
The Fairy Chemical Imidazole-4-carboxamide Inhibits the Expression of Axl, PD-L1, and PD-L2 and Improves Response to Cisplatin in Melanoma

 , , ,
, , , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ethical Statement
2.3. Mouse Model of Melanoma and Treatment Schedule
2.4. Apoptosis and MTT Assay
2.5. CT Examination
2.6. Euthanasia and Collection of Samples
2.7. Immunohistochemistry
2.8. Preparation of AHX, AOH, and ICA
2.9. Cell Lines and Reagents
2.10. Cell Culture
2.11. Cell Stimulation
2.12. Evaluation of Gene Expression
2.13. Western Blotting
2.14. Flow Cytometry Analysis
2.15. Statistical Analysis
3. Results
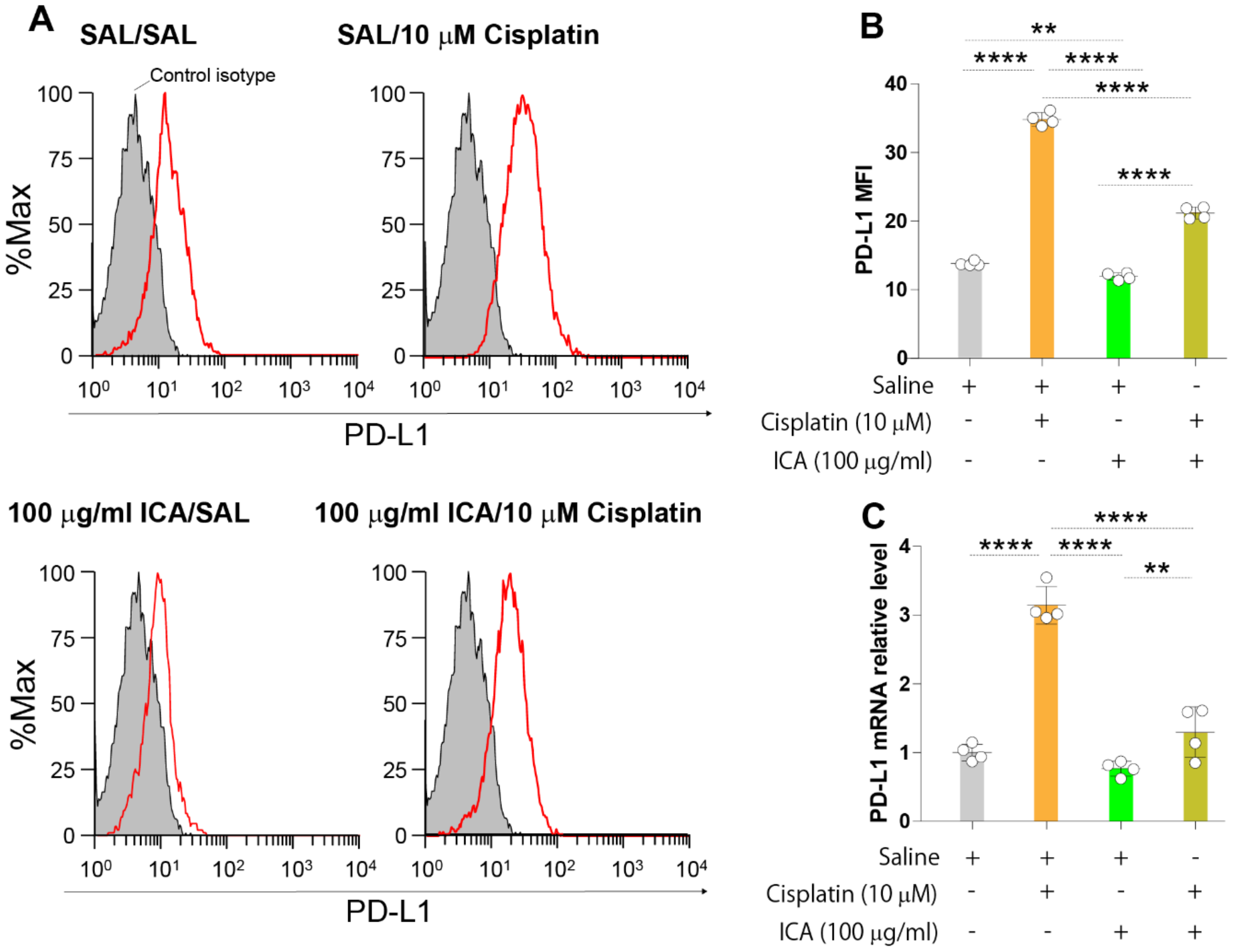
3.1. ICA Decreases PD-L1 Expression Upregulation Induced by Cisplatin in B16F10 Cells
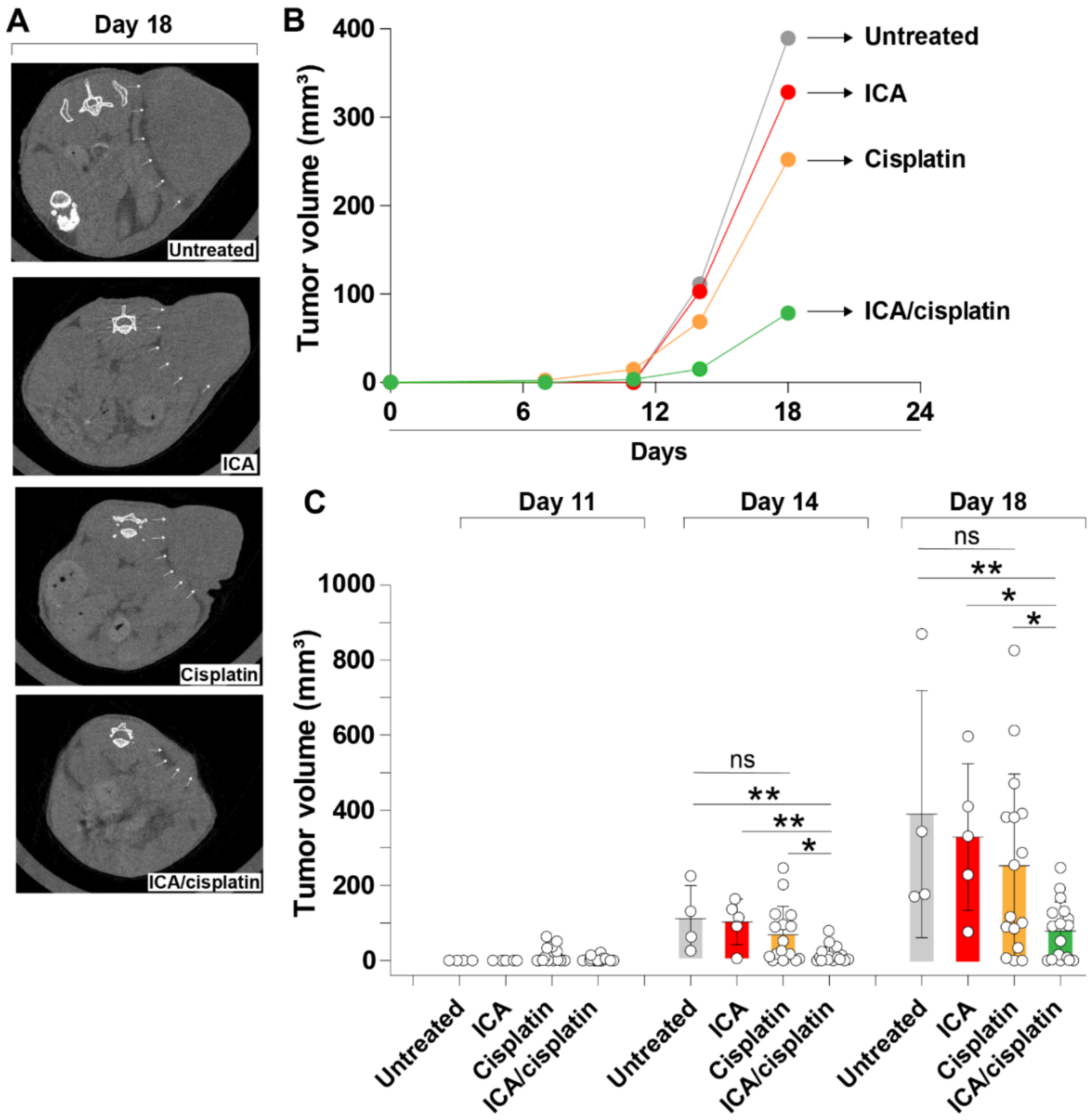
3.2. The Fairy Chemical ICA Inhibits Tumor Growth in a Mouse Xenograft Melanoma Model
3.3. ICA Potentiates Cisplatin Activity
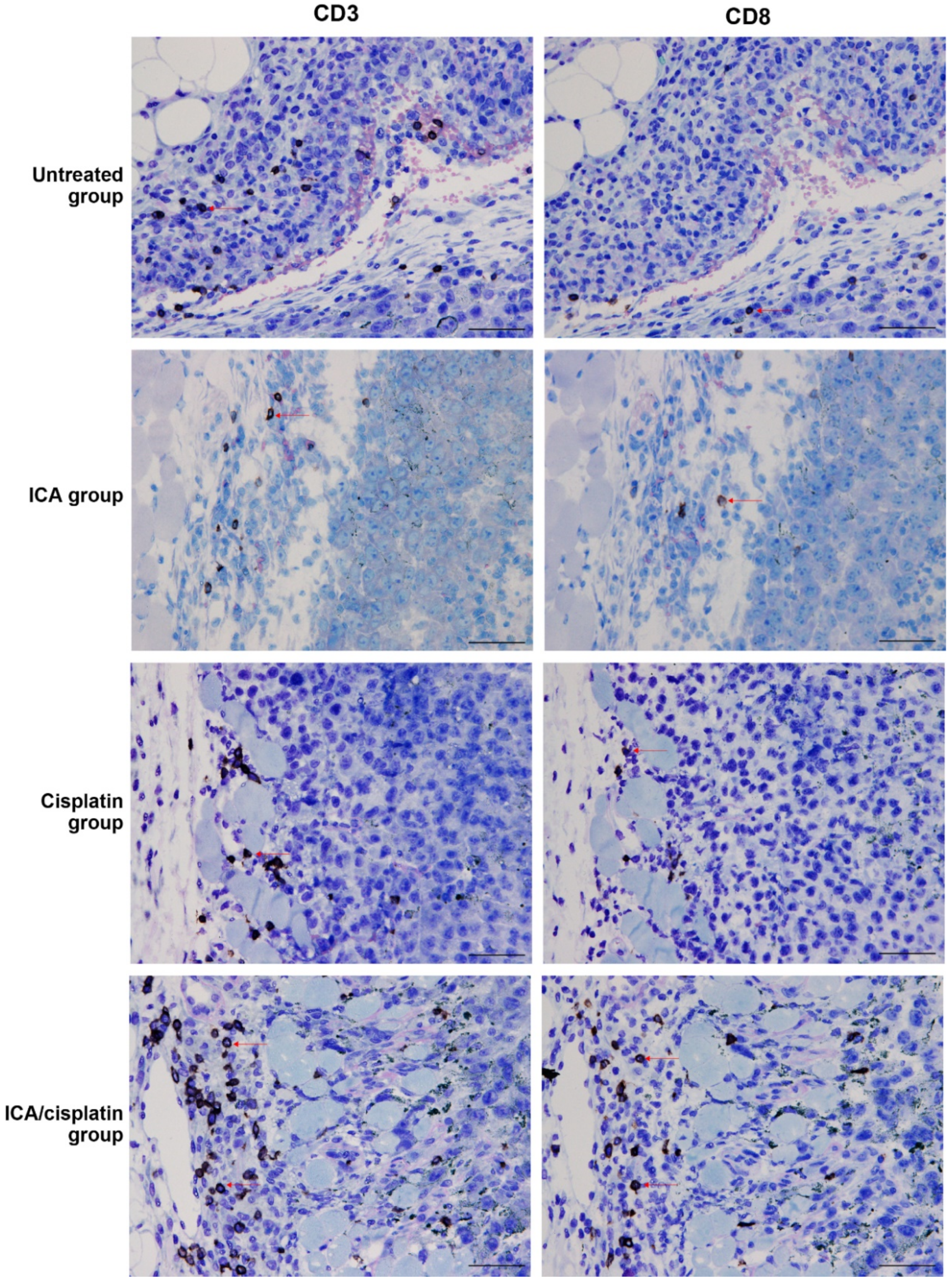
3.4. Enhanced Infiltration of CD8+ Cells in Mice Treated with ICA and Cisplatin
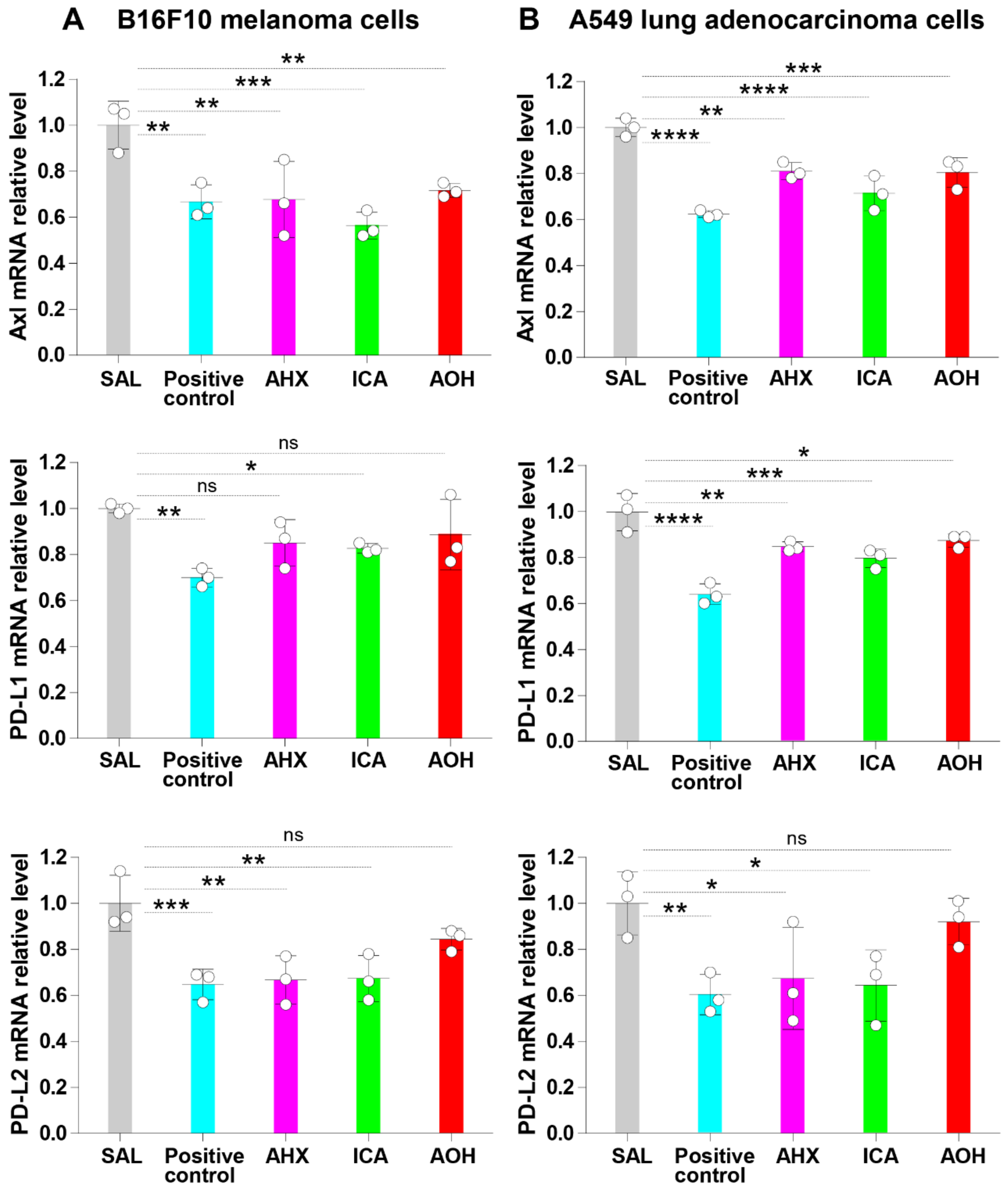
3.5. Fairy Chemicals Inhibit the mRNA Relative Expression of Axl, PD-L1, and PD-L2
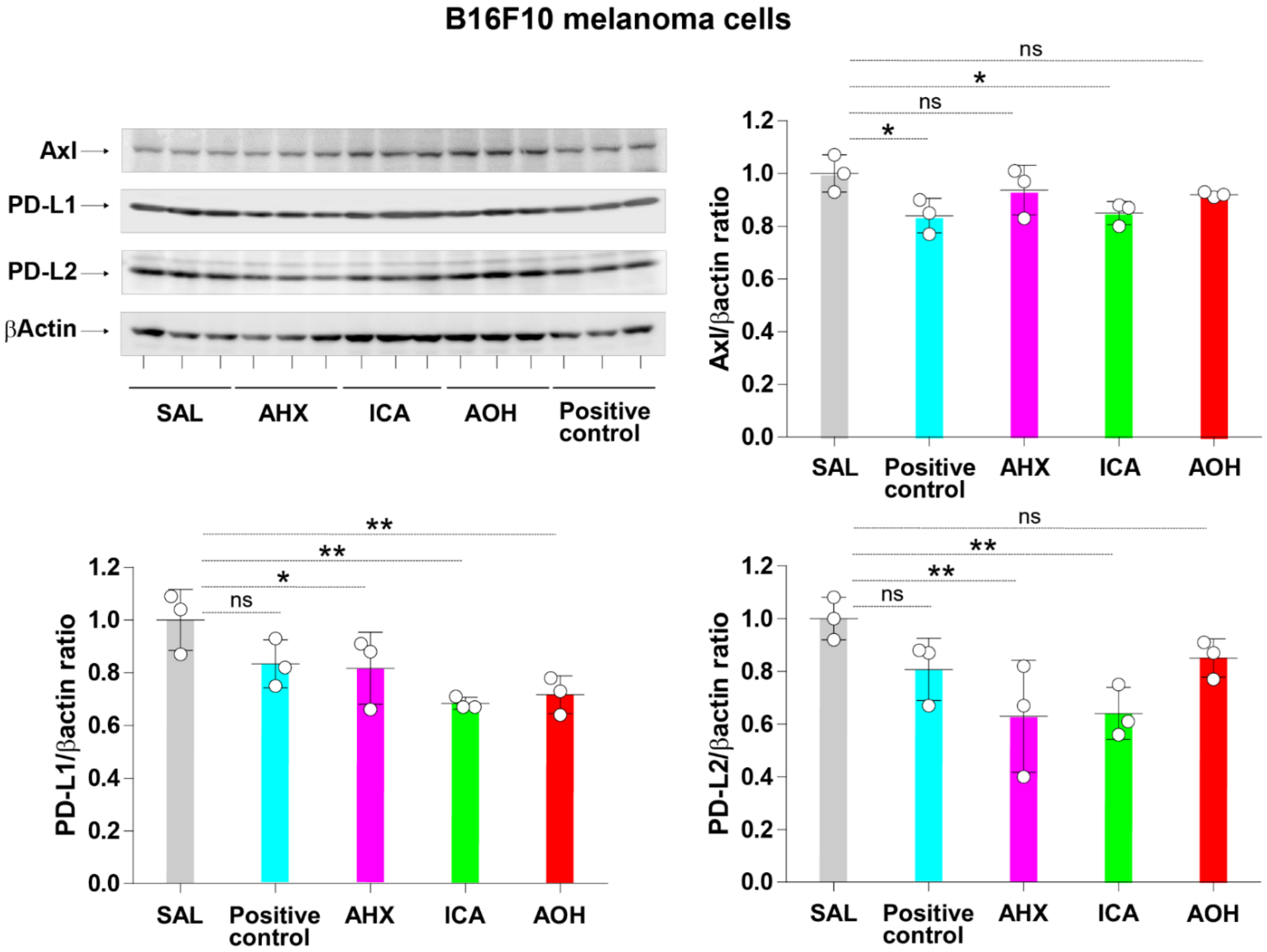
3.6. Inhibition of Axl, PD-L1, and PD-L2 Protein Expression by Fairy Chemicals in Melanoma Cells
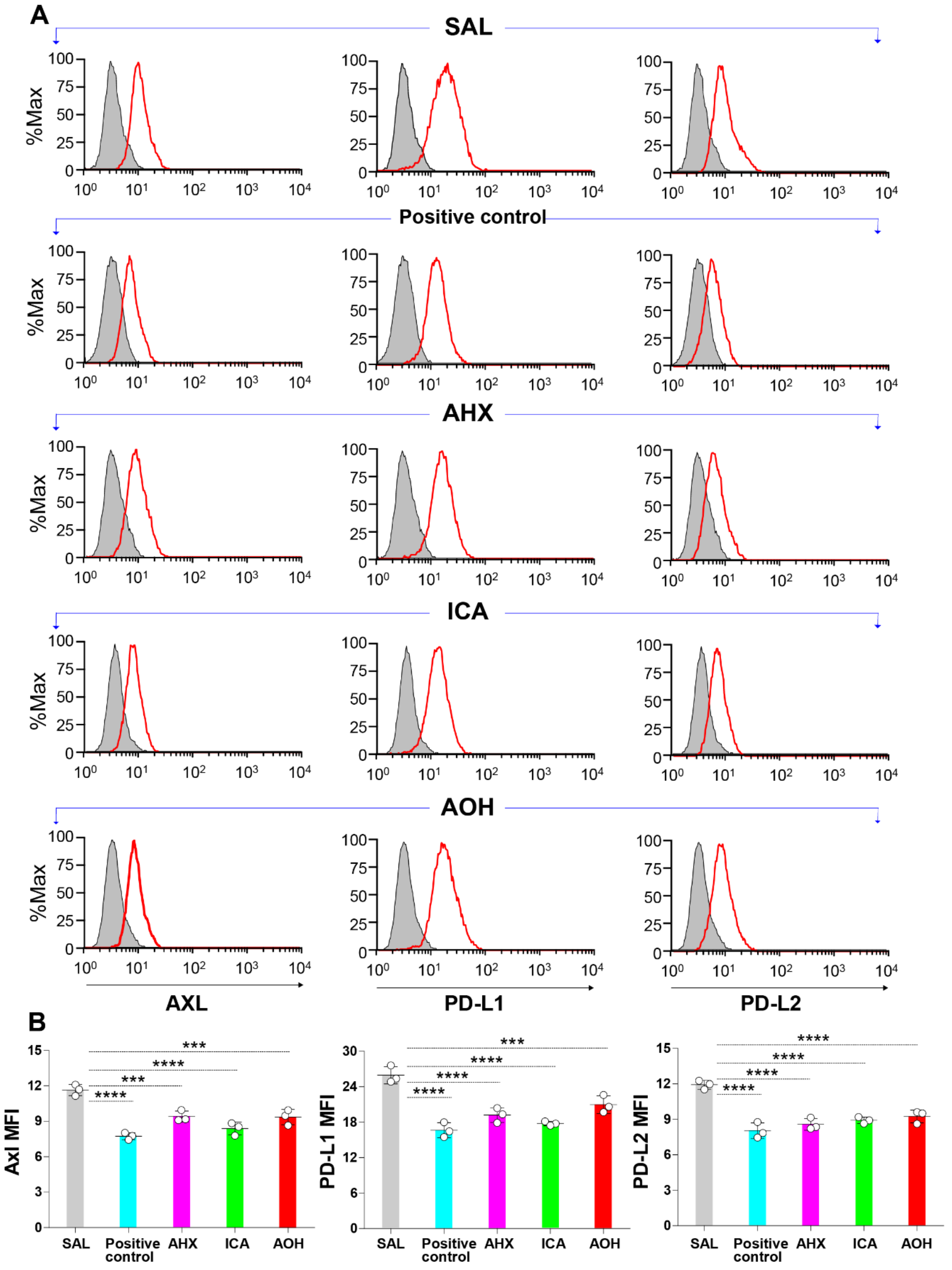
3.7. Significant Inhibition of the Surface Protein Expression of Axl and Immune Checkpoint Molecules by Fairy Chemicals in Melanoma Cells
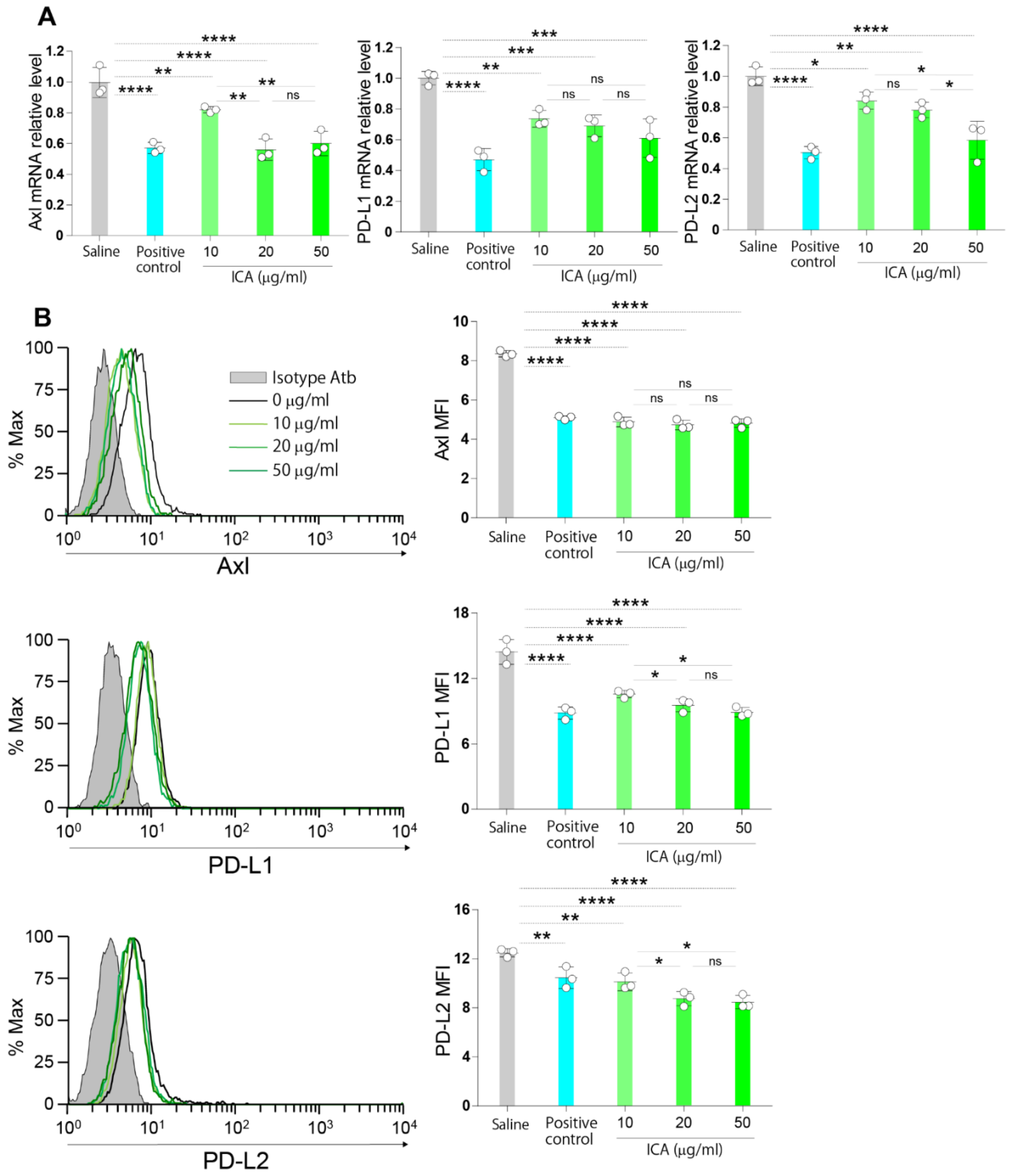
3.8. ICA Dose-Dependently Downregulates the Expression of Axl and Immune Checkpoint Molecules
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Aran, V.; Omerovic, J. Current Approaches in NSCLC Targeting K-RAS and EGFR. Int. J. Mol. Sci. 2019, 20, 5701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brahmer, J.R.; Tykodi, S.S.; Chow, L.Q.M.; Hwu, W.-J.; Topalian, S.L.; Hwu, P.; Drake, C.G.; Camacho, L.H.; Kauh, J.; Odunsi, K.; et al. Safety and Activity of Anti-PD-L1 Antibody in Patients with Advanced Cancer. N. Engl. J. Med. 2012, 366, 2455–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, S.; Camidge, D.R.; Shaw, A.T.; Gadgeel, S.; Ahn, J.S.; Kim, D.W.; Ou, S.-H.I.; Pérol, M.; Dziadziuszko, R.; Rosell, R.; et al. Alectinib versus Crizotinib in Untreated ALK-Positive Non–Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 377, 829–838. [Google Scholar] [CrossRef]
