
Pathophysiological Heterogeneity of the BBSOA Neurodevelopmental Syndrome


Abstract
:1. NR2F1 as a Master Regulator of Brain Development and Function
2. From Distinct Cohorts to a Unified List of NR2F1-Haploinsufficient BBSOAS Patients
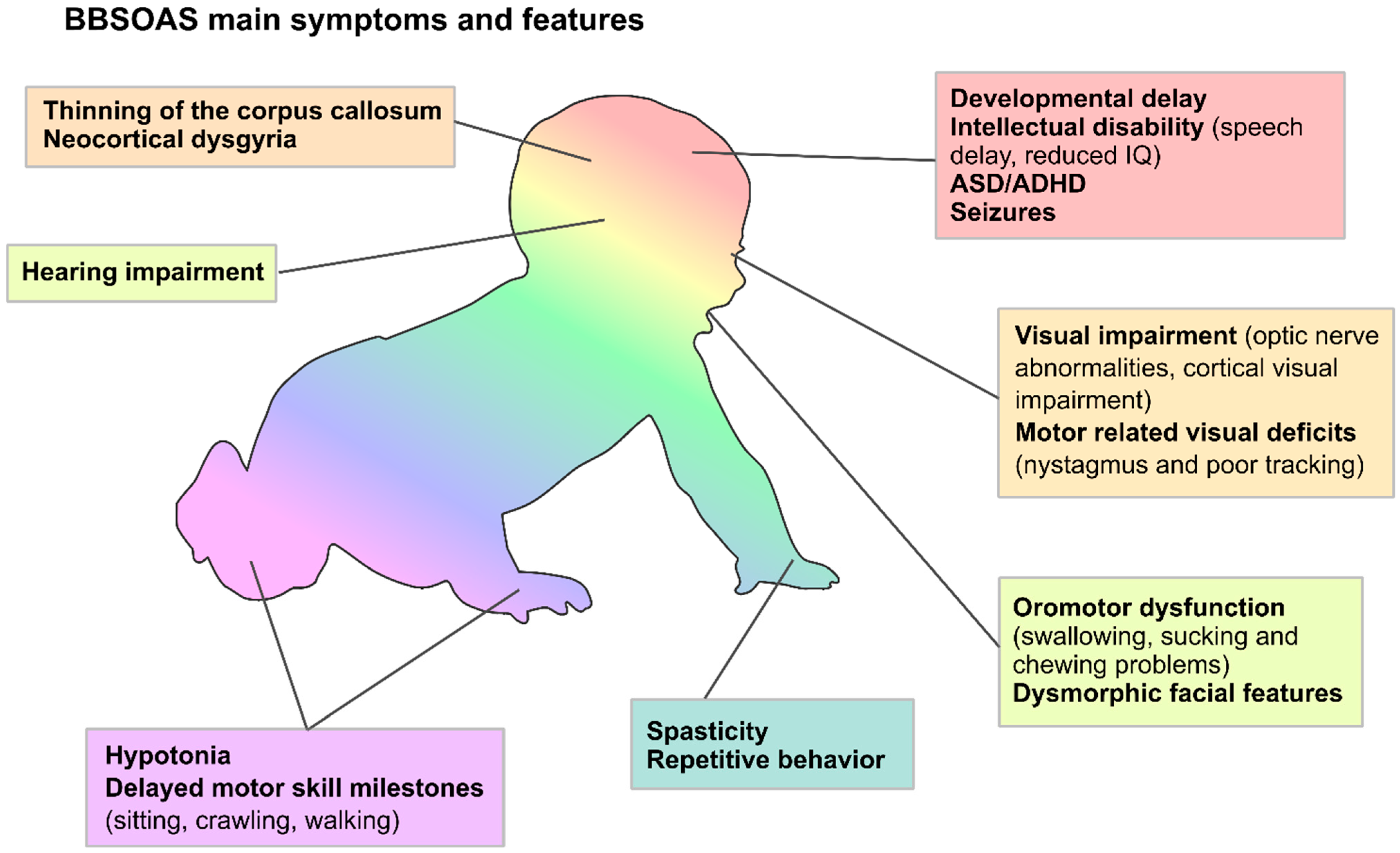
3. Towards a Consensus List of NR2F1-Related Symptoms and Standardized Therapeutic Interventions for BBSOAS Patients
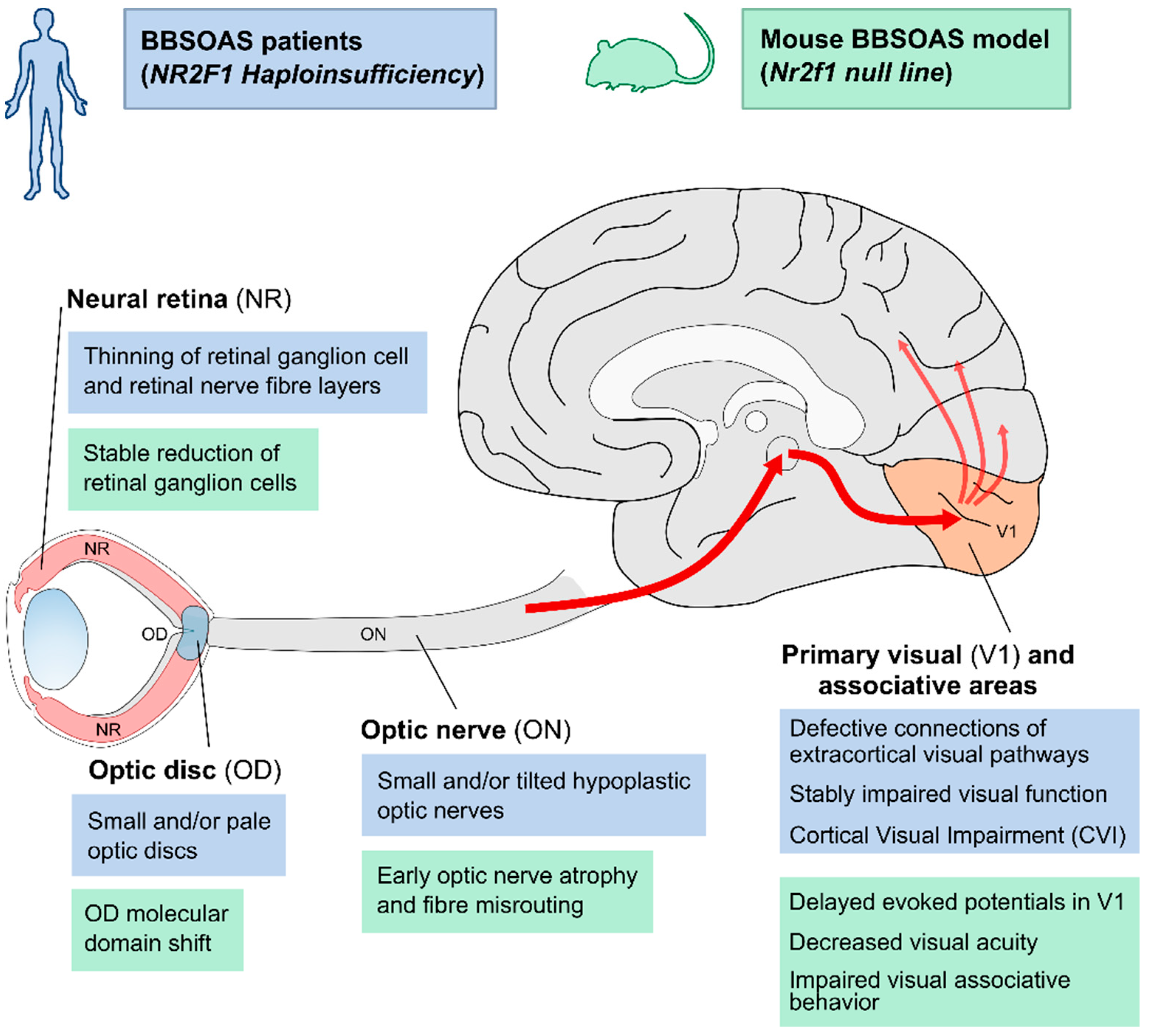
4. Optic Atrophy (OA), Optic Nerve Hypoplasia (ONH) and Non-Progressive Reduction in Visual Acuity: BBSOAS as a Non-Canonical Optic Neuropathy
5. A Genetic Control of Optic Disc Abnormalities in BBSOAS
6. When Intellectual Disability Meets Visual Disfunction: Cortical Visual Impairment (CVI) in BBSOAS Patients
7. The Many Converging Roads of Intellectual Disability in BBSOAS Patients: From Corpus-Callosum Thinning to Hippocampal and Neocortical Malformations
8. NR2F1 as an Autism Spectrum Disorder Gene
9. NR2F1 as a Susceptibility Gene for Infantile Epileptic Disorders
10. NR2F1 on the Move: Motor Dysfunction in BBSOAS Patients
11. Facial Dysmorphia
12. Feeding Problems and Mouth Stuffing, a Common but Poorly Characterized BBSOAS Feature
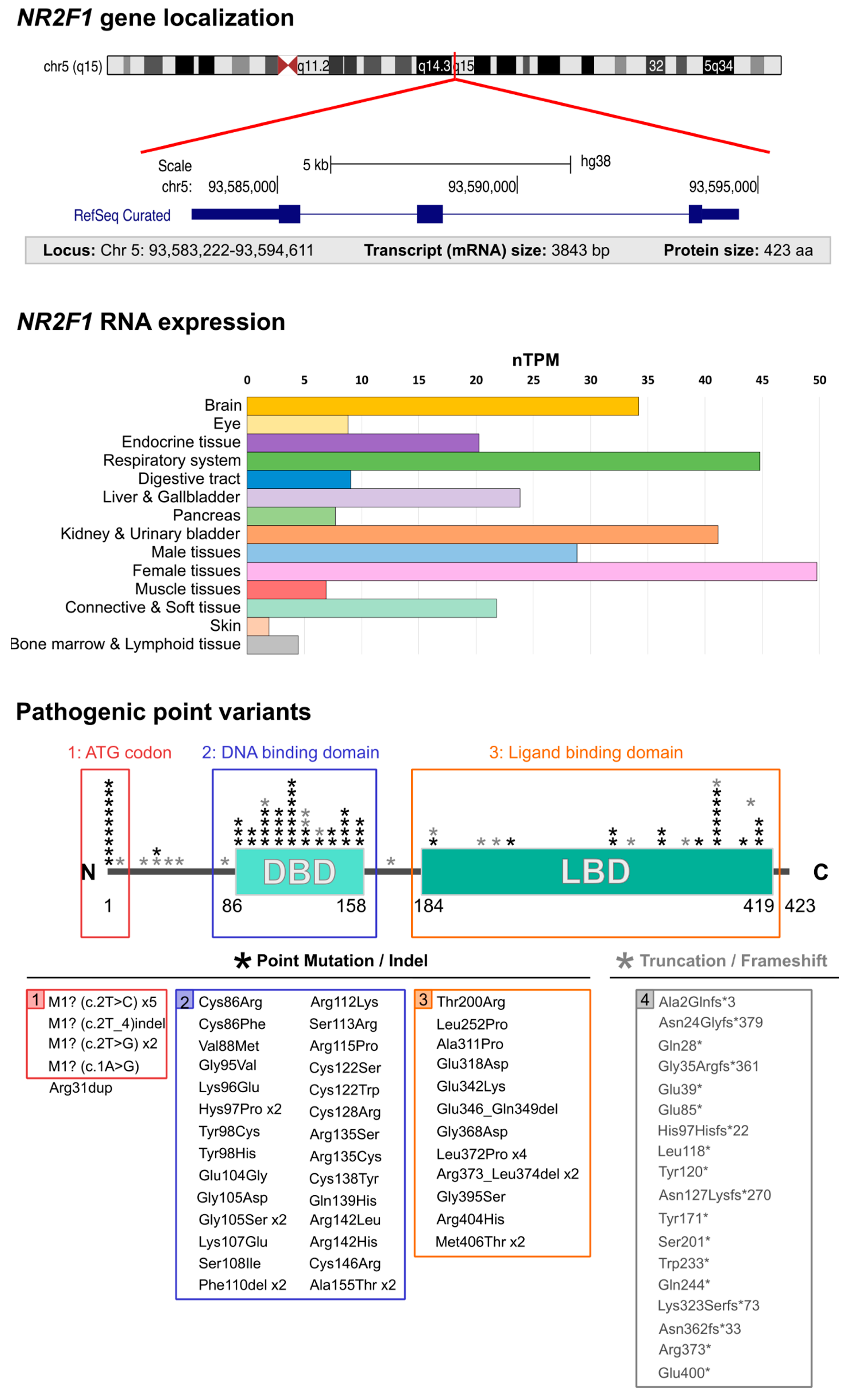
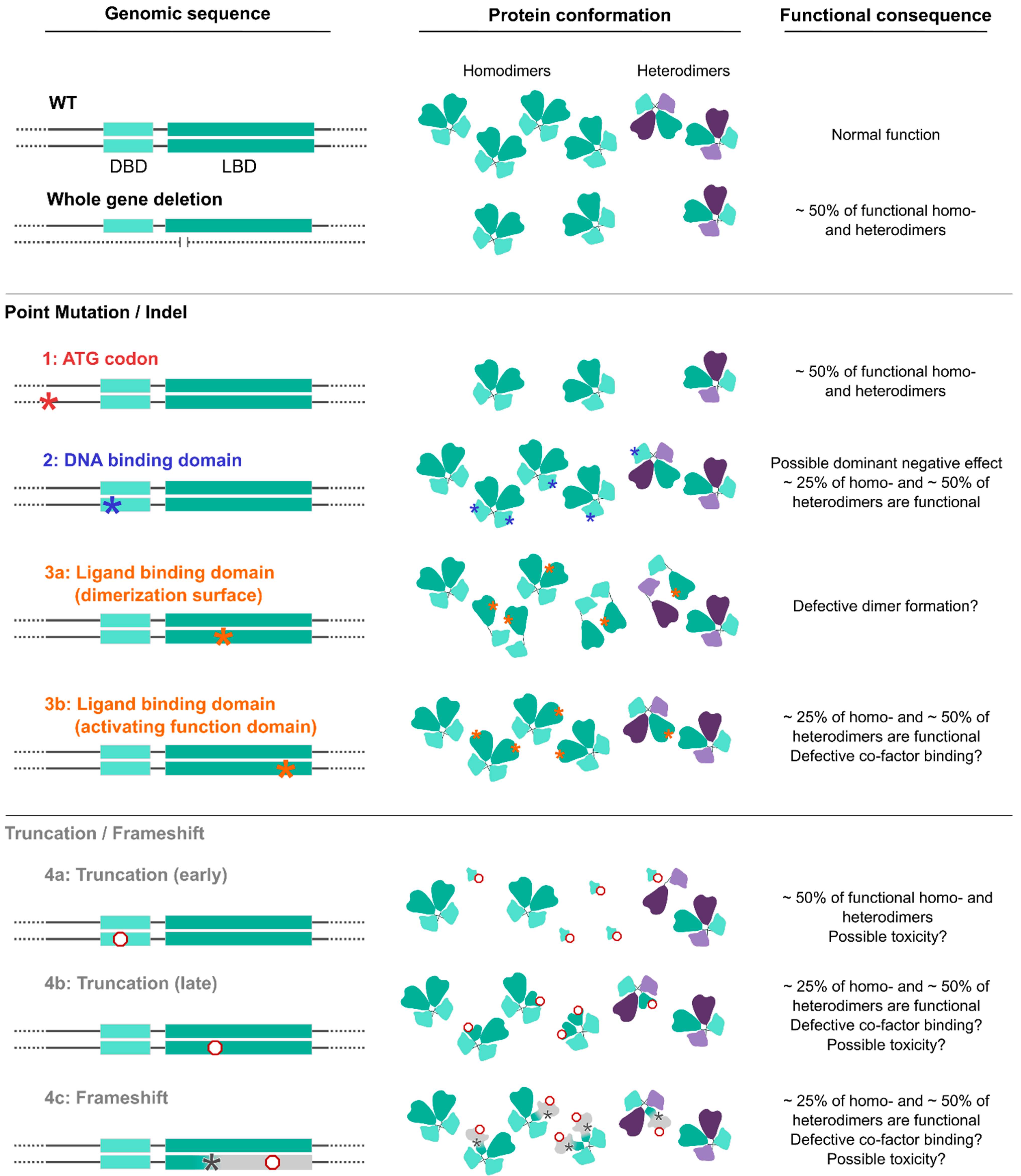
13. The Emerging BBSOAS Genotype–Phenotype Correlation from a Molecular Point of View: NR2F1 Dimers and the Dominant-Negative Effect
- Whole-gene deletions or small indels. (Severity index: 4.33). A few BBSOAS patients so far have been reported with chromosome deletions spanning from 582 Kb to 5 Mb in size, all resulting in whole-gene ablation and complete loss of one NR2F1 allele [1,27,28,29,30,38]. The likely consequence of such deletion is a halved NR2F1 protein production, i.e., haploinsufficiency, as was proven by protein quantification in skin fibroblast extracts [30]. However, it must be noted that large deletions and complex chromosomal aberrations can involve additional genes located next to the NR2F1 locus, such as FAM172A, POU5F2, MIR2277 and lnc-NR2F1, adding to the complexity of the condition, and possibly leading to supplementary congenital abnormalities, such as periventricular heterotopia and deafness, among others [27,28,29]. Interestingly, specific deletions only affecting the region adjacent to the NR2F1 gene, where the long non-coding (lnc)-NR2F1 RNA is located, were also shown to cause neurodevelopmental conditions with developmental and speech delay [213], probably due to the ability of lnc-NR2F1 to control the expression of autism-associated neural genes in a similar way to its corresponding protein-coding gene. These data suggest that (i) multiple genes could synergistically converge to cause similar neurological conditions, and that (ii) the loss of other genes together with NR2F1 in BBSOAS patients with large deletions could exacerbate the clinical features and further increase their heterogeneity.
