
Altered Phenotype and Enhanced Antibody-Producing Ability of Peripheral B Cells in Mice with Cd19-Driven Cre Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Immunizations
2.3. Cell Isolation and Culture
2.4. ELISA & ELISPOT Analysis
2.5. Flow Cytometric Analysis
2.6. Literature Survey
2.7. Statistical Analysis
3. Results
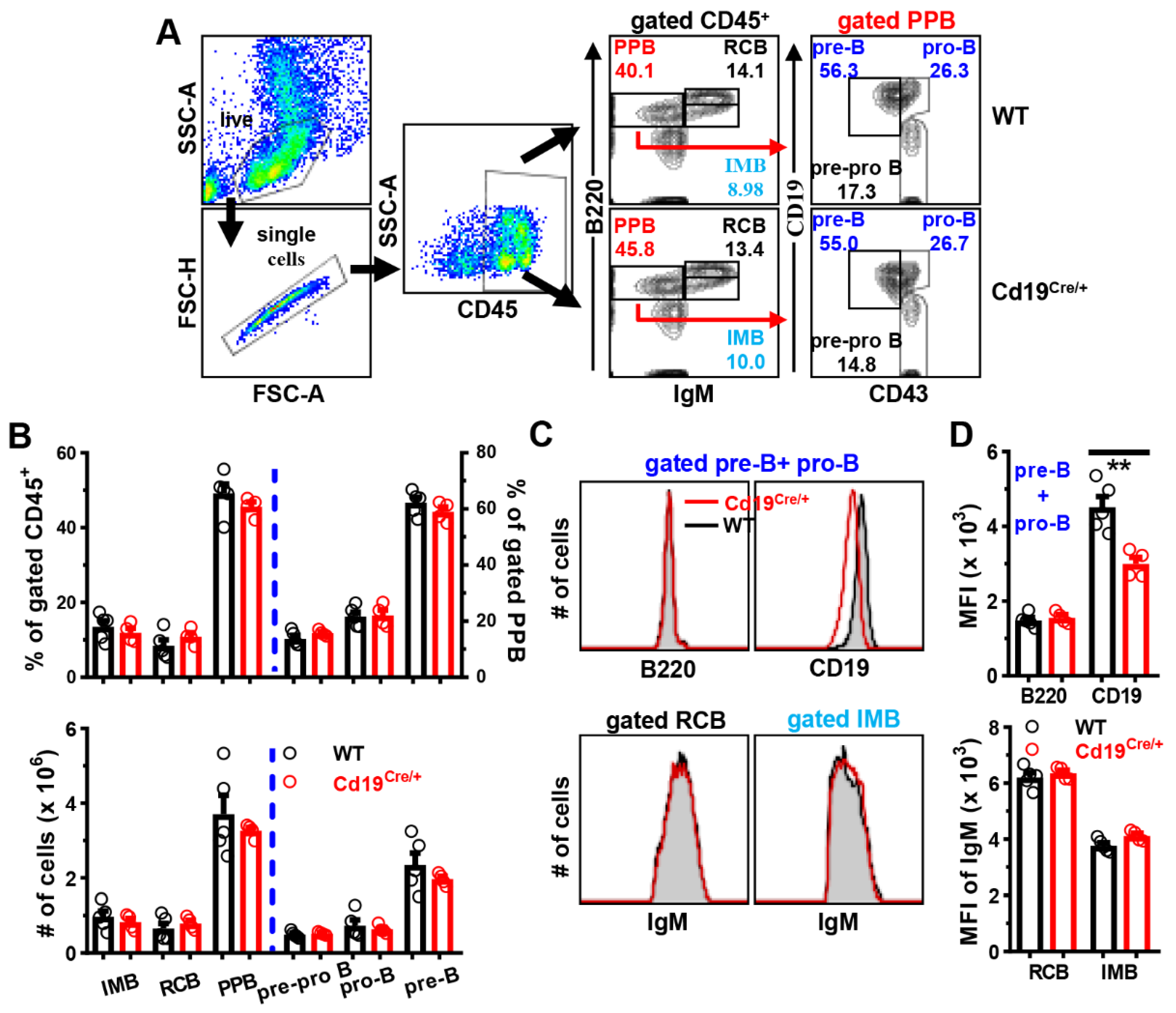
3.1. Comparable B Cell Development in the BM of Cd19Cre/+ Mice
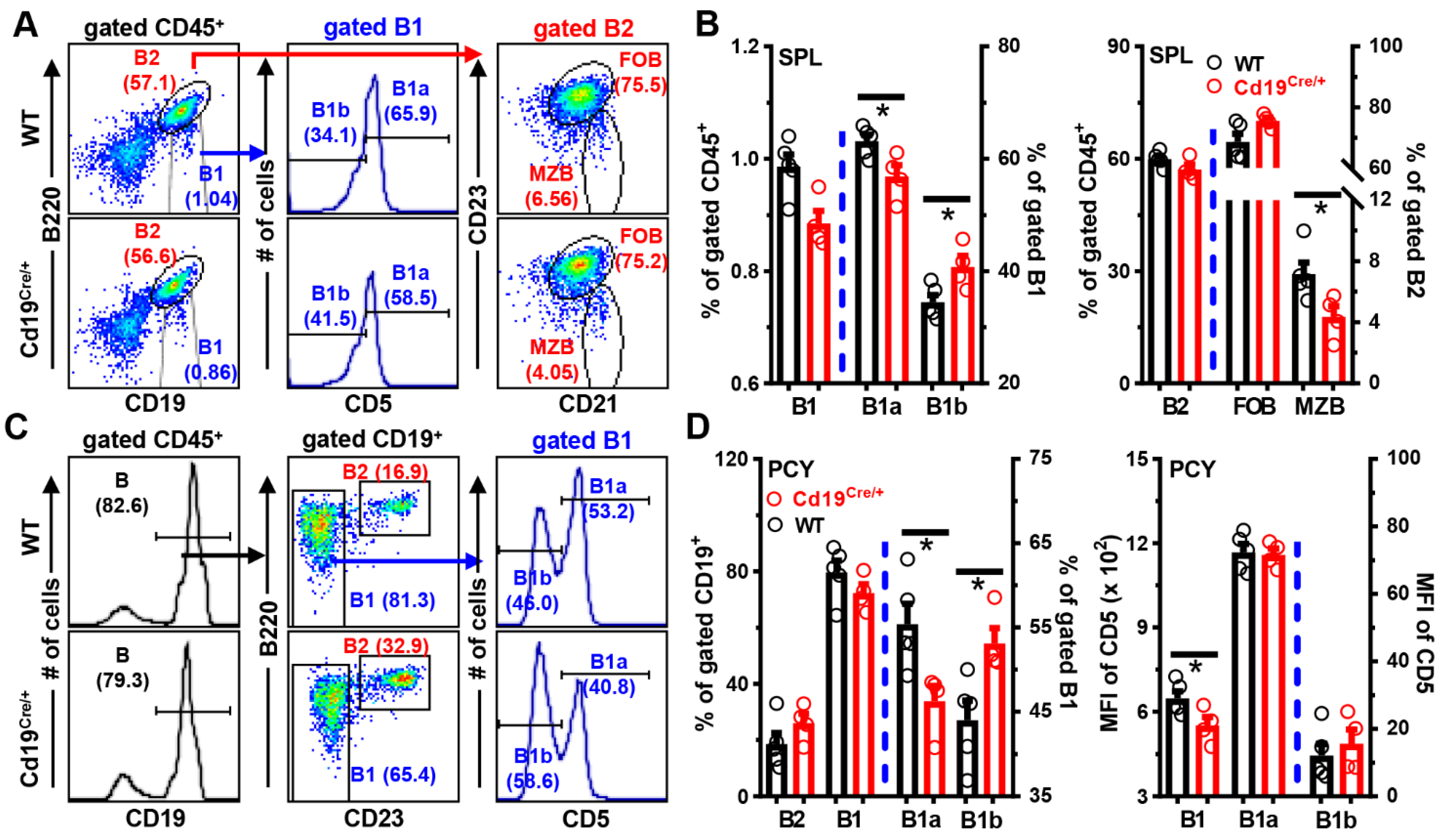
3.2. Mildly Disturbed Phenotypes of B Cells in the Periphery of Cd19Cre/+ Mice
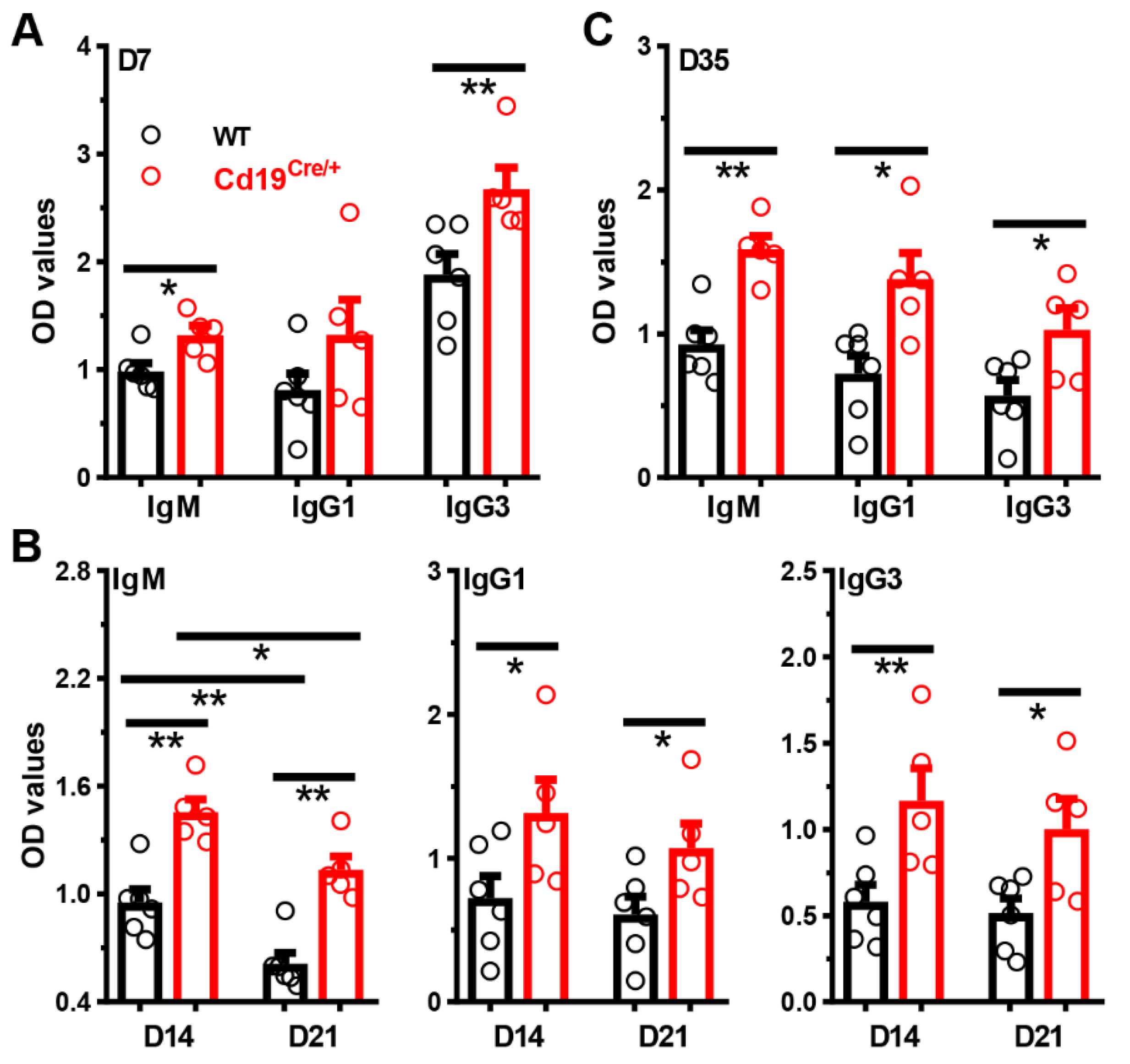
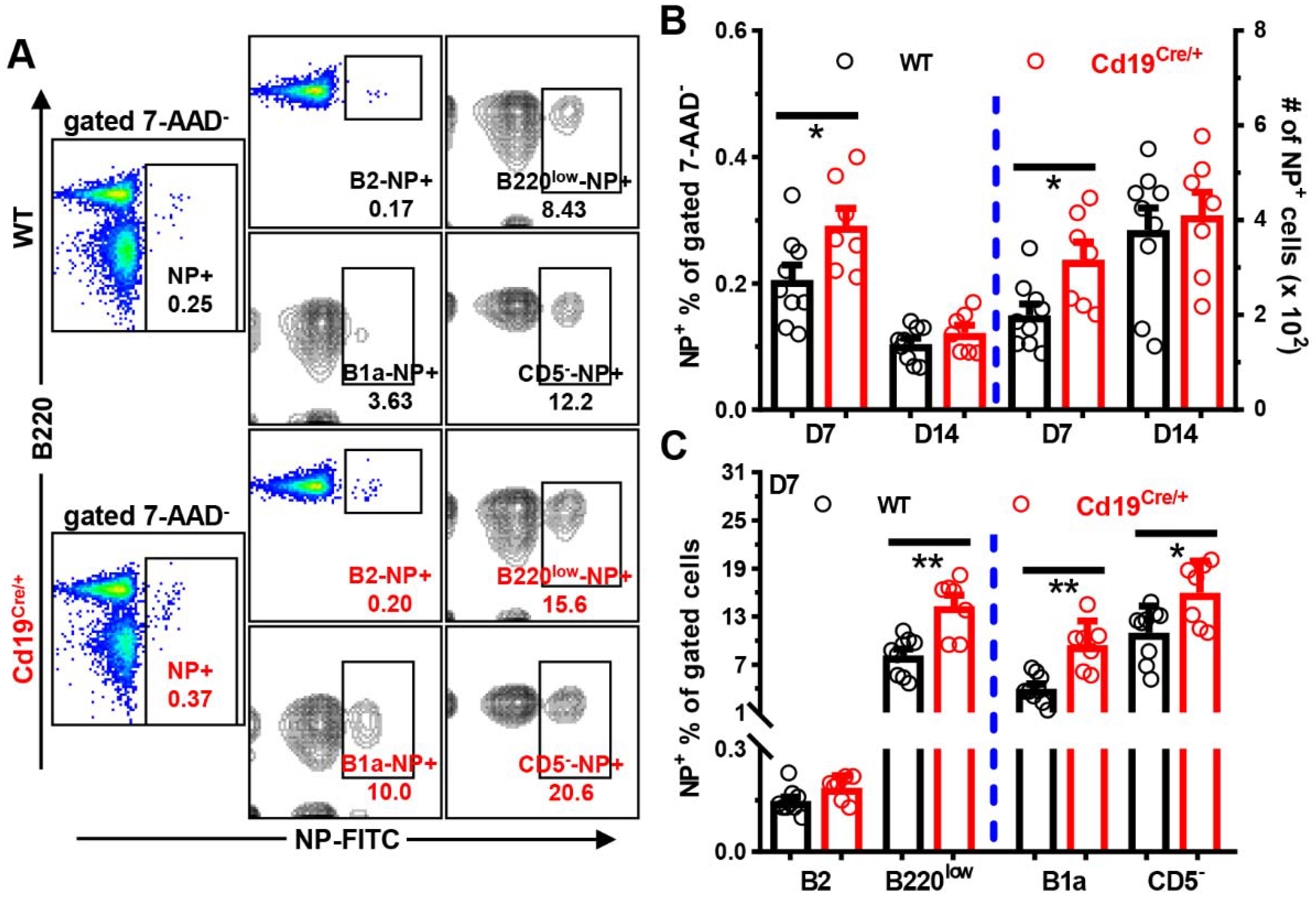
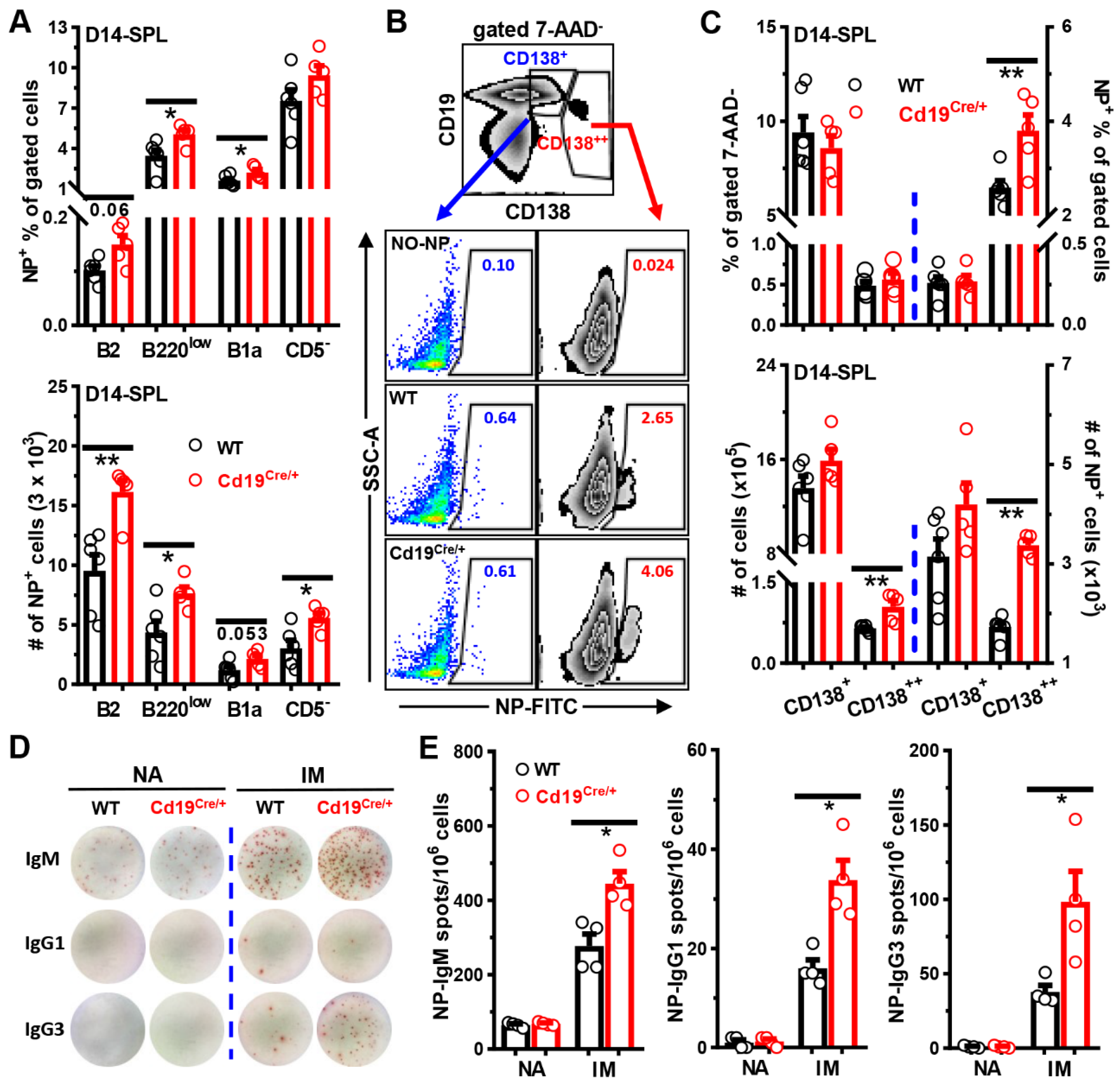
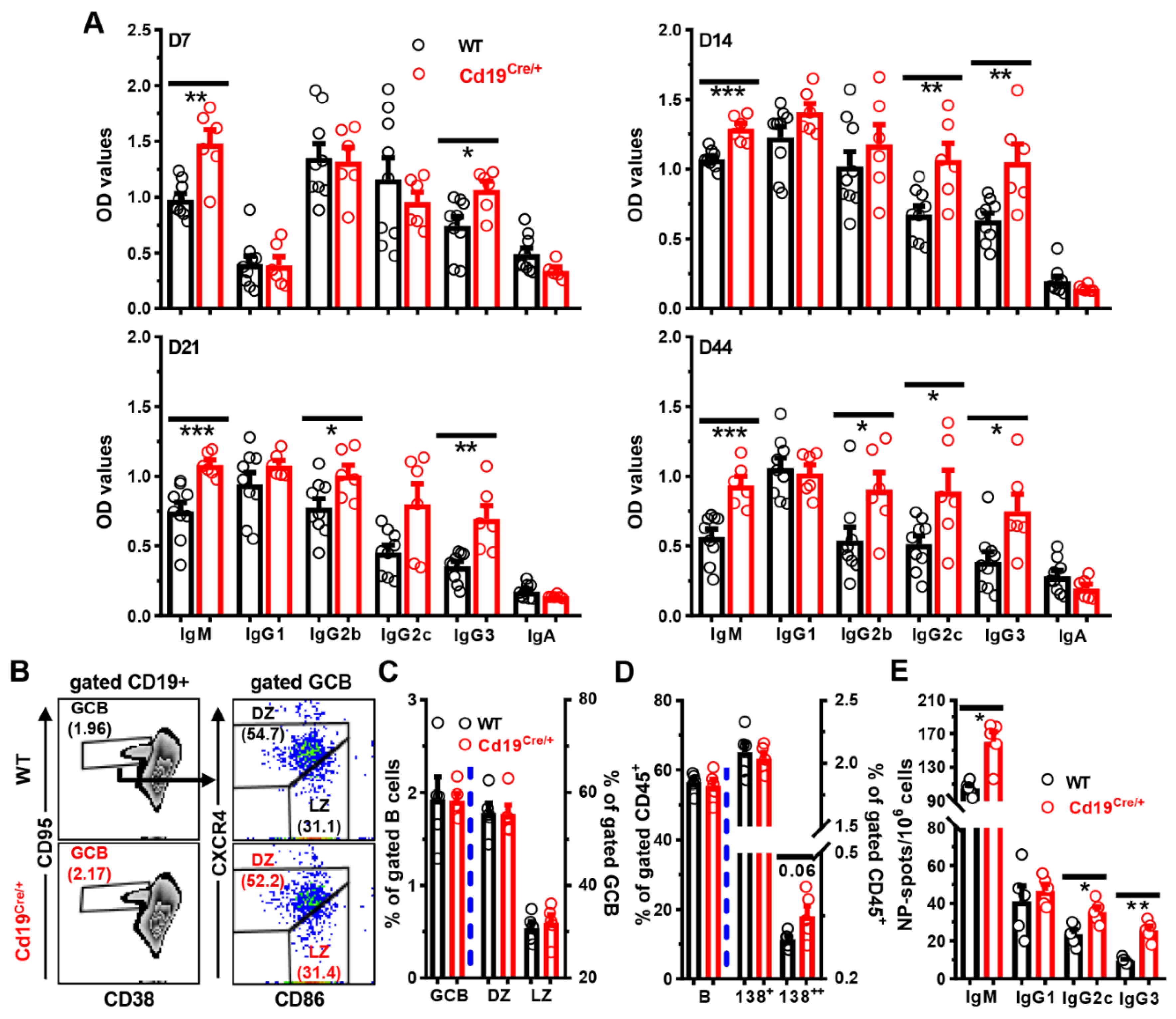
3.3. Increased Antibody Levels in Cd19Cre/+ Mice upon Immunization
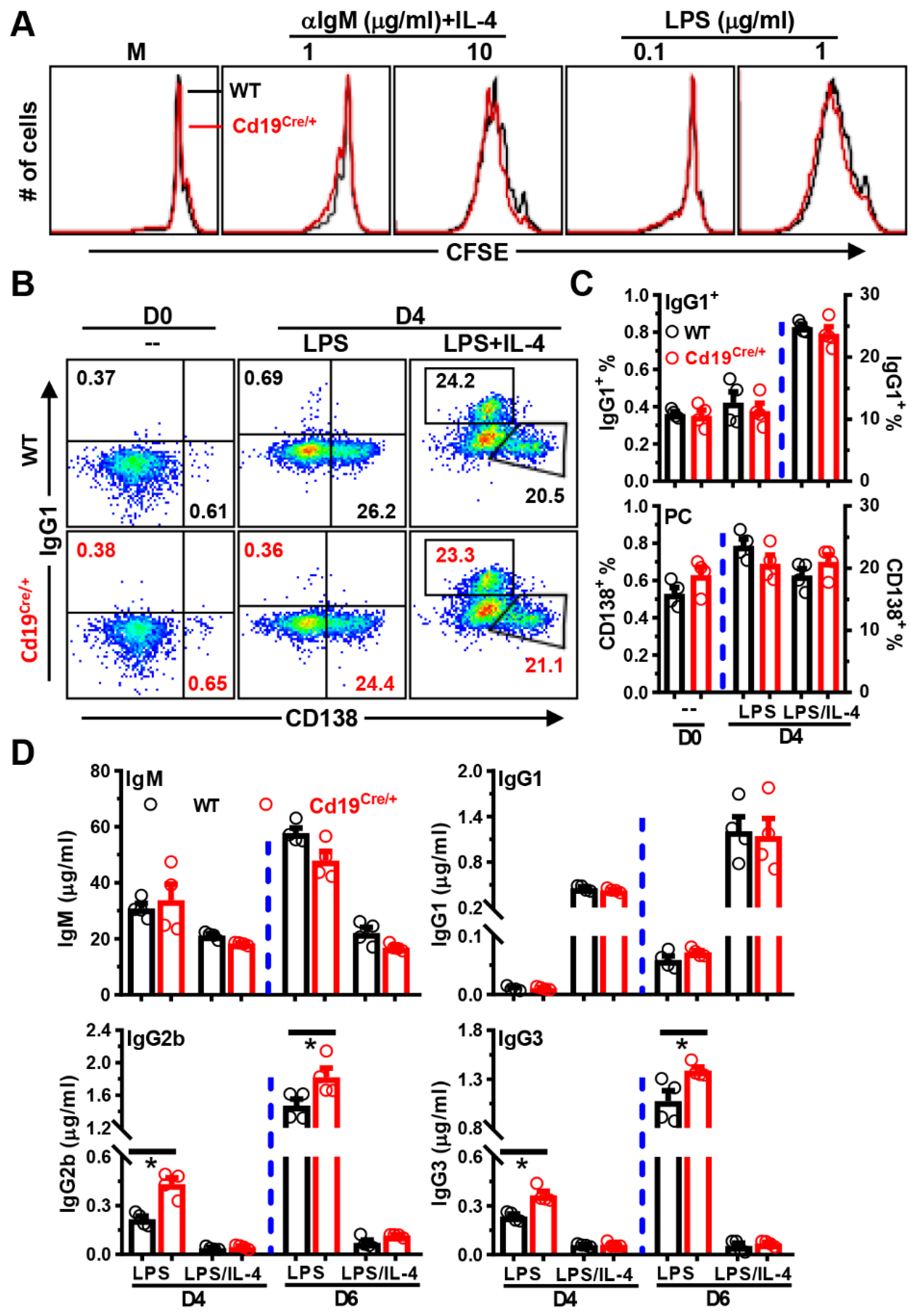
3.4. Increased Antibody-Producing Ability of B Cells from Cd19Cre/+ Mice upon LPS-Stimulation In Vitro
3.5. Survey of Published Literature Using Mice Containing Cd19Cre/+ Transgene Reveals That the Phenotype Is Confounded by Different Controls Used
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Orban, P.C.; Chui, D.; Marth, J.D. Tissue- and site-specific DNA recombination in transgenic mice. Proc. Natl. Acad. Sci. USA 1992, 89, 6861–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLellan, M.A.; Rosenthal, N.A.; Pinto, A.R. Cre-loxP-Mediated Recombination: General Principles and Experimental Considerations. Curr. Protoc. Mouse Biol. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Supprian, M.; Rajewsky, K. Vagaries of conditional gene targeting. Nat. Immunol. 2007, 8, 665–668. [Google Scholar] [CrossRef] [PubMed]
- Abram, C.L.; Roberge, G.L.; Hu, Y.; Lowell, C.A. Comparative analysis of the efficiency and specificity of myeloid-Cre deleting strains using ROSA-EYFP reporter mice. J. Immunol. Methods 2014, 408, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caton, M.L.; Smith-Raska, M.R.; Reizis, B. Notch-RBP-J signaling controls the homeostasis of CD8- dendritic cells in the spleen. J. Exp. Med. 2007, 204, 1653–1664. [Google Scholar] [CrossRef] [Green Version]
- Heffner, C.S.; Herbert Pratt, C.; Babiuk, R.P.; Sharma, Y.; Rockwood, S.F.; Donahue, L.R.; Eppig, J.T.; Murray, S.A. Supporting conditional mouse mutagenesis with a comprehensive cre characterization resource. Nat. Commun. 2012, 3, 1218. [Google Scholar] [CrossRef] [Green Version]
