
Intraocular Adeno-Associated Virus-Mediated Transgene Endothelin-1 Delivery to the Rat Eye Induces Functional Changes Indicative of Retinal Ischemia—A Potential Chronic Glaucoma Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. In Vivo Experiments
2.2.1. Acute ET-1 Injections
2.2.2. Vector Selection
2.2.3. Transgene ET-1 Administration
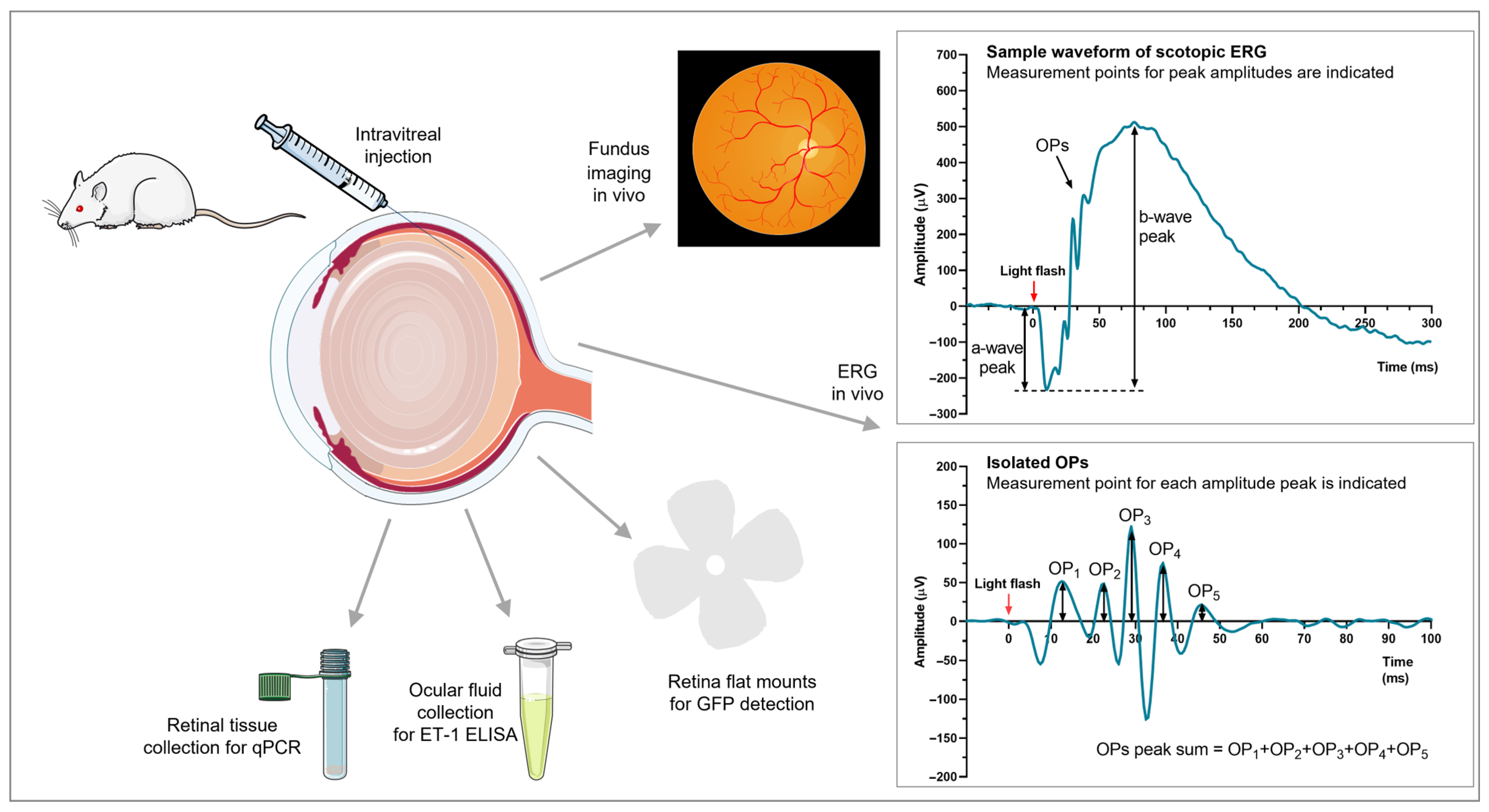
2.3. Intravitreal Injections
2.4. Fundus Imaging
2.5. Electroretinography
2.6. Post-Mortem Enucleation and Tissue Processing
2.6.1. Retina Flat Mounts and Cryosections
2.6.2. Ocular Fluid Collection and Retina for qRT-PCR
2.7. ELISA
2.8. Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results
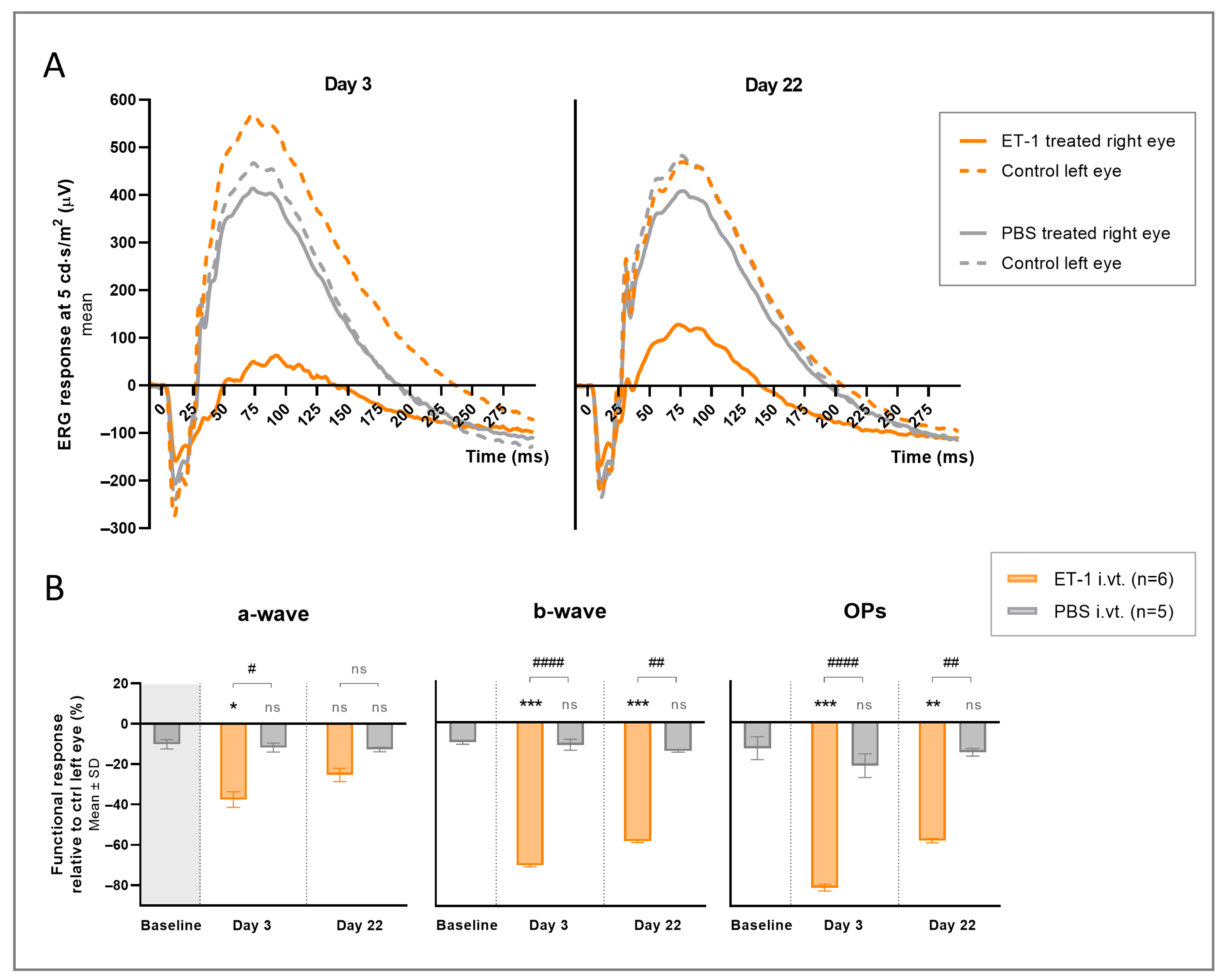
3.1. Acute Intravitreal ET-1 Administration
3.2. Vector Selection
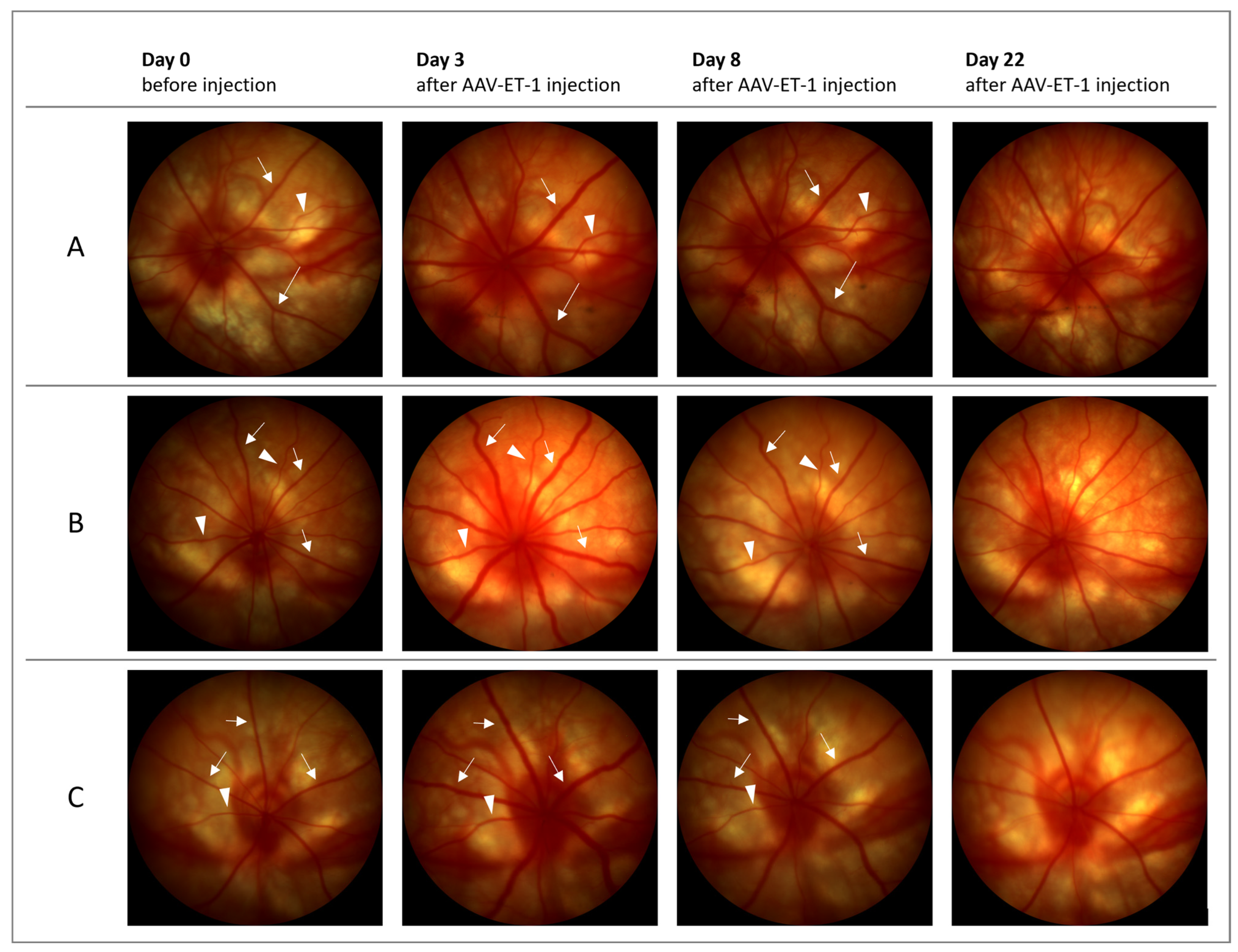
3.3. ET-1 Delivery to the Rat Retina via AAV-Mediated Gene Transfer
3.4. Compensatory Response
3.5. Follow-Up Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Resnikoff, S.; Pascolini, D.; Etya’ale, D.; Kocur, I.; Pararajasegaram, R.; Pokharel, G.P.; Mariotti, S.P. Global data on visual impairment in the year 2002. Bull. World Health Organ. 2004, 82, 844–851. [Google Scholar]
- Collaborative Normal-Tension Glaucoma Study Group. Comparison of glaucomatous progression between untreated patients with normal-tension glaucoma and patients with therapeutically reduced intraocular pressures. Am. J. Ophthalmol. 1998, 126, 487–497. [Google Scholar] [CrossRef]
- Killer, H.E.; Pircher, A. Normal tension glaucoma: Review of current understanding and mechanisms of the pathogenesis. Eye 2018, 32, 924–930. [Google Scholar] [CrossRef]
- Flammer, J.; Mozaffarieh, M. What is the present pathogenetic concept of glaucomatous optic neuropathy? Surv. Ophthalmol. 2007, 52 (Suppl. S2), S162–S173. [Google Scholar] [CrossRef]
- Garhöfer, G.; Fuchsjäger-Mayrl, G.; Vass, C.; Pemp, B.; Hommer, A.; Schmetterer, L. Retrobulbar blood flow velocities in open angle glaucoma and their association with mean arterial blood pressure. Investig. Opthalmol. Vis. Sci. 2010, 51, 6652–6657. [Google Scholar] [CrossRef] [Green Version]
- Berisha, F.; Feke, G.T.; Hirose, T.; McMeel, J.W.; Pasquale, L.R. Retinal blood flow and nerve fiber layer measurements in early-stage open-angle glaucoma. Am. J. Ophthalmol. 2008, 146, 466–472.e2. [Google Scholar] [CrossRef]