- Translation initiation variants (TIVs). (Severity index: 5.33). BBSOAS patients with missense variants falling in the translation initiation codon (ATG) show decreased NR2F1 protein resulting from reduced efficiency of both translational and transcriptional processes [30]. Notably, the third codon of the NR2F1 gene sequence is also an ATG, raising the possibility that an alternative initiation site could be present and potentially compensate for the loss or mutation of the main one. However, diminished NR2F1 levels measured in cells from these patients suggest that the second ATG site is not able to efficiently serve as an alternative start codon [30]. The production of a half dosage of the functional NR2F1 protein, without any mutated form competing for dimer formation, could make these patients more similar to BBSOAS individuals with whole-gene deletions than to patients carrying missense DBD/LBD variants, hence leading to milder phenotypes. However, this has been recently questioned by the report of a severe BBSOAS case owing to an NR2F1 start codon variant [85]. While the severity index shows an intermediate severity compared to the two groups, statistical analysis on larger cohorts will be necessary to better detail the phenotypes associated with TIVs.
- DBD missense variants (or DBD in-frame deletions). (Severity index: 5.62). The effect of missense mutations is tightly linked to the structural and functional importance of the affected amino acids and the protein region in which they are located. Based upon evolutionary conservation of distinct aminoacidic positions, bioinformatic prediction software can be used to identify highly conserved regions, and to evaluate whether they are intolerant towards variation and therefore potentially pathogenic [214,215,216]. BBSOAS pathogenic point mutations falling in the DBD and impacting the 3D structure of evolutionarily conserved sites in the zinc-finger domains have strong effects on NR2F1 structure and function, and consequently, on the clinical phenotype. As an example, substitution of a highly conserved zinc-finger motif of the DBD of NR2F1 leads to heavy changes in molecular structure and stability, as predicted in silico [36]. By contrast, missense variants located in less conserved regions adjacent to the DBD showed only a reduced transcriptional activity in the luciferase assay, in contrast to the almost abolished activity of DBD variants falling in crucial zinc-finger or structural sites [30]. As discussed above, the high penetrance of BBSOAS features following DBD missense mutations, compared to the complete loss of one allele by whole-gene deletion, could result from a possible dominant-negative effect of mutated NR2F1. Indeed, by forming non-functional dimers with the wild-type NR2F1 (produced by the normal allele), mutated NR2F1 proteins could ultimately affect 75% of the total pool of NR2F1 dimers in the cell (Figure 4). Heterodimer formation with other nuclear receptors (NRs) could be affected too, presumably resulting in a 50% loss of functional NR2F1-NRs heterodimers (Figure 4). The high number of reported DBD missense mutations, associated with the highest severity index among BBSOAS genetic groups, confirms the functional relevance of this region.
- LBD missense variants. (Severity index: 3.76). Patients with variants in the LBD manifest milder developmental delay, often lacking hypotonia, speech defects, seizures and repetitive behaviors [35], consistent with the lowest severity index among BBSOAS genetic categories. As the LBD is necessary for NR2F1 dimerization, it is reasonable to think that mutations falling on interacting surfaces could hamper dimer formation. While this could decrease NR2F1 activity in general, such a situation could also affect the formation of dimers between normal and mutated NR2F1, hence resembling whole-gene deletions, with no (or lower) dominant-negative effect. The LBD also contains the Activating Function domain 2 (AF-2), which is necessary for co-factor binding. While the exact nature of NR2F1 protein partners in neural cells remains elusive, the loss of such interactions following LBD mutations could be responsible for some BBSOAS features. Further studies will be necessary to dissect the specific function of distinct domains in the LBD and the impact of distinct mutations on dimer formation, as well as the identity of NR2F1 physiological co-factors. This will lead to further insights into NR2F1 molecular functions and how they correlate with patients’ symptoms.
- Truncation or frameshift followed by truncation variants. (Severity index: 5.18 and 5.29, respectively). Nonsense mutations result in premature termination of the protein (i.e., truncation of the peptide sequence) and can have profound impacts on gene function [217]. Obviously, the portion of the protein that is lost depends on the precise location of the aberrant termination site. The low number of described patients carrying NR2F1 truncations makes it challenging to gain a clear picture of the genotype–phenotype correlation for those pathological cases. However, it is reasonable to think that early truncations would trigger nonsense-mediated decay of the mRNA, resulting in loss-of-function, whereas late truncation variants that escape nonsense-mediated decay may create an abnormal, but stable, truncated protein, causing dominant-negative or neomorphic effects [218]. Hence, early variants leading to haploinsufficiency would be more similar to whole-gene deletions and may also protect against more detrimental phenotypic effects (i.e., dominant-negative effect), presumably leading to milder phenotypes [218,219]. Consistently, a recently reported patient presenting NR2F1 early truncation showed a mild phenotype [40], whereby one case has high-functioning ASD with superior verbal abilities [33]. By contrast, late variants that do not affect the dimerization region of the LBD could show a dominant-negative effect, similarly to what is reported for point variants, and ultimately leading to highly penetrant symptoms. Taking all truncation variants together, their collective severity index is more severe than that of deletions, similarly to variants affecting the starting codon.
14. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Bosch, D.G.M.; Boonstra, F.N.; Gonzaga-Jauregui, C.; Xu, M.; de Ligt, J.; Jhangiani, S.; Wiszniewski, W.; Muzny, D.M.; Yntema, H.G.; Pfundt, R.; et al. NR2F1 Mutations Cause Optic Atrophy with Intellectual Disability. Am. J. Hum. Genet. 2014, 94, 303–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C. COUP-TFI: An Intrinsic Factor for Early Regionalization of the Neocortex. Genes Dev. 2001, 15, 2054–2059. [Google Scholar] [CrossRef] [Green Version]
- Bertacchi, M.; Parisot, J.; Studer, M. The Pleiotropic Transcriptional Regulator COUP-TFI Plays Multiple Roles in Neural Development and Disease. Brain Res. 2018, 1705, 75–94. [Google Scholar] [CrossRef]
- Tang, K.; Tsai, S.Y.; Tsai, M.-J. COUP-TFs and Eye Development. Biochim. Biophys. Acta 2015, 1849, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Tocco, C.; Bertacchi, M.; Studer, M. Structural and Functional Aspects of the Neurodevelopmental Gene NR2F1: From Animal Models to Human Pathology. Front. Mol. Neurosci. 2021, 14, 767965. [Google Scholar] [CrossRef]
- Alfano, C.; Magrinelli, E.; Harb, K.; Hevner, R.F.; Studer, M. Postmitotic Control of Sensory Area Specification during Neocortical Development. Nat. Commun. 2014, 5, 5632. [Google Scholar] [CrossRef] [Green Version]
- Armentano, M.; Chou, S.-J.; Srubek Tomassy, G.; Leingärtner, A.; O’Leary, D.D.M.; Studer, M. COUP-TFI Regulates the Balance of Cortical Patterning between Frontal/Motor and Sensory Areas. Nat. Neurosci. 2007, 10, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Bertacchi, M.; Romano, A.L.; Loubat, A.; Tran Mau-Them, F.; Willems, M.; Faivre, L.; Khau van Kien, P.; Perrin, L.; Devillard, F.; Sorlin, A.; et al. NR2F1 Regulates Regional Progenitor Dynamics in the Mouse Neocortex and Cortical Gyrification in BBSOAS Patients. EMBO J. 2020, 39, e104163. [Google Scholar] [CrossRef]
- Faedo, A.; Tomassy, G.S.; Ruan, Y.; Teichmann, H.; Krauss, S.; Pleasure, S.J.; Tsai, S.Y.; Tsai, M.-J.; Studer, M.; Rubenstein, J.L.R. COUP-TFI Coordinates Cortical Patterning, Neurogenesis, and Laminar Fate and Modulates MAPK/ERK, AKT, and ß-Catenin Signaling. Cereb. Cortex 2008, 18, 2117–2131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomassy, G.S.; De Leonibus, E.; Jabaudon, D.; Lodato, S.; Alfano, C.; Mele, A.; Macklis, J.D.; Studer, M. Area-Specific Temporal Control of Corticospinal Motor Neuron Differentiation by COUP-TFI. Proc. Natl. Acad. Sci. USA 2010, 107, 3576–3581. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-A.; Wang, W.; Pedersen, S.E.; Raman, A.; Seymour, M.L.; Ruiz, F.R.; Xia, A.; van der Heijden, M.E.; Wang, L.; Yin, J.; et al. Nr2f1 Heterozygous Knockout Mice Recapitulate Neurological Phenotypes of Bosch-Boonstra-Schaaf Optic Atrophy Syndrome and Show Impaired Hippocampal Synaptic Plasticity. Hum. Mol. Genet. 2020, 29, 705–715. [Google Scholar] [CrossRef]
- Flore, G.; Di Ruberto, G.; Parisot, J.; Sannino, S.; Russo, F.; Illingworth, E.A.; Studer, M.; De Leonibus, E. Gradient COUP-TFI Expression Is Required for Functional Organization of the Hippocampal Septo-Temporal Longitudinal Axis. Cereb. Cortex 2016, 27, bhv336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisot, J.; Flore, G.; Bertacchi, M.; Studer, M. COUP-TFI Mitotically Regulates Production and Migration of Dentate Granule Cells and Modulates Hippocampal Cxcr4 Expression. Dev. Camb. Engl. 2017, 144, 2045–2058. [Google Scholar] [CrossRef] [Green Version]
- Lodato, S.; Tomassy, G.S.; De Leonibus, E.; Uzcategui, Y.G.; Andolfi, G.; Armentano, M.; Touzot, A.; Gaztelu, J.M.; Arlotta, P.; Menendez de la Prida, L.; et al. Loss of COUP-TFI Alters the Balance between Caudal Ganglionic Eminence- and Medial Ganglionic Eminence-Derived Cortical Interneurons and Results in Resistance to Epilepsy. J. Neurosci. 2011, 31, 4650–4662. [Google Scholar] [CrossRef]
- Touzot, A.; Ruiz-Reig, N.; Vitalis, T.; Studer, M. Molecular Control of Two Novel Migratory Paths for CGE-Derived Interneurons in the Developing Mouse Brain. Development 2016, 143, 1753–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertacchi, M.; Gruart, A.; Kaimakis, P.; Allet, C.; Serra, L.; Giacobini, P.; Delgado-García, J.M.; Bovolenta, P.; Studer, M. Mouse Nr2f1 Haploinsufficiency Unveils New Pathological Mechanisms of a Human Optic Atrophy Syndrome. EMBO Mol. Med. 2019, 11, e10291. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.; Xie, X.; Park, J.-I.; Jamrich, M.; Tsai, S.; Tsai, M.-J. COUP-TFs Regulate Eye Development by Controlling Factors Essential for Optic Vesicle Morphogenesis. Development 2010, 137, 725–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naka, H.; Nakamura, S.; Shimazaki, T.; Okano, H. Requirement for COUP-TFI and II in the Temporal Specification of Neural Stem Cells in CNS Development. Nat. Neurosci. 2008, 11, 1014–1023. [Google Scholar] [CrossRef]
- Okano, H.; Temple, S. Cell Types to Order: Temporal Specification of CNS Stem Cells. Curr. Opin. Neurobiol. 2009, 19, 112–119. [Google Scholar] [CrossRef]
- del Pino, I.; Tocco, C.; Magrinelli, E.; Marcantoni, A.; Ferraguto, C.; Tomagra, G.; Bertacchi, M.; Alfano, C.; Leinekugel, X.; Frick, A.; et al. COUP-TFI/Nr2f1 Orchestrates Intrinsic Neuronal Activity during Cortical Area Patterning. bioRxiv 2019. [CrossRef]
- Alzu’bi, A.; Lindsay, S.; Kerwin, J.; Looi, S.J.; Khalil, F.; Clowry, G.J. Distinct Cortical and Sub-Cortical Neurogenic Domains for GABAergic Interneuron Precursor Transcription Factors NKX2.1, OLIG2 and COUP-TFII in Early Fetal Human Telencephalon. Brain Struct. Funct. 2017, 222, 2309–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alzu’bi, A.; Lindsay, S.J.; Harkin, L.F.; McIntyre, J.; Lisgo, S.N.; Clowry, G.J. The Transcription Factors COUP-TFI and COUP-TFII Have Distinct Roles in Arealisation and GABAergic Interneuron Specification in the Early Human Fetal Telencephalon. Cereb. Cortex 2017, 27, 4971–4987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foglio, B.; Rossini, L.; Garbelli, R.; Regondi, M.C.; Mercurio, S.; Bertacchi, M.; Avagliano, L.; Bulfamante, G.; Coras, R.; Maiorana, A.; et al. Dynamic Expression of NR2F1 and SOX2 in Developing and Adult Human Cortex: Comparison with Cortical Malformations. Brain Struct. Funct. 2021, 226, 1303–1322. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Feng, S.; Tang, K. COUP—TF Genes, Human Diseases, and the Development of the Central Nervous System in Murine Models. In Current Topics in Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 125, pp. 275–301. ISBN 978-0-12-802172-9. [Google Scholar]
- Michaud, J.L.; Lachance, M.; Hamdan, F.F.; Carmant, L.; Lortie, A.; Diadori, P.; Major, P.; Meijer, I.A.; Lemyre, E.; Cossette, P.; et al. The Genetic Landscape of Infantile Spasms. Hum. Mol. Genet. 2014, 23, 4846–4858. [Google Scholar] [CrossRef] [Green Version]