- Thyagarajan, B.; Guimaraes, M.J.; Groth, A.C.; Calos, M.P. Mammalian genomes contain active recombinase recognition sites. Gene 2000, 244, 47–54. [Google Scholar] [CrossRef]
- Semprini, S.; Troup, T.J.; Kotelevtseva, N.; King, K.; Davis, J.R.; Mullins, L.J.; Chapman, K.E.; Dunbar, D.R.; Mullins, J.J. Cryptic loxP sites in mammalian genomes: Genome-wide distribution and relevance for the efficiency of BAC/PAC recombineering techniques. Nucleic Acids Res. 2007, 35, 1402–1410. [Google Scholar] [CrossRef]
- Loonstra, A.; Vooijs, M.; Beverloo, H.B.; Allak, B.A.; van Drunen, E.; Kanaar, R.; Berns, A.; Jonkers, J. Growth inhibition and DNA damage induced by Cre recombinase in mammalian cells. Proc. Natl. Acad. Sci. USA 2001, 98, 9209–9214. [Google Scholar] [CrossRef] [Green Version]
- Bersell, K.; Choudhury, S.; Mollova, M.; Polizzotti, B.D.; Ganapathy, B.; Walsh, S.; Wadugu, B.; Arab, S.; Kuhn, B. Moderate and high amounts of tamoxifen in alphaMHC-MerCreMer mice induce a DNA damage response, leading to heart failure and death. Dis. Model. Mech. 2013, 6, 1459–1469. [Google Scholar] [CrossRef]
- Pugach, E.K.; Richmond, P.A.; Azofeifa, J.G.; Dowell, R.D.; Leinwand, L.A. Prolonged Cre expression driven by the alpha-myosin heavy chain promoter can be cardiotoxic. J. Mol. Cell. Cardiol. 2015, 86, 54–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carow, B.; Gao, Y.; Coquet, J.; Reilly, M.; Rottenberg, M.E. lck-Driven Cre Expression Alters T Cell Development in the Thymus and the Frequencies and Functions of Peripheral T Cell Subsets. J. Immunol. 2016, 197, 2261–2268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Ristow, M.; Lin, X.; White, M.F.; Magnuson, M.A.; Hennighausen, L. RIP-Cre revisited, evidence for impairments of pancreatic beta-cell function. J. Biol. Chem. 2006, 281, 2649–2653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitelman, G.; Kedees, M. Mouse insulin cells expressing an inducible RIPCre transgene are functionally impaired. J. Biol. Chem. 2015, 290, 3647–3653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Wei, G.; Liu, D. CD19: A biomarker for B cell development, lymphoma diagnosis and therapy. Exp. Hematol. Oncol. 2012, 1, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Chen, G.; Zhao, Y.; Gao, X.M.; Wang, J. Regulation of the Development and Function of B Cells by ZBTB Transcription Factors. Front. Immunol. 2018, 9, 580. [Google Scholar] [CrossRef]