- Galassi, F.; Giambene, B.; Varriale, R. Systemic vascular dysregulation and retrobulbar hemodynamics in normal-tension glaucoma. Investig. Ophthalmol. Vis. Sci. 2011, 52, 4467–4471. [Google Scholar] [CrossRef] [Green Version]
- Su, W.-W.; Cheng, S.-T.; Ho, W.-J.; Tsay, P.-K.; Wu, S.-C.; Chang, S.H. Glaucoma is associated with peripheral vascular endothelial dysfunction. Ophthalmology 2008, 115, 1173–1178.e1. [Google Scholar] [CrossRef]
- Costa, V.P.; Harris, A.; Anderson, D.; Stodtmeister, R.; Cremasco, F.; Kergoat, H.; Lovasik, J.; Stalmans, I.; Zeitz, O.; Lanzl, I.; et al. Ocular perfusion pressure in glaucoma. Acta Ophthalmol. 2014, 92, e252–e266. [Google Scholar] [CrossRef]
- Zheng, Y.; Wong, T.Y.; Mitchell, P.; Friedman, D.S.; He, M.; Aung, T. Distribution of ocular perfusion pressure and its relationship with open-angle glaucoma: The singapore malay eye study. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3399–3404. [Google Scholar] [CrossRef] [Green Version]
- Stefánsson, E.; Olafsdottir, O.B.; Eliasdottir, T.S.; Vehmeijer, W.; Einarsdottir, A.B.; Bek, T.; Torp, T.L.; Grauslund, J.; Eysteinsson, T.; Karlsson, R.A.; et al. Retinal oximetry: Metabolic imaging for diseases of the retina and brain. Prog. Retin. Eye Res. 2019, 70, 1–22. [Google Scholar] [CrossRef]
- Osborne, N.N.; Casson, R.J.; Wood, J.P.; Chidlow, G.; Graham, M.; Melena, J. Retinal ischemia: Mechanisms of damage and potential therapeutic strategies. Prog. Retin. Eye Res. 2004, 23, 91–147. [Google Scholar] [CrossRef]
- Yorio, T.; Krishnamoorthy, R.; Prasanna, G. Endothelin: Is it a contributor to glaucoma pathophysiology? J. Glaucoma 2002, 11, 259–270. [Google Scholar] [CrossRef]
- Noske, W.; Hensen, J.; Wiederholt, M. Endothelin-like immunoreactivity in aqueous humor of patients with primary open-angle glaucoma and cataract. Graefe’s Arch. Clin. Exp. Ophthalmol. 1997, 235, 551–552. [Google Scholar] [CrossRef]
- Mihaylova, B.; Petkova, I.; Rankova-Yotova, C.; Dimitrova, G.; Tanev, I.; Ivanova, S.; Tsakova, A.; Svinarov, D. Plasma endothelin-1 and endothelin-A receptor concentrations in patients with primary open-angle glaucoma. Biotechnol. Biotechnol. Equip. 2017, 31, 782–787. [Google Scholar] [CrossRef] [Green Version]
