
Regulation of the Actin Cytoskeleton-Linked Ca2+ Signaling by Intracellular pH in Fertilized Eggs of Sea Urchin




Abstract
:1. Introduction
2. Materials and Methods
2.1. Gametes Collection, Modification of Seawater pH, and Embryos Observation
2.2. Visualization of Egg-Incorporated Sperm
2.3. Scanning Electron Microscopy (SEM) and Transmission Electron Microscopy (TEM)
2.4. Microinjection, Ca2+ Imaging, and Confocal Microscopy
2.5. Statistical Analysis
3. Results
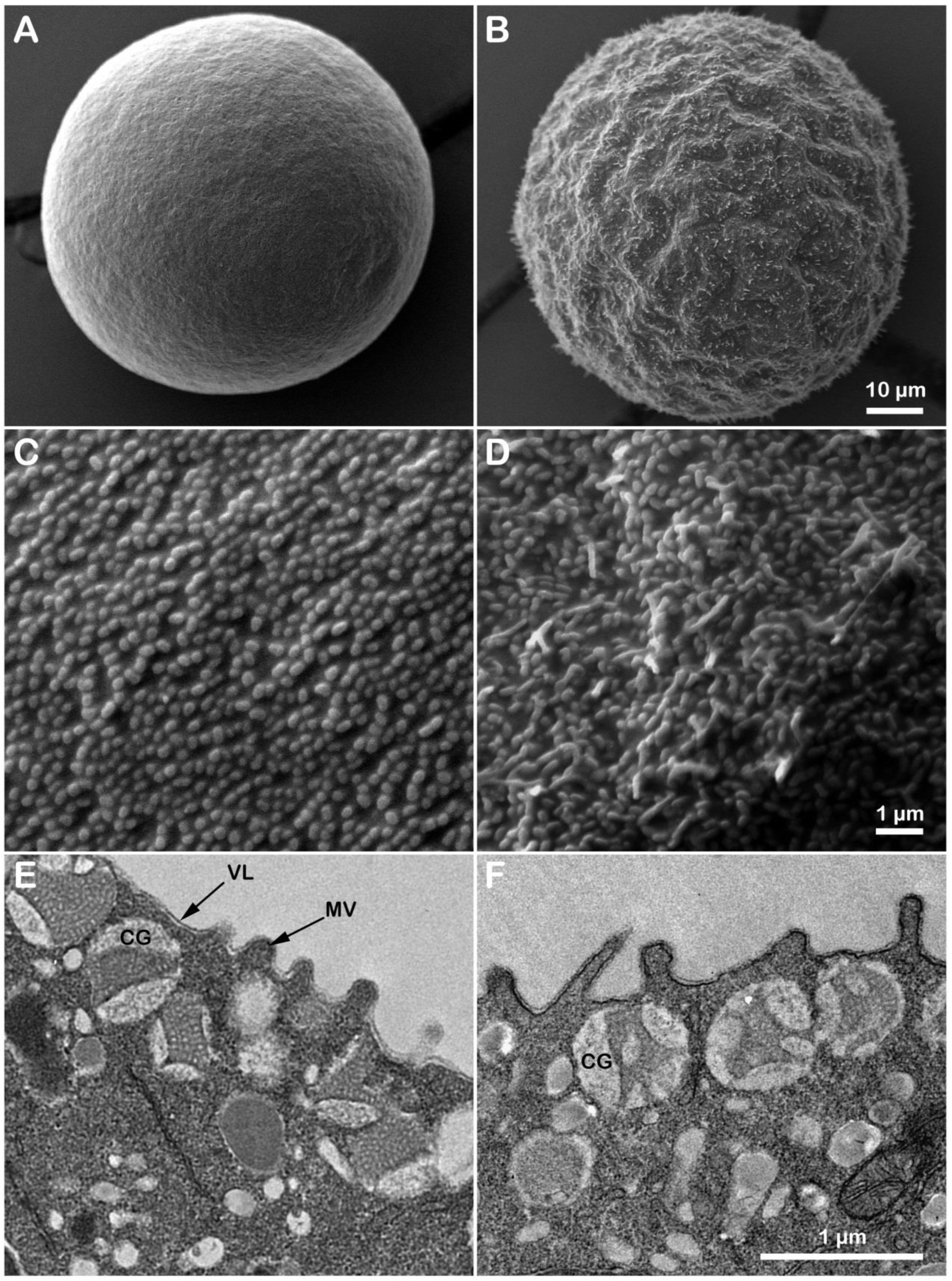
3.1. Microvillar Reorganization in Unfertilized P. lividus Eggs Exposed to NH40H-Seawater at pH 9
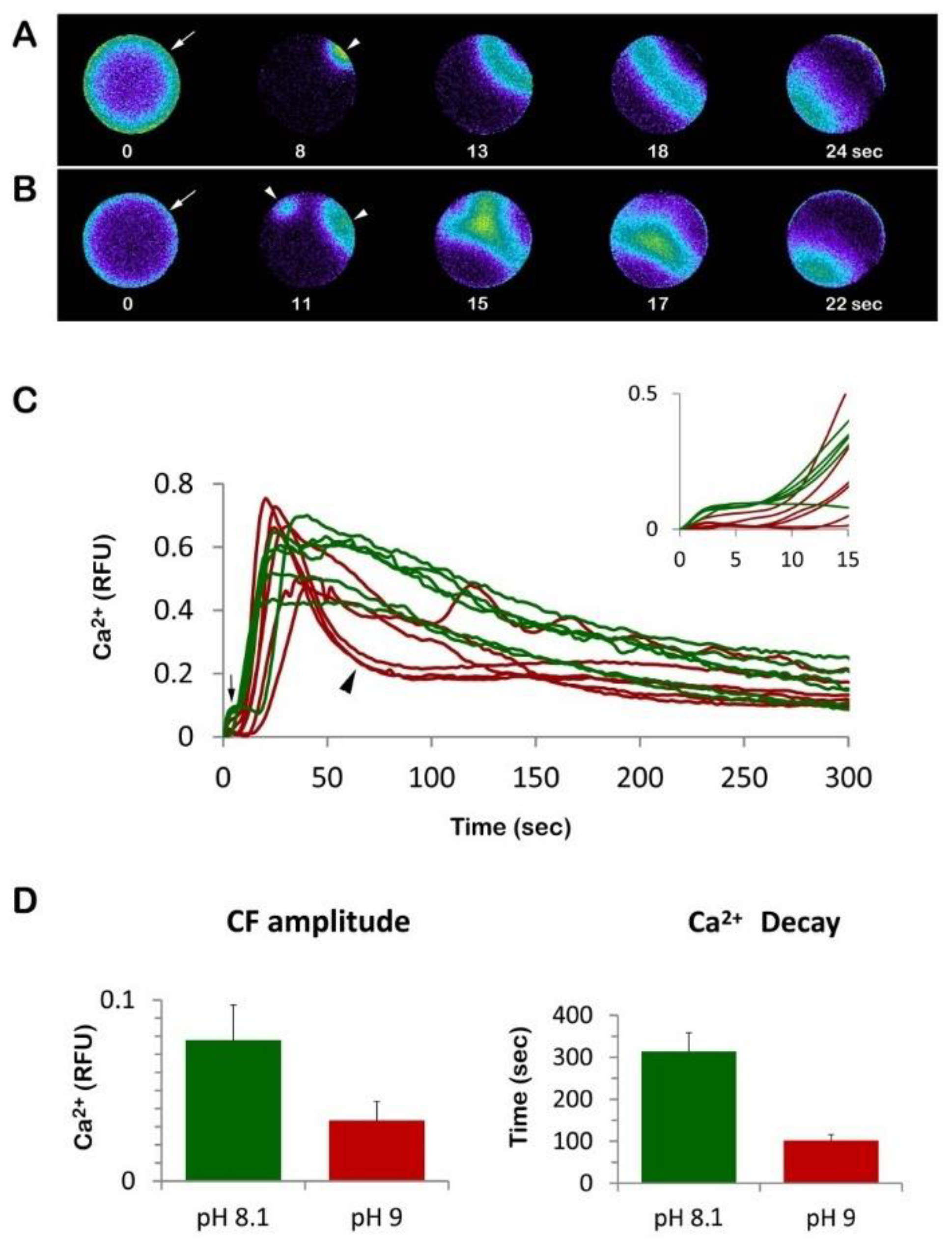
3.2. Eggs Incubated and Fertilized in Ammonia Seawater Display an Altered Ca2+ Response at Fertilization
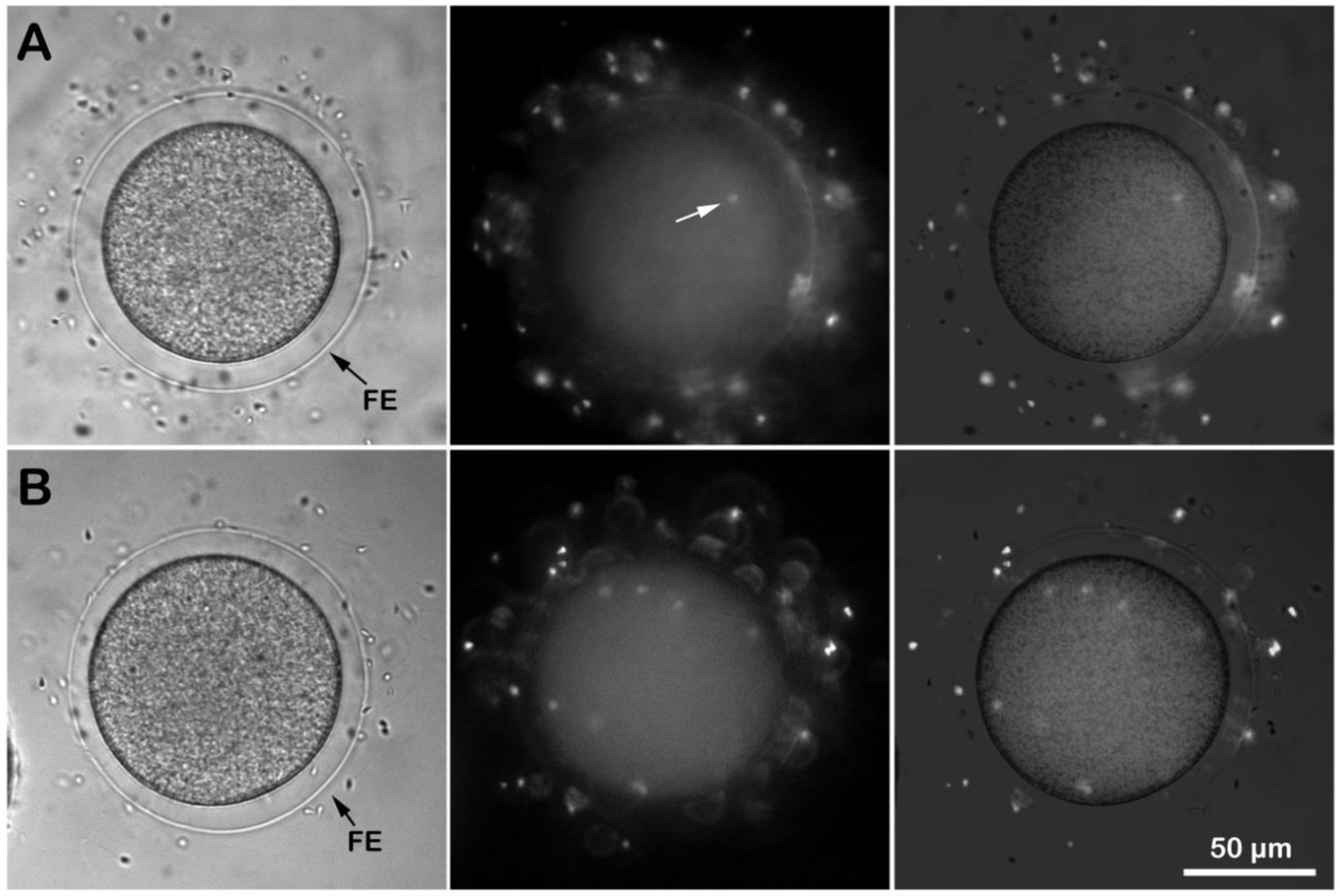
3.3. Eggs Fertilized in Ammonia Seawater Have a High Tendency of Polyspermy at Fertilization
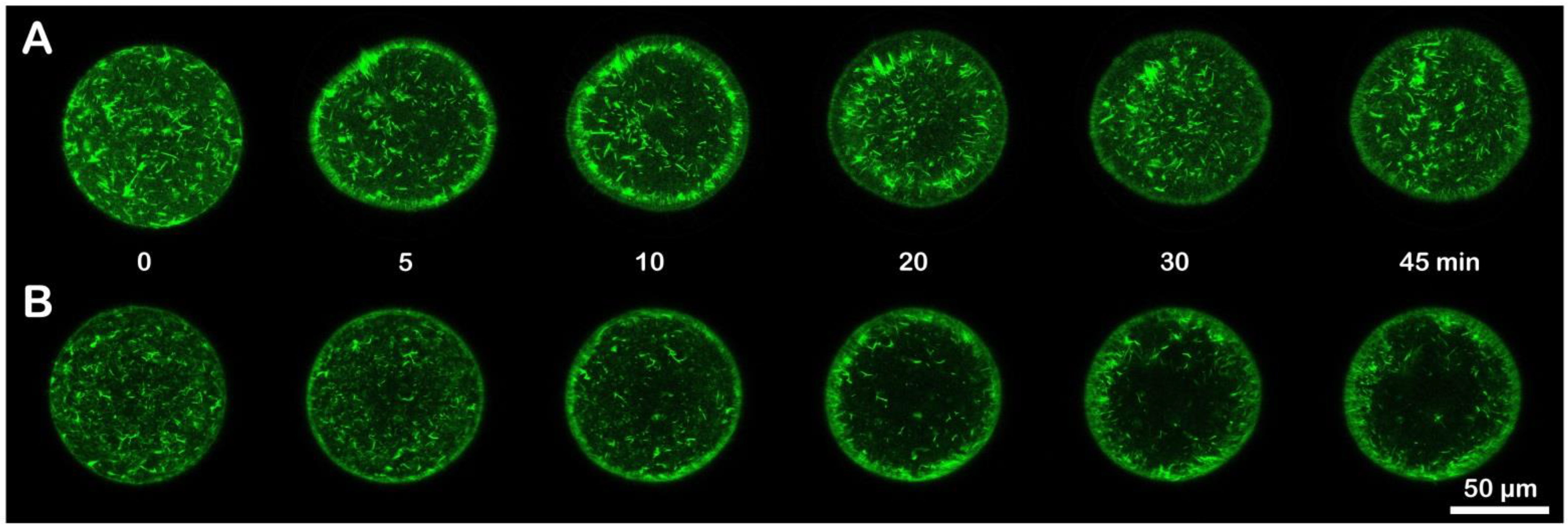
3.4. Effect of Ammonia Seawater on the Cortical F-Actin Changes in Fertilized Eggs
3.5. Effects of Ammonia Seawater on the Egg Surface Topography and Cortical Ultrastructure at Fertilization
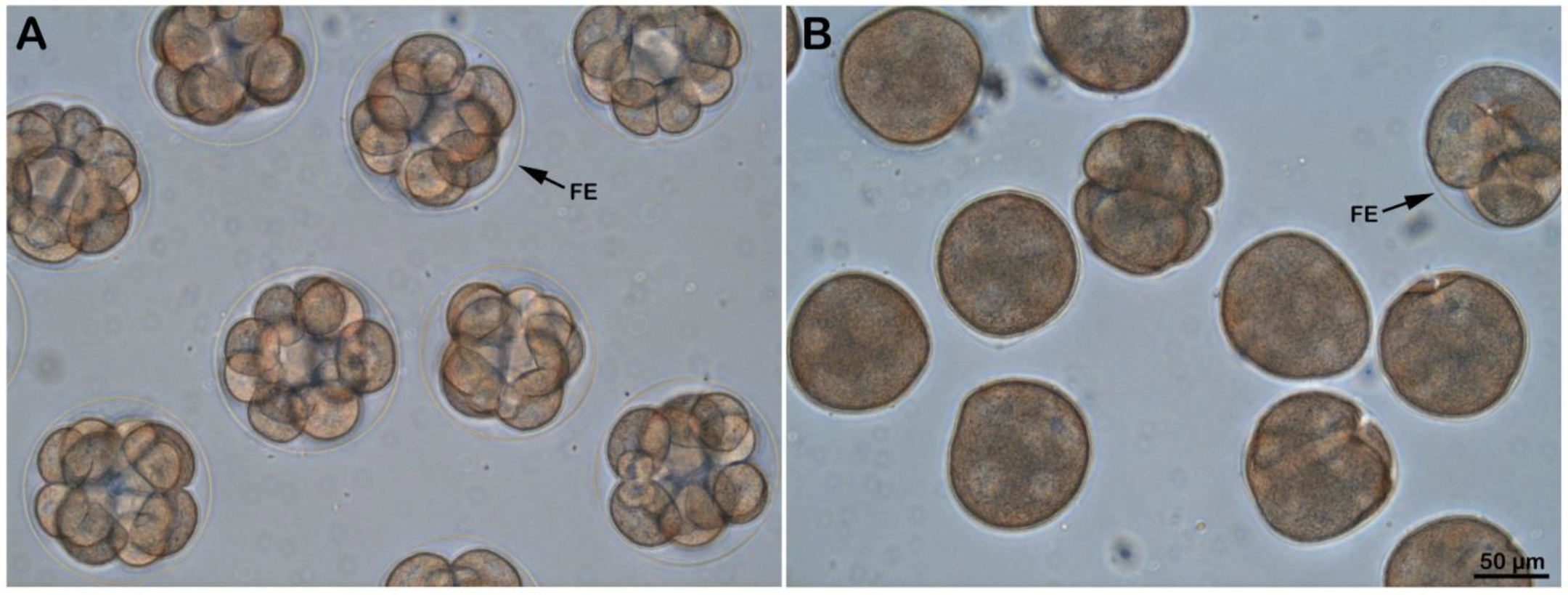
3.6. Effect of Ammonia Seawater on Embryonic Development
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hirohashi, N.; Vilela-Silva, A.-C.E.S.; Mourão, P.A.S.; Vacquier, V.D. Structural requirements for specie specific induction of the sperm acrosome reaction by sea urchin egg sulfated fucan. Biochem. Biophys. Res. Commun. 2002, 298, 403–407. [Google Scholar] [CrossRef]
- Dan, J.C. Studies on the acrosome. I reaction to egg-water and other stimuli. Biol. Bull. 1952, 103, 54–66. [Google Scholar] [CrossRef]
- Hoshi, M.; Moriyama, H.; Matsumoto, M. Structure of acrosome reaction-inducing substance in the jelly coat of starfish eggs: A mini review. Biochem. Biophys. Res. Commun. 2012, 425, 595–598. [Google Scholar] [CrossRef] [PubMed]
- Dale, B.; Dan-Sohkawa, M.; De Santis, A.; Hoshi, M. Fertilization of the starfish Astropecten aurantiacus. Exp. Cell Res. 1981, 132, 505–510. [Google Scholar] [CrossRef]
- Puppo, A.; Chun, J.T.; Gragnaniello, G.; Garante, E.; Santella, L. Alteration of the Cortical Actin Cytoskeleton Deregulates Ca2+ Signaling, Monospermic Fertilization, and Sperm Entry. PLoS ONE 2008, 3, e3588. [Google Scholar] [CrossRef] [Green Version]
- Santella, L.; Puppo, A.; Chun, J.T. The role of the actin cytoskeleton in calcium signaling in starfish oocytes. Int. J. Dev. Biol. 2008, 52, 571–584. [Google Scholar] [CrossRef] [Green Version]
- Santella, L.; Limatola, N.; Chun, J.T. Cellular and molecular aspects of oocyte maturation and fertilization: A perspective from the actin cytoskeleton. Zool. Lett. 2020, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Vacquier, V.D.; Moy, G.W. Isolation of bindin: The protein responsible for adhesion of sperm to sea urchin eggs. Proc. Natl. Acad. Sci. 1977, 74, 2456–2460. [Google Scholar] [CrossRef] [Green Version]