- Sanders, S.J.; Murtha, M.T.; Gupta, A.R.; Murdoch, J.D.; Raubeson, M.J.; Willsey, A.J.; Ercan-Sencicek, A.G.; DiLullo, N.M.; Parikshak, N.N.; Stein, J.L.; et al. De Novo Mutations Revealed by Whole-Exome Sequencing Are Strongly Associated with Autism. Nature 2012, 485, 237–241. [Google Scholar] [CrossRef]
- Al-Kateb, H.; Shimony, J.S.; Vineyard, M.; Manwaring, L.; Kulkarni, S.; Shinawi, M. NR2F1 Haploinsufficiency Is Associated with Optic Atrophy, Dysmorphism and Global Developmental Delay. Am. J. Med. Genet. A 2013, 161A, 377–381. [Google Scholar] [CrossRef]
- Brown, K.K.; Alkuraya, F.S.; Matos, M.; Robertson, R.L.; Kimonis, V.E.; Morton, C.C. NR2F1 Deletion in a Patient with a de Novo Paracentric Inversion, Inv(5)(Q15q33.2), and Syndromic Deafness. Am. J. Med. Genet. A 2009, 149A, 931–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, C.; Boys, A.; Parrini, E.; Mignon-Ravix, C.; McMahon, J.M.; Khantane, S.; Bertini, E.; Pallesi, E.; Missirian, C.; Zuffardi, O.; et al. Periventricular Heterotopia, Mental Retardation, and Epilepsy Associated with 5q14.3-Q15 Deletion. Neurology 2009, 72, 784–792. [Google Scholar] [CrossRef]
- Chen, C.-A.; Bosch, D.G.M.; Cho, M.T.; Rosenfeld, J.A.; Shinawi, M.; Lewis, R.A.; Mann, J.; Jayakar, P.; Payne, K.; Walsh, L.; et al. The Expanding Clinical Phenotype of Bosch-Boonstra-Schaaf Optic Atrophy Syndrome: 20 New Cases and Possible Genotype-Phenotype Correlations. Genet. Med. Off. J. Am. Coll. Med. Genet. 2016, 18, 1143–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrahams, B.S.; Arking, D.E.; Campbell, D.B.; Mefford, H.C.; Morrow, E.M.; Weiss, L.A.; Menashe, I.; Wadkins, T.; Banerjee-Basu, S.; Packer, A. SFARI Gene 2.0: A Community-Driven Knowledgebase for the Autism Spectrum Disorders (ASDs). Mol. Autism 2013, 4, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billiet, B.; Amati-Bonneau, P.; Desquiret-Dumas, V.; Guehlouz, K.; Milea, D.; Gohier, P.; Lenaers, G.; Mirebeau-Prunier, D.; den Dunnen, J.T.; Reynier, P.; et al. NR2F1 Database: 112 Variants and 84 Patients Support Refining the Clinical Synopsis of Bosch–Boonstra–Schaaf Optic Atrophy Syndrome. Hum. Mutat. 2021, 43, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Bojanek, E.K.; Mosconi, M.W.; Guter, S.; Betancur, C.; Macmillan, C.; Cook, E.H. Clinical and Neurocognitive Issues Associated with Bosch-Boonstra-Schaaf Optic Atrophy Syndrome: A Case Study. Am. J. Med. Genet. Part A 2019, 182, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Hino-Fukuyo, N.; Kikuchi, A.; Yokoyama, H.; Iinuma, K.; Hirose, M.; Haginoya, K.; Niihori, T.; Nakayama, K.; Aoki, Y.; Kure, S. Long-Term Outcome of a 26-Year-Old Woman with West Syndrome and an Nuclear Receptor Subfamily 2 Group F Member 1 Gene (NR2F1) Mutation. Seizure 2017, 50, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Kaiwar, C.; Zimmermann, M.T.; Ferber, M.J.; Niu, Z.; Urrutia, R.A.; Klee, E.W.; Babovic-Vuksanovic, D. Novel NR2F1 Variants Likely Disrupt DNA Binding: Molecular Modeling in Two Cases, Review of Published Cases, Genotype-Phenotype Correlation, and Phenotypic Expansion of the Bosch-Boonstra-Schaaf Optic Atrophy Syndrome. Cold Spring Harb. Mol. Case Stud. 2017, 3, a002162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mio, C.; Fogolari, F.; Pezzoli, L.; D’Elia, A.V.; Iascone, M.; Damante, G. Missense NR2F1 Variant in Monozygotic Twins Affected with the Bosch–Boonstra–Schaaf Optic Atrophy Syndrome. Mol. Genet. Genom. Med. 2020, 8, e1278. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Lee, J.S.; Lee, S.-T.; Kim, H.Y.; Han, S.-H.; Han, J. Targeted Panel Sequencing Identifies a Novel NR2F1 Mutations in a Patient with Bosch–Boonstra–Schaaf Optic Atrophy Syndrome. Ophthalmic Genet. 2019, 40, 359–361. [Google Scholar] [CrossRef] [PubMed]
- Rech, M.E.; McCarthy, J.M.; Chen, C.-A.; Edmond, J.C.; Shah, V.S.; Bosch, D.G.M.; Berry, G.T.; Williams, L.; Madan-Khetarpal, S.; Niyazov, D.; et al. Phenotypic Expansion of Bosch-Boonstra-Schaaf Optic Atrophy Syndrome and Further Evidence for Genotype-Phenotype Correlations. Am. J. Med. Genet. Part A 2020, 182, 1426–1437. [Google Scholar] [CrossRef] [Green Version]
- Walsh, S.; Gösswein, S.S.; Rump, A.; von der Hagen, M.; Hackmann, K.; Schröck, E.; Di Donato, N.; Kahlert, A.-K. Novel Dominant-Negative NR2F1 Frameshift Mutation and a Phenotypic Expansion of the Bosch-Boonstra-Schaaf Optic Atrophy Syndrome. Eur. J. Med. Genet. 2020, 63, 104019. [Google Scholar] [CrossRef]
- Zou, W.; Cheng, L.; Lu, S.; Wu, Z. A de Novo Nonsense Mutation in the N-Terminal of Ligand-Binding Domain of NR2F1 Gene Provoked a Milder Phenotype of BBSOAS. Ophthalmic Genet. 2020, 41, 88–89. [Google Scholar] [CrossRef]
- Jurkute, N.; Bertacchi, M.; Arno, G.; Tocco, C.; Kim, U.S.; Kruszewski, A.M.; Avery, R.A.; Bedoukian, E.C.; Han, J.; Ahn, S.J.; et al. Pathogenic NR2F1 Variants Cause a Developmental Ocular Phenotype Recapitulated in a Mutant Mouse Model. Brain Commun. 2021, 3, fcab162. [Google Scholar] [CrossRef]
- Hobbs, M.M.; Wolters, W.C.; Rayapati, A.O. Bosch-Boonstra-Schaaf Optic Atrophy Syndrome Presenting as New-Onset Psychosis in a 32-Year-Old Man: A Case Report and Literature Review. J. Psychiatr. Pract. 2020, 26, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hernández, E.; Rodríguez-García, M.E.; Chen, C.-A.; Cotrina-Vinagre, F.J.; Carnicero-Rodríguez, P.; Bellusci, M.; Schaaf, C.P.; Martínez-Azorín, F. Mitochondrial Involvement in a Bosch-Boonstra-Schaaf Optic Atrophy Syndrome Patient with a Novel de Novo NR2F1 Gene Mutation. J. Hum. Genet. 2018, 63, 525–528. [Google Scholar] [CrossRef] [PubMed]
- Starosta, R.T.; Tarnowski, J.; e Vairo, F.P.; Raymond, K.; Preston, G.; Morava, E. Bosch-Boonstra-Schaaf Optic Atrophy Syndrome (BBSOAS) Initially Diagnosed as ALG6-CDG: Functional Evidence for Benignity of the ALG6 c.391T>C (p.Tyr131His) Variant and Further Expanding the BBSOAS Phenotype. Eur. J. Med. Genet. 2020, 63, 103941. [Google Scholar] [CrossRef] [PubMed]
- Yu-Wai-Man, P.; Griffiths, P.G.; Chinnery, P.F. Mitochondrial Optic Neuropathies—Disease Mechanisms and Therapeutic Strategies. Prog. Retin. Eye Res. 2011, 30, 81–114. [Google Scholar] [CrossRef] [Green Version]
- Jurkute, N.; Majander, A.; Bowman, R.; Votruba, M.; Abbs, S.; Acheson, J.; Lenaers, G.; Amati-Bonneau, P.; Moosajee, M.; Arno, G.; et al. Clinical Utility Gene Card for: Inherited Optic Neuropathies Including next-Generation Sequencing-Based Approaches. Eur. J. Hum. Genet. 2018, 27, 494–502. [Google Scholar] [CrossRef]
- Yu-Wai-Man, P.; Bailie, M.; Atawan, A.; Chinnery, P.F.; Griffiths, P.G. Pattern of Retinal Ganglion Cell Loss in Dominant Optic Atrophy Due to OPA1 Mutations. Eye 2011, 25, 596–602. [Google Scholar] [CrossRef] [Green Version]
- Pang, Y.; Frantz, K.A.; Roberts, D.K. Association of Refractive Error with Optic Nerve Hypoplasia. Ophthalmic Physiol. Opt. J. Br. Coll. Ophthalmic Opt. Optom. 2015, 35, 570–576. [Google Scholar] [CrossRef]
- Dahl, S.; Pettersson, M.; Eisfeldt, J.; Schröder, A.K.; Wickström, R.; Fahnehjelm, K.T.; Anderlid, B.M.; Lindstrand, A. Whole Genome Sequencing Unveils Genetic Heterogeneity in Optic Nerve Hypoplasia. PLoS ONE 2020, 15, 1–16. [Google Scholar] [CrossRef]
- Karahan, E.; Tulin Berk, A. Ocular, Neurologic and Systemic Findings of the Cases with Optic Nerve Hypoplasia. Open Ophthalmol. J. 2016, 10, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Mosier, M.A.; Lieberman, M.F.; Green, W.R.; Knox, D.L. Hypoplasia of the Optic Nerve. Arch. Ophthalmol. Chic. Ill 1960 1978, 96, 1437–1442. [Google Scholar] [CrossRef]
- Taylor, D. Developmental Abnormalities of the Optic Nerve and Chiasm. Eye Lond. Engl. 2007, 21, 1271–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, S.; Jain, S.; Sodhi, H.B.S.; Rastogi, A. Kamlesh Optic Nerve Hypoplasia. Oman J. Ophthalmol. 2013, 6, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Pilat, A.; Sibley, D.; McLean, R.J.; Proudlock, F.A.; Gottlob, I. High-Resolution Imaging of the Optic Nerve and Retina in Optic Nerve Hypoplasia. Ophthalmology 2015, 122, 1330–1339. [Google Scholar] [CrossRef] [Green Version]
- Hoyt, C.S.; Good, W.V. Do We Really Understand the Difference between Optic Nerve Hypoplasia and Atrophy? Eye 1992, 6, 201–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.; Al-Hayouti, H.; Oladiwura, D.; Karim, R.; Sawczenko, A.; Dahlmann-Noor, A. Optic Atrophy in Children: Current Causes and Diagnostic Approach. Eur. J. Ophthalmol. 2020, 30, 1499–1505. [Google Scholar] [CrossRef] [PubMed]
- McClard, C.K.; Shah, V. Pediatric Optic Disc Pallor. Int. Ophthalmol. Clin. 2018, 58, 125–145. [Google Scholar] [CrossRef] [PubMed]
- Yu-Wai-Man, P.; Votruba, M.; Burté, F.; La Morgia, C.; Barboni, P.; Carelli, V.; Burte, F.; La Morgia, C.; Barboni, P.; Carelli, V.; et al. A Neurodegenerative Perspective on Mitochondrial Optic Neuropathies. Acta Neuropathol. 2016, 132, 789–806. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, P.E.; Lopez, M.J.; Czyz, C.N. Embryology, Eye Malformations. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Ryabets-Lienhard, A.; Stewart, C.; Borchert, M.; Geffner, M.E. The Optic Nerve Hypoplasia Spectrum: Review of the Literature and Clinical Guidelines. Adv. Pediatr. 2016, 63, 127–146. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Filion, P.; Borchert, M. Optic Nerve Hypoplasia Syndrome: A Review of the Epidemiology and Clinical Associations. Curr. Treat. Options Neurol. 2013, 15, 78–89. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Filion, P.; Epport, K.; Nelson, M.; Azen, C.; Geffner, M.E.; Fink, C.; Borchert, M. Neuroradiographic, Endocrinologic, and Ophthalmic Correlates of Adverse Developmental Outcomes in Children with Optic Nerve Hypoplasia: A Prospective Study. Pediatrics 2008, 121, e653–e659. [Google Scholar] [CrossRef] [Green Version]
- Azuma, N.; Yamaguchi, Y.; Handa, H.; Tadokoro, K.; Asaka, A.; Kawase, E.; Yamada, M. Mutations of the PAX6 Gene Detected in Patients with a Variety of Optic-Nerve Malformations. Am. J. Hum. Genet. 2003, 72, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.-A.; Yin, J.; Lewis, R.A.; Schaaf, C.P. Genetic Causes of Optic Nerve Hypoplasia. J. Med. Genet. 2017, 54, 441–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelberman, D. Mutations within Sox2/SOX2 Are Associated with Abnormalities in the Hypothalamo-Pituitary-Gonadal Axis in Mice and Humans. J. Clin. Investig. 2006, 116, 2442–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfano, C. The Nuclear Receptors COUP-TF: A Long-Lasting Experience in Forebrain Assembly. Cell. Mol. Life Sci. 2014, 71, 43–62. [Google Scholar] [CrossRef]
- Yamaguchi, H.; Zhou, C.; Lin, S.-C.; Durand, B.; Tsai, S.Y.; Tsai, M.-J. The Nuclear Orphan Receptor COUP-TFI Is Important for Differentiation of Oligodendrocytes. Dev. Biol. 2004, 266, 238–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eleuteri, C.; Olla, S.; Veroni, C.; Umeton, R.; Mechelli, R.; Romano, S.; Buscarinu, M.; Ferrari, F.; Calò, G.; Ristori, G.; et al. A Staged Screening of Registered Drugs Highlights Remyelinating Drug Candidates for Clinical Trials. Sci. Rep. 2017, 7, 45780. [Google Scholar] [CrossRef] [Green Version]
- Harlow, D.E.; Honce, J.M.; Miravalle, A.A. Remyelination Therapy in Multiple Sclerosis. Front. Neurol. 2015, 6, 257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najm, F.J.; Madhavan, M.; Zaremba, A.; Shick, E.; Karl, R.T.; Factor, D.C.; Miller, T.E.; Nevin, Z.S.; Kantor, C.; Sargent, A.; et al. Drug-Based Modulation of Endogenous Stem Cells Promotes Functional Remyelination In Vivo. Nature 2015, 522, 216–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porcu, G.; Serone, E.; De Nardis, V.; Di Giandomenico, D.; Lucisano, G.; Scardapane, M.; Poma, A.; Ragnini-Wilson, A. Clobetasol and Halcinonide Act as Smoothened Agonists to Promote Myelin Gene Expression and RxRγ Receptor Activation. PLoS ONE 2015, 10, e0144550. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Tang, W.; Luan, Z.; Yang, Y.; Wang, Z.; Zhang, Y.; Wang, Q.; Suo, L.; Huang, Z.; Wang, X.; et al. Protective Effect of Miconazole on Rat Myelin Sheaths Following Premature Infant Cerebral White Matter Injury. Exp. Ther. Med. 2018, 15, 2443–2449. [Google Scholar] [CrossRef] [PubMed]
- Abel, A.; McClelland, C.; Lee, M.S. Critical Review: Typical and Atypical Optic Neuritis. Surv. Ophthalmol. 2019, 64, 770–779. [Google Scholar] [CrossRef] [PubMed]
- Redler, Y.; Levy, M. Rodent Models of Optic Neuritis. Front. Neurol. 2020, 11, 580951. [Google Scholar] [CrossRef]
- Teixeira, L.B.C.; Hoeve, J.N.V.; Mayer, J.A.; Dubielzig, R.R.; Smith, C.M.; Radcliff, A.B.; Duncan, I.D. Modeling the Chronic Loss of Optic Nerve Axons and the Effects on the Retinal Nerve Fiber Layer Structure in Primary Disorder of Myelin. Investig. Opthalmol. Vis. Sci. 2016, 57, 4859. [Google Scholar] [CrossRef] [PubMed]
- Satoh, S.; Tang, K.; Iida, A.; Inoue, M.; Kodama, T.; Tsai, S.Y.; Tsai, M.-J.; Furuta, Y.; Watanabe, S. The Spatial Patterning of Mouse Cone Opsin Expression Is Regulated by Bone Morphogenetic Protein Signaling through Downstream Effector COUP-TF Nuclear Receptors. J. Neurosci. 2009, 29, 12401–12411. [Google Scholar] [CrossRef] [PubMed]