- Rickert, R.C.; Rajewsky, K.; Roes, J. Impairment of T-cell-dependent B-cell responses and B-1 cell development in CD19-deficient mice. Nature 1995, 376, 352–355. [Google Scholar] [CrossRef]
- Rickert, R.C.; Roes, J.; Rajewsky, K. B lymphocyte-specific, Cre-mediated mutagenesis in mice. Nucleic Acids Res. 1997, 25, 1317–1318. [Google Scholar] [CrossRef]
- Grabow, S.; Kelly, G.L.; Delbridge, A.R.; Kelly, P.N.; Bouillet, P.; Adams, J.M.; Strasser, A. Critical B-lymphoid cell intrinsic role of endogenous MCL-1 in c-MYC-induced lymphomagenesis. Cell Death Dis. 2016, 7, e2132. [Google Scholar] [CrossRef]
- Hart, G.T.; Wang, X.; Hogquist, K.A.; Jameson, S.C. Kruppel-like factor 2 (KLF2) regulates B-cell reactivity, subset differentiation, and trafficking molecule expression. Proc. Natl. Acad. Sci. USA 2011, 108, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Tabor, D.E.; Gould, K.A. Estrogen receptor alpha promotes lupus in (NZBxNZW)F1 mice in a B cell intrinsic manner. Clin. Immunol. 2017, 174, 41–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krop, I.; de Fougerolles, A.R.; Hardy, R.R.; Allison, M.; Schlissel, M.S.; Fearon, D.T. Self-renewal of B-1 lymphocytes is dependent on CD19. Eur. J. Immunol. 1996, 26, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.; Kearney, J.F. Positive selection from newly formed to marginal zone B cells depends on the rate of clonal production, CD19, and btk. Immunity 2000, 12, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Otero, D.C.; Anzelon, A.N.; Rickert, R.C. CD19 function in early and late B cell development: I. Maintenance of follicular and marginal zone B cells requires CD19-dependent survival signals. J. Immunol. 2003, 170, 73–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, Y.; Zhao, H.; Wang, Y.; Carter, R.H. Cutting edge: Primary and secondary effects of CD19 deficiency on cells of the marginal zone. J. Immunol. 2009, 182, 7343–7347. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.B.; Silverman, M.; Monroe, J.G. Transitional B cells: Step by step towards immune competence. Trends Immunol. 2003, 24, 343–349. [Google Scholar] [CrossRef]