- Vacquier, V.D. The quest for the sea urchin egg receptor for sperm. Biochem. Biophys. Res. Commun. 2012, 425, 583–587. [Google Scholar] [CrossRef]
- Kamei, N.; Glabe, C.G. The species-specific egg receptor for sea urchin sperm adhesion is EBR1, a novel ADAMTS protein. Genes Dev. 2003, 17, 2502–2507. [Google Scholar] [CrossRef] [Green Version]
- Wessel, G.M.; Wada, Y.; Yajima, M.; Kiyomoto, M. Bindin is essential for fertilization in the sea urchin. Proc. Natl. Acad. Sci. USA 2021, 118, e2109636118. [Google Scholar] [CrossRef] [PubMed]
- Steinhardt, R.A.; Lundin, L.; Mazia, D. Bioelectric responses of the echinoderm egg to fertilization. Proc. Natl. Acad. Sci. USA 1971, 68, 2426–2430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, E.L.; de Armendi, J. Membrane potential, action potential and activation potential of eggs of the sea urchin, Lytechinus variegatus. Exp. Cell Res. 1979, 122, 203–218. [Google Scholar] [CrossRef]
- Just, E.E. The fertilization reaction in Echinarachnius parma. I. Cortical response of the egg to insemination. Biol. Bull. 1919, 36, 1–10. [Google Scholar] [CrossRef]
- Just, E.E. The Biology of the Cell Surface; P. Blakiston’s Son & Co., Inc.: Philadelphia, PA, USA, 1939. [Google Scholar] [CrossRef]
- Jaffe, L.A. Fast block to polyspermy in sea urchin eggs is electrically mediated. Nature 1976, 261, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Limatola, N.; Vasilev, F.; Chun, J.T.; Santella, L. Sodium-mediated fast electrical depolarization does not prevent polyspermic fertilization in Paracentrotus lividus eggs. Zygote 2019, 27, 241–249. [Google Scholar] [CrossRef]
- Santella, L.; Limatola, N.; Chun, J.T. Calcium and actin in the saga of awakening oocytes. Biochem. Biophys. Res. Commun. 2015, 460, 104–113. [Google Scholar] [CrossRef]
- Dale, B.; De Belice, L.J.; Taglietti, V. Membrane noise and conductance increase during single spermatozoon-egg interactions. Nature 1978, 275, 217–219. [Google Scholar] [CrossRef]
- Shen, S.S.; Buck, W.R. Sources of calcium in sea urchin eggs during the fertilization response. Dev. Biol. 1993, 157, 157–169. [Google Scholar] [CrossRef]
- Créton, R.; Jaffe, L.F. Role of calcium influx during the latent period in sea urchin. Develop. Growth Differ. 1995, 37, 703–709. [Google Scholar] [CrossRef]
- Vasilev, F.; Limatola, N.; Chun, J.T.; Santella, L. Contributions of suboolemmal acidic vesicles and microvilli to the intracellular Ca2+ increase in the sea urchin eggs at fertilization. Int. J. Biol. Sci. 2019, 15, 757–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciapa, B.; Epel, D. A rapid change in phosphorylation on tyrosine accompanies fertilization of sea urchin eggs. FEBS Lett. 1991, 295, 167–170. [Google Scholar] [CrossRef] [Green Version]