- Henry, E.; Newby, D.E.; Webb, D.J.; Hadoke, P.W.F.; O’Brien, C.J. Altered endothelin-1 vasoreactivity in patients with untreated normal-pressure glaucoma. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2528–2532. [Google Scholar] [CrossRef] [Green Version]
- Buckley, C.; Hadoke, P.W.F.; Henry, E.; O’Brien, C. Systemic vascular endothelial cell dysfunction in normal pressure glaucoma. Br. J. Ophthalmol. 2002, 86, 227–232. [Google Scholar] [CrossRef]
- Sasaoka, M.; Taniguchi, T.; Shimazawa, M.; Ishida, N.; Shimazaki, A.; Hara, H. Intravitreal injection of endothelin-1 caused optic nerve damage following to ocular hypoperfusion in rabbits. Exp. Eye Res. 2006, 83, 629–637. [Google Scholar] [CrossRef]
- Oku, H.; Sugiyama, T.; Kojima, S.; Watanabe, T.; Azuma, I. Experimental optic cup enlargement caused by endothelin-1-induced chronic optic nerve head ischemia. Surv. Ophthalmol. 1999, 44 (Suppl. S1), S74–S84. [Google Scholar] [CrossRef]
- Leonova, J.; Thorlin, T.; Åberg, N.D.; Eriksson, P.S.; Rönnbäck, L.; Hansson, E. Endothelin-1 decreases glutamate uptake in primary cultured rat astrocytes. Am. J. Physiol. Cell Physiol. 2001, 281, C1495–C1503. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Oku, H.; Fukuhara, M.; Kojima, S.; Komori, A.; Ichikawa, M.; Katsumura, K.; Kobayashi, M.; Sugiyama, T.; Ikeda, T. Endothelin-1 enhances glutamate-induced retinal cell death, possibly through ETA receptors. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4684–4690. [Google Scholar] [CrossRef] [Green Version]
- Cheung, S.S.F.; Leung, J.W.C.; Lam, A.K.M.; Lam, K.S.L.; Chung, S.S.M.; Lo, A.C.Y.; Chung, S.K. Selective over-expression of endothelin-1 in endothelial cells exacerbates inner retinal edema and neuronal death in ischemic retina. PLoS ONE 2011, 6, e26184. [Google Scholar] [CrossRef] [Green Version]
- McGrady, N.R.; Minton, A.Z.; Stankowska, D.L.; He, S.; Jefferies, H.B.; Krishnamoorthy, R.R. Upregulation of the endothelin A (ETA) receptor and its association with neurodegeneration in a rodent model of glaucoma. BMC Neurosci. 2017, 18, 27. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; LeVatte, T.L.; Archibald, M.L.; Chauhan, B.C. Increase in endothelin B receptor expression in optic nerve astrocytes in endothelin-1 induced chronic experimental optic neuropathy. Exp. Eye Res. 2009, 88, 378–385. [Google Scholar] [CrossRef]
