
Transcriptional and Post-Transcriptional Regulation of Autophagy


 ,
,
Abstract
:1. Introduction
2. Regulation of Autophagy by Transcriptional and Post-Transcriptional Modifications
2.1. Transcription Factors Regulate Autophagy at RNA Level
2.2. Regulation of Autophagy by Non-Coding RNAs
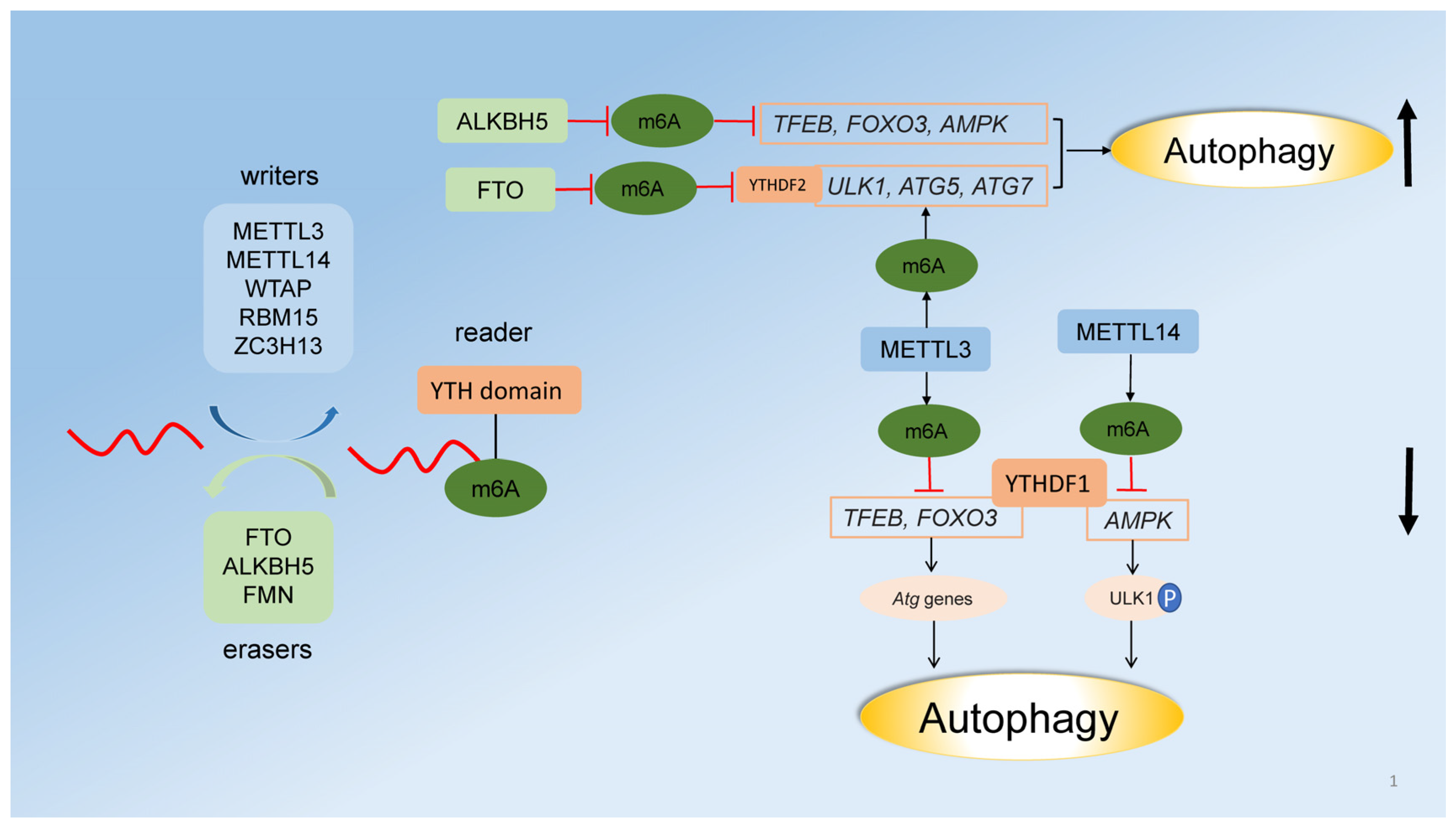
2.3. Regulation of Autophagy by RNA Methylation
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Levine, B.; Kroemer, G. Autophagy in the Pathogenesis of Disease. Cell 2008, 132, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Fullgrabe, J.; Klionsky, D.J.; Joseph, B. The return of the nucleus: Transcriptional and epigenetic control of autophagy. Nat. Rev. Mol. Cell Bio. 2014, 15, 65–74. [Google Scholar] [CrossRef]
- Cardenal-Muñoz, E.; Arafah, S.; López-Jiménez, A.T.; Kicka, S.; Falaise, A.; Bach, F.; Schaad, O.; King, J.S.; Hagedorn, M.; Soldati, T. Mycobacterium marinum antagonistically induces an autophagic response while repressing the autophagic flux in a TORC1- and ESX-1-dependent manner. PLoS Pathog. 2017, 13, e1006344. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Li, K.; Wu, W.; Wu, K.; Yi, H.; Li, W.; Xiao, Y.; Zhong, Y.; Cao, Y.; Tian, L. Steroid hormone 20-hydroxyecdysone induces the transcription and complex assembly of V-ATPases to facilitate autophagy in Bombyx mori. Insect Biochem. Molec. 2020, 116, 103255. [Google Scholar] [CrossRef]
- Luhr, M.; Torgersen, M.L.; Szalai, P.; Hashim, A.; Brech, A.; Staerk, J.; Engedal, N. The kinase PERK and the transcription factor ATF4 play distinct and essential roles in autophagy resulting from tunicamycin-induced ER stress. J. Biol. Chem. 2019, 294, 8197–8217. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Wagner, M.; Xiao, R.; Kim, K.H.; Feng, D.; Lazar, M.A.; Moore, D.D. Nutrient-sensing nuclear receptors coordinate autophagy. Nature 2014, 516, 112–115. [Google Scholar] [CrossRef]
- Rodriguez-Muela, N.; Germain, F.; Marino, G.; Fitze, P.S.; Boya, P. Autophagy promotes survival of retinal ganglion cells after optic nerve axotomy in mice. Cell Death Differ. 2012, 19, 162–169. [Google Scholar] [CrossRef]
- Kong, Y.; Huang, T.; Zhang, H.; Zhang, Q.; Ren, J.; Guo, X.; Fan, H.; Liu, L. The lncRNA NEAT1/miR-29b/Atg9a axis regulates IGFBPrP1-induced autophagy and activation of mouse hepatic stellate cells. Life Sci. 2019, 237, 116902. [Google Scholar] [CrossRef]
- Feng, Y.; Klionsky, D.J. Autophagic membrane delivery through ATG9. Cell Res. 2017, 27, 161–162. [Google Scholar] [CrossRef]
- Conte, A.; Paladino, S.; Bianco, G.; Fasano, D.; Gerlini, R.; Tornincasa, M.; Renna, M.; Fusco, A.; Tramontano, D.; Pierantoni, G.M. High mobility group A1 protein modulates autophagy in cancer cells. Cell Death Differ. 2017, 24, 1948–1962. [Google Scholar] [CrossRef]
- Tian, L.; Ma, L.; Guo, E.; Deng, X.; Ma, S.; Xia, Q.; Cao, Y.; Li, S. 20-Hydroxyecdysone upregulates Atg genes to induce autophagy in the Bombyx fat body. Autophagy 2013, 9, 1172–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Luo, M.; Li, K.; Dai, Y.; Yi, H.; Zhong, Y.; Cao, Y.; Tettamanti, G.; Tian, L. Cholesterol derivatives induce dephosphorylation of the histone deacetylases Rpd3/HDAC1 to upregulate autophagy. Autophagy 2021, 17, 512–528. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, I.; van de Vlekkert, D.; Wolf, E.; Finkelstein, D.; Neale, G.; Machado, E.; Mosca, R.; Campos, Y.; Tillman, H.; Roussel, M.F.; et al. MYC competes with MiT/TFE in regulating lysosomal biogenesis and autophagy through an epigenetic rheostat. Nat. Commun. 2019, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pastore, N.; Vainshtein, A.; Herz, N.J.; Huynh, T.; Brunetti, L.; Klisch, T.J.; Mutarelli, M.; Annunziata, P.; Kinouchi, K.; Brunetti-Pierri, N.; et al. Nutrient-sensitive transcription factors TFEB and TFE3 couple autophagy and metabolism to the peripheral clock. EMBO J. 2019, 38, e101347. [Google Scholar] [CrossRef]
