in postmenopausal women assessed for osteoporosis. Menopause 2018, 25, 1110–1115. [Google Scholar] [CrossRef]
- Kelly, R.R.; McDonald, L.T.; Jensen, N.R.; Sidles, S.J.; LaRue, A.C. Impacts of psychological stress on osteoporosis: Clinical implications and treatment interactions. Front. Psychiatry 2019, 10, 200. [Google Scholar] [CrossRef]
- Haider, I.T.; Lobos, S.M.; Simonian, N.; Schnitzer, T.J.; Edwards, W.B. Bone fragility after spinal cord injury: Reductions in stiffness and bone mineral at the distal femur and proximal tibia as a function of time. Osteoporos. Int. 2018, 29, 2703–2715. [Google Scholar] [CrossRef]
- Battaglino, R.A.; Lazzari, A.A.; Garshick, E.; Morse, L.R. Spinal cord injury-induced osteoporosis: Pathogenesis and emerging therapies. Curr. Osteoporos. Rep. 2012, 10, 278–285. [Google Scholar] [CrossRef]
- Malochet-Guinamand, S.; Durif, F.; Thomas, T. Parkinson’s disease: A risk factor for osteoporosis. Jt. Bone Spine 2015, 82, 406–410. [Google Scholar] [CrossRef]
- Kang, H.G.; Park, H.Y.; Ryu, H.U.; Suk, S.-H. Bone mineral loss and cognitive impairment: The PRESENT project. Medicine 2018, 97, e12755. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.-E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Prim. 2017, 3, 17013. [Google Scholar] [CrossRef]
- Yuan, J.; Meloni, B.P.; Shi, T.; Bonser, A.; Papadimitriou, J.M.; Mastaglia, F.L.; Zhang, C.; Zheng, M.; Gao, J. The potential influence of bone-derived modulators on the progression of Alzheimer’s disease. J. Alzheimer’s Dis. 2019, 69, 59–70. [Google Scholar] [CrossRef]
- Torsney, K.M.; Noyce, A.J.; Doherty, K.M.; Bestwick, J.P.; Dobson, R.; Lees, A.J. Bone health in Parkinson’s disease: A systematic review and meta-analysis. J. Neurol. Neurosurg. Psychiatry 2014, 85, 1159–1166. [Google Scholar] [CrossRef] [Green Version]
- Huat, T.J.; Camats-Perna, J.; Newcombe, E.A.; Valmas, N.; Kitazawa, M.; Medeiros, R. Metal toxicity links to Alzheimer’s disease and neuroinflammation. J. Mol. Biol. 2019, 431, 1843–1868. [Google Scholar] [CrossRef]
- Dalle Carbonare, L.; Valenti, M.T.; Del Forno, F.; Caneva, E.; Pietrobelli, A. Vitamin D: Daily vs. monthly use in children and elderly—What is going on? Nutrients 2017, 9, 652. [Google Scholar] [CrossRef] [Green Version]
- Moretti, R.; Morelli, M.E.; Caruso, P. Vitamin D in neurological diseases: A rationale for a pathogenic impact. Int. J. Mol. Sci. 2018, 19, 2245. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Zhou, R.; Zhang, Z.; Li, K. The association between vitamin D status, vitamin D supplementation, sunlight exposure, and Parkinson’s disease: A systematic review and meta-analysis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 666. [Google Scholar] [CrossRef]
- Wu, H.; Pang, Q. The effect of vitamin D and calcium supplementation on falls in older adults. Der Orthopäde 2017, 46, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Bahi-Buisson, N.; Nectoux, J.; Rosas-Vargas, H.; Milh, M.; Boddaert, N.; Girard, B.; Cances, C.; Ville, D.; Afenjar, A.; Rio, M. Key clinical features to identify girls with CDKL5 mutations. Brain 2008, 131, 2647–2661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fehr, S.; Wilson, M.; Downs, J.; Williams, S.; Murgia, A.; Sartori, S.; Vecchi, M.; Ho, G.; Polli, R.; Psoni, S. The CDKL5 disorder is an independent clinical entity associated with early-onset encephalopathy. Eur. J. Hum. Genet. 2013, 21, 266–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, T.; Varela, D.; Martins, G.; Conceição, N.; Cancela, M.L. Cdkl5 mutant zebrafish shows skeletal and neuronal alterations mimicking human CDKL5 deficiency disorder. Sci. Rep. 2022, 12, 9325. [Google Scholar] [CrossRef] [PubMed]
- Cheishvili, D.; Parashar, S.; Mahmood, N.; Arakelian, A.; Kremer, R.; Goltzman, D.; Szyf, M.; Rabbani, S.A. Identification of an epigenetic signature of osteoporosis in blood DNA of postmenopausal women. J. Bone Miner. Res. 2018, 33, 1980–1989. [Google Scholar] [CrossRef] [Green Version]
- Pontikoglou, C.; Deschaseaux, F.; Sensebé, L.; Papadaki, H.A. Bone marrow mesenchymal stem cells: Biological properties and their role in hematopoiesis and hematopoietic stem cell transplantation. Stem Cell Rev. Rep. 2011, 7, 569–589. [Google Scholar] [CrossRef]
- Hwang, N.S.; Zhang, C.; Hwang, Y.S.; Varghese, S. Mesenchymal stem cell differentiation and roles in regenerative medicine. Wiley Interdiscip. Rev. Syst. Biol. Med. 2009, 1, 97–106. [Google Scholar] [CrossRef]
- Deng, Q.; Li, P.; Che, M.; Liu, J.; Biswas, S.; Ma, G.; He, L.; Wei, Z.; Zhang, Z.; Yang, Y. Activation of hedgehog signaling in mesenchymal stem cells induces cartilage and bone tumor formation via Wnt/β-Catenin. Elife 2019, 8, e50208. [Google Scholar] [CrossRef]
- Vega, O.A.; Lucero, C.M.; Araya, H.F.; Jerez, S.; Tapia, J.C.; Antonelli, M.; Salazar-Onfray, F.; Las Heras, F.; Thaler, R.; Riester, S.M. Wnt/β-catenin signaling activates expression of the bone-related transcription factor RUNX2 in select human osteosarcoma cell types. J. Cell. Biochem. 2017, 118, 3662–3674. [Google Scholar] [CrossRef]
- Zhao, X.; Tang, L.; Le, T.P.; Nguyen, B.H.; Chen, W.; Zheng, M.; Yamaguchi, H.; Dawson, B.; You, S.; Martinez-Traverso, I.M. Yap and Taz promote osteogenesis and prevent chondrogenesis in neural crest cells in vitro and in vivo. Sci. Signal. 2022, 15, eabn9009. [Google Scholar] [CrossRef]
- Dalle Carbonare, L.; Antoniazzi, F.; Gandini, A.; Orsi, S.; Bertacco, J.; Li Vigni, V.; Minoia, A.; Griggio, F.; Perduca, M.; Mottes, M. Two Novel C-Terminus RUNX2 Mutations in Two Cleidocranial Dysplasia (CCD) Patients Impairing p53 Expression. Int. J. Mol. Sci. 2021, 22, 10336. [Google Scholar] [CrossRef]
- Valenti, M.T.; Mottes, M.; Cheri, S.; Deiana, M.; Micheletti, V.; Cosaro, E.; Davì, M.V.; Francia, G.; Dalle Carbonare, L. Runx2 overexpression compromises bone quality in acromegalic patients. Endocr.-Relat. Cancer 2018, 25, 269–277. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B.; Van Wijnen, A.J.; Stein, J.L.; Montecino, M.; Javed, A.; Zaidi, S.K.; Young, D.W.; Choi, J.-Y.; Pockwinse, S.M. Runx2 control of organization, assembly and activity of the regulatory machinery for skeletal gene expression. Oncogene 2004, 23, 4315–4329. [Google Scholar] [CrossRef] [Green Version]
- Depew, M.J.; Liu, J.K.; Long, J.E.; Presley, R.; Meneses, J.J.; Pedersen, R.A.; Rubenstein, J. Dlx5 regulates regional development of the branchial arches and sensory capsules. Development 1999, 126, 3831–3846. [Google Scholar] [CrossRef]