- Inoue, M.; Iida, A.; Satoh, S.; Kodama, T.; Watanabe, S. COUP-TFI and -TFII Nuclear Receptors Are Expressed in Amacrine Cells and Play Roles in Regulating the Differentiation of Retinal Progenitor Cells. Exp. Eye Res. 2010, 90, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Jonas, J.B.; Budde, W.M.; Panda-Jonas, S. Ophthalmoscopic Evaluation of the Optic Nerve Head. Surv. Ophthalmol. 1999, 43, 293–320. [Google Scholar] [CrossRef]
- Rodenbeck, S.J.; Mackay, D.D. Examining the Ocular Fundus in Neurology. Curr. Opin. Neurol. 2019, 32, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Cekić, S.; Stanković-Babić, G.; Višnjić, Z.; Jovanović, I.; Risimić, D. Optic Disc Abnormalities—Diagnosis, Evolution and Influence on Visual Acuity. Bosn. J. Basic Med. Sci. 2010, 10, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutton, G.N. Congenital Disorders of the Optic Nerve: Excavations and Hypoplasia. Eye 2004, 18, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Jeng-Miller, K.W.; Cestari, D.M.; Gaier, E.D. Congenital Anomalies of the Optic Disc: Insights from Optical Coherence Tomography Imaging. Curr. Opin. Ophthalmol. 2017, 28, 579–586. [Google Scholar] [CrossRef]
- Anglo, M. Topical Review: Optic Disc Pits and Associated Maculopathy. Optom. Vis. Sci. 2020, 97, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Uzel, M.M.; Karacorlu, M. Optic Disk Pits and Optic Disk Pit Maculopathy: A Review. Surv. Ophthalmol. 2019, 64, 595–607. [Google Scholar] [CrossRef]
- Gazdagh, G.; Mawby, R.; Self, J.E.; Baralle, D. Deciphering Developmental Disorders Study A Severe Case of Bosch– Boonstra–Schaaf Optic Atrophy Syndrome with a Novel Description of Coloboma and Septo-optic Dysplasia, Owing to a Start Codon Variant in the NR2F1 Gene. Am. J. Med. Genet. A 2022, 188, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Deiner, M.S.; Kennedy, T.E.; Fazeli, A.; Serafini, T.; Tessier-Lavigne, M.; Sretavan, D.W. Netrin-1 and DCC Mediate Axon Guidance Locally at the Optic Disc: Loss of Function Leads to Optic Nerve Hypoplasia. Neuron 1997, 19, 575–589. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, J. Proper Patterning of the Optic Fissure Requires the Sequential Activity of BMP7 and SHH. Development 2006, 133, 3179–3190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, G.W.; Edwards, A.O.; SchimentiI, L.A.; Manligas, G.S.; Zhang, Y.; Ritter, R. Renal-Coloboma Syndrome: Report of a Novel PAX2 Gene Mutation. Am. J. Ophthalmol. 2001, 132, 910–914. [Google Scholar] [CrossRef]
- Eccles, M.R.; Schimmenti, L.A. Renal-Coloboma Syndrome: A Multi-System Developmental Disorder Caused by PAX2 Mutations: PAX2 Mutation in Renal-Coloboma Syndrome. Clin. Genet. 1999, 56, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Good, V.; Jan, J.E.; Barrovich, A.J.; Hoyt, S. Cortical Visual Impairment in Children. 14. Surv. Ophthalmol. 1994, 38, 351–364. [Google Scholar] [CrossRef]
- Good, W.V.; Jan, J.E.; Burden, S.K.; Skoczenski, A.; Candy, R. Recent Advances in Cortical Visual Impairment. Dev. Med. Child Neurol. 2007, 43, 56–60. [Google Scholar] [CrossRef]
- Hoyt, C.S. Visual Function in the Brain-Damaged Child. Eye 2003, 17, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Huo, R.; Burden, S.K.; Hoyt, C.S.; Good, W.V. Chronic Cortical Visual Impairment in Children: Aetiology, Prognosis, and Associated Neurological Deficits. Br. J. Ophthalmol. 1999, 83, 670–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jan, J.E.; Groenveld, M.; Sykanda, A.M.; Hoyt, C.S. Behavioural Characteristics Of Children With Permanent Cortical Visual Impairment. Dev. Med. Child Neurol. 2008, 29, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Malkowicz, D.E.; Myers, G.; Leisman, G. Rehabilitation of Cortical Visual Impairment in Children. Int. J. Neurosci. 2006, 116, 1015–1033. [Google Scholar] [CrossRef] [PubMed]
- McConnell, E.L.; Saunders, K.J.; Little, J. What Assessments Are Currently Used to Investigate and Diagnose Cerebral Visual Impairment (CVI) in Children? A Systematic Review. Ophthalmic Physiol. Opt. 2021, 41, 224–244. [Google Scholar] [CrossRef] [PubMed]
- Philip, S.S.; Dutton, G.N. Identifying and Characterising Cerebral Visual Impairment in Children: A Review: Cerebral Visual Impairment in Children: A Review. Clin. Exp. Optom. 2014, 97, 196–208. [Google Scholar] [CrossRef]
- Sakki, H.E.A.; Dale, N.J.; Sargent, J.; Perez-Roche, T.; Bowman, R. Is There Consensus in Defining Childhood Cerebral Visual Impairment? A Systematic Review of Terminology and Definitions. Br. J. Ophthalmol. 2018, 102, 424–432. [Google Scholar] [CrossRef] [Green Version]
- Chou, S.-J.; Babot, Z.; Leingartner, A.; Studer, M.; Nakagawa, Y.; O’Leary, D.D.M. Geniculocortical Input Drives Genetic Distinctions Between Primary and Higher-Order Visual Areas. Science 2013, 340, 1239–1242. [Google Scholar] [CrossRef] [Green Version]
- Cadwell, C.R.; Bhaduri, A.; Mostajo-Radji, M.A.; Keefe, M.G.; Nowakowski, T.J. Development and Arealization of the Cerebral Cortex. Neuron 2019, 103, 980–1004. [Google Scholar] [CrossRef]
- Alfano, C.; Studer, M. Neocortical Arealization: Evolution, Mechanisms, and Open Questions. Dev. Neurobiol. 2013, 73, 411–447. [Google Scholar] [CrossRef]
- Hofman, J.; Hutny, M.; Sztuba, K.; Paprocka, J. Corpus Callosum Agenesis: An Insight into the Etiology and Spectrum of Symptoms. Brain Sci. 2020, 10, 625. [Google Scholar] [CrossRef]
- Paul, L.K.; Brown, W.S.; Adolphs, R.; Tyszka, J.M.; Richards, L.J.; Mukherjee, P.; Sherr, E.H. Agenesis of the Corpus Callosum: Genetic, Developmental and Functional Aspects of Connectivity. Nat. Rev. Neurosci. 2007, 8, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Schell-Apacik, C.C.; Wagner, K.; Bihler, M.; Ertl-Wagner, B.; Heinrich, U.; Klopocki, E.; Kalscheuer, V.M.; Muenke, M.; von Voss, H. Agenesis and Dysgenesis of the Corpus Callosum: Clinical, Genetic and Neuroimaging Findings in a Series of 41 Patients. Am. J. Med. Genet. A 2008, 146A, 2501–2511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfano, C.; Viola, L.; Heng, J.I.-T.; Pirozzi, M.; Clarkson, M.; Flore, G.; De Maio, A.; Schedl, A.; Guillemot, F.; Studer, M. COUP-TFI Promotes Radial Migration and Proper Morphology of Callosal Projection Neurons by Repressing Rnd2 Expression. Development 2011, 138, 4685–4697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armentano, M.; Filosa, A.; Andolfi, G.; Studer, M. COUP-TFI Is Required for the Formation of Commissural Projections in the Forebrain by Regulating Axonal Growth. Development 2006, 133, 4151–4162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenembaum, S.N. Acute Disseminated Encephalomyelitis. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2013; Volume 112, pp. 1253–1262. ISBN 978-0-444-52910-7. [Google Scholar]
- Broadbent, N.J.; Squire, L.R.; Clark, R.E. Spatial Memory, Recognition Memory, and the Hippocampus. Proc. Natl. Acad. Sci. USA 2004, 101, 14515–14520. [Google Scholar] [CrossRef] [Green Version]
- Hannula, D.E.; Libby, L.A.; Yonelinas, A.P.; Ranganath, C. Medial Temporal Lobe Contributions to Cued Retrieval of Items and Contexts. Neuropsychologia 2013, 51, 2322–2332. [Google Scholar] [CrossRef] [Green Version]
- Kumaran, D.; Maguire, E.A. The Human Hippocampus: Cognitive Maps or Relational Memory? J. Neurosci. 2005, 25, 7254–7259. [Google Scholar] [CrossRef]
- Mátéffyová, A.; Otáhal, J.; Tsenov, G.; Mareš, P.; Kubová, H. Intrahippocampal Injection of Endothelin-1 in Immature Rats Results in Neuronal Death, Development of Epilepsy and Behavioral Abnormalities Later in Life. Eur. J. Neurosci. 2006, 24, 351–360. [Google Scholar] [CrossRef]
- Ramos, J.M.J. Hippocampal Damage Impairs Long-Term Spatial Memory in Rats: Comparison between Electrolytic and Neurotoxic Lesions. Physiol. Behav. 2008, 93, 1078–1085. [Google Scholar] [CrossRef]
- Wan, J.; Shen, C.M.; Wang, Y.; Wu, Q.Z.; Wang, Y.L.; Liu, Q.; Sun, Y.M.; Cao, J.P.; Wu, Y.Q. Repeated Exposure to Propofol in the Neonatal Period Impairs Hippocampal Synaptic Plasticity and the Recognition Function of Rats in Adulthood. Brain Res. Bull. 2021, 169, 63–72. [Google Scholar] [CrossRef]
- Zhao, A.; Fang, F.; Li, B.; Chen, Y.; Qiu, Y.; Wu, Y.; Xu, W.; Deng, Y. Visual Abnormalities Associate With Hippocampus in Mild Cognitive Impairment and Early Alzheimer’s Disease. Front. Aging Neurosci. 2021, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.B.; Moser, E.I.; Forrest, E.; Andersen, P.; Morris, R.G.M. Spatial Learning with a Minislab in the Dorsal Hippocampus. Proc. Natl. Acad. Sci. USA 1995, 92, 9697–9701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberhuber, M.; Hope, T.M.H.; Seghier, M.L.; Parker Jones, O.; Prejawa, S.; Green, D.W.; Price, C.J. Four Functionally Distinct Regions in the Left Supramarginal Gyrus Support Word Processing. Cereb. Cortex 2016, 26, 4212–4226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seghier, M.L. The Angular Gyrus: Multiple Functions and Multiple Subdivisions. Neuroscientist 2013, 19, 43–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoeckel, C.; Gough, P.M.; Watkins, K.E.; Devlin, J.T. Supramarginal Gyrus Involvement in Visual Word Recognition. Cortex 2009, 45, 1091–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkovich, A.J.; Guerrini, R.; Kuzniecky, R.I.; Jackson, G.D.; Dobyns, W.B. A Developmental and Genetic Classification for Malformations of Cortical Development: Update 2012. Brain 2012, 135, 1348–1369. [Google Scholar] [CrossRef] [PubMed]
- Guerrini, R.; Dobyns, W.B. Malformations of Cortical Development: Clinical Features and Genetic Causes. Lancet Neurol. 2014, 13, 710–726. [Google Scholar] [CrossRef] [Green Version]
- Juric-Sekhar, G.; Hevner, R.F. Malformations of Cerebral Cortex Development: Molecules and Mechanisms. Annu. Rev. Pathol. Mech. Dis. 2019, 14, 293–318. [Google Scholar] [CrossRef]
- Parrini, E.; Conti, V.; Dobyns, W.B.; Guerrini, R. Genetic Basis of Brain Malformations. Mol. Syndromol. 2016, 7, 220–233. [Google Scholar] [CrossRef] [Green Version]
- Hansen, D.V.; Lui, J.H.; Parker, P.R.L.; Kriegstein, A.R. Neurogenic Radial Glia in the Outer Subventricular Zone of Human Neocortex. Nature 2010, 464, 554–561. [Google Scholar] [CrossRef]
- Shitamukai, A.; Konno, D.; Matsuzaki, F. Oblique Radial Glial Divisions in the Developing Mouse Neocortex Induce Self-Renewing Progenitors Outside the Germinal Zone That Resemble Primate Outer Subventricular Zone Progenitors. J. Neurosci. 2011, 31, 3683–3695. [Google Scholar] [CrossRef] [PubMed]
- Vaid, S.; Camp, J.G.; Hersemann, L.; Eugster Oegema, C.; Heninger, A.-K.; Winkler, S.; Brandl, H.; Sarov, M.; Treutlein, B.; Huttner, W.B.; et al. A Novel Population of Hopx-Dependent Basal Radial Glial Cells in the Developing Mouse Neocortex. Development 2018, 145, dev169276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tsai, J.-W.; LaMonica, B.; Kriegstein, A.R. A New Subtype of Progenitor Cell in the Mouse Embryonic Neocortex. Nat. Neurosci. 2011, 14, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, F.K.; Fei, J.-F.; Mora-Bermúdez, F.; Taverna, E.; Haffner, C.; Fu, J.; Anastassiadis, K.; Stewart, A.F.; Huttner, W.B. Sustained Pax6 Expression Generates Primate-like Basal Radial Glia in Developing Mouse Neocortex. PLoS Biol. 2015, 13, e1002217. [Google Scholar] [CrossRef] [Green Version]
- Fietz, S.A.; Kelava, I.; Vogt, J.; Wilsch-Bräuninger, M.; Stenzel, D.; Fish, J.L.; Corbeil, D.; Riehn, A.; Distler, W.; Nitsch, R.; et al. OSVZ Progenitors of Human and Ferret Neocortex Are Epithelial-like and Expand by Integrin Signaling. Nat. Neurosci. 2010, 13, 690–699. [Google Scholar] [CrossRef]
- Bershteyn, M.; Nowakowski, T.J.; Pollen, A.A.; Di Lullo, E.; Nene, A.; Wynshaw-Boris, A.; Kriegstein, A.R. Human IPSC-Derived Cerebral Organoids Model Cellular Features of Lissencephaly and Reveal Prolonged Mitosis of Outer Radial Glia. Cell Stem Cell 2017, 20, 435–449.e4. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Muffat, J.; Omer, A.; Bosch, I.; Lancaster, M.A.; Sur, M.; Gehrke, L.; Knoblich, J.A.; Jaenisch, R. Induction of Expansion and Folding in Human Cerebral Organoids. Cell Stem Cell 2017, 20, 385–396.e3. [Google Scholar] [CrossRef] [Green Version]
- Qian, X.; Jacob, F.; Song, M.M.; Nguyen, H.N.; Song, H.; Ming, G. Generation of Human Brain Region–Specific Organoids Using a Miniaturized Spinning Bioreactor. Nat. Protoc. 2018, 13, 565–580. [Google Scholar] [CrossRef]
- Autism Sequencing Consortium; Lim, E.T.; Uddin, M.; De Rubeis, S.; Chan, Y.; Kamumbu, A.S.; Zhang, X.; D’Gama, A.M.; Kim, S.N.; Hill, R.S.; et al. Rates, Distribution and Implications of Postzygotic Mosaic Mutations in Autism Spectrum Disorder. Nat. Neurosci. 2017, 20, 1217–1224. [Google Scholar] [CrossRef] [Green Version]
- Parikshak, N.N.; Luo, R.; Zhang, A.; Won, H.; Lowe, J.K.; Chandran, V.; Horvath, S.; Geschwind, D.H. Integrative Functional Genomic Analyses Implicate Specific Molecular Pathways and Circuits in Autism. Cell 2013, 155, 1008–1021. [Google Scholar] [CrossRef] [Green Version]