- Titov, A.; Zmievskaya, E.; Ganeeva, I.; Valiullina, A.; Petukhov, A.; Rakhmatullina, A.; Miftakhova, R.; Fainshtein, M.; Rizvanov, A.; Bulatov, E. Adoptive Immunotherapy beyond CAR T-Cells. Cancers 2021, 13, 743. [Google Scholar] [CrossRef]
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Carvajal, R.D.; Sosman, J.A.; Atkins, M.B.; et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N. Engl. J. Med. 2012, 366, 2443–2454. [Google Scholar] [CrossRef]
- Chang, E.; Pelosof, L.; Lemery, S.; Gong, Y.; Goldberg, K.B.; Farrell, A.T.; Keegan, P.; Veeraraghavan, J.; Wei, G.; Blumenthal, G.M.; et al. Systematic Review of PD-1/PD-L1 Inhibitors in Oncology: From Personalized Medicine to Public Health. Oncologist 2021, 26, e1786–e1799. [Google Scholar] [CrossRef]
- Murtuza, A.; Bulbul, A.; Shen, J.P.; Keshavarzian, P.; Woodward, B.D.; Lopez-Diaz, F.J.; Lippman, S.M.; Husain, H. Novel Third-Generation EGFR Tyrosine Kinase Inhibitors and Strategies to Overcome Therapeutic Resistance in Lung Cancer. Cancer Res. 2019, 79, 689–698. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wu, X. Study and analysis of antitumor resistance mechanism of PD1/PD-L1 immune checkpoint blocker. Cancer Med. 2020, 9, 8086–8121. [Google Scholar] [CrossRef]
- Hong, J.; Peng, D.; Chen, Z.; Sehdev, V.; Belkhiri, A. ABL Regulation by AXL Promotes Cisplatin Resistance in Esophageal Cancer. Cancer Res. 2012, 73, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Greger, J.; Shi, H.; Liu, Y.; Greshock, J.; Annan, R.; Halsey, W.; Sathe, G.M.; Martin, A.-M.; Gilmer, T.M. Novel Mechanism of Lapatinib Resistance in HER2-Positive Breast Tumor Cells: Activation of AXL. Cancer Res. 2009, 69, 6871–6878. [Google Scholar] [CrossRef] [Green Version]
- Van der Meer, J.H.; Van der Poll, T.; Van ’t Veer, C. TAM receptors, Gas6, and protein S: Roles in inflammation and hemostasis. Blood 2014, 123, 2460–2469. [Google Scholar] [CrossRef]