- Runft, L.L.; Jaffe, L.A.; Mehlmann, L.M. Egg Activation at Fertilization: Where It All Begins. Dev. Biol. 2002, 245, 237–254. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, M. Calcium at fertilization and in early development. Physiol. Rev. 2006, 86, 25–88. [Google Scholar] [CrossRef]
- Jaffe, L.F. Sources of calcium in egg activation: A review and hypothesis. Dev. Biol. 1983, 99, 265–276. [Google Scholar] [CrossRef]
- Dale, B.; DeFelice, L.J.; Ehrenstein, G. Injection of a soluble sperm fraction into sea-urchin eggs triggers the cortical reaction. Experientia 1985, 41, 1068–1070. [Google Scholar] [CrossRef]
- Santella, L.; Kyozuka, K.; Genazzani, A.A.; De Riso, L.; Carafoli, E. Nicotinic acid adenine dinucleotide phosphate-induced Ca2+ release: Interactions among distinct Ca2+ mobilizing mechanisms in starfish oocytes. J. Biol. Chem. 2000, 275, 8301–8306. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.; Kyozuka, K.; Gragnaniello, G.; Carafoli, E.; Santella, L. NAADP+ initiates the Ca2+ response during fertilization of starfish oocytes. FASEB J. 2001, 15, 2257–2267. [Google Scholar] [CrossRef]
- Churchill, G.C.; Okada, Y.; Thomas, J.M.; Genazzani, A.A.; Patel, S.; Galione, A. NAADP mobilizes Ca(2+) from reserve granules, lysosome-related organelles, in sea urchin eggs. Cell 2002, 111, 703–708. [Google Scholar] [CrossRef] [Green Version]
- Moccia, F.; Lim, D.; Nusco, G.A.; Ercolano, E.; Santella, L. NAADP activates a Ca2+ current that is dependent on F-actin cytoskeleton. FASEB J. 2003, 17, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Santella, L.; Lim, D.; Moccia, F. Calcium and fertilization: The beginning of life. Trends Biochem. Sci. 2004, 29, 400–408. [Google Scholar] [CrossRef]
- Churchill, G.C.; O’Neill, J.S.; Masgrau, R.; Patel, S.; Thomas, J.M.; Genazzani, A.A.; Galione, A. Sperm deliver a new second messenger: NAADP. Curr. Biol. 2003, 13, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Epel, D. A primary metabolic change of fertilization: Interconversion of pyridine nucleotides. Biochim. Biophys. Res. Commun. 1964, 17, 62–68. [Google Scholar] [CrossRef]
- Heinecke, J.W.; Shapiro, B.M. Respiratory burst oxidase of fertilization. Proc. Nat. Acad. Sci. USA 1989, 86, 1259–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santella, L.; Chun, J.T. Actin, more than just a housekeeping protein at the scene of fertilization. Sci. China Life Sci. 2011, 54, 733–743. [Google Scholar] [CrossRef] [Green Version]
- Santella, L.; Vasilev, F.; Chun, J.T. Fertilization in echinoderms. Biochem. Biophys. Res. Commun. 2012, 425, 588–594. [Google Scholar] [CrossRef]