- The DDD Study; Homozygosity Mapping Collaborative for Autism; UK10K Consortium; The Autism Sequencing Consortium; De Rubeis, S.; He, X.; Goldberg, A.P.; Poultney, C.S.; Samocha, K.; Ercument Cicek, A.; et al. Synaptic, Transcriptional and Chromatin Genes Disrupted in Autism. Nature 2014, 515, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Cotney, J.; Muhle, R.A.; Sanders, S.J.; Liu, L.; Willsey, A.J.; Niu, W.; Liu, W.; Klei, L.; Lei, J.; Yin, J.; et al. The Autism-Associated Chromatin Modifier CHD8 Regulates Other Autism Risk Genes during Human Neurodevelopment. Nat. Commun. 2015, 6, 6404. [Google Scholar] [CrossRef] [PubMed]
- Watt, N.; Wetherby, A.M.; Barber, A.; Morgan, L. Repetitive and Stereotyped Behaviors in Children with Autism Spectrum Disorders in the Second Year of Life. J. Autism Dev. Disord. 2008, 38, 1518–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molloy, A.; Rowe, F.J. Manneristic Behaviors of Visually Impaired Children. Strabismus 2011, 19, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Kerr, M.; Gil-Nagel, A.; Glynn, M.; Mula, M.; Thompson, R.; Zuberi, S.M. Treatment of Behavioral Problems in Intellectually Disabled Adult Patients with Epilepsy. Epilepsia 2013, 54, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Furniss, F.; Biswas, A.B. Recent Research on Aetiology, Development and Phenomenology of Self-Injurious Behaviour in People with Intellectual Disabilities: A Systematic Review and Implications for Treatment: Aetiology & Phenomenology of SIB in ID: A Systematic Review. J. Intellect. Disabil. Res. 2012, 56, 453–475. [Google Scholar] [CrossRef]
- Inoue, M. Assessments and Interventions to Address Challenging Behavior in Individuals with Intellectual Disability and Autism Spectrum Disorder in Japan: A Consolidated Review. Yonago Acta Med. 2019, 62, 169–181. [Google Scholar] [CrossRef] [Green Version]
- Matson, J.L.; Dempsey, T.; Rivet, T.T. The Interrelationships of Psychopathology Symptoms on Social Skills in Adults with Autism or PDD-NOS and Intellectual Disability. J. Dev. Phys. Disabil. 2009, 21, 39–55. [Google Scholar] [CrossRef]
- Oliver, C.; Licence, L.; Richards, C. Self-Injurious Behaviour in People with Intellectual Disability and Autism Spectrum Disorder. Curr. Opin. Psychiatry 2017, 30, 97–101. [Google Scholar] [CrossRef]
- Smith, K.R.M.; Matson, J.L. Social Skills: Differences among Adults with Intellectual Disabilities, Co-Morbid Autism Spectrum Disorders and Epilepsy. Res. Dev. Disabil. 2010, 31, 1366–1372. [Google Scholar] [CrossRef]
- Lowe, K.; Allen, D.; Jones, E.; Brophy, S.; Moore, K.; James, W. Challenging Behaviours: Prevalence and Topographies. J. Intellect. Disabil. Res. 2007, 51, 625–636. [Google Scholar] [CrossRef]
- Jacob, S.; Landeros-Weisenberger, A.; Leckman, J.F. Autism Spectrum and Obsessive–Compulsive Disorders: OC Behaviors, Phenotypes and Genetics. Autism Res. 2009, 2, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Muehlmann, A.M.; Lewis, M.H. Abnormal Repetitive Behaviours: Shared Phenomenology and Pathophysiology: Abnormal Repetitive Behaviours. J. Intellect. Disabil. Res. 2012, 56, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Amaral, D.G.; Schumann, C.M.; Nordahl, C.W. Neuroanatomy of Autism. Trends Neurosci. 2008, 31, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.; Sohn, I.-J.; Kim, N.; Sim, H.J.; Cheon, K.-A. Characteristics of Brains in Autism Spectrum Disorder: Structure, Function and Connectivity across the Lifespan. Exp. Neurobiol. 2015, 24, 273–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.K.; Andrews, D.S.; Ozonoff, S.; Solomon, M.; Rogers, S.; Amaral, D.G.; Nordahl, C.W. Longitudinal Evaluation of Cerebral Growth Across Childhood in Boys and Girls With Autism Spectrum Disorder. Biol. Psychiatry 2020, 90, 286–294. [Google Scholar] [CrossRef]
- Adhya, D.; Swarup, V.; Nagy, R.; Dutan, L.; Shum, C.; Valencia-Alarcón, E.P.; Jozwik, K.M.; Mendez, M.A.; Horder, J.; Loth, E.; et al. Atypical Neurogenesis in Induced Pluripotent Stem Cells From Autistic Individuals. Biol. Psychiatry 2021, 89, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Mariani, J.; Coppola, G.; Zhang, P.; Abyzov, A.; Provini, L.; Tomasini, L.; Amenduni, M.; Szekely, A.; Palejev, D.; Wilson, M.; et al. FOXG1-Dependent Dysregulation of GABA/Glutamate Neuron Differentiation in Autism Spectrum Disorders. Cell 2015, 162, 375–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Port, R.G.; Oberman, L.M.; Roberts, T.P. Revisiting the Excitation/Inhibition Imbalance Hypothesis of ASD through a Clinical Lens. Br. J. Radiol. 2019, 92, 20180944. [Google Scholar] [CrossRef]
- Siegel-Ramsay, J.E.; Romaniuk, L.; Whalley, H.C.; Roberts, N.; Branigan, H.; Stanfield, A.C.; Lawrie, S.M.; Dauvermann, M.R. Glutamate and Functional Connectivity—Support for the Excitatory-Inhibitory Imbalance Hypothesis in Autism Spectrum Disorders. Psychiatry Res. Neuroimaging 2021, 313, 111302. [Google Scholar] [CrossRef]
- Willsey, A.J.; Sanders, S.J.; Li, M.; Dong, S.; Tebbenkamp, A.T.; Muhle, R.A.; Reilly, S.K.; Lin, L.; Fertuzinhos, S.; Miller, J.A.; et al. Coexpression Networks Implicate Human Midfetal Deep Cortical Projection Neurons in the Pathogenesis of Autism. Cell 2013, 155, 997–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Yu, F.; Zhu, J.; Han, S.; Chen, J.; Wu, X.; Chen, Y.; Shen, T.; Liao, J.; Guo, W.; et al. Imbalance of Excitatory/Inhibitory Neuron Differentiation in Neurodevelopmental Disorders with an NR2F1 Point Mutation. Cell Rep. 2020, 31, 107521. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, B.; Velasco, S.; Kedaigle, A.J.; Pigoni, M.; Quadrato, G.; Deo, A.J.; Adiconis, X.; Uzquiano, A.; Sartore, R.; Yang, S.M.; et al. Autism Genes Converge on Asynchronous Development of Shared Neuron Classes. Nature 2022, 602, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Bjørnæs, H.; Stabell, K.; Henriksen, O.; Løyning, Y. The Effects of Refractory Epilepsy on Intellectual Functioning in Children and Adults. A Longitudinal Study. Seizure 2001, 10, 250–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, J.S. MRI Studies. Do Seizures Damage the Brain? In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2002; Volume 135, pp. 253–261. ISBN 978-0-444-50814-0. [Google Scholar]
- Mameniškienė, R.; Puteikis, K.; Jasionis, A.; Jatužis, D. A Review of Accelerated Long-Term Forgetting in Epilepsy. Brain Sci. 2020, 10, 945. [Google Scholar] [CrossRef]
- van Rijckevorsel, K. Cognitive Problems Related to Epilepsy Syndromes, Especially Malignant Epilepsies. Seizure 2006, 15, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shields, W.D. Infantile Spasms: Little Seizures, BIG Consequences. Epilepsy Curr. 2006, 6, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutula, T.P.; Hagen, J.; Pitkänen, A. Do Epileptic Seizures Damage the Brain? Curr. Opin. Neurol. 2003, 16, 189–195. [Google Scholar] [CrossRef]
- Pisani, F.; Fusco, C.; Spagnoli, C. Linking Acute Symptomatic Neonatal Seizures, Brain Injury and Outcome in Preterm Infants. Epilepsy Behav. 2020, 112, 107406. [Google Scholar] [CrossRef] [PubMed]
- Sutula, T.P.; Pitkanen, A. More Evidence for Seizure-Induced Neuron Loss: Is Hippocampal Sclerosis Both Cause and Effect of Epilepsy? Neurology 2001, 57, 169–170. [Google Scholar] [CrossRef]
- Scharfman, H.E. The Neurobiology of Epilepsy. Curr. Neurol. Neurosci. Rep. 2007, 7, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Bozzi, Y.; Provenzano, G.; Casarosa, S. Neurobiological Bases of Autism-Epilepsy Comorbidity: A Focus on Excitation/Inhibition Imbalance. Eur. J. Neurosci. 2018, 47, 534–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, E.M.; Campbell, D.B.; Stanwood, G.D.; Davis, C.; Noebels, J.L.; Levitt, P. Genetic Disruption of Cortical Interneuron Development Causes Region- and GABA Cell Type-Specific Deficits, Epilepsy, and Behavioral Dysfunction. J. Neurosci. 2003, 23, 622–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lévesque, M.; Biagini, G.; Avoli, M. Neurosteroids and Focal Epileptic Disorders. Int. J. Mol. Sci. 2020, 21, 9391. [Google Scholar] [CrossRef] [PubMed]
- Verhoog, Q.P.; Holtman, L.; Aronica, E.; van Vliet, E.A. Astrocytes as Guardians of Neuronal Excitability: Mechanisms Underlying Epileptogenesis. Front. Neurol. 2020, 11. [Google Scholar] [CrossRef]
- Anwar, H.; Khan, Q.U.; Nadeem, N.; Pervaiz, I.; Ali, M.; Cheema, F.F. Epileptic Seizures. Discov. Craiova Rom. 2020, 8, e110. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, H.; Huang, J.; Xu, M.; Xiao, C.; He, S. Regulation of Axon Initial Segment Diameter by COUP-TFI Fine-Tunes Action Potential Generation. Neurosci. Bull. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Aghakhani, Y. The Role of Periventricular Nodular Heterotopia in Epileptogenesis. Brain 2005, 128, 641–651. [Google Scholar] [CrossRef] [Green Version]
- Khoo, H.M.; Gotman, J.; Hall, J.A.; Dubeau, F. Treatment of Epilepsy Associated with Periventricular Nodular Heterotopia. Curr. Neurol. Neurosci. Rep. 2020, 20, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Meroni, A.; Galli, C.; Bramerio, M.; Tassi, L.; Colombo, N.; Cossu, M.; Lo Russo, G.; Garbelli, R.; Spreafico, R. Nodular Heterotopia: A Neuropathological Study of 24 Patients Undergoing Surgery for Drug-Resistant Epilepsy. Epilepsia 2009, 50, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Valton, L.; Guye, M.; McGonigal, A.; Marquis, P.; Wendling, F.; Régis, J.; Chauvel, P.; Bartolomei, F. Functional Interactions in Brain Networks Underlying Epileptic Seizures in Bilateral Diffuse Periventricular Heterotopia. Clin. Neurophysiol. 2008, 119, 212–223. [Google Scholar] [CrossRef]
- Balciuniene, J.; DeChene, E.T.; Akgumus, G.; Romasko, E.J.; Cao, K.; Dubbs, H.A.; Mulchandani, S.; Spinner, N.B.; Conlin, L.K.; Marsh, E.D.; et al. Use of a Dynamic Genetic Testing Approach for Childhood-Onset Epilepsy. JAMA Netw. Open 2019, 2, e192129. [Google Scholar] [CrossRef]
- Dimassi, S.; Labalme, A.; Ville, D.; Calender, A.; Mignot, C.; Boutry-Kryza, N.; de Bellescize, J.; Rivier-Ringenbach, C.; Bourel-Ponchel, E.; Cheillan, D.; et al. Whole-Exome Sequencing Improves the Diagnosis Yield in Sporadic Infantile Spasm Syndrome. Clin. Genet. 2016, 89, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Hino-Fukuyo, N.; Kikuchi, A.; Arai-Ichinoi, N.; Niihori, T.; Sato, R.; Suzuki, T.; Kudo, H.; Sato, Y.; Nakayama, T.; Kakisaka, Y.; et al. Genomic Analysis Identifies Candidate Pathogenic Variants in 9 of 18 Patients with Unexplained West Syndrome. Hum. Genet. 2015, 134, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Fournier, K.A.; Hass, C.J.; Naik, S.K.; Lodha, N.; Cauraugh, J.H. Motor Coordination in Autism Spectrum Disorders: A Synthesis and Meta-Analysis. J. Autism Dev. Disord. 2010, 40, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
- Jeste, S.S.; Geschwind, D.H. Clinical Trials for Neurodevelopmental Disorders: At a Therapeutic Frontier. Sci. Transl. Med. 2016, 8, 321fs1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.B.; Enticott, P.G.; Rinehart, N.J. Motor Development and Delay: Advances in Assessment of Motor Skills in Autism Spectrum Disorders. Curr. Opin. Neurol. 2018, 31, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Gerber, R.J.; Wilks, T.; Erdie-Lalena, C. Developmental Milestones: Motor Development. Pediatr. Rev. 2010, 31, 267–277. [Google Scholar] [CrossRef]
- Mosconi, M.W.; Kay, M.; D’Cruz, A.M.; Seidenfeld, A.; Guter, S.; Stanford, L.D.; Sweeney, J.A. Impaired Inhibitory Control Is Associated with Higher-Order Repetitive Behaviors in Autism Spectrum Disorders. Psychol. Med. 2009, 39, 1559–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donkelaar, H.J.; Lammens, M.; Wesseling, P.; Hori, A.; Keyser, A.; Rotteveel, J. Development and Malformations of the Human Pyramidal Tract. J. Neurol. 2004, 251, 1429–1442. [Google Scholar] [CrossRef]
- Roessmann, U.; Hori, A. Agyria (Lissencephaly) with Anomalous Pyramidal Crossing. Case Report and Review of Literature. J. Neurol. Sci. 1985, 69, 357–364. [Google Scholar] [CrossRef]
- Chow, C.W.; Halliday, J.L.; Anderson, R.M.D.; Danks, D.M.; Fortune, D.W. Congenital Absence of Pyramids and Its Significance in Genetic Diseases. Acta Neuropathol. 1985, 65, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Marcorelles, P.; Laquerriere, A. Neuropathology of Holoprosencephaly. Am. J. Med. Genet. C Semin. Med. Genet. 2010, 154C, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Welniarz, Q.; Dusart, I.; Roze, E. The Corticospinal Tract: Evolution, Development, and Human Disorders. Dev. Neurobiol. 2017, 810–829. [Google Scholar] [CrossRef]
- Tocco, C.; Øvsthus, M.; Bjaalie, J.G.; Leergaard, T.B.; Studer, M. The Topography of Corticopontine Projections Is Controlled by Postmitotic Expression of the Area-Mapping Gene Nr2f1. Development 2022, 149, dev200026. [Google Scholar] [CrossRef]