- Berclaz, G.; Altermatt, H.J.; Rohrbach, V.; Kieffer, I.; Dreher, E.; Andres, A.-C. Estrogen dependent expression of the receptor tyrosine kinase axl in normal and malignant human breast. Ann. Oncol. 2001, 12, 819–824. [Google Scholar] [CrossRef]
- Davra, V.; Kimani, S.G.; Calianese, D.; Birge, R.B. Ligand Activation of TAM Family Receptors-Implications for Tumor Biology and Therapeutic Response. Cancers 2016, 8, 107. [Google Scholar] [CrossRef]
- Gjerdrum, C.; Tiron, C.; Høiby, T.; Stefansson, I.; Haugen, H.; Sandal, T.; Collett, K.; Li, S.; McCormack, E.; Gjertsen, B.T.; et al. Axl is an essential epithelial-to-mesenchymal transition-induced regulator of breast cancer metastasis and patient survival. Proc. Natl. Acad. Sci. USA 2010, 107, 1124–1129. [Google Scholar] [CrossRef] [Green Version]
- Holland, S.J.; Pan, A.; Franci, C.; Hu, Y.; Chang, B.; Li, W.; Duan, M.; Torneros, A.; Yu, J.; Heckrodt, T.J.; et al. R428, a Selective Small Molecule Inhibitor of Axl Kinase, Blocks Tumor Spread and Prolongs Survival in Models of Metastatic Breast Cancer. Cancer Res. 2010, 70, 1544–1554. [Google Scholar] [CrossRef] [Green Version]
- Tsukita, Y.; Fujino, N.; Miyauchi, E.; Saito, R.; Fujishima, F.; Itakura, K.; Kyogoku, Y.; Okutomo, K.; Yamada, M.; Okazaki, T.; et al. Axl kinase drives immune checkpoint and chemokine signalling pathways in lung adenocarcinomas. Mol. Cancer 2019, 18, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Feeney, M.J.; Dwyer, J.; Hasler-Lewis, C.M.; Milner, J.A.; Noakes, M.; Rowe, S.; Wach, M.; Beelman, R.B.; Caldwell, J.; Cantorna, M.T.; et al. Mushrooms and Health Summit proceedings. J. Nutr. 2014, 144, 1128S–1136S. [Google Scholar] [CrossRef] [Green Version]
- Köhler, J.R.; Casadevall, A.; Perfect, J. The Spectrum of Fungi That Infects Humans. Cold Spring Harb. Perspect. Med. 2014, 5, a019273. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, H.E.; Parrent, J.L.; Jackson, J.A.; Moncalvo, J.-M.; Vilgalys, R. Fungal Community Analysis by Large-Scale Sequencing of Environmental Samples. Appl. Environ. Microbiol. 2005, 71, 5544–5550. [Google Scholar] [CrossRef] [Green Version]