- Boeckx, C.; Benítez-Burraco, A. Osteogenesis and neurogenesis: A robust link also for language evolution. Front. Cell. Neurosci. 2015, 9, 291. [Google Scholar] [CrossRef] [Green Version]
- Pleasure, S.J.; Anderson, S.; Hevner, R.; Bagri, A.; Marin, O.; Lowenstein, D.H.; Rubenstein, J.L. Cell migration from the ganglionic eminences is required for the development of hippocampal GABAergic interneurons. Neuron 2000, 28, 727–740. [Google Scholar] [CrossRef]
- Benes, F.M.; Lim, B.; Matzilevich, D.; Walsh, J.P.; Subburaju, S.; Minns, M. Regulation of the GABA cell phenotype in hippocampus of schizophrenics and bipolars. Proc. Natl. Acad. Sci. USA 2007, 104, 10164–10169. [Google Scholar] [CrossRef] [Green Version]
- Reale, M.E.; Webb, I.C.; Wang, X.; Baltazar, R.M.; Coolen, L.M.; Lehman, M.N. The transcription factor Runx2 is under circadian control in the suprachiasmatic nucleus and functions in the control of rhythmic behavior. PLoS ONE 2013, 8, e54317. [Google Scholar] [CrossRef] [Green Version]
- Talkowski, M.E.; Rosenfeld, J.A.; Blumenthal, I.; Pillalamarri, V.; Chiang, C.; Heilbut, A.; Ernst, C.; Hanscom, C.; Rossin, E.; Lindgren, A.M. Sequencing chromosomal abnormalities reveals neurodevelopmental loci that confer risk across diagnostic boundaries. Cell 2012, 149, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Ruzicka, W.B.; Subburaju, S.; Benes, F.M. Circuit-and diagnosis-specific DNA methylation changes at γ-aminobutyric acid–related genes in postmortem human hippocampus in schizophrenia and bipolar disorder. JAMA Psychiatry 2015, 72, 541–551. [Google Scholar] [CrossRef]
- Jeong, J.H.; Jin, J.S.; Kim, H.N.; Kang, S.M.; Liu, J.C.; Lengner, C.J.; Otto, F.; Mundlos, S.; Stein, J.L.; Van Wijnen, A.J. Expression of Runx2 transcription factor in non-skeletal tissues, sperm and brain. J. Cell. Physiol. 2008, 217, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Subburaju, S.; Benes, F.M. Induction of the GABA cell phenotype: An in vitro model for studying neurodevelopmental disorders. PLoS ONE 2012, 7, e33352. [Google Scholar] [CrossRef] [PubMed]
- Valenti, M.T.; Serafini, P.; Innamorati, G.; Gili, A.; Cheri, S.; Bassi, C.; Dalle Carbonare, L. Runx2 expression: A mesenchymal stem marker for cancer. Oncol. Lett. 2016, 12, 4167–4172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroeter, M.; Zickler, P.; Denhardt, D.T.; Hartung, H.-P.; Jander, S. Increased thalamic neurodegeneration following ischaemic cortical stroke in osteopontin-deficient mice. Brain 2006, 129, 1426–1437. [Google Scholar] [CrossRef] [PubMed]
- Soulet, D.; Rivest, S. Bone-marrow-derived microglia: Myth or reality? Curr. Opin. Pharmacol. 2008, 8, 508–518. [Google Scholar] [CrossRef]
- Cartier, N.; Lewis, C.-A.; Zhang, R.; Rossi, F. The role of microglia in human disease: Therapeutic tool or target? Acta Neuropathol. 2014, 128, 363–380. [Google Scholar] [CrossRef] [Green Version]
- El Khoury, J.; Luster, A.D. Mechanisms of microglia accumulation in Alzheimer’s disease: Therapeutic implications. Trends Pharmacol. Sci. 2008, 29, 626–632. [Google Scholar] [CrossRef]
- Huang, S.; Li, Z.; Liu, Y.; Gao, D.; Zhang, X.; Hao, J.; Yang, F. Neural regulation of bone remodeling: Identifying novel neural molecules and pathways between brain and bone. J. Cell. Physiol. 2019, 234, 5466–5477. [Google Scholar] [CrossRef]
- Rajpar, I.; Tomlinson, R.E. Function of peripheral nerves in the development and healing of tendon and bone. Proc. Semin. Cell Dev. Biol. 2022, 123, 48–56. [Google Scholar] [CrossRef]
- Freese, J.L.; Pino, D.; Pleasure, S.J. Wnt signaling in development and disease. Neurobiol. Dis. 2010, 38, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Kele-Olovsson, J.M. Regulation Of Midbrain Dopaminergic Neuron Development by Wnts, Sfrps And bHLH Proteins; Karolinska Institutet (Sweden): Solna, Sweden, 2007. [Google Scholar]
- Kane, L.A.; Lazarou, M.; Fogel, A.I.; Li, Y.; Yamano, K.; Sarraf, S.A.; Banerjee, S.; Youle, R.J. PINK1 phosphorylates ubiquitin to activate Parkin E3 ubiquitin ligase activity. J. Cell Biol. 2014, 205, 143–153. [Google Scholar] [CrossRef]
- Zhang, W.; Hou, W.; Chen, M.; Chen, E.; Xue, D.; Ye, C.; Li, W.; Pan, Z. Upregulation of parkin accelerates osteoblastic differentiation of bone marrow-derived mesenchymal stem cells and bone regeneration by enhancing autophagy and β-Catenin signaling. Front. Cell Dev. Biol. 2020, 907, 576104. [Google Scholar] [CrossRef]
- Takeda, S.; Elefteriou, F.; Levasseur, R.; Liu, X.; Zhao, L.; Parker, K.L.; Armstrong, D.; Ducy, P.; Karsenty, G. Leptin regulates bone formation via the sympathetic nervous system. Cell 2002, 111, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Elefteriou, F.; Ahn, J.D.; Takeda, S.; Starbuck, M.; Yang, X.; Liu, X.; Kondo, H.; Richards, W.G.; Bannon, T.W.; Noda, M. Leptin regulation of bone resorption by the sympathetic nervous system and CART. Nature 2005, 434, 514–520. [Google Scholar] [CrossRef]
- Sato, S.; Hanada, R.; Kimura, A.; Abe, T.; Matsumoto, T.; Iwasaki, M.; Inose, H.; Ida, T.; Mieda, M.; Takeuchi, Y. Central control of bone remodeling by neuromedin U. Nat. Med. 2007, 13, 1234–1240. [Google Scholar] [CrossRef]
- Hohmann, E.L.; Elde, R.P.; Rysavy, J.A.; Einzig, S.; Gebhard, R.L. Innervation of periosteum and bone by sympathetic vasoactive intestinal peptide-containing nerve fibers. Science 1986, 232, 868–871. [Google Scholar] [CrossRef]
- Wan, Q.Q.; Qin, W.P.; Ma, Y.X.; Shen, M.J.; Li, J.; Zhang, Z.B.; Chen, J.H.; Tay, F.R.; Niu, L.N.; Jiao, K. Crosstalk between bone and nerves within bone. Adv. Sci. 2021, 8, 2003390. [Google Scholar] [CrossRef]
- Zhu, Y.; Ma, Y.; Elefteriou, F. Cortical bone is an extraneuronal site of norepinephrine uptake in adult mice. Bone Rep. 2018, 9, 188–198. [Google Scholar] [CrossRef]
- Robles, H.; Park, S.; Joens, M.S.; Fitzpatrick, J.A.; Craft, C.S.; Scheller, E.L. Characterization of the bone marrow adipocyte niche with three-dimensional electron microscopy. Bone 2019, 118, 89–98. [Google Scholar] [CrossRef]
- Mulcrone, P.L.; Campbell, J.P.; Clément-Demange, L.; Anbinder, A.L.; Merkel, A.R.; Brekken, R.A.; Sterling, J.A.; Elefteriou, F. Skeletal colonization by breast cancer cells is stimulated by an osteoblast and β2AR-dependent neo-angiogenic switch. J. Bone Miner. Res. 2017, 32, 1442–1454. [Google Scholar] [CrossRef]