- Greig, L.C.; Woodworth, M.B.; Galazo, M.J.; Padmanabhan, H.; Macklis, J.D. Molecular Logic of Neocortical Projection Neuron Specification, Development and Diversity. Nat. Rev. Neurosci. 2013, 14, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Contesse, T.; Ayrault, M.; Mantegazza, M.; Studer, M.; Deschaux, O. Hyperactive and Anxiolytic-like Behaviors Result from Loss of COUP-TFI/Nr2f1 in the Mouse Cortex. Genes Brain Behav. 2019, 18, e12556. [Google Scholar] [CrossRef]
- Iverson, J.M. Developing Language in a Developing Body: The Relationship between Motor Development and Language Development. J. Child Lang. 2010, 37, 229–261. [Google Scholar] [CrossRef]
- Esposito, G.; Venuti, P.; Maestro, S.; Muratori, F. An Exploration of Symmetry in Early Autism Spectrum Disorders: Analysis of Lying. Brain Dev. 2009, 31, 131–138. [Google Scholar] [CrossRef]
- Bhat, A.N.; Landa, R.J.; Galloway, J.C. Current Perspectives on Motor Functioning in Infants, Children, and Adults with Autism Spectrum Disorders. Phys. Ther. 2011, 91, 1116–1129. [Google Scholar] [CrossRef] [Green Version]
- Karasik, L.B.; Tamis-Lemonda, C.S.; Adolph, K.E. Transition from Crawling to Walking and Infants’ Actions with Objects and People. Child Dev. 2011, 82, 1199–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barske, L.; Rataud, P.; Behizad, K.; Del Rio, L.; Cox, S.G.; Crump, J.G. Essential Role of Nr2f Nuclear Receptors in Patterning the Vertebrate Upper Jaw. Dev. Cell 2018, 44, 337–347.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLaurier, A. Evolution and Development of the Fish Jaw Skeleton. Wiley Interdiscip. Rev. Dev. Biol. 2019, 8, e337. [Google Scholar] [CrossRef] [PubMed]
- Rada-Iglesias, A.; Bajpai, R.; Prescott, S.; Brugmann, S.A.; Swigut, T.; Wysocka, J. Epigenomic Annotation of Enhancers Predicts Transcriptional Regulators of Human Neural Crest. Cell Stem Cell 2012, 11, 633–648. [Google Scholar] [CrossRef] [Green Version]
- Manikandan, M.; Abuelreich, S.; Elsafadi, M.; Alsalman, H.; Almalak, H.; Siyal, A.; Hashmi, J.A.; Aldahmash, A.; Kassem, M.; Alfayez, M.; et al. NR2F1 Mediated Down-Regulation of Osteoblast Differentiation Was Rescued by Bone Morphogenetic Protein-2 (BMP-2) in Human MSC. Differentiation 2018, 104, 36–41. [Google Scholar] [CrossRef]
- Ingulli, E.G.; Mak, R.H. Growth in Children with Chronic Kidney Disease: Role of Nutrition, Growth Hormone, Dialysis, and Steroids. Curr. Opin. Pediatr. 2014, 26, 187–192. [Google Scholar] [CrossRef]
- Putnick, D.L.; Bell, E.M.; Ghassabian, A.; Robinson, S.L.; Sundaram, R.; Yeung, E. Feeding Problems as an Indicator of Developmental Delay in Early Childhood. J. Pediatr. 2021, S0022347621010751. [Google Scholar] [CrossRef]
- Boyle, K.B.; Anderson, J.M. Hypotonia and Poor Feeding in a Newborn. NeoReviews 2008, 9, e602–e604. [Google Scholar] [CrossRef]
- Martin, K.; Inman, J.; Kirschner, A.; Deming, K.; Gumbel, R.; Voelker, L. Characteristics of Hypotonia in Children: A Consensus Opinion of Pediatric Occupational and Physical Therapists. Pediatr. Phys. Ther. 2005, 17, 275–282. [Google Scholar] [CrossRef]
- Prasad, A.N.; Prasad, C. The Floppy Infant: Contribution of Genetic and Metabolic Disorders. Brain Dev. 2003, 25, 457–476. [Google Scholar] [CrossRef]
- Qiu, Y.; Pereira, F.A.; DeMayo, F.J.; Lydon, J.P.; Tsai, S.Y.; Tsai, M.-J. Null Mutation of MCOUP-TFI Results in Defects in Morphogenesis of the Glossopharyngeal Ganglion, Axonal Projection, and Arborization. Genes Dev. 1997, 11, 1925–1937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, B.D.; Sanchez, R.; Rahman, T.; Rodriguez-Tirado, C.; Moritsch, S.; Martinez, A.R.; Miles, B.; Farias, E.; Mezei, M.; Nobre, A.R.; et al. An NR2F1-Specific Agonist Suppresses Metastasis by Inducing Cancer Cell Dormancy. J. Exp. Med. 2022, 219, e20210836. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.; Chen, C.-C.; Orr, A.; Barreto, P.; Tamamis, P.; Safe, S. Activation of COUP-TFI by a Novel Diindolylmethane Derivative. Cells 2019, 8, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourguet, W.; Ruff, M.; Chambon, P.; Gronemeyer, H. Crystal Structure of the Ligand-Binding Domain of the Human Nuclear Receptor RXR-α. Nature 1995, 375, 377–382. [Google Scholar] [CrossRef]
- Egea, P.F.; Mitschler, A.; Rochel, N.; Ruff, M.; Chambon, P.; Moras, D. Crystal Structure of the Human RXRa Ligand-Binding Domain Bound to Its Natural Ligand: 9-Cis Retinoic Acid. 10. EMBO J. 2000, 19, 2592–2601. [Google Scholar] [CrossRef]
- Germain, P.; Staels, B.; Dacquet, C.; Spedding, M.; Laudet, V. Overview of Nomenclature of Nuclear Receptors. Pharmacol. Rev. 2006, 58, 685–704. [Google Scholar] [CrossRef]
- Wang, Z.; Benoit, G.; Liu, J.; Prasad, S.; Aarnisalo, P.; Liu, X.; Xu, H.; Walker, N.P.C.; Perlmann, T. Structure and Function of Nurr1 Identifies a Class of Ligand-Independent Nuclear Receptors. Nature 2003, 423, 555–560. [Google Scholar] [CrossRef]
- Tsai, S.Y.; Tsai, M.-J. Chick Ovalbumin Upstream Promoter-Transcription Factors (COUP-TFs): Coming of Age. Endocr. Rev. 1997, 18, 229–240. [Google Scholar]
- Ang, C.E.; Ma, Q.; Wapinski, O.L.; Fan, S.; Flynn, R.A.; Lee, Q.Y.; Coe, B.; Onoguchi, M.; Olmos, V.H.; Do, B.T.; et al. The Novel LncRNA Lnc-NR2F1 Is pro-Neurogenic and Mutated in Human Neurodevelopmental Disorders. eLife 2019, 8, 41770. [Google Scholar] [CrossRef]
- Cooper, G.M.; Goode, D.L.; Ng, S.B.; Sidow, A.; Bamshad, M.J.; Shendure, J.; Nickerson, D.A. Single-Nucleotide Evolutionary Constraint Scores Highlight Disease-Causing Mutations. Nat. Methods 2010, 7, 250–251. [Google Scholar] [CrossRef] [Green Version]
- Gussow, A.B.; Petrovski, S.; Wang, Q.; Allen, A.S.; Goldstein, D.B. The Intolerance to Functional Genetic Variation of Protein Domains Predicts the Localization of Pathogenic Mutations within Genes. Genome Biol. 2016, 17, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kircher, M.; Witten, D.M.; Jain, P.; O’Roak, B.J.; Cooper, G.M.; Shendure, J. A General Framework for Estimating the Relative Pathogenicity of Human Genetic Variants. Nat. Genet. 2014, 46, 310–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivas, M.A.; Pirinen, M.; Conrad, D.F.; Lek, M.; Tsang, E.K.; Karczewski, K.J.; Maller, J.B.; Kukurba, K.R.; DeLuca, D.S.; Fromer, M.; et al. Effect of Predicted Protein-Truncating Genetic Variants on the Human Transcriptome. Science 2015, 348, 666–669. [Google Scholar] [CrossRef] [Green Version]
- Holbrook, J.A.; Neu-Yilik, G.; Hentze, M.W.; Kulozik, A.E. Nonsense-Mediated Decay Approaches the Clinic. Nat. Genet. 2004, 36, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.N.; Pearce, D.A. Nonsense-Mediated Decay in Genetic Disease: Friend or Foe? Mutat. Res. Mutat. Res. 2014, 762, 52–64. [Google Scholar] [CrossRef] [Green Version]
- DeBoever, C.; Tanigawa, Y.; Lindholm, M.E.; McInnes, G.; Lavertu, A.; Ingelsson, E.; Chang, C.; Ashley, E.A.; Bustamante, C.D.; Daly, M.J.; et al. Medical Relevance of Protein-Truncating Variants across 337,205 Individuals in the UK Biobank Study. Nat. Commun. 2018, 9, 1612. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.; Jensen, M.; Amini, H.; Hormozdiari, F.; Penn, O.; Shifman, S.; Girirajan, S.; Hormozdiari, F. Dissecting the Genetic Basis of Comorbid Epilepsy Phenotypes in Neurodevelopmental Disorders. Genome Med. 2019, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parenti, I.; Rabaneda, L.G.; Schoen, H.; Novarino, G. Neurodevelopmental Disorders: From Genetics to Functional Pathways. Trends Neurosci. 2020, 43, 608–621. [Google Scholar] [CrossRef] [PubMed]
- Hammoudi, D.S.; Lee, S.S.F.; Madison, A.; Mirabella, G.; Buncic, J.R.; Logan, W.J.; Snead, O.C.; Westall, C.A. Reduced Visual Function Associated with Infantile Spasms in Children on Vigabatrin Therapy. Investig. Opthalmol. Vis. Sci. 2005, 46, 514. [Google Scholar] [CrossRef] [PubMed]
- Holmes, G.L. Effect of Seizures on the Developing Brain and Cognition. Semin. Pediatr. Neurol. 2016, 23, 120–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-C.; Chang, Y.-C. The Long-Term Effects of Febrile Seizures on the Hippocampal Neuronal Plasticity—Clinical and Experimental Evidence. Brain Dev. 2009, 31, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Shields, W.D. Catastrophic Epilepsy in Childhood. Epilepsia 2000, 41, S2–S6. [Google Scholar] [CrossRef] [PubMed]
- Eldomery, M.K.; Coban-Akdemir, Z.; Harel, T.; Rosenfeld, J.A.; Gambin, T.; Stray-Pedersen, A.; Küry, S.; Mercier, S.; Lessel, D.; Denecke, J.; et al. Lessons Learned from Additional Research Analyses of Unsolved Clinical Exome Cases. Genome Med. 2017, 9, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jezela-Stanek, A.; Ciara, E.; Jurkiewicz, D.; Kucharczyk, M.; Jędrzejowska, M.; Chrzanowska, K.H.; Krajewska-Walasek, M.; Żemojtel, T. The Phenotype-driven Computational Analysis Yields Clinical Diagnosis for Patients with Atypical Manifestations of Known Intellectual Disability Syndromes. Mol. Genet. Genom. Med. 2020, 8, e1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochtus, A.; Olson, H.E.; Smith, L.; Keith, L.G.; El Achkar, C.; Taylor, A.; Mahida, S.; Park, M.; Kelly, M.; Shain, C.; et al. Genetic Diagnoses in Epilepsy: The Impact of Dynamic Exome Analysis in a Pediatric Cohort. Epilepsia 2020, 61, 249–258. [Google Scholar] [CrossRef]
- Fokkema, I.F.A.C.; Velde, K.J.; Slofstra, M.K.; Ruivenkamp, C.A.L.; Vogel, M.J.; Pfundt, R.; Blok, M.J.; Lekanne Deprez, R.H.; Waisfisz, Q.; Abbott, K.M.; et al. Dutch Genome Diagnostic Laboratories Accelerated and Improved Variant Interpretation and Increased Accuracy by Sharing Data. Hum. Mutat. 2019, 40, 2230–2238. [Google Scholar] [CrossRef] [PubMed]
- McInerney-Leo, A.M.; Duncan, E.L. Massively Parallel Sequencing for Rare Genetic Disorders: Potential and Pitfalls. Front. Endocrinol. 2021, 11, 628946. [Google Scholar] [CrossRef]
- Sun, H.; Shen, X.-R.; Fang, Z.-B.; Jiang, Z.-Z.; Wei, X.-J.; Wang, Z.-Y.; Yu, X.-F. Next-Generation Sequencing Technologies and Neurogenetic Diseases. Life 2021, 11, 361. [Google Scholar] [CrossRef] [PubMed]
- Vissers, L.E.L.M.; van Nimwegen, K.J.M.; Schieving, J.H.; Kamsteeg, E.-J.; Kleefstra, T.; Yntema, H.G.; Pfundt, R.; van der Wilt, G.J.; Krabbenborg, L.; Brunner, H.G.; et al. A Clinical Utility Study of Exome Sequencing versus Conventional Genetic Testing in Pediatric Neurology. Genet. Med. 2017, 19, 1055–1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niguidula, N.; Alamillo, C.; Shahmirzadi Mowlavi, L.; Powis, Z.; Cohen, J.S.; Farwell Hagman, K.D. Clinical Whole-Exome Sequencing Results Impact Medical Management. Mol. Genet. Genom. Med. 2018, 6, 1068–1078. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.Y.; Dillon, O.J.; Stark, Z.; Schofield, D.; Alam, K.; Shrestha, R.; Chong, B.; Phelan, D.; Brett, G.R.; Creed, E.; et al. Diagnostic Impact and Cost-Effectiveness of Whole-Exome Sequencing for Ambulant Children With Suspected Monogenic Conditions. JAMA Pediatr. 2017, 171, 855. [Google Scholar] [CrossRef] [PubMed]
- D’Gama, A.M.; Walsh, C.A. Somatic Mosaicism and Neurodevelopmental Disease. Nat. Neurosci. 2018, 21, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Cheroni, C.; Caporale, N.; Testa, G. Autism Spectrum Disorder at the Crossroad between Genes and Environment: Contributions, Convergences, and Interactions in ASD Developmental Pathophysiology. Mol. Autism 2020, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Hertz-Picciotto, I.; Schmidt, R.J.; Krakowiak, P. Understanding Environmental Contributions to Autism: Causal Concepts and the State of Science. Autism Res. 2018, 11, 554–586. [Google Scholar] [CrossRef] [PubMed]
- Imbriani, G.; Panico, A.; Grassi, T.; Idolo, A.; Serio, F.; Bagordo, F.; De Filippis, G.; De Giorgi, D.; Antonucci, G.; Piscitelli, P.; et al. Early-Life Exposure to Environmental Air Pollution and Autism Spectrum Disorder: A Review of Available Evidence. Int. J. Environ. Res. Public. Health 2021, 18, 1204. [Google Scholar] [CrossRef] [PubMed]
- Masini, E.; Loi, E.; Vega-Benedetti, A.F.; Carta, M.; Doneddu, G.; Fadda, R.; Zavattari, P. An Overview of the Main Genetic, Epigenetic and Environmental Factors Involved in Autism Spectrum Disorder Focusing on Synaptic Activity. Int. J. Mol. Sci. 2020, 21, 8290. [Google Scholar] [CrossRef] [PubMed]
- Kruse, S.W.; Suino-Powell, K.; Zhou, X.E.; Kretschman, J.E.; Reynolds, R.; Vonrhein, C.; Xu, Y.; Wang, L.; Tsai, S.Y.; Tsai, M.-J.; et al. Identification of COUP-TFII Orphan Nuclear Receptor as a Retinoic Acid–Activated Receptor. PLoS Biol. 2008, 6, e227. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.M. Strategies for Protecting Oligodendrocytes and Enhancing Remyelination in Multiple Sclerosis. Discov. Med. 2013, 16, 53–63. [Google Scholar]
- Winters, J.J.; Ferguson, C.J.; Lenk, G.M.; Giger-Mateeva, V.I.; Shrager, P.; Meisler, M.H.; Giger, R.J. Congenital CNS Hypomyelination in the Fig4 Null Mouse Is Rescued by Neuronal Expression of the PI(3,5)P2 Phosphatase Fig4. J. Neurosci. 2011, 31, 17736–17751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barateiro, A.; Fernandes, A. Temporal Oligodendrocyte Lineage Progression: In Vitro Models of Proliferation, Differentiation and Myelination. Biochim. Biophys. Acta BBA Mol. Cell Res. 2014, 1843, 1917–1929. [Google Scholar] [CrossRef] [Green Version]