- Ramoutsakis, I.A.; Papadakis, C.E.; Helidonis, E.S. Therapeutic Methods Used for Otolaryngological Problems during the Byzantine Period. Ann. Otol. Rhinol. Laryngol. 2002, 111, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Anusiya, G.; Prabu, U.G.; Yamini, N.V.; Sivarajasekar, N.; Rambabu, K.; Bharath, G.; Banat, F. A review of the therapeutic and biological effects of edible and wild mushrooms. Bioengineered 2021, 12, 11239–11268. [Google Scholar] [CrossRef] [PubMed]
- Malya, I.Y.; Wu, J.; Harada, E.; Toda, M.; D’Alessandro-Gabazza, C.N.; Yasuma, T.; Gabazza, E.; Choi, J.-H.; Hirai, H.; Kawagishi, H. Plant growth regulators and Axl and immune checkpoint inhibitors from the edible mushroom Leucopaxillus giganteus. Biosci. Biotechnol. Biochem. 2020, 84, 1332–1338. [Google Scholar] [CrossRef] [PubMed]
- Ridwan, A.Y.; Wu, J.; Harada, E.; D´alessandro-Gabazza, C.N.; Toda, M.; Yasuma, T.; Gabazza, E.; Choi, J.-H.; Hirai, H.; Kawagishi, H. Axl and immune checkpoints inhibitors from fruiting bodies of Pleurocybella porrigens. J. Antibiot. 2020, 73, 733–736. [Google Scholar] [CrossRef]
- Yasuma, T.; Toda, M.; Kobori, H.; Tada, N.; D’Alessandro-Gabazza, C.; Gabazza, E. Subcritical Water Extracts from Agaricus blazei Murrill’s Mycelium Inhibit the Expression of Immune Checkpoint Molecules and Axl Receptor. J. Fungi 2021, 7, 590. [Google Scholar] [CrossRef]
- Kawagishi, H. Fairy chemicals–a candidate for a new family of plant hormones and possibility of practical use in agriculture*. Biosci. Biotechnol. Biochem. 2018, 82, 752–758. [Google Scholar] [CrossRef]
- Kawagishi, H. Are fairy chemicals a new family of plant hormones? Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 29–38. [Google Scholar] [CrossRef] [Green Version]
- Kawagishi, H. Chemical studies on bioactive compounds related to higher fungi. Biosci. Biotechnol. Biochem. 2021, 85, zbaa072. [Google Scholar] [CrossRef]
- Wu, J.; Kawagishi, H. Plant growth regulators from mushrooms. J. Antibiot. 2020, 73, 657–665. [Google Scholar] [CrossRef]
- Choi, J.-H.; Abe, N.; Tanaka, H.; Fushimi, K.; Nishina, Y.; Morita, A.; Kiriiwa, Y.; Motohashi, R.; Hashizume, D.; Koshino, H.; et al. Plant-Growth Regulator, Imidazole-4-Carboxamide, Produced by the Fairy Ring Forming Fungus Lepista sordida. J. Agric. Food Chem. 2010, 58, 9956–9959. [Google Scholar] [CrossRef]