- Hirai, T.; Tanaka, K.; Togari, A. β-adrenergic receptor signaling regulates Ptgs2 by driving circadian gene expression in osteoblasts. J. Cell Sci. 2014, 127, 3711–3719. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Liang, H.; Huang, B.; Xiang, L.; Wang, T.; Xiong, Y.; Yang, B.; Guo, Y.; Gong, P. Beta-adrenergic signaling affect osteoclastogenesis via osteocytic MLO-Y4 cells’ RANKL production. Biochem. Biophys. Res. Commun. 2017, 488, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Nyman, J.S.; Tao, H.; Moss, H.H.; Yang, X.; Elefteriou, F. β2-Adrenergic receptor signaling in osteoblasts contributes to the catabolic effect of glucocorticoids on bone. Endocrinology 2011, 152, 1412–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Song, Y.; Li, J.; Lei, X.; Zhang, S.; Gao, Y.; Cheng, P.; Liu, B.; Miao, S.; Bi, L. Blockade of adrenergic β-receptor activation through local delivery of propranolol from a 3D collagen/polyvinyl alcohol/hydroxyapatite scaffold promotes bone repair in vivo. Cell Prolif. 2020, 53, e12725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bellier, J.-P.; Kimura, H. Peripheral type of choline acetyltransferase: Biological and evolutionary implications for novel mechanisms in cholinergic system. J. Chem. Neuroanat. 2011, 42, 225–235. [Google Scholar] [CrossRef]
- Bajayo, A.; Bar, A.; Denes, A.; Bachar, M.; Kram, V.; Attar-Namdar, M.; Zallone, A.; Kovács, K.J.; Yirmiya, R.; Bab, I. Skeletal parasympathetic innervation communicates central IL-1 signals regulating bone mass accrual. Proc. Natl. Acad. Sci. USA 2012, 109, 15455–15460. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Abe, T.; Chida, D.; Nakamoto, N.; Hori, N.; Kokabu, S.; Sakata, Y.; Tomaru, Y.; Iwata, T.; Usui, M. Functional role of acetylcholine and the expression of cholinergic receptors and components in osteoblasts. FEBS Lett. 2010, 584, 817–824. [Google Scholar] [CrossRef] [Green Version]
- Brazill, J.M.; Beeve, A.T.; Craft, C.S.; Ivanusic, J.J.; Scheller, E.L. Nerves in bone: Evolving concepts in pain and anabolism. J. Bone Miner. Res. 2019, 34, 1393–1406. [Google Scholar] [CrossRef]
- Oostinga, D.; Steverink, J.G.; van Wijck, A.J.; Verlaan, J.-J. An understanding of bone pain: A narrative review. Bone 2020, 134, 115272. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, H.; Zhang, G.; He, Y.; Zhang, P.; Sun, Z.; Gao, Y.; Tan, Y. Calcitonin gene-related peptide reduces Porphyromonas gingivalis LPS-induced TNF-α release and apoptosis in osteoblasts. Mol. Med. Rep. 2018, 17, 3246–3254. [Google Scholar]
- Heffner, M.A.; Genetos, D.C.; Christiansen, B.A. Bone adaptation to mechanical loading in a mouse model of reduced peripheral sensory nerve function. PLoS ONE 2017, 12, e0187354. [Google Scholar] [CrossRef] [Green Version]
- Kosmidis, S.; Polyzos, A.; Harvey, L.; Youssef, M.; Denny, C.A.; Dranovsky, A.; Kandel, E.R. RbAp48 protein is a critical component of GPR158/OCN signaling and ameliorates age-related memory loss. Cell Rep. 2018, 25, 959–973.e956. [Google Scholar] [CrossRef]
- Bhusal, A.; Rahman, M.H.; Lee, W.-H.; Bae, Y.C.; Lee, I.-K.; Suk, K. Paradoxical role of lipocalin-2 in metabolic disorders and neurological complications. Biochem. Pharmacol. 2019, 169, 113626. [Google Scholar] [CrossRef]
- Laszczyk, A.; Nettles, D.; Pollock, T.; Fox, S.; Garcia, M.; Wang, J.; Quarles, L.; King, G. FGF-23 deficiency impairs hippocampal-dependent cognitive function. eNeuro 2019, 6, e0469. [Google Scholar] [CrossRef]
- Hanada, R.; Leibbrandt, A.; Hanada, T.; Kitaoka, S.; Furuyashiki, T.; Fujihara, H.; Trichereau, J.; Paolino, M.; Qadri, F.; Plehm, R. Central control of fever and female body temperature by RANKL/RANK. Nature 2009, 462, 505–509. [Google Scholar] [CrossRef]
- Zhang, J.; Fujita, Y.; Chang, L.; Pu, Y.; Qu, Y.; Wang, S.; Hashimoto, K. Beneficial effects of anti-RANKL antibody in depression-like phenotype, inflammatory bone markers, and bone mineral density in male susceptible mice after chronic social defeat stress. Behav. Brain Res. 2020, 379, 112397. [Google Scholar] [CrossRef]
- Zhang, D.-D.; Cao, Y.; Mu, J.-Y.; Liu, Y.-M.; Gao, F.; Han, F.; Zhai, F.-F.; Zhou, L.-X.; Ni, J.; Yao, M. Inflammatory biomarkers and cerebral small vessel disease: A community-based cohort study (P3-3.001). AAN Enterprises 2022, 98, 1915. [Google Scholar] [CrossRef]
- Levey, A.I.; Qiu, D.; Zhao, L.; Hu, W.T.; Duong, D.M.; Higginbotham, L.; Dammer, E.B.; Seyfried, N.T.; Wingo, T.S.; Hales, C.M. A phase II study repurposing atomoxetine for neuroprotection in mild cognitive impairment. Brain 2022, 145, 1924–1938. [Google Scholar] [CrossRef]
- Ross, R.D.; Shah, R.C.; Leurgans, S.; Bottiglieri, T.; Wilson, R.S.; Sumner, D.R. Circulating Dkk1 and TRAIL are associated with cognitive decline in community-dwelling, older adults with cognitive concerns. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2018, 73, 1688. [Google Scholar] [CrossRef]
- Sheehan-Rooney, K.; Swartz, M.E.; Lovely, C.B.; Dixon, M.J.; Eberhart, J.K. Bmp and Shh signaling mediate the expression of satb2 in the pharyngeal arches. PLoS ONE 2013, 8, e59533. [Google Scholar] [CrossRef] [Green Version]
- Konopka, G.; Bomar, J.M.; Winden, K.; Coppola, G.; Jonsson, Z.O.; Gao, F.; Peng, S.; Preuss, T.M.; Wohlschlegel, J.A.; Geschwind, D.H. Human-specific transcriptional regulation of CNS development genes by FOXP2. Nature 2009, 462, 213–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.-H.; Xu, Y.-J.; Gao, J.; Yan, S.-G.; Zhao, J.; Tu, Q.; Zhang, J.; Duan, X.-J.; Sommer, C.A.; Mostoslavsky, G. Critical-size calvarial bone defects healing in a mouse model with silk scaffolds and SATB2-modified iPSCs. Biomaterials 2011, 32, 5065–5076. [Google Scholar] [PubMed]
- Miguez, A.; Ducret, S.; Di Meglio, T.; Parras, C.; Hmidan, H.; Haton, C.; Sekizar, S.; Mannioui, A.; Vidal, M.; Kerever, A. Opposing roles for Hoxa2 and Hoxb2 in hindbrain oligodendrocyte patterning. J. Neurosci. 2012, 32, 17172–17185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisen, M.J.; Meglio, T.D.; Pasqualetti, M.; Ducret, S.; Brunet, J.-F.; Chedotal, A.; Rijli, F.M. Hox paralog group 2 genes control the migration of mouse pontine neurons through slit-robo signaling. PLoS Biol. 2008, 6, e142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobreva, G.; Chahrour, M.; Dautzenberg, M.; Chirivella, L.; Kanzler, B.; Fariñas, I.; Karsenty, G.; Grosschedl, R. SATB2 is a multifunctional determinant of craniofacial patterning and osteoblast differentiation. Cell 2006, 125, 971–986. [Google Scholar] [CrossRef] [Green Version]
- Tavella, S.; Bobola, N. Expressing Hoxa2 across the entire endochondral skeleton alters the shape of the skeletal template in a spatially restricted fashion. Differentiation 2010, 79, 194–202. [Google Scholar] [CrossRef]