- Chanoumidou, K.; Mozafari, S.; Baron-Van Evercooren, A.; Kuhlmann, T. Stem Cell Derived Oligodendrocytes to Study Myelin Diseases. Glia 2020, 68, 705–720. [Google Scholar] [CrossRef]
- Czepiel, M.; Boddeke, E.; Copray, S. Human Oligodendrocytes in Remyelination Research: Human Oligodendrocytes in Remyelination. Glia 2015, 63, 513–530. [Google Scholar] [CrossRef]
- Madhavan, M.; Nevin, Z.S.; Shick, H.E.; Garrison, E.; Clarkson-Paredes, C.; Karl, M.; Clayton, B.L.L.; Factor, D.C.; Allan, K.C.; Barbar, L.; et al. Induction of Myelinating Oligodendrocytes in Human Cortical Spheroids. Nat. Methods 2018, 15, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Marton, R.M.; Miura, Y.; Sloan, S.A.; Li, Q.; Revah, O.; Levy, R.J.; Huguenard, J.R.; Pașca, S.P. Differentiation and Maturation of Oligodendrocytes in Human Three-Dimensional Neural Cultures. Nat. Neurosci. 2019, 22, 484–491. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | LOVD Database ID; Patient ID | Variant Type | Variant (Protein) | MRI (General; Optic Nerve and Cortical Morphology) | DD | ID | Visual System Defect(s) and Visual Deficit | EOE/S | ASD Behavioral Abnormalities | Hypotonia | Severity Index |
|---|---|---|---|---|---|---|---|---|---|---|---|
| BR09 | BE18, #1 | De novo deletion (400–500 Kb MD at breakpoints following paracentric inversion) | Deleted | Cranial nerve abnormalities | Yes | ND | ND | ND | ND | Yes | 3 |
| CA09 | BE18, #2 | De novo deletion | Deleted | PH | Yes | Yes; speech delay | Coloboma | FS | ND | Yes | 6 |
| CA09 | BE18, #3 | De novo deletion | Deleted | PH | Yes | Yes; speech delay | ND | IS | ND | ND | 4 |
| CA09 | BE18, #4 | De novo deletion | Deleted | PH; HPM; MCP; polymicrogyria | Yes | Yes; speech delay | ND | Yes | ND | Yes | 5 |
| AK13 | BE18, #5; RE20, #27 | De novo deletion (582 Kb) | Deleted; (del. includes FLJ42709, FAM172A, POU5F2, and MIR2277) | OA (small OC) | Yes, DMD | No but speech delay | OA; GVI | No | ADHD | Yes | 6 |
| SA13 | BE18, #6; RE20, #53 | De novo MM in LBD | p.Arg404His | ND | ND | ND | ND | ND | ASD | ND | 1 |
| BO14 | LOVD: NR2F1_000001; BO14, #2; BE18, #8; RE20, #13 | De novo MM in DBD | p.Ser113Arg | OA (small OD and OC) | Yes | No or ND | OA; P/SOD; CVI; GVI | ND | ND | Yes | 4 |
| BO14 | LOVD: NR2F1_000002; BO14, #1; BE18, #7; RE20, #14 | De novo MM in DBD | p.Arg115Pro | normal | No | Yes (IQ 48) | OA; P/SOD; small ON; CVI; GVI | ND | ND | ND | 2 |
| BO14 | LOVD: NR2F1_000003; BO14, #3; BE18, #9; RE20, #48 | De novo MM in LBD | p.Leu252Pro | ND | Yes | Yes (IQ 55–65) | P/SOD; CVI; GVI | ND | ND | Yes | 4 |
| BO14 | BO14, #4; BE18, #10; RE20, #28 | Deletion (0.83 Mb) | Deleted; (del. includes FAM172A, KIAA0825) | ND | No | Mild (IQ 61–74) | P/SOD; CVI; GVI | ND | ND | ND | 2 |
| BO14 | BO14, #5; BE18, #11; RE20, #24 | De novo deletion (2.83 Mb) | Deleted; (del. includes FAM172A, KIAA0825, ANKRD31) | Normal or ND | Yes | No (IQ ND) | P/SOD; small ON; CVI; GVI | ND | ND | ND | 2 |
| BO14 | LOVD: NR2F1_000004; BO14, #6; CH16, #12; RE20, #12 | De novo MM in DBD | p.Arg112Lys | Normal or ND | Yes | Yes (IQ 52) | OA; P/SOD; mild GVI | ND | ASD; OCD | ND | 4 |
| HF15; HF17 | LOVD: NR2F1_000018; CH16, #14; RE20, #18 | De novo MM in DBD | p.Arg135Cys | Normal or ND | Yes, DMD | Yes (DQ < 20); speech delay; non-verbal | Bilateral OA | West Syndrome; IS and FS; generalized tonic seizures | ASD traits | ND | 5 |
| DI16 | LOVD: NR2F1_000057; CH16, #15; RE20, #11 | De novo in-frame deletion in DBD | p.Phe110del | CC thinning; LV asymmetry; septum pellucidum agenesis; abnormal gyration | Yes | Severe; non-verbal | No or ND | West Syndrome; spasms at 3 mo; IS; hypsarrythmic EEG; electroclinical spasms | ASD | Global | 6 |
| CH16 | LOVD: NR2F1_000041; CH16, #1; BE18, #16; RE20, #16 | De novo MM in DBD | p.Cys128Arg | CC thinning; WM reduction; leukodystrophy | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; GVI | Epilepsy with staring spells and generalized tonic-clonic seizures | ASD; self-injurious behavior; head-banging | Yes | 7 |
| MI14; CH16 | LOVD: NR2F1_000075; CH16, #2; BE18, #17; RE20, #17 | De novo MM in DBD | p.Arg135Ser | CC thinning; WM reduction; ON malformation and OC bilateral hypoplasia | Yes, DMD | Yes (IQ ND); speech delay | Mild OA; ONH; CVI; GVI | IS; Occipital lobe epilepsy | ASD; head-banging | Profound, axial and appendicular | 7 |
| CH16; EL17 | LOVD: NR2F1_000007; CH16, #3; BE18, #18; RE20, #19 | De novo MM in DBD | p.Cys138Tyr | WM reduction | Global | Yes | OA; GVI | FS; Abnormal EEG during sleep | ASD; RB (persistent head-banging) | No | 6 |
| CH16 | LOVD: NR2F1_000058; CH16, #4; BE18, #19; RE20, #21 | De novo MM in DBD | p.Arg142Leu | CC thinning at 7 mo | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; P/SOD; small ON; CVI; GVI | IS; atonic seizures with markedly abnormal EEG | ND | Yes | 6 |
| CH16 | LOVD: NR2F1_000010; CH16, #5; BE18 #20; RE20 #22 | De novo MM in DBD | p.Cys146Arg | CC thinning and septo-optic dysplasia | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; GVI | No | ASD traits; RB (self-stimulatory behaviors) | Yes | 6 |
| CH16 | LOVD: NR2F1_000024; CH16, #6; BE18, #21; RE20, #47; | De novo MM in DBD | p.Ala155Thr | Normal or ND | No | Mild speech delay (pronunciation; dysarthria) | No or ND | No | NO or ND | Yes | 2 |
| CH16 | LOVD: NR2F1_000078; CH16, #7; BE18, #22; RE20, #51 | De novo MM in LBD | p.Gly368Asp | Normal or ND | Yes | Yes (IQ ND); speech delay | No or ND | First generalized seizure at 18 yo | ASD; RB; aggressive behavior | No | 4 |
| CH16 | LOVD: NR2F1_000057; CH16, #8; BE18, #23; RE20, #10 | De novo in-frame deletion in DBD | p.Phe110del | CC thinning | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; ONH; CVI; GVI | IS | No or ND | Yes | 6 |
| CH16 | LOVD: NR2F1_000053; CH16, #9; CH16, #24; RE20, #40 | De novo frameshift truncation | p.Gly35Argfs*361 | Normal or ND | Yes, DMD | Yes (IQ 55–69; verbal IQ 35–40 at 16 yo); speech delay | OA; P/SOD; CVI; GVI | Few seizures at 3–4 yo | ASD; RB including PDD-NOS at 22 yo; ADHD | Yes | 6 |
| CH16 | LOVD: NR2F1_000055; CH16, #10; BE18, #25; RE20, #41 | De novo frameshift truncation | p.His97Hisfs*22 | ND | Yes | Yes (IQ ND); speech delay | OA; ONH; GVI | No | ASD; OCB/RB; ADHD | Yes | 5 |
| CH16 | LOVD: NR2F1_000052; CH16, #11; BE18, #26; RE20, #36 | De novo TIV | p? | Normal or ND | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; ONH; GVI | No | ASD; RB (head banging) | Yes | 5 |
| CH16 | LOVD: NR2F1_000052; CH16, #12; BE18, #27; #37 in RE20, #37 | De novo TIV | p? | CC thinning; ONH | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; ONH; P/SOD; CVI; GVI | Yes | ASD traits | Yes | 7 |
| CH16 | LOVD: NR2F1_000048; CH16, #13; BE18, #28; RE20, #35 | De novo TIV | p.M1? | ND | Yes, DMD | Yes (FSIQ in the 40 s); speech delay | OA; ONH; CVI; GVI | No | ASD; head banging | Substantial, central | 5 |
| CH16 | LOVD: NR2F1_000048; CH16, #14; BE18, #29; RE20, #38 | De novo TIV | p.M1? | CC and CS thinning; pyramidal decussation agenesis; right vs. left fiber directionality asymmetry | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; coloboma; ONH; GVI | Seizure at 3 yo, complex partial, left parietal | OCB (hand stereotypes); ADHD | Yes | 7 |
| CH16 | LOVD: NR2F1_000042; CH16, #15; BE18, #30; RE20, #39 | De novo TIV | p? | Cerebral malformations; bilateral HPM | Yes, DMD | Yes (IQ ND); speech delay; non-verbal | OA; GVI | Tonic-clonic seizures at 13 and 18 yo | OCB | Yes | 7 |
| CH16 | CH16, #16; BE18, #31; RE20, #29 | De novo deletion (0.2 Mb) | Deleted; (del. includes FAM172A, partial) | ND | Yes, DMD | Yes (IQ ND); speech delay | OA; P/SOD; pigmented maculae; GVI | No | ADHD | Yes | 5 |
| CH16 | CH16, #17; BE18, #32; RE20, #31 | Deletion (0.9 Mb) | Deleted; (del. includes FAM172A; KIAA0825, partial) | ND | Yes | Yes (IQ ND); speech delay | OA; P/SOD; GVI | No | ASD; ADHD | No | 4 |
| CH16 | CH16, #18; BE18, #33; RE20, #32 | Parental (son of CH16, #17), deletion (0.9 Mb) | Deleted | CC agenesis; DM of the EC and IC anterior limb; focal abnormality of the right CB | Yes, DMD | Yes (IQ ND); speech delay | OA; P/SOD; GVI | No | ASD traits; ADHD | Axial | 6 |
| CH16 | CH16, #19; BE18, #34; RE20, #30 | Deletion (1.2 Mb) | Deleted; (del. Includes FAM172A, KIAA0825, ANKRD31) | ND | Yes | Yes (verbal IQ 96; non-verbal IQ 70) | OA; GVI | No | ASD; PDD-NOS | No | 4 |
| CH16 | CH16, #20; BE18, #35; RE20, #23 | Deletion (5.0 Mb) | Deleted; (del. Includes FAM172A, KIAA0825, ANKRD32, MCTP30) | MCP | Yes | Yes (IQ ND) | No or ND | No | ND | Low muscle tone, normal mass and strength | 4 |
| KA17 | LOVD: NR2F1_000039; BE18, #36; RE20, #1 | De novo MM in DBD | p.Cys86Phe | CC thinning; WM reduction; MCP | Yes, DMD | Yes (DQ < 25 at 14 yo); speech delay; non-verbal | OA; mild bilateral ONH; CVI; GVI | One episode of IS; left occipital onset seizure in EEG; FS | Severe ASD; RB (self-stimulating, self-injurious behavior); limited social interaction | Yes | 7 |
| KA17 | LOVD: NR2F1_000079; BE18, #37; RE20, #52 | De novo MM in LBD | p.Leu372Pro | ND | Yes, DMD | Yes (IQ ND); speech delay | OA; GVI | ND | RB; ADHD | Yes | 5 |
| VI17 | LOVD: NR2F1_000017 | MM in DBD | p.Gly105Asp | Cerebral malformations | ND | Yes (IQ ND) | No or ND | ND | ND | ND | 2 |
| MH18 | LOVD: NR2F1_000040; BE18, #38; RE20, #5 | De novo MM in DBD | p.Lys96Glu | CC thinning | Yes | Yes (IQ ca. 30–50); speech delay | Mild OA; CVI; GVI | No | ND | Yes | 5 |
| PA19 | LOVD: NR2F1_000038; RE20, #45; JU21, #10 | Truncation | p.Tyr171* | CC thinning | Yes | Yes; mild (IQ ca. 77–80) | OA; GVI | No | Behavioral disorders; ADHD | ND | 5 |
| BO20 | LOVD: NR2F1_000037; RE20, #43 | De novo truncation | p.Gln28* | ONH | No | No (verbal IQ 141; nonverbal IQ 63) | OA; ONH; CVI; mild GVI | No; EEG showed rare isolated sharp waves from central regions | ASD (hand flapping and toe walking at 24 mo); behavioral disorders; ADHD | Yes | 5 |
| BE20 | LOVD: NR2F1_000059; BE20, #1 | De novo MM in DBD | p.Arg142His | CC thinning; OA (OC and nerve thinning); abnormal gyration | Yes | Yes | OA; amblyopia | IS at 8 mo | ASD and ADHD traits | Yes | 7 |
| BE20 | LOVD: NR2F1_000060; BE20, #2 | De novo truncation | p.Gln244* | CC thinning; ventricular asymmetry and enlargement; abnormal gyration; polymicrogyria | Yes | Yes | No or ND | ND | Behavioral disorders | Yes | 5 |
| BE20 | LOVD: NR2F1_000054; BE20, #3; JU21, #4 | De novo truncation | p.Glu39* | CC and OC thinning; CB malformation; ectopic nodular heterotopy; abnormal gyration | Yes | Yes (speech difficulties) | Severe bilateral OA; LVA | 3–4 ES/y | Stereotypical movements; RB; ADHD | Yes | 7 |
| BE20 | LOVD: NR2F1_000048; BE20, #4 | De novo TIV | p? | CC thinning; cortical malformation; abnormal gyration | Yes | Yes (speech difficulties) | OA | ND | ASD and ADHD traits; behavioral disorders | No | 5 |
| BE20 | LOVD: NR2F1_000056; BE20, #5 | De novo MM in DBD | p.Tyr98His | CC thinning; OC hypoplasia; abnormal gyration | Yes | Yes | OA | ND | ASD traits; behavioral disorders; stereotypical movements | Yes | 6 |
| BE20 | LOVD: NR2F1_000061; BE20, #6 | De novo frameshift truncation | p.Lys323Serfs*73 | Short CC; ON and chiasm thinning; hypoplastic olfactory lobes; abnormal gyration | Yes, DMD | Yes (speech difficulties) | OA; LVA | ND | ASD traits | No | 5 |
| ZO20 | LOVD: NR2F1_000085 | De novo truncation | p.Ser201* | ND | Mild/moderate | Mild/moderate | Bilateral P/SOD; LVA | ND | ND | ND | 2 |
| HO20 | LOVD: NR2F1_000084; RE20, #44 | De novo truncation | p.Glu85* | Normal or ND | Yes | Yes (IQ 69) | OA; GVI | Spells of behavioral arrest and non-responsiveness | ASD; auditory hallucinations and delusions; crying episodes | Yes | 6 |
| WA20 | LOVD: NR2F1_000051 | Frameshift truncation | p.Asn362fs*33 | CC; ON and OC hypoplasia; mild MCP | Apparent at 8 mo | Speech delay | Severe GVI | Myoclonic astatic seizures at 2½ yo | ASD | ND | 6 |
| MI20 | LOVD: NR2F1_000034; MI20, #1 | De novo MM in DBD | p.Gly105Ser | Benign enlargement of the subarachnoid spaces (BESS) | Yes, DMD | Speech delay; non-verbal until 2 yo | Bilateral OA; GVI | Myoclonic epilepsy diagnosed at 3 yo | RB | ND | 6 |
| MI20 | LOVD: NR2F1_000034; MI20, #2 | De novo MM in DBD | p.Gly105Ser | LV enlargement; intraventricular arachnoid cyst | Yes, DMD | Speech delay; non-verbal until 2 yo | Bilateral OA; GVI | Myoclonic epilepsy diagnosed at 4 yo | RB | ND | 6 |
| ST20 | LOVD: NR2F1_000035 | MM in DBD | p.Lys107Glu | CC; ON; OC and optic tracts atrophy; complex pituitary cyst. | Marked and global; DMD | ND | Declining visual acuity; legally blind by 10 yo | 1–3 yo + 30 episodes of FS; occasionally with myoclonus and generalized seizures | Aggressive behavior; depression; hallucinations | Yes | 6 |
| RE20 | LOVD: NR2F1_000065; RE20, #33 | De novo TIV | p.M1? | ND | No | ND | No or ND | No | ND | Yes | 1 |
| RE20 | LOVD: NR2F1_000048; RE20, #34 | De novo TIV | p.M1? | ND | Yes, DMD | Speech delay; non-verbal | OA; small ON; CVI; GVI | FS | ASD | Yes | 5 |
| RE20 | LOVD: NR2F1_000067; RE20, #2 | De novo MM in DBD | p.Cys86Arg | CC thinning | Yes | Speech delay; non-verbal | OA; CVI; GVI | IS | ASD traits | Yes | 7 |