- Choi, J.-H.; Kikuchi, A.; Pumkaeo, P.; Hirai, H.; Tokuyama, S.; Kawagishi, H. Bioconversion of AHX to AOH by resting cells of Burkholderia contaminans CH-1. Biosci. Biotechnol. Biochem. 2016, 80, 2045–2050. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Choi, J.H.; Takemura, H.; Kotajima, M.; Wu, J.; Tokuyama, S.; Hirai, H.; Asakawa, T.; Ouchi, H.; Inai, M.; et al. Biosynthesis of the Fairy Chemicals, 2-Azahypoxanthine and Imidazole-4-carboxamide, in the Fairy Ring-Forming Fungus Lepista sordida. J. Nat. Prod. 2020, 83, 2469–2476. [Google Scholar] [CrossRef]
- Choi, J.H.; Ohnishi, T.; Yamakawa, Y.; Takeda, S.; Sekiguchi, S.; Maruyama, W.; Yamashita, K.; Suzuki, T.; Morita, A.; Ikka, T.; et al. The source of "fairy rings": 2-azahypoxanthine and its metabolite found in a novel purine metabolic pathway in plants. Angew. Chem. Int. Ed. Engl. 2014, 53, 1552–1555. [Google Scholar] [CrossRef]
- Lee, D.; Miwa, Y.; Wu, J.; Shoda, C.; Jeong, H.; Kawagishi, H.; Tsubota, K.; Kurihara, T. A Fairy Chemical Suppresses Retinal Angiogenesis as a HIF Inhibitor. Biomolecules 2020, 10, 1405. [Google Scholar] [CrossRef]
- Mollo, M.R.; Antonini, D.; Cirillo, L.; Missero, C. Research Techniques Made Simple: Skin Carcinogenesis Models: Xenotransplantation Techniques. J. Investig. Dermatol. 2016, 136, e13–e17. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro-Gabazza, C.N.; Kobayashi, T.; Yasuma, T.; Toda, M.; Kim, H.; Fujimoto, H.; Hataji, O.; Takeshita, A.; Nishihama, K.; Okano, T.; et al. A Staphylococcus pro-apoptotic peptide induces acute exacerbation of pulmonary fibrosis. Nat. Commun. 2020, 11, 1539. [Google Scholar] [CrossRef]
- Ikeuchi, K.; Fujii, R.; Sugiyama, S.; Asakawa, T.; Inai, M.; Hamashima, Y.; Choi, J.-H.; Suzuki, T.; Kawagishi, H.; Kan, T. Practical synthesis of natural plant-growth regulator 2-azahypoxanthine, its derivatives, and biotin-labeled probes. Org. Biomol. Chem. 2014, 12, 3813–3815. [Google Scholar] [CrossRef] [Green Version]