- Garcez, R.C.; Le Douarin, N.M.; Creuzet, S.E. Combinatorial activity of Six1-2-4 genes in cephalic neural crest cells controls craniofacial and brain development. Cell. Mol. Life Sci. 2014, 71, 2149–2164. [Google Scholar] [CrossRef]
- Zhao, H.; Zhou, W.; Yao, Z.; Wan, Y.; Cao, J.; Zhang, L.; Zhao, J.; Li, H.; Zhou, R.; Li, B. Foxp1/2/4 regulate endochondral ossification as a suppresser complex. Dev. Biol. 2015, 398, 242–254. [Google Scholar]
- MuhChyi, C.; Juliandi, B.; Matsuda, T.; Nakashima, K. Epigenetic regulation of neural stem cell fate during corticogenesis. Int. J. Dev. Neurosci. 2013, 31, 424–433. [Google Scholar] [CrossRef]
- Borrell, V.; Cárdenas, A.; Ciceri, G.; Galcerán, J.; Flames, N.; Pla, R.; Nóbrega-Pereira, S.; García-Frigola, C.; Peregrín, S.; Zhao, Z. Slit/Robo signaling modulates the proliferation of central nervous system progenitors. Neuron 2012, 76, 338–352. [Google Scholar] [CrossRef] [Green Version]
- Long, Q.; Qiu, B.; Wang, K.; Yang, J.; Jia, C.; Xin, W.; Wang, P.; Han, R.; Fei, Z.; Liu, W. Genetically engineered bone marrow mesenchymal stem cells improve functional outcome in a rat model of epilepsy. Brain Res. 2013, 1532, 1–13. [Google Scholar] [CrossRef]
- Sugita, S.; Hosaka, Y.; Okada, K.; Mori, D.; Yano, F.; Kobayashi, H.; Taniguchi, Y.; Mori, Y.; Okuma, T.; Chang, S.H. Transcription factor Hes1 modulates osteoarthritis development in cooperation with calcium/calmodulin-dependent protein kinase 2. Proc. Natl. Acad. Sci. USA 2015, 112, 3080–3085. [Google Scholar] [CrossRef] [Green Version]
- Otto, E.; Knapstein, P.-R.; Jahn, D.; Appelt, J.; Frosch, K.-H.; Tsitsilonis, S.; Keller, J. Crosstalk of brain and bone—Clinical observations and their molecular bases. Int. J. Mol. Sci. 2020, 21, 4946. [Google Scholar] [CrossRef]
- Owen, R.; Reilly, G.C. In vitro models of bone remodelling and associated disorders. Front. Bioeng. Biotechnol. 2018, 6, 134. [Google Scholar] [CrossRef] [Green Version]
- Blesa, J.S.; Przedborski, S. Parkinson’s disease: Animal models and dopaminergic cell vulnerability. Front. Neuroanat 2014, 8, 155. [Google Scholar] [CrossRef] [Green Version]
- Oliveros Anerillas, L.; Kingham, P.J.; Lammi, M.J.; Wiberg, M.; Kelk, P. Three-dimensional osteogenic differentiation of bone marrow mesenchymal stem cells promotes matrix metallopeptidase 13 (MMP13) expression in Type I collagen hydrogels. Int. J. Mol. Sci. 2021, 22, 13594. [Google Scholar] [CrossRef]
- Fatehullah, A.; Tan, S.H.; Barker, N. Organoids as an in vitro model of human development and disease. Nat. Cell Biol. 2016, 18, 246–254. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Kim, Y.H.; Hebisch, M.; Sliwinski, C.; Lee, S.; D’Avanzo, C.; Chen, H.; Hooli, B.; Asselin, C.; Muffat, J. A three-dimensional human neural cell culture model of Alzheimer’s disease. Nature 2014, 515, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Tibbitt, M.W.; Anseth, K.S. Hydrogels as extracellular matrix mimics for 3D cell culture. Biotechnol. Bioeng. 2009, 103, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Amelian, A.; Wasilewska, K.; Megias, D.; Winnicka, K. Application of standard cell cultures and 3D in vitro tissue models as an effective tool in drug design and development. Pharmacol. Rep. 2017, 69, 861–870. [Google Scholar] [CrossRef]
- Bolognin, S.; Fossépré, M.; Qing, X.; Jarazo, J.; Ščančar, J.; Moreno, E.L.; Nickels, S.L.; Wasner, K.; Ouzren, N.; Walter, J. 3D cultures of Parkinson’s disease-specific dopaminergic neurons for high content phenotyping and drug testing. Adv. Sci. 2019, 6, 1800927. [Google Scholar] [CrossRef] [PubMed]
- Oun, A.; Sabogal-Guaqueta, A.M.; Galuh, S.; Alexander, A.; Kortholt, A.; Dolga, A. The multifaceted role of LRRK2 in Parkinson’s disease: From human iPSC to organoids. Neurobiol. Dis. 2022, 173, 105837. [Google Scholar] [CrossRef] [PubMed]
- Fett, M.E.; Pilsl, A.; Paquet, D.; Van Bebber, F.; Haass, C.; Tatzelt, J.; Schmid, B.; Winklhofer, K.F. Parkin is protective against proteotoxic stress in a transgenic zebrafish model. PLoS ONE 2010, 5, e11783. [Google Scholar] [CrossRef] [PubMed]
- Baptista, L.S.; Kronemberger, G.S.; Côrtes, I.; Charelli, L.E.; Matsui, R.A.M.; Palhares, T.N.; Sohier, J.; Rossi, A.M.; Granjeiro, J.M. Adult stem cells spheroids to optimize cell colonization in scaffolds for cartilage and bone tissue engineering. Int. J. Mol. Sci. 2018, 19, 1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, M.N.; Ren, G.; Young, K.; Pina, S.; Reis, R.L.; Oliveira, J.M. Scaffold fabrication technologies and structure/function properties in bone tissue engineering. Adv. Funct. Mater. 2021, 31, 2010609. [Google Scholar] [CrossRef]
- Kargozar, S.; Mozafari, M.; Hamzehlou, S.; Brouki Milan, P.; Kim, H.-W.; Baino, F. Bone tissue engineering using human cells: A comprehensive review on recent trends, current prospects, and recommendations. Appl. Sci. 2019, 9, 174. [Google Scholar]
- Heo, J.H.; Kang, D.; Seo, S.J.; Jin, Y. Engineering the extracellular matrix for organoid culture. Int. J. Stem Cells 2022, 15, 60–69. [Google Scholar] [CrossRef]
- Chen, S.; Chen, X.; Geng, Z.; Su, J. The horizon of bone organoid: A perspective on construction and application. Bioact. Mater. 2022, 18, 15–25. [Google Scholar] [CrossRef]
- Ye, W.; Luo, C.; Li, C.; Huang, J.; Liu, F. Organoids to study immune functions, immunological diseases and immunotherapy. Cancer Lett. 2020, 477, 31–40. [Google Scholar]
- Grebenyuk, S.; Ranga, A. Engineering organoid vascularization. Front. Bioeng. Biotechnol. 2019, 7, 39. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Nikkhah, M.; Annabi, N. Biomaterials, Cells, and Patho-physiology: Building Better Organoids and On-Chip Technologies. Biomaterials 2019, 198, 1–2. [Google Scholar] [CrossRef]
- Torisawa, Y.-s.; Spina, C.S.; Mammoto, T.; Mammoto, A.; Weaver, J.C.; Tat, T.; Collins, J.J.; Ingber, D.E. Bone marrow–on–a–chip replicates hematopoietic niche physiology in vitro. Nat. Methods 2014, 11, 663–669. [Google Scholar] [CrossRef] [Green Version]
- Tonelli, F.; Bek, J.W.; Besio, R.; De Clercq, A.; Leoni, L.; Salmon, P.; Coucke, P.J.; Willaert, A.; Forlino, A. Zebrafish: A resourceful vertebrate model to investigate skeletal disorders. Front. Endocrinol. 2020, 11, 489. [Google Scholar] [CrossRef]