- Santella, L.; Limatola, N.; Vasilev, F.; Chun, J.T. Maturation and fertilization of echinoderm eggs: Role of actin cytoskeleton dynamics. Biochem. Biophys. Res. Commun. 2018, 506, 361–371. [Google Scholar] [CrossRef]
- Chun, J.T.; Vasilev, F.; Limatola, N.; Santella, L. Fertilization in Starfish and Sea Urchin: Roles of Actin. Cell Differ. 2018, 65, 33–47. [Google Scholar] [CrossRef]
- Chun, J.T.; Limatola, N.; Vasilev, F.; Santella, L. Early events of fertilization in sea urchin eggs are sensitive to actin-binding organic molecules. Biochem. Biophys. Res. Commun. 2014, 450, 1166–1174. [Google Scholar] [CrossRef]
- Limatola, N.; Chun, J.T.; Santella, L. Effects of Salinity and pH of Seawater on the Reproduction of the Sea Urchin Paracentrotus lividus. Biol. Bull. 2020, 239, 13–23. [Google Scholar] [CrossRef]
- Limatola, N.; Vasilev, F.; Santella, L.; Chun, J.T. Nicotine Induces Polyspermy in Sea Urchin Eggs through a Non-Cholinergic Pathway Modulating Actin Dynamics. Cells 2020, 9, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilney, L.G.; Jaffe, L.A. Actin, microvilli, and the fertilization cone of sea urchin eggs. J. Cell Biol. 1980, 87, 771–782. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.T.; Puppo, A.; Vasilev, F.; Gragnaniello, G.; Garante, E.; Santella, L. The biphasic increase of PIP2 in the fertilized eggs of starfish: New roles in actin polymerization and Ca2+ signaling. PLoS ONE 2010, 5, e14100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilev, F.; Limatola, N.; Park, D.R.; Kim, U.H.; Santella, L.; Chun, J.T. Disassembly of subplasmalemmal actin filaments induces cytosolic Ca2+ increases in Astropecten aranciacus eggs. Cell Physiol. Biochem. 2018, 48, 2011–2034. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.D.; Epel, E.; Paul, M. Intracellular pH and activation of sea urchin eggs after fertilisation. Nature 1976, 262, 661–664. [Google Scholar] [CrossRef]
- Shen, S.S.; Steinhardt, R.A. Direct measurement of intracellular pH during metabolic derepression of the sea urchin egg. Nature 1978, 272, 253–254. [Google Scholar] [CrossRef]
- Steinhardt, R.A.; Mazia, D. Development of K+-conductance and membrane potentials in unfertilized sea urchin eggs after exposure to NH4OH. Nature 1973, 241, 400–401. [Google Scholar] [CrossRef]
- Epel, D. Protein synthesis in sea urchin eggs: A “late” response to fertilization. Proc. Nat. Acad. Sci USA. 1967, 57, 899–906. [Google Scholar] [CrossRef] [Green Version]
- Mazia, D. Chromosome cycles turned on in unfertilized sea urchin eggs exposed to NH40H. Proc. Nat. Acad. Sci. USA 1974, 71, 690–693. [Google Scholar] [CrossRef] [Green Version]
- Wilt, F.H.; Mazia, D. The stimulation of cytoplasmic polyadenylylation in sea urchin eggs by ammonia. Dev. Biol. 1974, 37, 422–424. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Brandriff, B. DNA synthesis in unfertilized sea urchin eggs can be turned on and turned off by the addition and removal of procaine hydrochloride. Dev. Biol. 1975, 47, 12–31. [Google Scholar] [CrossRef]