- Wang, X.; Baldridge, W.H.; Chauhan, B.C. Acute endothelin-1 application induces reversible fast axonal transport blockade in adult rat optic nerve. Investig. Ophthalmol. Vis. Sci. 2008, 49, 961–967. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, G.; Krishnamoorthy, R.; Clark, A.F.; Wordinger, R.J.; Yorio, T. Human optic nerve head astrocytes as a target for endothelin-1. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2704–2713. [Google Scholar]
- Bourque, S.L.; Davidge, S.T.; Adams, M.A. The interaction between endothelin-1 and nitric oxide in the vasculature: New perspectives. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1288–R1295. [Google Scholar] [CrossRef] [Green Version]
- Venkataraman, S.T.; Flanagan, J.G.; Hudson, C. Vascular reactivity of optic nerve head and retinal blood vessels in glaucoma—A review. Microcirculation 2010, 17, 568–581. [Google Scholar] [CrossRef]
- Cioffi, G.A.; Orgül, S.; Onda, E.; Bacon, D.R.; Van Buskirk, E.M. An in vivo model of chronic optic nerve ischemia: The dose-dependent effects of endothelin-1 on the optic nerve microvasculature. Curr. Eye Res. 1995, 14, 1147–1153. [Google Scholar] [CrossRef]
- Orgül, S.; Cioffi, G.A.; Wilson, D.J.; Bacon, D.R.; Van Buskirk, E.M. An endothelin-1 induced model of optic nerve ischemia in the rabbit. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1860–1869. [Google Scholar]
- Masuzawa, K.; Jesmin, S.; Maeda, S.; Kaji, Y.; Oshika, T.; Zaedi, S.; Shimojo, N.; Yaji, N.; Miyauchi, T.; Goto, K. A model of retinal ischemia-reperfusion injury in rats by subconjunctival injection of endothelin-1. Exp. Biol. Med. 2006, 231, 1085–1089. [Google Scholar]
- Chauhan, B.C.; LeVatte, T.L.; Jollimore, C.A.; Yu, P.K.; Reitsamer, H.A.; Kelly, M.E.M.; Yu, D.-Y.; Tremblay, F.; Archibald, M.L. Model of endothelin-1-induced chronic optic neuropathy in rat. Investig. Ophthalmol. Vis. Sci. 2004, 45, 144–152. [Google Scholar] [CrossRef]
- Bursell, S.E.; Clermont, A.C.; Oren, B.; King, G.L. The in vivo effect of endothelins on retinal circulation in nondiabetic and diabetic rats. Investig. Ophthalmol. Vis. Sci. 1995, 36, 596–607. [Google Scholar]