- Finley, M.L.; Kidd, K.A.; Curry, R.A.; Lescord, G.L.; Clayden, M.G.; O’Driscoll, N.J. A comparison of mercury biomagnification through lacustrine food webs supporting brook trout (Salvelinus fontinalis) and other salmonid fishes. Front. Environ. Sci. 2016, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Bergen, D.J.; Kague, E.; Hammond, C.L. Zebrafish as an emerging model for osteoporosis: A primary testing platform for screening new osteo-active compounds. Front. Endocrinol. 2019, 10, 6. [Google Scholar] [CrossRef]
- Flinn, L.; Mortiboys, H.; Volkmann, K.; Köster, R.W.; Ingham, P.W.; Bandmann, O. Complex I deficiency and dopaminergic neuronal cell loss in parkin-deficient zebrafish (Danio rerio). Brain 2009, 132, 1613–1623. [Google Scholar] [CrossRef] [Green Version]
- Razali, K.; Othman, N.; Mohd Nasir, M.H.; Doolaanea, A.A.; Kumar, J.; Ibrahim, W.N.; Mohamed Ibrahim, N.; Mohamed, W.M. The Promise of the zebrafish model for Parkinson’s disease: Today’s science and tomorrow’s treatment. Front. Genet. 2021, 12, 655550. [Google Scholar] [CrossRef]
- Kõks, S.; Dogan, S.; Tuna, B.G.; González-Navarro, H.; Potter, P.; Vandenbroucke, R.E. Mouse models of ageing and their relevance to disease. Mech. Ageing Dev. 2016, 160, 41–53. [Google Scholar] [CrossRef]
- Jilka, R.L. The relevance of mouse models for investigating age-related bone loss in humans. J. Gerontol. Ser. A: Biomed. Sci. Med. Sci. 2013, 68, 1209–1217. [Google Scholar] [CrossRef]
- Klæstrup, I.H.; Just, M.K.; Holm, K.L.; Alstrup, A.K.O.; Romero-Ramos, M.; Borghammer, P.; Van Den Berge, N. Impact of aging on animal models of Parkinson’s disease. Front. Aging Neurosci. 2022, 14, 909273. [Google Scholar] [CrossRef]
- Moore, D.J.; Dawson, T.M. Value of genetic models in understanding the cause and mechanisms of Parkinson’s disease. Curr. Neurol. Neurosci. Rep. 2008, 8, 288–296. [Google Scholar] [PubMed]
- Van der Vlag, M.; Havekes, R.; Heckman, P.R. The contribution of Parkin, PINK1 and DJ-1 genes to selective neuronal degeneration in Parkinson’s disease. Eur. J. Neurosci. 2020, 52, 3256–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, E.E.S.; Ho, P.W.-L.; Liu, H.-F.; Pang, S.Y.-Y.; Leung, C.-T.; Malki, Y.; Choi, Z.Y.-K.; Ramsden, D.B.; Ho, S.-L. LRRK2 mutant knock-in mouse models: Therapeutic relevance in Parkinson’s disease. Transl. Neurodegener. 2022, 11, 10. [Google Scholar] [PubMed]
- Yang, T.-X.; Zhu, Y.-F.; Wang, C.-C.; Yang, J.-Y.; Xue, C.-H.; Huang, Q.-R.; Wang, Y.-M.; Zhang, T.-T. Epa-enriched plasmalogen attenuates the cytotoxic effects of lps-stimulated microglia on the sh-sy5y neuronal cell line. Brain Res. Bull. 2022, 186, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.Á.; Renaud, A.; Gamblin, A.-l.; D’arros, C.; Nedellec, S.; Trichet, V.; Layrolle, P. 3D cell culture and osteogenic differentiation of human bone marrow stromal cells plated onto jet-sprayed or electrospun micro-fiber scaffolds. Biomed. Mater. 2015, 10, 045019. [Google Scholar] [PubMed]
- Noroozi, R.; Shamekhi, M.A.; Mahmoudi, R.; Zolfagharian, A.; Asgari, F.; Mousavizadeh, A.; Bodaghi, M.; Hadi, A.; Haghighipour, N. In vitro static and dynamic cell culture study of novel bone scaffolds based on 3D-printed PLA and cell-laden alginate hydrogel. Biomed. Mater. 2022, 17, 045024. [Google Scholar] [CrossRef]
- Ohori-Morita, Y.; Niibe, K.; Limraksasin, P.; Nattasit, P.; Miao, X.; Yamada, M.; Mabuchi, Y.; Matsuzaki, Y.; Egusa, H. Novel Mesenchymal Stem Cell Spheroids with Enhanced Stem Cell Characteristics and Bone Regeneration Ability. Stem Cells Transl. Med. 2022, 11, 434–449. [Google Scholar]
- Yuan, P.; Zhang, M.; Tong, L.; Morse, T.M.; McDougal, R.A.; Ding, H.; Chan, D.; Cai, Y.; Grutzendler, J. PLD3 affects axonal spheroids and network defects in Alzheimer’s disease. Nature 2022, 612, 328–337. [Google Scholar]
- Rabadan, M.; De La Cruz, E.D.; Rao, S.B.; Chen, Y.; Gong, C.; Crabtree, G.; Xu, B.; Markx, S.; Gogos, J.A.; Yuste, R. An in vitro model of neuronal ensembles. Nat. Commun. 2022, 13, 3340. [Google Scholar] [CrossRef]
- Spitz, S.; Bolognin, S.; Brandauer, K.; Fuessl, J.; Schuller, P.; Schobesberger, S.; Jordan, C.; Schaedl, B.; Grillari, J.; Wanzenboeck, H.D. Development of a multi-sensor integrated midbrain organoid-on-a-chip platform for studying Parkinson’s disease. bioRxiv 2022. [Google Scholar] [CrossRef]
- Shin, N.; Kim, Y.; Ko, J.; Choi, S.W.; Hyung, S.; Lee, S.E.; Park, S.; Song, J.; Jeon, N.L.; Kang, K.S. Vascularization of iNSC spheroid in a 3D spheroid-on-a-chip platform enhances neural maturation. Biotechnol. Bioeng. 2022, 119, 566–574. [Google Scholar] [CrossRef]
- Brighi, C.; Cordella, F.; Chiriatti, L.; Soloperto, A.; Di Angelantonio, S. Retinal and brain organoids: Bridging the gap between in vivo physiology and in vitro micro-physiology for the study of alzheimer’s diseases. Front. Neurosci. 2020, 14, 655. [Google Scholar] [CrossRef]
- Gonzalez, C.; Armijo, E.; Bravo-Alegria, J.; Becerra-Calixto, A.; Mays, C.E.; Soto, C. Modeling amyloid beta and tau pathology in human cerebral organoids. Mol. Psychiatry 2018, 23, 2363–2374. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, T.; Ding, J.; Li, Z. Bone-on-a-chip platforms and integrated biosensors: Towards advanced in vitro bone models with real-time biosensing. Biosens. Bioelectron. 2022, 219, 114798. [Google Scholar] [CrossRef]
- Cha, C. Microfluidic Biotechnology for “Bone-on-a-Chip”. Biofabr. Orthop. Methods Tech. Appl. 2022, 1, 181–209. [Google Scholar]
- Gan, S.; Huang, Z.; Liu, N.; Su, R.; Xie, G.; Zhong, B.; Zhang, K.; Wang, S.; Hu, X.; Zhang, J. Micro RNA-140-5p impairs zebrafish embryonic bone development via targeting BMP-2. FEBS Lett. 2016, 590, 1438–1446. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Martínez, L.; Guerrero-Peña, L.; Suárez-Bregua, P.; Naranjo, S.; Tena, J.J.; Rotllant, J. Neural Regulation of Bone Mineral Homeostasis in Fish: Functional and Transcriptional Characterization of pth4 Neurons. In Proceedings of the 6th International Symposium on Genomics in Aquaculture, Granada, Spain, 4–6 May 2022. [Google Scholar]
- Kawanishi, S.; Takata, K.; Itezono, S.; Nagayama, H.; Konoya, S.; Chisaki, Y.; Toda, Y.; Nakata, S.; Yano, Y.; Kitamura, Y. Bone-marrow-derived microglia-like cells ameliorate brain amyloid pathology and cognitive impairment in a mouse model of Alzheimer’s disease. J. Alzheimer’s Dis. 2018, 64, 563–585. [Google Scholar] [CrossRef]
- LLabre, J.E.; Gil, C.; Amatya, N.; Lagalwar, S.; Possidente, B.; Vashishth, D. Degradation of Bone Quality in a Transgenic Mouse Model of Alzheimer′ s Disease. J. Bone Miner. Res. 2022, 37, 2548–2565. [Google Scholar] [CrossRef]