- Sakurai, N.; Maeda, M.; Lee, S.U.; Ishikawa, Y.; Li, M.; Williams, J.C.; Wang, L.; Su, L.; Suzuki, M.; Saito, T.I.; et al. The LRF transcription factor regulates mature B cell development and the germinal center response in mice. J. Clin. Investig. 2011, 121, 2583–2598. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Brooks, S.R.; Li, X.; Anzelon, A.N.; Rickert, R.C.; Carter, R.H. The physiologic role of CD19 cytoplasmic tyrosines. Immunity 2002, 17, 501–514. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, A.; Beard, L.J.; Feldman, R.G. IgG subclass distribution of antibodies to bacterial and viral antigens. Pediatr. Infect. Dis. J. 1990, 9, S16–S24. [Google Scholar] [CrossRef]
- Arenzana, T.L.; Smith-Raska, M.R.; Reizis, B. Transcription factor Zfx controls BCR-induced proliferation and survival of B lymphocytes. Blood 2009, 113, 5857–5867. [Google Scholar] [CrossRef] [Green Version]
- Homig-Holzel, C.; Hojer, C.; Rastelli, J.; Casola, S.; Strobl, L.J.; Muller, W.; Quintanilla-Martinez, L.; Gewies, A.; Ruland, J.; Rajewsky, K.; et al. Constitutive CD40 signaling in B cells selectively activates the noncanonical NF-kappaB pathway and promotes lymphomagenesis. J. Exp. Med. 2008, 205, 1317–1329. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Kim, T.S.; Jacob, A.; Walsh, M.C.; Kadono, Y.; Fuentes-Panana, E.; Yoshioka, T.; Yoshimura, A.; Yamamoto, M.; Kaisho, T.; et al. TRAF6 is required for generation of the B-1a B cell compartment as well as T cell-dependent and -independent humoral immune responses. PLoS ONE 2009, 4, e4736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.Y.; Wolfram, P.; Canty, K.; Harley, B.; Nombela-Arrieta, C.; Pivarnik, G.; Manis, J.; Beggs, H.E.; Silberstein, L.E. Focal adhesion kinase regulates the localization and retention of pro-B cells in bone marrow microenvironments. J. Immunol. 2013, 190, 1094–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramon, S.; Bancos, S.; Thatcher, T.H.; Murant, T.I.; Moshkani, S.; Sahler, J.M.; Bottaro, A.; Sime, P.J.; Phipps, R.P. Peroxisome proliferator-activated receptor gamma B cell-specific-deficient mice have an impaired antibody response. J. Immunol. 2012, 189, 4740–4747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veillette, A.; Zhang, S.; Shi, X.; Dong, Z.; Davidson, D.; Zhong, M.C. SAP expression in T cells, not in B cells, is required for humoral immunity. Proc. Natl. Acad. Sci. USA 2008, 105, 1273–1278. [Google Scholar] [CrossRef] [Green Version]