- Nagata, A.; Omachi, K.; Higashide, T.; Shirae, S.; Shimazaki, A.; Nakamura, M.; Ishida, N.; Sugiyama, K. OCT evaluation of neuroprotective effects of tafluprost on retinal injury after intravitreal injection of endothelin-1 in the rat eye. Investig. Ophthalmol. Vis. Sci. 2014, 55, 1040–1047. [Google Scholar] [CrossRef] [Green Version]
- Lau, J.; Dang, M.; Hockmann, K.; Ball, A.K. Effects of acute delivery of endothelin-1 on retinal ganglion cell loss in the rat. Exp. Eye Res. 2006, 82, 132–145. [Google Scholar] [CrossRef]
- Arfuzir, N.N.N.; Agarwal, R.; Iezhitsa, I.; Agarwal, P.; Ismail, N.M. Dose-Dependent Effects of Endothelin-1 on Retinal and Optic Nerve Morphology in Sprague Dawley Rats. Neurochem. J. 2019, 13, 73–80. [Google Scholar] [CrossRef]
- Mi, X.-S.; Zhang, X.; Feng, Q.; Lo, A.C.Y.; Chung, S.K.; So, K.-F. Progressive retinal degeneration in transgenic mice with overexpression of endothelin-1 in vascular endothelial cells. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4842–4851. [Google Scholar] [CrossRef] [Green Version]
- Dang, T.M.; Tsai, T.I.; Vingrys, A.J.; Bui, B.V. Post-receptoral contributions to the rat scotopic electroretinogram a-wave. Doc. Ophthalmol. 2011, 122, 149–156. [Google Scholar] [CrossRef]
- Robson, J.G.; Saszik, S.M.; Ahmed, J.; Frishman, L.J. Rod and cone contributions to the a-wave of the electroretinogram of the macaque. J. Physiol. 2003, 547, 509–530. [Google Scholar] [CrossRef]
- Robson, J.G.; Frishman, L.J. Dissecting the dark-adapted electroretinogram. Doc. Ophthalmol. 1998, 95, 187–215. [Google Scholar] [CrossRef]
- Möller, A.; Eysteinsson, T. Modulation of the components of the rat dark-adapted electroretinogram by the three subtypes of GABA receptors. Vis. Neurosci. 2003, 20, 535–542. [Google Scholar] [CrossRef]
- Dong, C.-J.; Agey, P.; Hare, W.A. Origins of the electroretinogram oscillatory potentials in the rabbit retina. Vis. Neurosci. 2004, 21, 533–543. [Google Scholar] [CrossRef]
- Bretag, A.H. Synthetic interstial fluid for isolated mammalian tissue. Life Sci. 1969, 8, 319–329. [Google Scholar] [CrossRef]
- Smith, C.A.; Chauhan, B.C. In vivo imaging of adeno-associated viral vector labelled retinal ganglion cells. Sci. Rep. 2018, 8, 1490. [Google Scholar] [CrossRef] [Green Version]