- Settembre, C.; De Cegli, R.; Mansueto, G.; Saha, P.K.; Vetrini, F.; Visvikis, O.; Huynh, T.; Carissimo, A.; Palmer, D.; Klisch, T.J.; et al. TFEB controls cellular lipid metabolism through a starvation-induced autoregulatory loop. Nat. Cell Biol. 2013, 15, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, S.; Goodwin, J.G.; Chauhan, S.; Manyam, G.; Wang, J.; Kamat, A.M.; Boyd, D.D. ZKSCAN3 Is a Master Transcriptional Repressor of Autophagy. Mol. Cell 2013, 50, 16–28. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Brault, J.J.; Schild, A.; Cao, P.; Sandri, M.; Schiaffino, S.; Lecker, S.H.; Goldberg, A.L. FoxO3 coordinately activates protein degradation by the autophagic/lysosomal and proteasomal pathways in atrophying muscle cells. Cell Metab. 2007, 6, 472–483. [Google Scholar] [CrossRef] [Green Version]
- Milan, G.; Romanello, V.; Pescatore, F.; Armani, A.; Paik, J.; Frasson, L.; Seydel, A.; Zhao, J.; Abraham, R.; Goldberg, A.L.; et al. Regulation of autophagy and the ubiquitin–proteasome system by the FOXO transcriptional network during muscle atrophy. Nat. Commun. 2015, 6, 6670. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Zhao, Y. Other Molecular Mechanisms Regulating Autophagy. In Advances in Experimental Medicine and Biology; Qin, Z.H., Ed.; Science Press: Beijing, China, 2019; Volume 1206, pp. 260–270. [Google Scholar]
- Shin, H.R.; Kim, H.; Oh, S.; Lee, J.; Kee, M.; Ko, H.; Kweon, M.; Won, K.; Baek, S.H. AMPK–SKP2–CARM1 signalling cascade in transcriptional regulation of autophagy. Nature 2016, 534, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Kwon, S.; Byun, S.; Xiao, Z.; Park, S.; Wu, S.; Chiang, C.; Kemper, B.; Kemper, J.K. Critical role of RanBP2-mediated SUMOylation of Small Heterodimer Partner in maintaining bile acid homeostasis. Nat. Commun. 2016, 7, 12179. [Google Scholar] [CrossRef] [Green Version]
- Byun, S.; Kim, Y.C.; Zhang, Y.; Kong, B.; Guo, G.; Sadoshima, J.; Ma, J.; Kemper, B.; Kemper, J.K. A postprandial FGF19-SHP-LSD1 regulatory axis mediates epigenetic repression of hepatic autophagy. EMBO J. 2017, 36, 1755–1769. [Google Scholar] [CrossRef]
- Ahmed, M.; Lai, T.H.; Hwang, J.S.; Zada, S.; Pham, T.M.; Kim, D.R. Transcriptional Regulation of Autophagy Genes via Stage-Specific Activation of CEBPB and PPARG during Adipogenesis: A Systematic Study Using Public Gene Expression and Transcription Factor Binding Datasets. Cells 2019, 8, 1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polager, S.; Ofir, M.; Ginsberg, D. E2F1 regulates autophagy and the transcription of autophagy genes. Oncogene 2008, 27, 4860–4864. [Google Scholar] [CrossRef] [Green Version]
- Mu, N.; Lei, Y.; Wang, Y.; Wang, Y.; Duan, Q.; Ma, G.; Liu, X.; Su, L. Inhibition of SIRT1/2 upregulates HSPA5 acetylation and induces pro-survival autophagy via ATF4-DDIT4-mTORC1 axis in human lung cancer cells. Apoptosis 2019, 24, 798–811. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; Li, S. E93 predominantly transduces 20-hydroxyecdysone signaling to induce autophagy and caspase activity in Drosophila fat body. Insect Biochem. Molec. 2014, 45, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Demontis, F.; Perrimon, N. FOXO/4E-BP signaling in Drosophila muscles regulates organism-wide proteostasis during aging. Cell 2010, 143, 813–825. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gordesky-Gold, B.; Leney-Greene, M.; Weinbren, N.L.; Tudor, M.; Cherry, S. Inflammation-Induced, STING-Dependent Autophagy Restricts Zika Virus Infection in the Drosophila Brain. Cell Host Microbe 2018, 24, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Tettamanti, G.; Casartelli, M. Cell death during complete metamorphosis. Philos. Trans. R. Soc. Lond. B 2019, 374, 20190065. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Qian, X.; Lu, Z. Local histone acetylation by ACSS2 promotes gene transcription for lysosomal biogenesis and autophagy. Autophagy 2017, 13, 1790–1791. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Nolan, T.M.; Yin, Y.; Bassham, D.C. Identification of transcription factors that regulate ATG8 expression and autophagy in Arabidopsis. Autophagy 2020, 16, 123–139. [Google Scholar] [CrossRef]
- Lapierre, L.R.; Kumsta, C.; Sandri, M.; Ballabio, A.; Hansen, M. Transcriptional and epigenetic regulation of autophagy in aging. Autophagy 2015, 11, 867–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Huang, J.; Liu, Y.; Li, X.; Zhou, S.; Qian, S.; Liu, Y.; Zhu, H.; Huang, H.; Dang, Y.; et al. Transactivation of Atg4b by C/EBPβ Promotes Autophagy To Facilitate Adipogenesis. Mol. Cell Biol. 2013, 33, 3180–3190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fawcett, T.W.; Martindale, J.L.; Guyton, K.Z.; Hai, T.; Holbrook, N.J. Complexes containing activating transcription factor (ATF)/cAMP-responsive-element-binding protein (CREB) interact with the CCAAT/enhancer-binding protein (C/EBP)-ATF composite site to regulate Gadd153 expression during the stress response. Biochem. J. 1999, 339, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Copetti, T.; Bertoli, C.; Dalla, E.; Demarchi, F.; Schneider, C. p65/RelA ModulatesBECN1 Transcription and Autophagy. Mol. Cell Biol. 2009, 29, 2594–2608. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.; Zhang, H.; Levine, A.J.; Jin, S. The coordinate regulation of the p53 and mTOR pathways in cells. Proc. Natl. Acad. Sci. USA 2005, 102, 8204–8209. [Google Scholar] [CrossRef] [Green Version]
- Tasdemir, E.; Maiuri, M.C.; Galluzzi, L.; Vitale, I.; Djavaheri-Mergny, M.; D’Amelio, M.; Criollo, A.; Morselli, E.; Zhu, C.; Harper, F.; et al. Regulation of autophagy by cytoplasmic p53. Nat. Cell Biol. 2008, 10, 676–687. [Google Scholar] [CrossRef] [Green Version]
- You, L.; Wang, Z.; Li, H.; Shou, J.; Jing, Z.; Xie, J.; Sui, X.; Pan, H.; Han, W. The role of STAT3 in autophagy. Autophagy 2015, 11, 729–739. [Google Scholar] [CrossRef] [Green Version]
- McCormick, J.; Suleman, N.; Scarabelli, T.M.; Knight, R.A.; Latchman, D.S.; Stephanou, A. STAT1 deficiency in the heart protects against myocardial infarction by enhancing autophagy. J. Cell Mol. Med. 2012, 16, 386–393. [Google Scholar] [CrossRef]
- Liu, X.; Dai, F.; Guo, E.; Li, K.; Ma, L.; Tian, L.; Cao, Y.; Zhang, G.; Palli, S.R.; Li, S. 20-Hydroxyecdysone (20E) Primary Response Gene E93 Modulates 20E Signaling to Promote Bombyx Larval-Pupal Metamorphosis. J. Biol. Chem. 2015, 290, 27370–27383. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Yan, Y.; Qian, L.; Gong, Z. Long non-coding RNAs act as regulators of cell autophagy in diseases (Review). Oncol. Rep. 2017, 37, 1359–1366. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Wang, Z.; Zhang, W.; Zhang, L. Non-coding RNAs regulate autophagy process via influencing the expression of associated protein. Prog. Biophys. Mol. Biol. 2020, 151, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Frankel, L.B.; Wen, J.; Lees, M.; Hoyer-Hansen, M.; Farkas, T.; Krogh, A.; Jaattela, M.; Lund, A.H. microRNA-101 is a potent inhibitor of autophagy. EMBO J. 2011, 30, 4628–4641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.; Wen, X.; Bao, J.; Liu, B. MicroRNA-modulated autophagic signaling networks in cancer. Int. J. Biochem. Cell Biol. 2012, 44, 733–736. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, H.; Qi, Y.; Kan, Y.; Ge, Z. Effects of miR-103a-3p on the autophagy and apoptosis of cardiomyocytes by regulating Atg5. Int. J. Mol. Med. 2019, 43, 1951–1960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Wang, X.; Song, M.; He, Z.; Cui, G.; Peng, G.; Dieterich, C.; Antebi, A.; Jing, N.; Shen, Y. A secreted microRNA disrupts autophagy in distinct tissues of Caenorhabditis elegans upon ageing. Nat. Commun. 2019, 10, 4827. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Chen, D.; He, Y.; Meléndez, A.; Feng, Z.; Hong, Q.; Bai, X.; Li, Q.; Cai, G.; Wang, J.; et al. MiR-34 modulates Caenorhabditis elegans lifespan via repressing the autophagy gene atg9. Age 2013, 35, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.M.; Jiang, M.T.; Lin, P.; Yang, H.; Dang, Y.W.; Yu, Q.; Liao, D.Y.; Luo, D.Z.; Chen, G. Potential ceRNA networks involved in autophagy suppression of pancreatic cancer caused by chloroquine diphosphate: A study based on differentially expressed circRNAs, lncRNAs, miRNAs and mRNAs. Int. J. Oncol. 2019, 54, 600–626. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Diling, C.; Yinrui, G.; Longkai, Q.; Xiaocui, T.; Yadi, L.; Xin, Y.; Guoyan, H.; Ou, S.; Tianqiao, Y.; Dongdong, W.; et al. Circular RNA NF1-419 enhances autophagy to ameliorate senile dementia by binding Dynamin-1 and Adaptor protein 2 B1 in AD-like mice. Aging 2019, 11, 12002–12031. [Google Scholar] [CrossRef]
- Li, X.X.; Xiao, L.; Chung, H.K.; Ma, X.X.; Liu, X.; Song, J.L.; Jin, C.Z.; Rao, J.N.; Gorospe, M.; Wang, J.Y. Interaction between HuR and circPABPN1 Modulates Autophagy in the Intestinal Epithelium by Altering ATG16L1 Translation. Mol. Cell Biol. 2020, 40. [Google Scholar] [CrossRef]
- Palanisamy, K.; Tsai, T.H.; Yu, T.M.; Sun, K.T.; Yu, S.H.; Lin, F.Y.; Wang, I.K.; Li, C.Y. RNA-binding protein, human antigen R regulates hypoxia-induced autophagy by targeting ATG7/ATG16L1 expressions and autophagosome formation. J. Cell Physiol. 2019, 234, 7448–7458. [Google Scholar] [CrossRef]
- Xie, Z.Y.; Wang, F.F.; Xiao, Z.H.; Liu, S.F.; Lai, Y.L.; Tang, S.L. Long noncoding RNA XIST enhances ethanol-induced hepatic stellate cells autophagy and activation via miR-29b/HMGB1 axis. Iubmb Life 2019, 71, 1962–1972. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Wang, S.; Jin, L.; Weng, M.; Zhou, D.; Wang, J.; Tang, Z.; Quan, Z. Long non-coding RNA GBCDRlnc1 induces chemoresistance of gallbladder cancer cells by activating autophagy. Mol. Cancer 2019, 18, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Li, X.; Cai, Q.; Zhang, C.; Yu, Q.; Jiang, Y.; Lee, J.; Hawke, D.; Wang, Y.; Xia, Y.; et al. Phosphoglycerate Kinase 1 Phosphorylates Beclin1 to Induce Autophagy. Mol. Cell 2017, 65, 917–931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Wang, P.; Zhang, J.; Wu, H.; Sui, S.; Zhang, J.; Wang, Q.; Qiao, K.; Yang, W.; Xu, H.; et al. Ai-lncRNA EGOT enhancing autophagy sensitizes paclitaxel cytotoxicity via upregulation of ITPR1 expression by RNA-RNA and RNA-protein interactions in human cancer. Mol. Cancer 2019, 18, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, W.; Wang, P.; Li, S. RNA interference of long noncodingRNA HOTAIR suppresses autophagy and promotes apoptosis and sensitivity to cisplatin in oral squamous cell carcinoma. J. Oral. Pathol. Med. 2018, 47, 930–937. [Google Scholar] [CrossRef]