- Ouchi, H.; Asakawa, T.; Ikeuchi, K.; Inai, M.; Choi, J.-H.; Kawagishi, H.; Kan, T. Synthesis of double-13C-labeled imidazole derivatives. Tetrahedron Lett. 2018, 59, 3516–3518. [Google Scholar] [CrossRef]
- Deguchi, H.; Takeya, H.; Wada, H.; Gabazza, E.C.; Hayashi, N.; Urano, H.; Suzuki, K. Dilazep, an antiplatelet agent, inhibits tissue factor expression in endothelial cells and monocytes. Blood 1997, 90, 2345–2356. [Google Scholar] [CrossRef] [Green Version]
- Fournel, L.; Wu, Z.; Stadler, N.; Damotte, D.; Lococo, F.; Boulle, G.; Ségal-Bendirdjian, E.; Bobbio, A.; Icard, P.; Trédaniel, J.; et al. Cisplatin increases PD-L1 expression and optimizes immune check-point blockade in non-small cell lung cancer. Cancer Lett. 2019, 464, 5–14. [Google Scholar] [CrossRef]
- Tsai, T.F.; Lin, J.F.; Lin, Y.C.; Chou, K.Y.; Chen, H.E.; Ho, C.Y.; Chen, P.C.; Hwang, T.I. Cisplatin contributes to programmed death-ligand 1 expression in bladder cancer through ERK1/2-AP-1 signaling pathway. Biosci. Rep. 2019, 39, BSR20190362. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Cai, S.; Deng, Y.; Zhang, Z.; Zhou, X.; Su, Y.; Xu, D. PD-1/PD-L1 enhanced cisplatin resistance in gastric cancer through PI3K/AKT mediated P-gp expression. Int. Immunopharmacol. 2021, 94, 107443. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Recent developments in mushrooms as anti-cancer therapeutics: A review. 3 Biotech 2011, 2, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venturella, G.; Ferraro, V.; Cirlincione, F.; Gargano, M.L. Medicinal Mushrooms: Bioactive Compounds, Use, and Clinical Trials. Int. J. Mol. Sci. 2021, 22, 634. [Google Scholar] [CrossRef]
- Zhu, J.; Xu, J.; Jiang, L.; Huang, J.; Yan, J.; Chen, Y.; Yang, Q. Improved antitumor activity of cisplatin combined with Ganoderma lucidum polysaccharides in U14 cervical carcinoma-bearing mice. Kaohsiung J. Med Sci. 2019, 35, 222–229. [Google Scholar] [CrossRef] [Green Version]
- DePeaux, K.; Delgoffe, G.M. Metabolic barriers to cancer immunotherapy. Nat. Rev. Immunol. 2021, 21, 785–797. [Google Scholar] [CrossRef]
- Gargano, M.L.; Van Griensven, L.J.L.D.; Isikhuemhen, O.S.; Lindequist, U.; Venturella, G.; Wasser, S.P.; Zervakis, G. Medicinal mushrooms: Valuable biological resources of high exploitation potential. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2017, 151, 548–565. [Google Scholar] [CrossRef]