- Winkler, M.M.; Grainger, J.L. Mechanism of action of NH4Cl and other weak bases in the activation of sea urchin eggs. Nature 1978, 273, 536–538. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Johnson, J.D.; Epel, D. Fertilization acid of sea urchin eggs is not a consequence of cortical granule exocytosis. J. Exp. Zool. 1976, 197, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Begg, D.A.; Rebhun, L.I. pH regulates the polymerization of actin in the sea urchin egg cortex. J. Cell Biol. 1979, 83, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Winkler, M.M.; Steinhardt, R.A.; Grainger, J.L.; Minning, L. Dual ionic controls for the activation of protein synthesis at fertilization. Nature 1980, 287, 558–560. [Google Scholar] [CrossRef]
- Byrd, W.; Belisle, B.W. Microvillar elongation following parthenogenetic activation of sea urchin eggs. Exp. Cell Res. 1985, 159, 211–223. [Google Scholar] [CrossRef]
- Mazia, D.; Schatten, G.; Steinhardt, R. Turning on of activities in unfertilized sea urchin eggs: Correlation with changes of the surface. Proc. Natl. Acad. Sci. USA 1975, 72, 4469–4473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epel, D. The Program of Fertilization. Sci. Am. 1977, 237, 128–139. Available online: http://www.jstor.org/stable/24953924 (accessed on 11 February 2022). [CrossRef] [PubMed]
- Limatola, N.; Chun, J.T.; Cherraben, S.; Schmitt, J.L.; Lehn, J.M.; Santella, L. Effects of Dithiothreitol on fertilization and early development in sea urchin. Cells. 2021, 10, 3573. [Google Scholar] [CrossRef]
- Santella, L.; Limatola, N.; Chun, J.T. Actin cytoskeleton and fertilization in starfish eggs. In Sexual Reproduction in Animals and Plants—Part II: Gametogenesis, Gamete Recognition, Activation, and Evolution; Sawada, H., Inuoe, N., Iwano, M., Eds.; Springer Open: Tokyo, Japan, 2014; pp. 141–155. [Google Scholar] [CrossRef] [Green Version]
- Terasaki, M. Actin filament translocations in sea urchin eggs. Cell Motil. Cytoskelet. 1996, 34, 48–56. [Google Scholar] [CrossRef]
- Rothschild, M.; Swann, M. The fertilization reaction in the sea-urchin egg; a propagated response to sperm attachment. J. Exp. Biol. 1949, 26, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Paul, M.; Epel, D. Fertilization-associated light-scattering changes in eggs of the sea urchin Strongilocentrotus purpuratus. Exp. Cell Res. 1971, 65, 281–288. [Google Scholar] [CrossRef]
- Carroll, E.J.; Epel, D. Isolation and biological activity of the proteases released by sea urchin eggs following fertilization. Dev. Biol. 1975, 44, 22–32. [Google Scholar] [CrossRef]