- Tang, B.; Li, N.; Gu, J.; Zhuang, Y.; Li, Q.; Wang, H.; Fang, Y.; Yu, B.; Zhang, J.; Xie, Q.; et al. Compromised autophagy byMIR30B benefits the intracellular survival ofHelicobacter pylori. Autophagy 2014, 8, 1045–1057. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Wang, F.; Hu, S.; Yin, C.; Li, X.; Zhao, S.; Wang, J.; Yan, X. MiR-20a and miR-106b negatively regulate autophagy induced by leucine deprivation via suppression of ULK1 expression in C2C12 myoblasts. Cell Signal. 2012, 24, 2179–2186. [Google Scholar] [CrossRef]
- Hu, J.; Meng, Y.; Zhang, Z.; Yan, Q.; Jiang, X.; Lv, Z.; Hu, L. MARCH5 RNA promotes autophagy, migration, and invasion of ovarian cancer cells. Autophagy 2017, 13, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Liu, C.; Zhou, L.; Wang, J.; Wang, M.; Zhao, B.; Zhao, W.; Xu, S.; Fan, L.; Zhang, X.; et al. APF lncRNA regulates autophagy and myocardial infarction by targeting miR-188-3p. Nat. Commun. 2015, 6, 6779. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Yang, Y.; Ba, Z.; Li, S.; Chen, H.; Hou, X.; Ma, L.; He, P.; Jiang, L.; Li, L.; et al. MicroRNA-93 Regulates Hypoxia-Induced Autophagy by Targeting ULK1. Oxid. Med. Cell Longev. 2017, 2017, 2709053. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Wu, F.; Dong, F.; Chuang, A.Y.; Messer, J.S.; Boone, D.L.; Kwon, J.H. Human autophagy geneATG16L1 is post-transcriptionally regulated byMIR142-3p. Autophagy 2014, 10, 468–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etna, M.P.; Sinigaglia, A.; Grassi, A.; Giacomini, E.; Romagnoli, A.; Pardini, M.; Severa, M.; Cruciani, M.; Rizzo, F.; Anastasiadou, E.; et al. Mycobacterium tuberculosis-induced miR-155 subverts autophagy by targeting ATG3 in human dendritic cells. PLoS Pathog. 2018, 14, e1006790. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, W.N.; Xu, S.B.; Wu, H.; Dai, B.; Jian, D.D.; Yang, M.; Wu, Y.T.; Feng, Q.; Zhu, J.H.; et al. MicroRNA-214-3p: A link between autophagy and endothelial cell dysfunction in atherosclerosis. Acta Physiol. 2018, 222, e12973. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Han, X.; Hu, Z.; Chen, L. The PVT1/miR-216b/Beclin-1 regulates cisplatin sensitivity of NSCLC cells via modulating autophagy and apoptosis. Cancer Chemoth. Pharm. 2019, 83, 921–931. [Google Scholar] [CrossRef] [PubMed]
- Jian, X.; Xiao-yan, Z.; Bin, H.; Yu-feng, Z.; Bo, K.; Zhi-nong, W.; Xin, N. MiR-204 regulate cardiomyocyte autophagy induced by hypoxia-reoxygenation through LC3-II. Int. J. Cardiol. 2011, 148, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Ge, D.; Han, L.; Huang, S.; Peng, N.; Wang, P.; Jiang, Z.; Zhao, J.; Su, L.; Zhang, S.; Zhang, Y.; et al. Identification of a novel MTOR activator and discovery of a competing endogenous RNA regulating autophagy in vascular endothelial cells. Autophagy 2014, 10, 957–971. [Google Scholar] [CrossRef]
- Pan, B.; Feng, B.; Chen, Y.; Huang, G.; Wang, R.; Chen, L.; Song, H. MiR-200b regulates autophagy associated with chemoresistance in human lung adenocarcinoma. Oncotarget 2015, 6, 32805–32820. [Google Scholar] [CrossRef]