- Konno, S.; Chu, K.; Feuer, N.; Phillips, J.; Choudhury, M. Potent Anticancer Effects of Bioactive Mushroom Extracts (Phellinus linteus) on a Variety of Human Cancer Cells. J. Clin. Med. Res. 2015, 7, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shomali, N.; Onar, O.; Karaca, B.; Demirtas, N.; Cihan, A.C.; Akata, I.; Yildirim, O. Antioxidant, Anticancer, Antimicrobial, and Antibiofilm Properties of the Culinary-Medicinal Fairy Ring Mushroom, Marasmius oreades (Agaricomycetes). Int. J. Med. Mushrooms 2019, 21, 571–582. [Google Scholar] [CrossRef]
- Ba, D.M.; Ssentongo, P.; Beelman, R.B.; Muscat, J.; Gao, X.; Richie, J.P. Higher Mushroom Consumption Is Associated with Lower Risk of Cancer: A Systematic Review and Meta-Analysis of Observational Studies. Adv. Nutr. Int. Rev. J. 2021, 12, 1691–1704. [Google Scholar] [CrossRef]
- Delmanto, R.D.; De Lima, P.L.A.; Sugui, M.M.; Da Eira, A.F.; Salvadori, D.M.F.; Speit, G.; Ribeiro, L.R. Antimutagenic effect of Agaricus blazei Murrill mushroom on the genotoxicity induced by cyclophosphamide. Mutat. Res. Toxicol. Environ. Mutagen. 2001, 496, 15–21. [Google Scholar] [CrossRef]
- Kothari, D.; Patel, S.; Kim, S.-K. Anticancer and other therapeutic relevance of mushroom polysaccharides: A holistic appraisal. Biomed. Pharmacother. 2018, 105, 377–394. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Liang, H.; Luo, L. Antitumor polysaccharides from mushrooms: A review on the structural characteristics, antitumor mechanisms and immunomodulating activities. Carbohydr. Res. 2016, 424, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Hafizi, S.; Dahlback, B. Gas6 and protein S. Vitamin K-dependent ligands for the Axl receptor tyrosine kinase subfamily. FEBS J. 2006, 273, 5231–5244. [Google Scholar] [CrossRef]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef]
- Son, H.-Y.; Jeong, H.-K. Immune Evasion Mechanism and AXL. Front. Oncol. 2021, 11, 651–668. [Google Scholar] [CrossRef]
- Wu, X.; Liu, X.; Koul, S.; Lee, C.Y.; Zhang, Z.; Halmos, B. AXL kinase as a novel target for cancer therapy. Oncotarget 2014, 5, 9546–9563. [Google Scholar] [CrossRef] [Green Version]