| RE20 | LOVD: NR2F1_000068; RE20, #3; RO20, #170 | De novo MM in DBD | p.Val88Met | Normal or ND | Yes | Speech delay; non-verbal | OA; CVI; GVI | Onset at 9 wo; IS; focal and partial seizures; myoclonic jerks | ASD; RB (head banging) | Yes | 6 |
| RE20 | LOVD: NR2F1_000009; RE20, #4 | MM in DBD | p.Gly95Val | ND | Yes, DMD | Yes (IQ 56); speech delay | P/SOD; small ON; CVI; GVI | IS and absence seizures | ASD traits | Yes | 6 |
| RE20 | LOVD: NR2F1_000069; RE20, #6 | MM in DBD | p.Hys97Pro | Slightly decreased brain volume | Yes | Yes (IQ ND); speech delay; non-verbal | OA; CVI; GVI | Myoclonic seizures | ASD (severe) | Yes | 7 |
| RE20 | LOVD: NR2F1_000070; RE20, #7 | De novo MM in DBD | p.Tyr98Cys | Abnormal | Yes | Speech delay | P/SOD; ONH; CVI; GVI | Myoclonic; absence seizures | ASD; RB (head banging); ADHD | Yes | 7 |
| RE20 | LOVD: NR2F1_000071; RE20, #8 | De novo MM in DBD | p.Glu104Gly | ND | Yes | Speech delay; non-verbal | OA; CVI; GVI | No | ASD traits | Yes | 5 |
| RE20 | LOVD: NR2F1_000072; RE20, #9 | MM in DBD | p.Ser108Ile | ON thinning and small OC | Yes | Yes (IQ ND); speech delay; non-verbal | OA; small ON; CVI; GVI | No | ASD traits | Yes | 6 |
| RE20 | LOVD: NR2F1_000073; RE20, #15 | MM in DBD | p.Cys122Ser | ND | Yes, DMD | Speech delay; non-verbal | OA; GVI | IS | ASD traits | Yes | 5 |
| RE20 | LOVD: NR2F1_000074; RE20, #42 | frameshift truncation | p.Asn127Lysfs*270 | ND | Yes, DMD | Speech delay | ONH; CVI; GVI | Yes | ASD; auditory hallucinations | Yes | 6 |
| RE20 | LOVD: NR2F1_000076; RE20, #20 | De novo MM in DBD | p.Gln139His | CC thinning; DM; ON thinning and small OC | Yes, DMD | Speech delay | OA; CVI; GVI | No | ASD | Yes | 6 |
| RE20 | LOVD: NR2F1_000077; RE20, #49 | MM in LBD | p.Ala311Pro | Normal or ND | Yes | Mild (FSIQ 80 below average); speech delay | P/SOD; mild GVI | Generalized Myoclonic and absence seizures | ASD | Yes | 6 |
| RE20 | De novo MM in LBD | p.Glu318Asp | Abnormal | No but mild DMD | No (IQ 94; performance IQ 54) | OA; CVI; GVI | Atonic; Rolandic epilepsy | ASD | No | 4 | |
| RE20 | LOVD: NR2F1_000021; RE20, #46 | De novo truncation | p.Arg373* | CC; ON and OC thinning | Yes, DMD | Mild (DQ ca. 60–70); speech delay | P/SOD; ONH; CVI; GVI | No | ASD | Yes | 6 |
| RE20 | LOVD: NR2F1_000036; RE20, #54 | De novo MM in LBD | p.Met406Thr | Small ON; Abnormal MRI | Yes, DMD | Yes (IQ ND); speech delay | CVI; GVI | No | ASD | Yes | 6 |
| RE20 | LOVD: NR2F1_000063; RE20, #25 | Maternal, deletion (2.5 Mb) | Deleted; (del. includes FLJ42709, FAM172A, MIR2277, POU5F2, KIAA0825, MIR1974, ANKRD32, MCTP1, FAM81B, TTC37) | ND | Yes | Yes (IQ ND); speech delay | OA; CVI; GVI | Absence and tonic seizures | ASD; PDD-NOS; OCD; pacing and hitting | Yes | 5 |
| RE20 | LOVD: NR2F1_000064; RE20, #26 | Deletion (0.97 Mb) | Deleted | ND | Yes | Speech delay | OA; small ON; CVI; GVI | No | ASD | Mild | 5 |
| JU21 | JU21, #1 | Frameshift truncation | p.Ala2Glnfs*3 | CC thinning; abnormal gyration | Yes; mild | ND | OA; ONH; LVA | Occasional epileptic-like state during light sleep; FS at 4 yo; convulsions at 8 yo | ASD; ADHD | Hypotonia and hyperlaxity | 6 |
| JU21 | JU21, #2 | Frameshift truncation | p.Asn24Glyfs*379 | Normal or ND | Yes | Yes (IQ ND); speech difficulties; learning disability | OA; ONH; CVI; LVA | No | ND | ND | 3 |
| JU21 | JU21, #3 | AA duplication | p.Arg31dup | Normal or ND | Yes | Yes (IQ ND); learning disability | OA; CVI; LVA | No | ND | ND | 3 |
| JU21 | LOVD: NR2F1_000069; JU21, #5 | De novo MM in DBD | p.Hys97Pro | CC; ON and OC thinning; periventricular leukomalacia; MCP | Yes; global (delayed visual maturation) | Yes (IQ ND); learning disability | OA; LVA | One episode of FS | ASD | Moderate/severe | 7 |
| JU21 | JU21, #6 | De novo truncation | p.Leu118* | CC mild foreshortening | Yes; global | Yes (IQ ND); learning disability | OA; LVA | No | ND | Yes | 5 |
| JU21 | JU21, #7 | (Likely) de novo truncation | p.Tyr120* | WM abnormalities | Yes; pervasive; global apraxia | Yes (IQ ND); speech delay; learning disability | OA; LVA | Myoclonic epilepsy; focal impaired awareness seizures | ASD; ADHD | ND | 6 |
| BA19; JU21 | LOVD: NR2F1_000086; JU21, #8 | De novo MM in DBD | p.Cys122Trp | WM reduction; CC thinning | Yes; global | Yes (IQ ND); learning disability | OA; CVI; LVA | IS; Myoclonic epilepsy | ASD; ADHD | Yes | 7 |
| JU21 | JU21, #9 | De novo MM in DBD | p.Ala155Thr | WM reduction; ON thinning | Yes | Yes (IQ ND); speech delay; learning disability | CVI; LVA | No | ASD; anxiety; limited attention span | ND | 5 |
| JU21 | JU21, #11 | De novo MM in LBD | p.Thr200Arg | Lateral and third ventricles enlargement; MCP | Global | Yes (IQ ND); learning disability | Central, steady, maintained | No | ND | Yes | 5 |
| JU21 | JU21, #12 | De novo truncation | p.Trp233* | CC; ON and OC thinning; WM delayed maturation; brain abnormalities | Yes | Yes (IQ ND); speech difficulties | OA; microphthalmia; small ON head; CVI | No | Limited concentration and short attention span | ND | 5 |
| JU21 | LOVD: NR2F1_000082; JU21, #13 | De novo MM in LBD | p.Glu342Lys | Normal CC e ON; OC atrophy and defective rotation; Normal gyration | No | No | OA; ONH; LVA | No | ND | ND | 2 |
| JU21 | JU21, #14 | (Likely) de novo deletion in LBD | p.Glu346_Gln349del | ND | Yes; walking delay | Yes (IQ ND); speech delay; dyslexia; learning disability | ONH; CVI; LVA | No | ND | ND | 3 |
| JU21 | LOVD: NR2F1_000079; JU21, #15 | Familial MM in LBD | p.Leu372Pro | ND | Yes; walking delay | Yes (IQ ND); speech delay | Small ON head; CVI; LVA | No | ND | ND | 3 |
| JU21 | LOVD: NR2F1_000079; JU21, #16 | Familial MM in LBD | p.Leu372Pro | Normal or ND | Yes; walking delay | Yes (IQ ND); speech delay; | OA; ONH; CVI; LVA | One episode of FS | ND | ND | 4 |
| JU21 | LOVD: NR2F1_000079; JU21, #17 | Familial MM in LBD | p.Leu372Pro | Normal or ND | Yes; walking delay | Yes (IQ ND); speech delay; | OA; CVI; LVA | No | ND | ND | 3 |
| JU21 | JU21, #18 | Familial deletion in LBD | p.Arg373_Leu374del | ND | No | ND | OA; ONH; LVA | No | ND | ND | 1 |
| JU21 | JU21, #19 | Familial deletion in LBD | p.Arg373_Leu374del | ON atrophy | No | ND | OA; ONH; LVA | No | ND | ND | 2 |
| JU21 | JU21, #20 | De novo MM in LBD | p.Gly395Ser | ON atrophy; WM loss | Yes | Yes (IQ ND); learning disability | CVI; LVA | No | ND | Generalized | 5 |
| JU21 | LOVD: NR2F1_000083; JU21, #21 | De novo truncation | p.Glu400* | CC thinning; ON and OC atrophy; abnormal gyration | Yes | Yes (IQ ND); learning disability | OA; ONH; LVA | No | ASD; behavioral disorders | ND | 5 |
| JU21 | JU21, #22 | De novo whole-gene deletion (599 Kb) | deleted; (del. includes FAM172A; NR2F1-AS1, partial; KIAA0825, last exon) | CC thickening; ON atrophy; cerebral vascular system abnormalities | Yes | Yes (IQ ND); mild speech delay; learning disability | OA; LVA | No | ND | ND | 4 |
| JS20 | LOVD: NR2F1_000036 | De novo MM in LBD | p.Met406Thr | DM | Yes | Severe (IQ ND); speech delay; non-verbal | OA; suspected ON dysplasia; GVI | Seizures from 4 mo | Short attention span | ND | 6 |
| GA21 | LOVD: NR2F1_000048 | De novo TIV | p.M1? | Brain abnormalities; ON; OC and optic tract hypoplasia | Yes | Severe (IQ ND) | Right iris and chorioretinal coloboma; small ON; bilateral P/SOD; LVA | EEG at 12 yo showed possible occipital seizures | ND | Yes | 6 |
| BBSOAS Main Feature(s) | Clinical Description(s) |
|---|---|
| Developmental delay (DD) | Delay in reaching language, social or motor skills milestones |
| Intellectual disability (ID) | Significantly reduced ability to understand new or complex information and to learn and apply new skills (impaired intelligence). IQ ranging from profound ID with IQ < 20, to moderate (35 < IQ < 49) or mild ID (50 < IQ < 69) |
| Visual impairment | Optic nerve abnormalities and/or brain-based vision impairment: |
| Optic nerve atrophy or pallor | |
| Optic nerve hypoplasia | |
| Cortical visual impairment (difficulty locating objects in a crowded field and following rapidly moving images and scenes). | |
| Alacrima (abnormal amount of reflex tearing) | |
| Manifest latent nystagmus and poor tracking; congenital | |
| Significant refractive errors | |
| Amblyopia | |
| Hypotonia | Low muscle tone; congenital |
| Oromotor dysfunction | Swallowing, sucking and chewing problems; congenital |
| Repetitive behavior | Hand flapping, head banging and more |
| Autism spectrum disorder (ASD) | ASD or autistic traits |
| Seizures | Infantile and/or febrile; occipital seizures |
| Attention-deficit hyperactivity disorder (ADHD) | Inattention, impulsivity and hyperactivity |
| Hearing impairment | Abnormal hearing |
| Dysmorphic facial features | Mild and inconsistent |
| Thin corpus callosum and neocortical dysgyria | Hypoplasia of the corpus callosum and abnormal pattern of cortical convolutions and sulci (dysgyria in temporal and parietal areas) on brain MRI |
| Recommended Clinical Exam(s) | Exam Description |
|---|---|
| Developmental assessment | Identify areas of impairment and allow for early intervention. |
| Psychological evaluation for autism | ADI-R (Autism Diagnostic Interview, Revised) and ADOS (Autism Diagnostic Observation Schedule) testing performed by a certified clinical psychologist. |
| Brain MRI | Recommended at age three years or older. |
| Full, dilated eye examination | Fundus photography and OCT scan of RNFL to document optic nerve health performed by an ophthalmologist every two years. |
| Visual acuity tests | As appropriate for patient’s age and understanding |
| Full hearing evaluation | Every two years |
| Recommended Therapeutic Approach(es) | Therapy Description |
|---|---|
| Visual therapy | Focused on CVI |
| Physical therapy | Aiming to increase strength and to improve gross motor skills. |
| Occupational therapy | Aiming to improve fine motor skills and coordination. |
| Speech therapy | Consideration of sign language and alternative communication devices |
| ABA (Applied behavioral analysis) therapy | If ASD is diagnosed |
| Anti-convulsive treatment | If epilepsy is present |
| Overall prevalence (N = 92) | Variants in the DBD (N = 32) | Variants in the LBD (N = 17) | Deletions (N = 15) | TIV (N = 9) | Truncations (N = 11) | Frameshift (N = 7) | |
|---|---|---|---|---|---|---|---|
| Average severity index | 4.94 | 5.62 | 3.76 | 4.33 | 5.33 | 5.18 | 5.29 |
| Phenotypic feature | |||||||
| Morphology | |||||||
| Myelin defects | 14.13% (13/92) | 25.00% (8/32) | 11.76% (2/17) | 6.67% (1/15) | 0.00% (0/9) | 18.18% (2/11) | 0.00% (0/7) |
| CC malformations | 32.61% (30/92) | 46.88% (15/32) | 0.00% (0/17) | 13.33% (2/15) | 33.33% (3/9) | 63.64% (7/11) | 42.86% (3/7) |
| Development and behavior | |||||||
| DD | 88.04% (81/92) | 90.62% (29/32) | 70.59% (12/17) | 93.33% (14/15) | 88.89% (8/9) | 90.91% (10/11) | 100% (7/7) |
| DMD | 30.43% (28/92) | 40.63% (13/32) | 11.76% (2/17) | 20.00% (3/15) | 66.67% 6/9) | 9.09% (1/11) | 42.86% (3/7) |
| ID/speech delay | 86.95% (80/92) | 93.75% (30/32) | 70.59% (12/17) | 86.67% (13/15) | 88.89% (8/9) | 90.91% (10/11) | 85.71% (6/7) |
| ASD | 38.04% (32/92) | 40.63% (13/32) | 29.41% (5/17) | 26.67% (4/15) | 33.33% (3/9) | 45.45% (5/11) | 71.43% (5/7) |
| ASD-like traits | 14.13% (13/92) | 28.13% (9/32) | 0.00% (0/17) | 6.67% (1/15) | 22.22% (2/9) | 0.00% (0/11) | 14.29% (1/7) |
| ADHD | 18.48% (17/92) | 9.38% (3/32) | 5.88% (1/17) | 26.67% (4/15) | 22.22% (2/9) | 36.36% (4/11) | 42.86% (3/7) |
| Visual system | |||||||
| CVI | 42.39% (39/92) | 53.13% (17/32) | 47.06% (8/17) | 26.67% (4/15) | 33.33% (3/9) | 27.27% (3/11) | 42.86% (3/7) |
| OA | 67.39% (62/92) | 78.13% (25/32) | 47.06% (8/17) | 53.33% (8/15) | 77.78% (7/15) | 72.73% (8/11) | 71.43% (5/7) |
| ONH | 21.74% (20/92) | 12.50% (4/32) | 29.41% (5/17) | 0.00% (0/15) | 44.44% (4/9) | 27.27% (3/11) | 57.14% (4/7) |
| P/SOD | 19.56% (18/92) | 18.75% (6/32) | 11.76% (2/17) | 33.33% (5/15) | 22.22% (2/9) | 18.18% (2/11) | 14.29% (1/7) |
| Others | |||||||
| Epilepsy | 45.65% (42/92) | 62.50% (20/32) | 29.41% (5/17) | 26.67% (4/15) | 55.56% (5/9) | 45.45% (5/11) | 42.86% (3/7) |
| Hypotonia | 61.96% (57/92) | 75.00% (24/32) | 35.29% (6/17) | 60.00% (9/15) | 88.89% (8/9) | 54.55% (6/11) | 57.14% (4/7) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertacchi, M.; Tocco, C.; Schaaf, C.P.; Studer, M. Pathophysiological Heterogeneity of the BBSOA Neurodevelopmental Syndrome. Cells 2022, 11, 1260. https://doi.org/10.3390/cells11081260
Bertacchi M, Tocco C, Schaaf CP, Studer M. Pathophysiological Heterogeneity of the BBSOA Neurodevelopmental Syndrome. Cells. 2022; 11(8):1260. https://doi.org/10.3390/cells11081260
Chicago/Turabian StyleBertacchi, Michele, Chiara Tocco, Christian P. Schaaf, and Michèle Studer. 2022. "Pathophysiological Heterogeneity of the BBSOA Neurodevelopmental Syndrome" Cells 11, no. 8: 1260. https://doi.org/10.3390/cells11081260
APA StyleBertacchi, M., Tocco, C., Schaaf, C. P., & Studer, M. (2022). Pathophysiological Heterogeneity of the BBSOA Neurodevelopmental Syndrome. Cells, 11(8), 1260. https://doi.org/10.3390/cells11081260