- Yang, L.; Wang, H.; Shen, Q.; Feng, L.; Jin, H. Long non-coding RNAs involved in autophagy regulation. Cell Death Dis. 2017, 8, e3073. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.; Wu, J.; Qiu, W.; Lyu, Q.; He, J.; Xie, W.; Xu, N.; Zhang, Y. MiR-15a and miR-16 induce autophagy and enhance chemosensitivity of Camptothecin. Cancer Biol. Ther. 2015, 16, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Zhang, Y.; Han, B.; Bai, Y.; Zhou, R.; Gan, G.; Chao, J.; Hu, G.; Yao, H. Circular RNA HIPK2 regulates astrocyte activation via cooperation of autophagy and ER stress by targeting MIR124-2HG. Autophagy 2017, 13, 1722–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Han, T.; Shi, S.; Chen, E. Long noncoding RNA HAGLROS regulates cell apoptosis and autophagy in lipopolysaccharides-induced WI-38 cells via modulating miR-100/NF-κB axis. Biochem. Biophys. Res. Commun. 2018, 500, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Lu, W.; Ge, D.; Meng, N.; Li, Y.; Su, L.; Zhang, S.; Zhang, Y.; Zhao, B.; Miao, J. A new microRNA signal pathway regulated by long noncoding RNA TGFB2-OT1 in autophagy and inflammation of vascular endothelial cells. Autophagy 2015, 11, 2172–2183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Su, L.; He, X.; Zhao, B.; Miao, J. Long noncoding RNA CA7-4 promotes autophagy and apoptosis via sponging MIR877-3P and MIR5680 in high glucose-induced vascular endothelial cells. Autophagy 2020, 16, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Wu, J.; Yu, X.; Zhou, J.; Yu, H.; Ma, L. Long non-coding RNA MALAT1 enhances the apoptosis of cardiomyocytes through autophagy inhibition by regulating TSC2-mTOR signaling. Biol. Res. 2019, 52. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Fu, H.; Liu, X.; Lei, Q.; Zhang, Y.; She, X.; Liu, Q.; Liu, Q.; Sun, Y.; Li, G.; et al. LINC00470 Coordinates the Epigenetic Regulation of ELFN2 to Distract GBM Cell Autophagy. Mol. Ther. 2018, 26, 2267–2281. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, Z.; Wu, S. Long non-coding RNA CTA sensitizes osteosarcoma cells to doxorubicin through inhibition of autophagy. Oncotarget 2017, 8, 31465–31477. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Wang, C.; Fei, W.; Fang, X.; Hu, X. Epitranscriptomic m6A modification in the stem cell field and its effects on cell death and survival. Am. J. Cancer Res. 2019, 9, 752–764. [Google Scholar]
- Yang, Y.; Hsu, P.J.; Chen, Y.S.; Yang, Y.G. Dynamic transcriptomic m6A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Li, Q.; Li, G.; Zhang, Q.; Zhuo, L.; Han, X.; Zhang, M.; Chen, X.; Pan, T.; Yan, L.; et al. The mechanism of m6A methyltransferase METTL3-mediated autophagy in reversing gefitinib resistance in NSCLC cells by β-elemene. Cell Death Dis. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Lin, Z.; Niu, Y.; Wan, A.; Chen, D.; Liang, H.; Chen, X.; Sun, L.; Zhan, S.; Chen, L.; Cheng, C.; et al. RNA m6A methylation regulates sorafenib resistance in liver cancer through FOXO3-mediated autophagy. EMBO J. 2020, 39, e103181. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Feng, X.; Zhang, H.; Luo, Y.; Huang, J.; Lin, M.; Jin, J.; Ding, X.; Wu, S.; Huang, H.; et al. METTL3 and ALKBH5 oppositely regulate m6A modification of TFEB mRNA, which dictates the fate of hypoxia/reoxygenation-treated cardiomyocytes. Autophagy 2019, 15, 1419–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Zhang, X.; Miao, Y.; Liang, P.; Zhu, K.; She, Y.; Wu, Y.; Liu, D.; Huang, J.; Ren, J.; et al. m6A RNA modification controls autophagy through upregulating ULK1 protein abundance. Cell Res. 2018, 28, 955–957. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lin, T.; Ho, M.; Yeh, J.; Tsai, M.; Hung, K.; Hsieh, I.; Wen, M. Regulation of autophagy in leukocytes through RNA N6-adenosine methylation in chronic kidney disease patients. Biochem. Biophys. Res. Commun. 2020, 527, 953–959. [Google Scholar] [CrossRef]