- Cawley, K.M.; Bustamante-Gomez, N.C.; Guha, A.G.; MacLeod, R.S.; Xiong, J.; Gubrij, I.; Liu, Y.; Mulkey, R.; Palmieri, M.; Thostenson, J.D.; et al. Local Production of Osteoprotegerin by Osteoblasts Suppresses Bone Resorption. Cell Rep. 2020, 32, 108052. [Google Scholar] [CrossRef]
- Guo, M.; Price, M.J.; Patterson, D.G.; Barwick, B.G.; Haines, R.R.; Kania, A.K.; Bradley, J.E.; Randall, T.D.; Boss, J.M.; Scharer, C.D. EZH2 Represses the B Cell Transcriptional Program and Regulates Antibody-Secreting Cell Metabolism and Antibody Production. J. Immunol. 2018, 200, 1039–1052. [Google Scholar] [CrossRef]
- Murga, M.; Lecona, E.; Kamileri, I.; Diaz, M.; Lugli, N.; Sotiriou, S.K.; Anton, M.E.; Mendez, J.; Halazonetis, T.D.; Fernandez-Capetillo, O. POLD3 Is Haploinsufficient for DNA Replication in Mice. Mol. Cell 2016, 63, 877–883. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Bod, L.; Pochet, N.; Kota, S.B.; Hu, D.; Madi, A.; Kilpatrick, J.; Shi, J.; Ho, A.; Zhang, H.; et al. Checkpoint Receptor TIGIT Expressed on Tim-1(+) B Cells Regulates Tissue Inflammation. Cell Rep. 2020, 32, 107892. [Google Scholar] [CrossRef]
- Xu, W.; Fukuyama, T.; Ney, P.A.; Wang, D.; Rehg, J.; Boyd, K.; van Deursen, J.M.; Brindle, P.K. Global transcriptional coactivators CREB-binding protein and p300 are highly essential collectively but not individually in peripheral B cells. Blood 2006, 107, 4407–4416. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Li, X.; Ma, Z.; Wang, C.; Yang, Q.; Byrne-Steele, M.; Hong, R.; Min, Q.; Zhou, G.; Cheng, Y.; et al. CTLA-4 expression by B-1a B cells is essential for immune tolerance. Nat. Commun. 2021, 12, 525. [Google Scholar] [CrossRef]
- Johnson, J.L.; Rosenthal, R.L.; Knox, J.J.; Myles, A.; Naradikian, M.S.; Madej, J.; Kostiv, M.; Rosenfeld, A.M.; Meng, W.; Christensen, S.R.; et al. The Transcription Factor T-bet Resolves Memory B Cell Subsets with Distinct Tissue Distributions and Antibody Specificities in Mice and Humans. Immunity 2020, 52, 842–855. [Google Scholar] [CrossRef] [PubMed]