- Cardoso, A.L.; Fernandes, A.; Aguilar-Pimentel, J.A.; de Angelis, M.H.; Guedes, J.R.; Brito, M.A.; Ortolano, S.; Pani, G.; Athanasopoulou, S.; Gonos, E.S. Towards frailty biomarkers: Candidates from genes and pathways regulated in aging and age-related diseases. Ageing Res. Rev. 2018, 47, 214–277. [Google Scholar] [CrossRef]
- Tsai, Y.-L.; Yen, C.-T.; Wang, Y.-F. Astrocyte Dysregulation and Calcium Ion Imbalance May Link the Development of Osteoporosis and Alzheimer’s Disease. J. Alzheimer’s Dis. 2022, 88, 439–445. [Google Scholar] [CrossRef]
- Li, S.; Yang, B.; Teguh, D.; Zhou, L.; Xu, J.; Rong, L. Amyloid β Peptide Enhances RANKL-Induced Osteoclast Activation through NF-κB, ERK, and Calcium Oscillation Signaling. Int. J. Mol. Sci. 2016, 17, 1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, B.-W.; Guo, J.-D.; Wu, M.-S.; Liu, Y.; Lu, M.; Zhou, Y.-H.; Han, H.-W. Osteoblast-derived lipocalin-2 regulated by miRNA-96-5p/Foxo1 advances the progression of Alzheimer’s disease. Epigenomics 2020, 12, 1501–1513. [Google Scholar] [CrossRef] [PubMed]
- Ximerakis, M.; Lipnick, S.L.; Innes, B.T.; Simmons, S.K.; Adiconis, X.; Dionne, D.; Mayweather, B.A.; Nguyen, L.; Niziolek, Z.; Ozek, C. Single-cell transcriptomic profiling of the aging mouse brain. Nat. Neurosci. 2019, 22, 1696–1708. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Wang, Z.; Wang, D.; Wang, Z.; Martens, Y.A.; Wu, L.; Xu, Y.; Wang, K.; Li, J.; Huang, R. Amyloid-beta modulates microglial responses by binding to the triggering receptor expressed on myeloid cells 2 (TREM2). Mol. Neurodegener. 2018, 13, 15. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Lv, Z.-Y.; Li, Y.-X.; Hao, Y.-L. Effects of TYROBP Deficiency on Neuroinflammation of a Alzheimer’s Disease Mouse Model Carrying a PSEN1 p. G378E Mutation. Chin. Med. Sci. J. Chung-Kuo I Hsueh K’o Hsueh Tsa Chih 2022. [Google Scholar] [CrossRef]
- Ulland, T.K.; Colonna, M. TREM2—A key player in microglial biology and Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 667–675. [Google Scholar] [CrossRef]
- Tsukasaki, M.; Takayanagi, H. Osteoimmunology: Evolving concepts in bone–immune interactions in health and disease. Nat. Rev. Immunol. 2019, 19, 626–642. [Google Scholar] [CrossRef]
- Varma, V.; Varma, S.; An, Y.; Hohman, T.; Seddighi, S.; Casanova, R.; Beri, A.; Dammer, E.; Seyfried, N.; Pletnikova, O. Alpha-2 macroglobulin in Alzheimer’s disease: A marker of neuronal injury through the RCAN1 pathway. Mol. Psychiatry 2017, 22, 13–23. [Google Scholar] [CrossRef]
- Xiong, L.; Pan, J.-X.; Guo, H.-h.; Mei, L.; Xiong, W.-C. Parkinson’s in the bone. Cell Biosci. 2021, 11, 190. [Google Scholar] [CrossRef]
- Allen, N.E.; Canning, C.G.; Almeida, L.R.S.; Bloem, B.R.; Keus, S.H.; Löfgren, N.; Nieuwboer, A.; Verheyden, G.S.; Yamato, T.P.; Sherrington, C. Interventions for preventing falls in Parkinson’s disease. Cochrane Database Syst. Rev. 2022, 6, CD011574. [Google Scholar]
- Berwick, D.C.; Harvey, K. The regulation and deregulation of Wnt signaling by PARK genes in health and disease. J. Mol. Cell Biol. 2014, 6, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Inestrosa, N.C.; Arenas, E. Emerging roles of Wnts in the adult nervous system. Nat. Rev. Neurosci. 2010, 11, 77–86. [Google Scholar] [CrossRef]
- L’Episcopo, F.; Tirolo, C.; Testa, N.; Caniglia, S.; Morale, M.C.; Serapide, M.F.; Pluchino, S.; Marchetti, B. Wnt/β-catenin signaling is required to rescue midbrain dopaminergic progenitors and promote neurorepair in ageing mouse model of Parkinson’s disease. Stem Cells 2014, 32, 2147–2163. [Google Scholar] [CrossRef] [Green Version]
- Manikandan, M.; Abuelreich, S.; Elsafadi, M.; Alsalman, H.; Almalak, H.; Siyal, A.; Hashmi, J.A.; Aldahmash, A.; Kassem, M.; Alfayez, M. NR2F1 mediated down-regulation of osteoblast differentiation was rescued by bone morphogenetic protein-2 (BMP-2) in human MSC. Differentiation 2018, 104, 36–41. [Google Scholar] [CrossRef]
- Walter, J.; Bolognin, S.; Poovathingal, S.K.; Magni, S.; Gérard, D.; Antony, P.M.; Nickels, S.L.; Salamanca, L.; Berger, E.; Smits, L.M. The Parkinson’s-disease-associated mutation LRRK2-G2019S alters dopaminergic differentiation dynamics via NR2F1. Cell Rep. 2021, 37, 109864. [Google Scholar] [CrossRef]
- Bisson, E.J.; Finlayson, M.L.; Ekuma, O.; Leslie, W.D.; Marrie, R.A. Multiple sclerosis is associated with low bone mineral density and osteoporosis. Neurol. Clin. Pract. 2019, 9, 391–399. [Google Scholar] [CrossRef]
- Biel, A.; Castanza, A.S.; Rutherford, R.; Fair, S.R.; Chifamba, L.; Wester, J.C.; Hester, M.E.; Hevner, R.F. AUTS2 syndrome: Molecular mechanisms and model systems. Front. Mol. Neurosci. 2022, 15, 858582. [Google Scholar] [CrossRef]
- Eisenberg, E.; Levanon, E.Y. Human housekeeping genes, revisited. TRENDS Genet. 2013, 29, 569–574. [Google Scholar] [CrossRef]
- Beunders, G.; Voorhoeve, E.; Golzio, C.; Pardo, L.M.; Rosenfeld, J.A.; Talkowski, M.E.; Simonic, I.; Lionel, A.C.; Vergult, S.; Pyatt, R.E. Exonic deletions in AUTS2 cause a syndromic form of intellectual disability and suggest a critical role for the C terminus. Am. J. Hum. Genet. 2013, 92, 210–220. [Google Scholar] [CrossRef]
- De Rubeis, S.; He, X.; Goldberg, A.P.; Poultney, C.S.; Samocha, K.; Ercument Cicek, A.; Kou, Y.; Liu, L.; Fromer, M.; Walker, S. Synaptic, transcriptional and chromatin genes disrupted in autism. Nature 2014, 515, 209–215. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.-l.; Lu, H.; Shelly, M.; Gao, H.; Poo, M.-m. Phosphorylation of E3 ligase Smurf1 switches its substrate preference in support of axon development. Neuron 2011, 69, 231–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benitez-Burraco, A.; Boeckx, C. Possible functional links among brain-and skull-related genes selected in modern humans. Front. Psychol. 2015, 6, 794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liedén, A.; Kvarnung, M.; Nilssson, D.; Sahlin, E.; Lundberg, E.S. Intragenic duplication—A novel causative mechanism for SATB2-associated syndrome. Am. J. Med. Genet. Part A 2014, 164, 3083–3087. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Qu, Z.; Tickner, J.; Xu, J.; Dai, K.; Zhang, X. The role of SATB2 in skeletogenesis and human disease. Cytokine Growth Factor Rev. 2014, 25, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.Q.; Gordon, J.A.; Beloti, M.M.; Croce, C.M.; Wijnen, A.J.v.; Stein, J.L.; Stein, G.S.; Lian, J.B. A network connecting Runx2, SATB2, and the miR-23a∼ 27a∼ 24-2 cluster regulates the osteoblast differentiation program. Proc. Natl. Acad. Sci. USA 2010, 107, 19879–19884. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Qian, Y.; Yang, F.; Wang, H.; Yu, Y. Lentiviral-mediated expression of SATB 2 promotes osteogenic differentiation of bone marrow stromal cells in vitro and in vivo. Eur. J. Oral Sci. 2014, 122, 190–197. [Google Scholar] [CrossRef]