- Wang, X.; Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Shi, H.; Wang, F.; Wang, Y. m6A mRNA methylation controls autophagy and adipogenesis by targeting Atg5 and Atg7. Autophagy 2020, 16, 1221–1235. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wei, J.; Cui, Y.; Park, G.; Shah, P.; Deng, Y.; Aplin, A.E.; Lu, Z.; Hwang, S.; He, C.; et al. m6A mRNA demethylase FTO regulates melanoma tumorigenicity and response to anti-PD-1 blockade. Nat. Commun. 2019, 10, 2782. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, J.; Xu, D.; Xiang, Z.; Ding, J.; Yang, X.; Li, D.; Han, X. m6A mRNA methylation regulates testosterone synthesis through modulating autophagy in Leydig cells. Autophagy 2021, 17, 457–475. [Google Scholar] [CrossRef]
- Roczniak-Ferguson, A.; Petit, C.S.; Froehlich, F.; Qian, S.; Ky, J.; Angarola, B.; Walther, T.C.; Ferguson, S.M. The Transcription Factor TFEB Links mTORC1 Signaling to Transcriptional Control of Lysosome Homeostasis. Sci. Signal. 2012, 5, ra42. [Google Scholar] [CrossRef] [Green Version]
- Perera, R.M.; Stoykova, S.; Nicolay, B.N.; Ross, K.N.; Fitamant, J.; Boukhali, M.; Lengrand, J.; Deshpande, V.; Selig, M.K.; Ferrone, C.R.; et al. Transcriptional control of autophagy-lysosome function drives pancreatic cancer metabolism. Nature 2015, 524, 361–365. [Google Scholar] [CrossRef]
- Ma, L.; Ma, Y.; Zhang, Z.; Wang, Q.; Liu, X. The signaling axis of Rac1-TFEB regulates autophagy and tumorigenesis. Anti-Cancer Drug 2019, 30, 1031–1037. [Google Scholar] [CrossRef]
- Djavaheri-Mergny, M.; Amelotti, M.; Mathieu, J.; Besançon, F.; Bauvy, C.; Souquère, S.; Pierron, G.; Codogno, P. NF-κB Activation Represses Tumor Necrosis Factor-α-induced Autophagy. J. Biol. Chem. 2006, 281, 30373–30382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talebian, S.; Daghagh, H.; Yousefi, B.; Ozkul, Y.; Ilkhani, K.; Seif, F.; Alivand, M.R. The role of epigenetics and non-coding RNAs in autophagy: A new perspective for thorough understanding. Mech. Ageing Dev. 2020, 190, 111309. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Bu, S. Clinical value of circular RNAs and autophagy-related miRNAs in the diagnosis and treatment of pancreatic cancer. Hepatob. Pancreat. Dis. 2019, 18, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Vlachos, I.S.; Karagkouni, D.; Georgakilas, G.; Kanellos, I.; Vergoulis, T.; Zagganas, K.; Tsanakas, P.; Floros, E.; Dalamagas, T.; et al. DIANA-LncBase v2: Indexing microRNA targets on non-coding transcripts. Nucleic. Acids Res. 2016, 44, D231–D238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horos, R.; Buscher, M.; Sachse, C.; Hentze, M.W. Vault RNA emerges as a regulator of selective autophagy. Autophagy 2019, 15, 1463–1464. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y. Epigenetic Cross-Talk between DNA Methylation and Histone Modifications in Human Cancers. Yonsei Med. J. 2009, 50, 455. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Transcription Factor | Function |
|---|---|
| Leucine zipper transcription factors (MiT/TFE) | MiT/TFE recognize promoters of lysosomal and Atg genes and represent transcriptional controllers of lysosomal biogenesis and autophagy [4,13]. |
| Nuclear receptors PPARα and FXR | PPARα and FXR oppositely control the expression of Atg7, Beclin1, Bnip3, and LC3 and autophagic vesicle formation [6]. |
| Small heterodimer partner (SHP) | SHP decreases mRNA levels of Atg genes and inhibits autophagy [21]. |
| Transcription factors FOXO/FOXA | Activation of FOXO/FOXA induces the expression of multiple Atg genes and lysosomal genes [17,32]. |
| CCAAT/enhancer binding protein beta (C/EBPβ) | C/EBPβ targets key Atg genes and induces the expression of Atg genes [23,33]. |