- Katada, Y.; Kobayashi, K.; Tsubota, K.; Kurihara, T. Evaluation of AAV-DJ vector for retinal gene therapy. PeerJ 2019, 7, e6317. [Google Scholar] [CrossRef]
- Daniels, D.M.; Shen, W.-Y.; Constable, I.J.; Rakoczy, P.E. Quantitative model demonstrating that recombinant adeno-associated virus and green fluorescent protein are non-toxic to the rat retina. Clin. Exp. Ophthalmol. 2003, 31, 439–444. [Google Scholar] [CrossRef]
- Ciulla, T.A.; Pawlyk, B.S.; Harris, A.; Oberoi, A.; Miller, J.W.; Sandberg, M.A. Endothelin-1-mediated retinal artery vasospasm and the rabbit electroretinogram. J. Ocul. Pharmacol. Ther. 2000, 16, 393–398. [Google Scholar] [CrossRef]
- Takei, K.; Sato, T.; Nonoyama, T.; Miyauchi, T.; Goto, K.; Hommura, S. A new model of transient complete obstruction of retinal vessels induced by endothelin-1 injection into the posterior vitreous body in rabbits. Graefe’s Arch. Clin. Exp. Ophthalmol. 1993, 231, 476–481. [Google Scholar] [CrossRef]
- De Schaepdrijver, L.; Simoens, P.; Lauwers, H.; De Geest, J.P. Retinal vascular patterns in domestic animals. Res. Vet. Sci. 1989, 47, 34–42. [Google Scholar] [CrossRef]
- Lee, D.; Jeong, H.; Miwa, Y.; Shinojima, A.; Katada, Y.; Tsubota, K.; Kurihara, T. Retinal dysfunction induced in a mouse model of unilateral common carotid artery occlusion. PeerJ 2021, 9, e11665. [Google Scholar] [CrossRef]
- Ogishima, H.; Nakamura, S.; Nakanishi, T.; Imai, S.; Kakino, M.; Ishizuka, F.; Tsuruma, K.; Shimazawa, M.; Hara, H. Ligation of the pterygopalatine and external carotid arteries induces ischemic damage in the murine retina. Investig. Ophthalmol. Vis. Sci. 2011, 52, 9710–9720. [Google Scholar] [CrossRef]
- Zhong, L. Electroretinography (ERG) amplitude changes in a rat retinal ischemia model. In Proceedings of the 2021 ARVO Annual Meeting, Online, 1–7 May 2021; p. 601. [Google Scholar]
- Rosenbaum, D.M.; Rosenbaum, P.S.; Singh, M.; Gupta, G.; Gupta, H.; Li, B.; Roth, S. Functional and morphologic comparison of two methods to produce transient retinal ischemia in the rat. J. Neuroophthalmol. 2001, 21, 62–68. [Google Scholar] [CrossRef] [Green Version]
- Chenda, S.; Lee, Y.J.; Park, J.Y.; Ohn, Y.H. Electroretinographic findings in patients with central retinal vein occlusion. J. Soonchunhyang Med. Sci. 2009, 15, 33–40. [Google Scholar]