- Darnell, J.E.; Kerr, I.M.; Stark, G.R. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef] [Green Version]
- Schindler, C.; Levy, D.; Decker, T. JAK-STAT Signaling: From Interferons to Cytokines. J. Biol. Chem. 2007, 282, 20059–20063. [Google Scholar] [CrossRef] [Green Version]
- Ohaegbulam, K.C.; Assal, A.; Lazar-Molnar, E.; Yao, Y.; Zang, X. Human cancer immunotherapy with antibodies to the PD-1 and PD-L1 pathway. Trends Mol. Med. 2014, 21, 24–33. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, C.; Yasuma, T.; D’Alessandro-Gabazza, C.N.; Toda, M.; Fridman D’Alessandro, V.; Inoue, R.; Fujimoto, H.; Kobori, H.; Tharavecharak, S.; Takeshita, A.; et al. The Fairy Chemical Imidazole-4-carboxamide Inhibits the Expression of Axl, PD-L1, and PD-L2 and Improves Response to Cisplatin in Melanoma. Cells 2022, 11, 374. https://doi.org/10.3390/cells11030374
Inoue C, Yasuma T, D’Alessandro-Gabazza CN, Toda M, Fridman D’Alessandro V, Inoue R, Fujimoto H, Kobori H, Tharavecharak S, Takeshita A, et al. The Fairy Chemical Imidazole-4-carboxamide Inhibits the Expression of Axl, PD-L1, and PD-L2 and Improves Response to Cisplatin in Melanoma. Cells. 2022; 11(3):374. https://doi.org/10.3390/cells11030374
Chicago/Turabian StyleInoue, Chisa, Taro Yasuma, Corina N. D’Alessandro-Gabazza, Masaaki Toda, Valeria Fridman D’Alessandro, Ryo Inoue, Hajime Fujimoto, Hajime Kobori, Suphachai Tharavecharak, Atsuro Takeshita, and et al. 2022. "The Fairy Chemical Imidazole-4-carboxamide Inhibits the Expression of Axl, PD-L1, and PD-L2 and Improves Response to Cisplatin in Melanoma" Cells 11, no. 3: 374. https://doi.org/10.3390/cells11030374
APA StyleInoue, C., Yasuma, T., D’Alessandro-Gabazza, C. N., Toda, M., Fridman D’Alessandro, V., Inoue, R., Fujimoto, H., Kobori, H., Tharavecharak, S., Takeshita, A., Nishihama, K., Okano, Y., Wu, J., Kobayashi, T., Yano, Y., Kawagishi, H., & Gabazza, E. C. (2022). The Fairy Chemical Imidazole-4-carboxamide Inhibits the Expression of Axl, PD-L1, and PD-L2 and Improves Response to Cisplatin in Melanoma. Cells, 11(3), 374. https://doi.org/10.3390/cells11030374