| Activating transcription factor 4(ATF4) | ATF4 is involved in the cellular stress response and autophagosome formation [5,34]. |
| Nuclear factor-kappa B (NF-κB) | NF-κB activates the expression of Atg genes and induces autophagy [28,35]. |
| Zinc-finger-family DNA-binding protein, ZKSCAN3 | ZKSCAN3 decreases mRNA levels of Atg genes and inhibits autophagy [16]. |
| Tumor suppressor p53 | In the nucleus, P53 transactivates Atg genes and induces autophagy by inhibiting mTOR; in the cytoplasm, P53 suppresses autophagy [36,37]. |
| Signal transducer and activator of transcription (STAT) | STAT3 phosphorylation upregulates BNIP3 expression; STAT1 suppresses the expression of Atg genes [38,39]. |
| Transcription factor E2F | Activation of E2F1 upregulates the expression of Atg genes [24]. |
| TGA9 (TGACG motif-binding protein 9) | TGA9 activates autophagy by upregulating the expression of Atg genes [31]. |
| E93 | Knockdown of E93 reduces the expression of several Atg genes in B. mori [40]. |
| EcR-USP | 20E-EcR-USP upregulates the transcription of Atg genes to induce autophagy [11]. |
| Non-Coding RNAs | Target Genes | Species | Impact on Autophagy |
|---|---|---|---|
| miR30b | Atg12, Beclin-1 | Helicobacter pylori | ↓[58] |
| miR-17 | Ulk1 | Mouse | ↓[59] |
| miR-30a | Beclin1, Atg12, Atg5 | Mouse | ↓[44,60] |
| miR-188-3p | Atg7 | Mouse | ↓[61] |
| miR-93, miR106b, miR142-3p | ULK1, ATG16L | Human | ↓[44,62,63] |
| miR-101 | ATG4D, LC3 | Human | ↓[43,44] |
| miR-155 | ATG3 | Human | ↓[64] |
| miR-214-3p | ATG5, ATG12 | Human | ↓[65] |
| miR-216b | BECLIN1 | Human | ↓[66] |
| miR-103a-3p | ATG5 | Human | ↓[45] |
| miR-183, miR-204 | LC3B1/LC3-II | Human | ↓[44,67] |
| miR-83, miR-29 | atg-4.2 / ATG4D, ATG9a | Caenorhabditis elegans/Human | ↓[8,46] |
| miR-34 | Atg9a/ATG9a | Caenorhabditis elegans/Human | ↓[47] |
| miR-4459 | ATG13 | Human | ↓[68] |
| miR-23b | ATG12 | Human | ↓[69] |
| miR-19a | BECLIN1, LC3 | Human | ↓[70] |
| miR-376b | ATG4C, BECLIN1 | Human | ↓[44] |
| miR-15a, miR-16 | Rictor (mTORC1) | Human | ↑[71] |
| circNF1-419 | Dynamin-1 | Mouse | ↑[50] |
| circHIPK2 | ATG5, BECLIN1-1 | Human | ↑[72] |
| circPABPN1 | ATG16l1 | Human | ↓[51] |
| lncRNA APF | Atg7 | Mouse | ↑[61] |
| lncRNA NEAT1, lncRNA XIST | Atg9a | Mouse | ↑[8,53] |
| lncRNA HAGLROS | PI3K-AKT-NF-κB | Human | ↑[73] |
| lncRNA TGFB2-OT1 | ATG3, ATG7, ATG13 | Human | ↑[74] |
| lncRNA CA7-4 | AMPK | Human | ↑[75] |
| lncRNA GBCDRlnc1 | BECLIN1, ATG5, ATG12 | Human | ↑[54,55] |
| lncRNA MALAT1 | Beclin1, LC3 | Mouse | ↓[76] |
| lncRNA LINC00470 | BECLIN1, ATG3, ATG7 | Human | ↓[77] |
| lncRNA CTA | Unknown | Human | ↓[78] |
| lncRNA HOTAIR | BECLIN1, LC3, ATG3, ATG7 | Human | ↓[57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Long, S.; Gan, Z.; Tettamanti, G.; Li, K.; Tian, L. Transcriptional and Post-Transcriptional Regulation of Autophagy. Cells 2022, 11, 441. https://doi.org/10.3390/cells11030441
Ma Q, Long S, Gan Z, Tettamanti G, Li K, Tian L. Transcriptional and Post-Transcriptional Regulation of Autophagy. Cells. 2022; 11(3):441. https://doi.org/10.3390/cells11030441
Chicago/Turabian StyleMa, Qiuqin, Shihui Long, Zhending Gan, Gianluca Tettamanti, Kang Li, and Ling Tian. 2022. "Transcriptional and Post-Transcriptional Regulation of Autophagy" Cells 11, no. 3: 441. https://doi.org/10.3390/cells11030441
APA StyleMa, Q., Long, S., Gan, Z., Tettamanti, G., Li, K., & Tian, L. (2022). Transcriptional and Post-Transcriptional Regulation of Autophagy. Cells, 11(3), 441. https://doi.org/10.3390/cells11030441