- Kasama, Y.; Sekiguchi, S.; Saito, M.; Tanaka, K.; Satoh, M.; Kuwahara, K.; Sakaguchi, N.; Takeya, M.; Hiasa, Y.; Kohara, M.; et al. Persistent expression of the full genome of hepatitis C virus in B cells induces spontaneous development of B-cell lymphomas in vivo. Blood 2010, 116, 4926–4933. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Lin, Y.; Mishra, A.; Shelly, C.; Gao, R.; Reeh, C.W.; Wang, P.Z.; Xi, R.; Liu, Y.; Wenzel, P.; et al. Bmi1 Maintains the Self-Renewal Property of Innate-like B Lymphocytes. J. Immunol. 2020, 204, 3262–3272. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Sun, Y.; Chen, J.; Zhang, Y.; Fan, S.; Huang, M.; Xie, X.; Cai, Y.; Shang, Y.; Gui, T.; et al. REV7 is required for processing AID initiated DNA lesions in activated B cells. Nat. Commun. 2020, 11, 2812. [Google Scholar] [CrossRef]
- Feyerabend, T.B.; Weiser, A.; Tietz, A.; Stassen, M.; Harris, N.; Kopf, M.; Radermacher, P.; Moller, P.; Benoist, C.; Mathis, D.; et al. Cre-mediated cell ablation contests mast cell contribution in models of antibody- and T cell-mediated autoimmunity. Immunity 2011, 35, 832–844. [Google Scholar] [CrossRef] [Green Version]
- Ohnmacht, C.; Schwartz, C.; Panzer, M.; Schiedewitz, I.; Naumann, R.; Voehringer, D. Basophils orchestrate chronic allergic dermatitis and protective immunity against helminths. Immunity 2010, 33, 364–374. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Ono, N.; Steeber, D.A.; Pisetsky, D.S.; Tedder, T.F. CD19 regulates B lymphocyte signaling thresholds critical for the development of B-1 lineage cells and autoimmunity. J. Immunol. 1996, 157, 4371–4378. [Google Scholar]
- Engel, P.; Zhou, L.J.; Ord, D.C.; Sato, S.; Koller, B.; Tedder, T.F. Abnormal B lymphocyte development, activation, and differentiation in mice that lack or overexpress the CD19 signal transduction molecule. Immunity 1995, 3, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Haas, K.M.; Poe, J.C.; Steeber, D.A.; Tedder, T.F. B-1a and B-1b cells exhibit distinct developmental requirements and have unique functional roles in innate and adaptive immunity to S. pneumoniae. Immunity 2005, 23, 7–18. [Google Scholar] [CrossRef] [Green Version]