- Huang, H.; Tindall, D.J. Dynamic FoxO transcription factors. J. Cell Sci. 2007, 120, 2479–2487. [Google Scholar] [CrossRef] [Green Version]
- Van Bon, B.; Hoischen, A.; Hehir-Kwa, J.; De Brouwer, A.; Ruivenkamp, C.; Gijsbers, A.; Marcelis, C.; De Leeuw, N.; Veltman, J.; Brunner, H. Intragenic deletion in DYRK1A leads to mental retardation and primary microcephaly. Clin. Genet. 2011, 79, 296–299. [Google Scholar] [CrossRef]
- Courcet, J.-B.; Faivre, L.; Malzac, P.; Masurel-Paulet, A.; Lopez, E.; Callier, P.; Lambert, L.; Lemesle, M.; Thevenon, J.; Gigot, N. The DYRK1A gene is a cause of syndromic intellectual disability with severe microcephaly and epilepsy. J. Med. Genet. 2012, 49, 731–736. [Google Scholar] [CrossRef]
- Lee, Y.; Ha, J.; Kim, H.J.; Kim, Y.-S.; Chang, E.-J.; Song, W.-J.; Kim, H.-H. Negative feedback Inhibition of NFATc1 by DYRK1A regulates bone homeostasis. J. Biol. Chem. 2009, 284, 33343–33351. [Google Scholar] [CrossRef] [Green Version]
- Shakibaei, M.; Shayan, P.; Busch, F.; Aldinger, C.; Buhrmann, C.; Lueders, C.; Mobasheri, A. Resveratrol mediated modulation of Sirt-1/Runx2 promotes osteogenic differentiation of mesenchymal stem cells: Potential role of Runx2 deacetylation. PLoS ONE 2012, 7, e35712. [Google Scholar] [CrossRef] [Green Version]
- Iyer, S.; Han, L.; Bartell, S.M.; Kim, H.-N.; Gubrij, I.; de Cabo, R.; O’Brien, C.A.; Manolagas, S.C.; Almeida, M. Sirtuin1 (Sirt1) promotes cortical bone formation by preventing β-catenin sequestration by FoxO transcription factors in osteoblast progenitors. J. Biol. Chem. 2014, 289, 24069–24078. [Google Scholar] [CrossRef] [Green Version]
- Joe, I.-S.; Jeong, S.-G.; Cho, G.-W. Resveratrol-induced SIRT1 activation promotes neuronal differentiation of human bone marrow mesenchymal stem cells. Neurosci. Lett. 2015, 584, 97–102. [Google Scholar] [CrossRef]
- Peng, X.-d.; Xu, P.-Z.; Chen, M.-L.; Hahn-Windgassen, A.; Skeen, J.; Jacobs, J.; Sundararajan, D.; Chen, W.S.; Crawford, S.E.; Coleman, K.G. Dwarfism, impaired skin development, skeletal muscle atrophy, delayed bone development, and impeded adipogenesis in mice lacking Akt1 and Akt2. Genes Dev. 2003, 17, 1352–1365. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, A.; Larson, E.A.; Klein, R.F.; Rotwein, P. Distinct actions of akt1 on skeletal architecture and function. PLoS ONE 2014, 9, e93040. [Google Scholar] [CrossRef] [Green Version]
- Dudek, H.; Datta, S.R.; Franke, T.F.; Birnbaum, M.J.; Yao, R.; Cooper, G.M.; Segal, R.A.; Kaplan, D.R.; Greenberg, M.E. Regulation of neuronal survival by the serine-threonine protein kinase Akt. Science 1997, 275, 661–665. [Google Scholar] [CrossRef]
- Bajwa, N.M.; Kesavan, C.; Mohan, S. Long-term consequences of traumatic brain injury in bone metabolism. Front. Neurol. 2018, 9, 115. [Google Scholar] [CrossRef] [Green Version]
- Jodoin, M.; Rouleau, D.M.; Therrien, E.; Chauny, J.-M.; Sandman, E.; Larson-Dupuis, C.; Leduc, S.; Gosselin, N.; De Beaumont, L. Investigating the incidence and magnitude of heterotopic ossification with and without joints involvement in patients with a limb fracture and mild traumatic brain injury. Bone Rep. 2019, 11, 100222. [Google Scholar] [CrossRef]
- Kalamakis, G.; Brüne, D.; Ravichandran, S.; Bolz, J.; Fan, W.; Ziebell, F.; Stiehl, T.; Catalá-Martinez, F.; Kupke, J.; Zhao, S. Quiescence modulates stem cell maintenance and regenerative capacity in the aging brain. Cell 2019, 176, 1407–1419.e1414. [Google Scholar] [CrossRef]
- Brunet, A.; Goodell, M.A.; Rando, T.A. Ageing and rejuvenation of tissue stem cells and their niches. Nat. Rev. Mol. Cell Biol. 2022, 24, 45–62. [Google Scholar] [CrossRef]
- Baror, R.; Neumann, B.; Segel, M.; Chalut, K.J.; Fancy, S.P.; Schafer, D.P.; Franklin, R.J. Transforming growth factor-beta renders ageing microglia inhibitory to oligodendrocyte generation by CNS progenitors. Glia 2019, 67, 1374–1384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desplats, P.; Lee, H.-J.; Bae, E.-J.; Patrick, C.; Rockenstein, E.; Crews, L.; Spencer, B.; Masliah, E.; Lee, S.-J. Inclusion formation and neuronal cell death through neuron-to-neuron transmission of α-synuclein. Proc. Natl. Acad. Sci. USA 2009, 106, 13010–13015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eglitis, M.A.; Mezey, É. Hematopoietic cells differentiate into both microglia and macroglia in the brains of adult mice. Proc. Natl. Acad. Sci. USA 1997, 94, 4080–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brazelton, T.R.; Rossi, F.M.; Keshet, G.I.; Blau, H.M. From marrow to brain: Expression of neuronal phenotypes in adult mice. Science 2000, 290, 1775–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mezey, E.; Chandross, K.J.; Harta, G.; Maki, R.A.; McKercher, S.R. Turning blood into brain: Cells bearing neuronal antigens generated in vivo from bone marrow. Science 2000, 290, 1779–1782. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-Y.; Yang, L.-P.; Zhao, L. Stem cell therapy for Alzheimer’s disease. World J. Stem Cells 2020, 12, 787. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, J.K.; Lee, H.; Carter, J.E.; Chang, J.W.; Oh, W.; Yang, Y.S.; Suh, J.-G.; Lee, B.-H.; Jin, H.K. Human umbilical cord blood-derived mesenchymal stem cells improve neuropathology and cognitive impairment in an Alzheimer’s disease mouse model through modulation of neuroinflammation. Neurobiol. Aging 2012, 33, 588–602. [Google Scholar] [CrossRef]
- Harach, T.; Jammes, F.; Muller, C.; Duthilleul, N.; Cheatham, V.; Zufferey, V.; Cheatham, D.; Lukasheva, Y.A.; Lasser, T.; Bolmont, T. Administrations of human adult ischemia-tolerant mesenchymal stem cells and factors reduce amyloid beta pathology in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2017, 51, 83–96. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, Z.-f.; Zhang, J.-y. Immunomodulatory role of mesenchymal stem cells in Alzheimer’s disease. Life Sci. 2020, 246, 117405. [Google Scholar] [CrossRef]
- Munoz, J.R.; Stoutenger, B.R.; Robinson, A.P.; Spees, J.L.; Prockop, D.J. Human stem/progenitor cells from bone marrow promote neurogenesis of endogenous neural stem cells in the hippocampus of mice. Proc. Natl. Acad. Sci. USA 2005, 102, 18171–18176, Erratum in Proc. Natl. Acad. Sci. USA 2006, 103, 2000–2002. [Google Scholar] [CrossRef] [Green Version]