- Schuel, H. Secretory functions of egg cortical granules in fertilization and development: Secretory functions of egg cortical granules in fertilization and development: A critical review. Gamete Res. 1978, 1, 299–382. [Google Scholar] [CrossRef]
- Wessel, G.M.; Brooks, J.M.; Green, E.; Haley, S.; Voronina, E.; Wong, J.; Zaydfudim, V.; Conner, S. The biology of cortical granules. Int. J. Cytol. 2001, 209, 117–206. [Google Scholar] [CrossRef]
- Shen, S.S.; Steinhardt, R.A. Intracellular pH controls the development of new potassium conductance after fertilization of the sea urchin egg. Exp. Cell Res. 1980, 125, 55–61. [Google Scholar] [CrossRef]
- Epel, D. The initiation of development. Cell Differ. Dev. 1990, 29, 1–12. [Google Scholar] [CrossRef]
- Mazia, D. The release of calcium in Arbacia eggs on fertilization. J. Cell. Comp. Physiol. 1937, 10, 291–304. [Google Scholar] [CrossRef]
- Steinhardt, R.A.; Epel, D. Activation of sea-urchin eggs by a calcium ionophore. Proc. Nat. Acad. Sci. USA 1974, 71, 1915–1919. [Google Scholar] [CrossRef] [Green Version]
- Yonemura, S.; Mabuchi, I. Wave of cortical actin polymerization in the sea urchin egg. Cell Motil. Cytoskelet. 1987, 7, 46–53. [Google Scholar] [CrossRef]
- Collins, F.; Epel, D. The role of calcium ions in the acrosome reaction of sea urchin sperm: Regulation of exocytosis. Exp. Cell Res. 1977, 106, 211222. [Google Scholar] [CrossRef]
- García-Soto, J.; Darszon, A. High pH-induced acrosome reaction and Ca2+ uptake in sea urchin sperm suspended in Na+-free seawater. Dev. Biol. 1985, 110, 338–345. [Google Scholar] [CrossRef]
- Dan, J.C. Studies on the Acrosome. III. Effect of Calcium Deficiency. Biol. Bull. 1954, 107, 335–349. [Google Scholar] [CrossRef]
- Tilney, L.G.; Kiehart, D.P.; Sardet, C.; Tilney, M. Polymerization of actin. IV. Role of Ca++ and H+ in the assembly of actin and in membrane fusion in the acrosomal reaction of echinoderm sperm. J. Cell Biol. 1978, 77, 536–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange, K. Microvillar Ca++ signaling: A new view of an old problem. J. Cell Physiol. 1999, 180, 19–34. [Google Scholar] [CrossRef]
- Lange, K. Microvillar ion channels: Cytoskeletal modulation of ion fluxes. J. Theor. Biol. 2000, 206, 561–584. [Google Scholar] [CrossRef]
- Limatola, N.; Chun, J.T.; Kyozuka, K.; Santella, L. Novel Ca2+ increases in the maturing oocytes of starfish during the germinal vesicle breakdown. Cell Calcium 2015, 58, 500–510. [Google Scholar] [CrossRef]
- Gillot, I.; Ciapa, B.; Payan, P.; Sardet, C. The calcium content of cortical granules and the loss of calcium from sea urchin eggs at fertilization. Dev. Biol. 1991, 146, 396–405. [Google Scholar] [CrossRef]
- McPherson, S.M.; McPherson, P.S.; Mathews, L.; Campbell, K.P.; Longo, F.J. Cortical localization of a calcium release channel in sea urchin eggs. J. Cell Biol. 1992, 116, 1111–1121. [Google Scholar] [CrossRef] [Green Version]