- Oliver, A.M.; Martin, F.; Gartland, G.L.; Carter, R.H.; Kearney, J.F. Marginal zone B cells exhibit unique activation, proliferative and immunoglobulin secretory responses. Eur. J. Immunol. 1997, 27, 2366–2374. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, B.; Quinn, W.J., 3rd; Hazard, K.; Erikson, J.; Allman, D. Characterization of marginal zone B cell precursors. J. Exp. Med. 2005, 202, 1225–1234. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Rajewsky, K. Homeostasis of peripheral B cells in the absence of B cell influx from the bone marrow. J. Exp. Med. 2001, 194, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.W.; Mrazek, M.D.; Yang, Y.; Herzenberg, L.A. Phenotypically distinct B cell development pathways map to the three B cell lineages in the mouse. Proc. Natl. Acad. Sci. USA 2006, 103, 6293–6298. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Steeber, D.A.; Tedder, T.F. The CD19 signal transduction molecule is a response regulator of B-lymphocyte differentiation. Proc. Natl. Acad. Sci. USA 1995, 92, 11558–11562. [Google Scholar] [CrossRef] [Green Version]
- Tsitsikov, E.N.; Gutierrez-Ramos, J.C.; Geha, R.S. Impaired CD19 expression and signaling, enhanced antibody response to type II T independent antigen and reduction of B-1 cells in CD81-deficient mice. Proc. Natl. Acad. Sci. USA 1997, 94, 10844–10849. [Google Scholar] [CrossRef] [Green Version]
- Rigley, K.P.; Callard, R.E. Inhibition of B cell proliferation with anti-CD19 monoclonal antibodies: Anti-CD19 antibodies do not interfere with early signaling events triggered by anti-IgM or interleukin 4. Eur. J. Immunol. 1991, 21, 535–540. [Google Scholar] [CrossRef]
- Pape, K.A.; Maul, R.W.; Dileepan, T.; Paustian, A.S.; Gearhart, P.J.; Jenkins, M.K. Naive B Cells with High-Avidity Germline-Encoded Antigen Receptors Produce Persistent IgM(+) and Transient IgG(+) Memory B Cells. Immunity 2018, 48, 1135–1143. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Steeber, D.A.; Jansen, P.J.; Tedder, T.F. CD19 expression levels regulate B lymphocyte development: Human CD19 restores normal function in mice lacking endogenous CD19. J. Immunol. 1997, 158, 4662–4669. [Google Scholar]
- Gardby, E.; Chen, X.J.; Lycke, N.Y. Impaired CD40-signalling in CD19-deficient mice selectively affects Th2-dependent isotype switching. Scand. J. Immunol. 2001, 53, 13–23. [Google Scholar] [CrossRef]
- Gardby, E.; Lycke, N.Y. CD19-deficient mice exhibit poor responsiveness to oral immunization despite evidence of unaltered total IgA levels, germinal centers and IgA-isotype switching in Peyer’s patches. Eur J. Immunol. 2000, 30, 1861–1871. [Google Scholar] [CrossRef]