- Kan, I.; Barhum, Y.; Melamed, E.; Offen, D. Mesenchymal stem cells stimulate endogenous neurogenesis in the subventricular zone of adult mice. Stem Cell Rev. Rep. 2011, 7, 404–412. [Google Scholar] [CrossRef]
- Segal-Gavish, H.; Karvat, G.; Barak, N.; Barzilay, R.; Ganz, J.; Edry, L.; Aharony, I.; Offen, D.; Kimchi, T. Mesenchymal stem cell transplantation promotes neurogenesis and ameliorates autism related behaviors in BTBR mice. Autism Res. 2016, 9, 17–32. [Google Scholar] [CrossRef]
- Cova, L.; Armentero, M.; Zennaro, E.; Calzarossa, C.; Bossolasco, P.; Busca, G.; Lambertenghi Deliliers, G.; Polli, E.; Nappi, G.; Silani, V.; et al. Multiple neurogenic and neurorescue effects of human mesenchymal stem cell after transplantation in an experimental model of Parkinson’s disease. Brain Res. 2010, 1311, 12–27. [Google Scholar] [CrossRef]
- Bao, X.; Wei, J.; Feng, M.; Lu, S.; Li, G.; Dou, W.; Ma, W.; Ma, S.; An, Y.; Qin, C. Transplantation of human bone marrow-derived mesenchymal stem cells promotes behavioral recovery and endogenous neurogenesis after cerebral ischemia in rats. Brain Res. 2011, 1367, 103–113. [Google Scholar] [CrossRef]
- Volkman, R.; Offen, D. Concise review: Mesenchymal stem cells in neurodegenerative diseases. Stem Cells 2017, 35, 1867–1880. [Google Scholar] [CrossRef] [Green Version]
- Park, H.-J.; Shin, J.Y.; Lee, B.R.; Kim, H.O.; Lee, P.H. Mesenchymal stem cells augment neurogenesis in the subventricular zone and enhance differentiation of neural precursor cells into dopaminergic neurons in the substantia nigra of a parkinsonian model. Cell Transplant. 2012, 21, 1629–1640. [Google Scholar] [CrossRef]
- Sanchez-Ramos, J.; Song, S.; Cardozo-Pelaez, F.; Hazzi, C.; Stedeford, T.; Willing, A.; Freeman, T.; Saporta, S.; Janssen, W.; Patel, N. Adult bone marrow stromal cells differentiate into neural cells in vitro. Exp. Neurol. 2000, 164, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Nakano, M.; Nagaishi, K.; Konari, N.; Saito, Y.; Chikenji, T.; Mizue, Y.; Fujimiya, M. Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by exosome transfer into damaged neurons and astrocytes. Sci. Rep. 2016, 6, 24805. [Google Scholar] [CrossRef] [Green Version]
- Börger, V.; Bremer, M.; Ferrer-Tur, R.; Gockeln, L.; Stambouli, O.; Becic, A.; Giebel, B. Mesenchymal stem/stromal cell-derived extracellular vesicles and their potential as novel immunomodulatory therapeutic agents. Int. J. Mol. Sci. 2017, 18, 1450. [Google Scholar] [CrossRef]
- Xiong, Y.; Mahmood, A.; Chopp, M. Emerging potential of exosomes for treatment of traumatic brain injury. Neural Regen. Res. 2017, 12, 19. [Google Scholar] [CrossRef]
- Yang, Y.; Ye, Y.; Su, X.; He, J.; Bai, W.; He, X. MSCs-derived exosomes and neuroinflammation, neurogenesis and therapy of traumatic brain injury. Front. Cell. Neurosci. 2017, 11, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Ling, Z.; Lu, L.; Zhao, J.; Chen, X.; Xu, P.; Zou, X. Regulatory Roles of Bone in Neurodegenerative Diseases. Front. Aging Neurosci. 2020, 12, 610581. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Li, Y.; Buller, B.; Katakowski, M.; Zhang, Y.; Wang, X.; Shang, X.; Zhang, Z.G.; Chopp, M. Exosome-mediated transfer of miR-133b from multipotent mesenchymal stromal cells to neural cells contributes to neurite outgrowth. Stem Cells 2012, 30, 1556–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Zhang, Z.G.; Lu, M.; Wang, X.; Shang, X.; Elias, S.B.; Chopp, M. MiR-146a promotes remyelination in a cuprizone model of demyelinating injury. Neuroscience 2017, 348, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Nakano, M.; Kobayashi, E.; Mizue, Y.; Chikenji, T.; Otani, M.; Nagaishi, K.; Fujimiya, M. An enriched environment prevents diabetes-induced cognitive impairment in rats by enhancing exosomal miR-146a secretion from endogenous bone marrow-derived mesenchymal stem cells. PLoS ONE 2018, 13, e0204252. [Google Scholar] [CrossRef] [Green Version]
- Yuyama, K.; Sun, H.; Mitsutake, S.; Igarashi, Y. Sphingolipid-modulated exosome secretion promotes clearance of amyloid-β by microglia. J. Biol. Chem. 2012, 287, 10977–10989. [Google Scholar] [CrossRef] [Green Version]
- Vatsa, P.; Negi, R.; Ansari, U.; Khanna, V.; Pant, A. Insights of Extracellular Vesicles of Mesenchymal Stem Cells: A Prospective Cell-Free Regenerative Medicine for Neurodegenerative Disorders. Mol. Neurobiol. 2022, 59, 459–474. [Google Scholar] [CrossRef]
- Barzilay, R.; Kan, I.; Ben-Zur, T.; Bulvik, S.; Melamed, E.; Offen, D. Induction of human mesenchymal stem cells into dopamine-producing cells with different differentiation protocols. Stem Cells Dev. 2008, 17, 547–554. [Google Scholar] [CrossRef]
- Liang, J.; Wu, S.; Zhao, H.; Li, S.-l.; Liu, Z.-x.; Wu, J.; Zhou, L. Human umbilical cord mesenchymal stem cells derived from Wharton’s jelly differentiate into cholinergic-like neurons in vitro. Neurosci. Lett. 2013, 532, 59–63. [Google Scholar] [CrossRef]
- Liu, X.; Li, D.; Jiang, D.; Fang, Y. Acetylcholine secretion by motor neuron-like cells from umbilical cord mesenchymal stem cells. Neural Regen. Res. 2013, 8, 2086. [Google Scholar]
- Brofiga, M.; Massobrio, P. Brain-on-a-Chip: Dream or Reality? Front. Neurosci. 2022, 16, 837623. [Google Scholar] [CrossRef]
- Kaur, A.; Nigam, K.; Tyagi, A.; Dang, S. A Preliminary Pharmacodynamic Study for the Management of Alzheimer’s Disease Using Memantine-Loaded PLGA Nanoparticles. AAPS PharmSciTech 2022, 23, 298. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minoia, A.; Dalle Carbonare, L.; Schwamborn, J.C.; Bolognin, S.; Valenti, M.T. Bone Tissue and the Nervous System: What Do They Have in Common? Cells 2023, 12, 51. https://doi.org/10.3390/cells12010051
Minoia A, Dalle Carbonare L, Schwamborn JC, Bolognin S, Valenti MT. Bone Tissue and the Nervous System: What Do They Have in Common? Cells. 2023; 12(1):51. https://doi.org/10.3390/cells12010051
Chicago/Turabian StyleMinoia, Arianna, Luca Dalle Carbonare, Jens Christian Schwamborn, Silvia Bolognin, and Maria Teresa Valenti. 2023. "Bone Tissue and the Nervous System: What Do They Have in Common?" Cells 12, no. 1: 51. https://doi.org/10.3390/cells12010051
APA StyleMinoia, A., Dalle Carbonare, L., Schwamborn, J. C., Bolognin, S., & Valenti, M. T. (2023). Bone Tissue and the Nervous System: What Do They Have in Common? Cells, 12(1), 51. https://doi.org/10.3390/cells12010051