- Morgan, A.J. Sea urchin eggs in the acid reign. Cell Calcium 2011, 50, 147–156. [Google Scholar] [CrossRef]
- Willoughby, D.; Thomas, R.C.; Schwiening, C.J. The effects of intracellular pH changes on resting cytosolic calcium in voltage-clamped snail neurons. J. Physiol. 2001, 530, 405–416. [Google Scholar] [CrossRef]
- Li, S.; Hao, B.; Lu, Y.; Yu, P.; Lee, H.-C.; Yue, J. Intracellular alkalinization induces cytosolic Ca2+ increases by inhibiting sarco/endoplasmic reticulum Ca2+-ATPase (SERCA). PLoS ONE 2012, 7, e31905. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.T.; Santella, L. The actin cytoskeleton in meiotic maturation and fertilization of starfish eggs. Biochem. Biophys. Res. Commun. 2009, 384, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, M. Weak bases mimic the fertilization-associated chloride conductance increase and induce morphological changes in the cortex of Xenopus laevis eggs. Cell Differ. Dev. 1989, 26, 39–51. [Google Scholar] [CrossRef]
- Moody, W., Jr. Effects of intracellular H+ on the electrical properties of excitable cells. Annu. Rev. Neurosci. 1984, 7, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Byrnes, W.M.; Newman, S.A. Ernest Everett Just: Egg and embryo as excitable systems. J. Exp. Zool. B Mol. Dev. Evol. 2014, 322, 191–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, J.M. An experimental study of polyspermy. Biol. Bull. 1936, 70, 361–384. [Google Scholar] [CrossRef]
- Fujimori, T.; Hirai, S. Polyspermy during the course of maturation. Biol. Bull. 1979, 157, 229–257. [Google Scholar] [CrossRef]
- Limatola, N.; Vasilev, F.; Chun, J.T.; Santella, L. Altered actin cytoskeleton in ageing eggs of starfish affects fertilization process. Exp. Cell Res. 2019, 381, 179–190. [Google Scholar] [CrossRef]
- Longo, F.J.; Woerner, M.; Chiba, K.; Hoshi, M. Cortical changes in starfish (Asterina pectinifera) oocytes during 1-methyladenine-induced maturation and fertilisation/activation. Zygote 1995, 3, 225–239. [Google Scholar] [CrossRef]
- Miyazaki, S.I.; Ohmori, H.; Sasaki, S. Potassium rectifications of the starfish oocyte membrane and their changes during oocyte maturation. J. Physiol. 1975, 246, 55–78. [Google Scholar] [CrossRef] [Green Version]
- Mohri, T.; Kyozuka, K. Starfish oocytes of A. pectinifera reveal marked differences in sperm-induced electrical and intracellular calcium changes during oocyte maturation and at fertilization. Mol. Reprod. Dev. 2022, 89, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.; Lange, K.; Santella, L. Activation of oocytes by latrunculin A. FASEB J. 2002, 16, 1050–1056. [Google Scholar] [CrossRef] [PubMed]