- Kim, H.M.; Park, K.H.; Woo, S.J. Correlation of electroretinography components with visual function and prognosis of central retinal artery occlusion. Sci. Rep. 2020, 10, 12146. [Google Scholar] [CrossRef]
- Grozdanic, S.D.; Sakaguchi, D.S.; Kwon, Y.H.; Kardon, R.H.; Sonea, I.M. Functional characterization of retina and optic nerve after acute ocular ischemia in rats. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2597–2605. [Google Scholar] [CrossRef] [Green Version]
- Bayer, A.U.; Danias, J.; Brodie, S.; Maag, K.P.; Chen, B.; Shen, F.; Podos, S.M.; Mittag, T.W. Electroretinographic abnormalities in a rat glaucoma model with chronic elevated intraocular pressure. Exp. Eye Res. 2001, 72, 667–677. [Google Scholar] [CrossRef]
- Gur, M.; Zeevi, Y.Y.; Bielik, M.; Neumann, E. Changes in the oscillatory potentials of the electroretinogram in glaucoma. Curr. Eye Res. 1987, 6, 457–466. [Google Scholar] [CrossRef]
- Vaegan; Graham, S.L.; Goldberg, I.; Millar, T.J. Selective reduction of oscillatory potentials and pattern electroretinograms after retinal ganglion cell damage by disease in humans or by kainic acid toxicity in cats. Doc. Ophthalmol. 1991, 77, 237–253. [Google Scholar] [CrossRef]
- Wilsey, L.J.; Fortune, B. Electroretinography in glaucoma diagnosis. Curr. Opin. Ophthalmol. 2016, 27, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Leydhecker, G. The electroretinogram in glaucomatous eyes. Br. J. Ophthalmol. 1950, 34, 550–554. [Google Scholar] [CrossRef] [Green Version]
- Kendell, K.R.; Quigley, H.A.; Kerrigan, L.A.; Pease, M.E.; Quigley, E.N. Primary open-angle glaucoma is not associated with photoreceptor loss. Investig. Ophthalmol. Vis. Sci. 1995, 36, 200–205. [Google Scholar]
- Velten, I.M.; Korth, M.; Horn, F.K. The a-wave of the dark adapted electroretinogram in glaucomas: Are photoreceptors affected? Br. J. Ophthalmol. 2001, 85, 397–402. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.S.; Zawadzki, R.J.; Lim, M.C.; Brandt, J.D.; Keltner, J.L.; Doble, N.; Werner, J.S. Evidence of outer retinal changes in glaucoma patients as revealed by ultrahigh-resolution in vivo retinal imaging. Br. J. Ophthalmol. 