- Matsushita, T.; Fujimoto, M.; Hasegawa, M.; Komura, K.; Takehara, K.; Tedder, T.F.; Sato, S. Inhibitory role of CD19 in the progression of experimental autoimmune encephalomyelitis by regulating cytokine response. Am. J. Pathol. 2006, 168, 812–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, R.; Fujimoto, M.; Ishiura, N.; Kuwano, Y.; Nakashima, H.; Yazawa, N.; Okochi, H.; Sato, S.; Tedder, T.F.; Tamaki, K. CD19 expression in B cells is important for suppression of contact hypersensitivity. Am. J. Pathol. 2007, 171, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Ishiura, N.; Nakashima, H.; Kuwano, Y.; Okochi, H.; Tamaki, K.; Sato, S.; Tedder, T.F.; Fujimoto, M. Regulatory B cells (B10 cells) have a suppressive role in murine lupus: CD19 and B10 cell deficiency exacerbates systemic autoimmunity. J. Immunol. 2010, 184, 4801–4809. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ding, Z.; Xu, H.; Heyman, B. Marginal zone B cells transport IgG3-immune complexes to splenic follicles. J. Immunol. 2014, 193, 1681–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Zhao, S.; Qin, X.-Y.; He, T.-T.; Hu, M.-M.; Gong, Z.; Wang, H.-M.; Gong, F.-Y.; Gao, X.-M.; Wang, J. Altered Phenotype and Enhanced Antibody-Producing Ability of Peripheral B Cells in Mice with Cd19-Driven Cre Expression. Cells 2022, 11, 700. https://doi.org/10.3390/cells11040700
Zhao Y, Zhao S, Qin X-Y, He T-T, Hu M-M, Gong Z, Wang H-M, Gong F-Y, Gao X-M, Wang J. Altered Phenotype and Enhanced Antibody-Producing Ability of Peripheral B Cells in Mice with Cd19-Driven Cre Expression. Cells. 2022; 11(4):700. https://doi.org/10.3390/cells11040700
Chicago/Turabian StyleZhao, Ying, Sai Zhao, Xiao-Yuan Qin, Ting-Ting He, Miao-Miao Hu, Zheng Gong, Hong-Min Wang, Fang-Yuan Gong, Xiao-Ming Gao, and Jun Wang. 2022. "Altered Phenotype and Enhanced Antibody-Producing Ability of Peripheral B Cells in Mice with Cd19-Driven Cre Expression" Cells 11, no. 4: 700. https://doi.org/10.3390/cells11040700
APA StyleZhao, Y., Zhao, S., Qin, X. -Y., He, T. -T., Hu, M. -M., Gong, Z., Wang, H. -M., Gong, F. -Y., Gao, X. -M., & Wang, J. (2022). Altered Phenotype and Enhanced Antibody-Producing Ability of Peripheral B Cells in Mice with Cd19-Driven Cre Expression. Cells, 11(4), 700. https://doi.org/10.3390/cells11040700