- Moccia, F. Latrunculin A depolarizes starfish oocytes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 148, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Loeb, J. The comparative efficiency of weak and strong bases in artificial parthenogenesis. J. Exp. Zool. 1912, 13, 577–590. [Google Scholar] [CrossRef]
- Boron, W.F.; Roos, A.; De Weer, P. NH4Cl and other weak bases in the activation of sea urchin eggs. Nature 1978, 274, 190. [Google Scholar] [CrossRef] [PubMed]
- Madshus, I.H. Regulation of intracellular pH in eukaryotic cells. Biochem. J. 1988, 250, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Just, E.E. The fertilization-reaction in Echinarachnius Parma. V. The existence in the inseminated egg of a period of special susceptibility to hypotonic seawater. Am. J. Physiol. 1922, 61, 516–527. Available online: http://dh.howard.edu/bio_fac (accessed on 11 February 2022). [CrossRef]
- Limatola, N.; Chun, J.T.; Santella, L. Fertilization and development of Arbacia lixula eggs are affected by osmolality conditions. BioSystems 2021, 206, 104448. [Google Scholar] [CrossRef]
- Busa, W.B.; Nuccitelli, R. Metabolic regulation via intracellular pH. Am. J. Physiol. 1984, 246, 409–438. [Google Scholar] [CrossRef] [Green Version]
- Nishigaki, T.; José, O.; González-Cota, A.; Romero, F.; Treviño, C.L.; Darszon, A. Intracellular pH in sperm physiology. Biochem Biophys. Res. Commun. 2014, 450, 1149–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molinari, G.; Nervo, E. Role of protons in calcium signaling. Biochem. J. 2021, 478, 895–910. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. lividus NSW | P. lividus pH 9 | A. lixula NSW | A. lixula pH 9 | |
|---|---|---|---|---|
| Mean | 0 | 100 * | 5 | 100 |
| SD | 0 | 0 | 0.01 | 0 |
| n | 4 | 4 | 2 | 2 |
| P. lividus NSW | P. lividus pH 9 | A. lixula NSW | A. lixula pH 9 | |
|---|---|---|---|---|
| Mean | 1 | 12.89 * | 1.07 | 6.07 * |
| SD | 0 | 5.79 | 0.35 | 1.33 |
| n | 80 | 80 | 40 | 40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limatola, N.; Chun, J.T.; Santella, L. Regulation of the Actin Cytoskeleton-Linked Ca2+ Signaling by Intracellular pH in Fertilized Eggs of Sea Urchin. Cells 2022, 11, 1496. https://doi.org/10.3390/cells11091496
Limatola N, Chun JT, Santella L. Regulation of the Actin Cytoskeleton-Linked Ca2+ Signaling by Intracellular pH in Fertilized Eggs of Sea Urchin. Cells. 2022; 11(9):1496. https://doi.org/10.3390/cells11091496
Chicago/Turabian StyleLimatola, Nunzia, Jong Tai Chun, and Luigia Santella. 2022. "Regulation of the Actin Cytoskeleton-Linked Ca2+ Signaling by Intracellular pH in Fertilized Eggs of Sea Urchin" Cells 11, no. 9: 1496. https://doi.org/10.3390/cells11091496
APA StyleLimatola, N., Chun, J. T., & Santella, L. (2022). Regulation of the Actin Cytoskeleton-Linked Ca2+ Signaling by Intracellular pH in Fertilized Eggs of Sea Urchin. Cells, 11(9), 1496. https://doi.org/10.3390/cells11091496