2011, 95, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Ortín-Martínez, A.; Salinas-Navarro, M.; Nadal-Nicolás, F.M.; Jiménez-López, M.; Valiente-Soriano, F.J.; García-Ayuso, D.; Bernal-Garro, J.M.; Avilés-Trigueros, M.; Agudo-Barriuso, M.; Villegas-Pérez, M.P.; et al. Laser-induced ocular hypertension in adult rats does not affect non-RGC neurons in the ganglion cell layer but results in protracted severe loss of cone-photoreceptors. Exp. Eye Res. 2015, 132, 17–33. [Google Scholar] [CrossRef] [Green Version]
- Vaegan; Graham, S.L.; Goldberg, I.; Buckland, L.; Hollows, F.C. Flash and pattern electroretinogram changes with optic atrophy and glaucoma. Exp. Eye Res. 1995, 60, 697–706. [Google Scholar] [CrossRef]
- Yin, Z.Q.; Vaegan; Millar, T.J.; Beaumont, P.; Sarks, S. Widespread choroidal insufficiency in primary open-angle glaucoma. J. Glaucoma 1997, 6, 23–32. [Google Scholar] [CrossRef]
- Louzada-Junior, P.; Dias, J.J.; Santos, W.F.; Lachat, J.J.; Bradford, H.F.; Coutinho-Netto, J. Glutamate release in experimental ischaemia of the retina: An approach using microdialysis. J. Neurochem. 1992, 59, 358–363. [Google Scholar] [CrossRef]
- Adachi, K.; Fujita, Y.; Morizane, C.; Akaike, A.; Ueda, M.; Satoh, M.; Masai, H.; Kashii, S.; Honda, Y. Inhibition of NMDA receptors and nitric oxide synthase reduces ischemic injury of the retina. Eur. J. Pharmacol. 1998, 350, 53–57. [Google Scholar] [CrossRef]
- Russo, R.; Cavaliere, F.; Varano, G.P.; Milanese, M.; Adornetto, A.; Nucci, C.; Bonanno, G.; Morrone, L.A.; Corasaniti, M.T.; Bagetta, G. Impairment of neuronal glutamate uptake and modulation of the glutamate transporter GLT-1 induced by retinal ischemia. PLoS ONE 2013, 8, e69250. [Google Scholar] [CrossRef] [Green Version]
- Izumi, Y.; Hammerman, S.B.; Kirby, C.O.; Benz, A.M.; Olney, J.W.; Zorumski, C.F. Involvement of glutamate in ischemic neurodegeneration in isolated retina. Vis. Neurosci. 2003, 20, 97–107. [Google Scholar] [CrossRef]
- Khodorova, A.; Richter, J.; Vasko, M.R.; Strichartz, G. Early and late contributions of glutamate and CGRP to mechanical sensitization by endothelin-1. J. Pain 2009, 10, 740–749. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, Y.; Takimoto, M.; Oda, K.; Früh, T.; Takai, M.; Okada, T.; Hori, S. Endothelin evokes efflux of glutamate in cultures of rat astrocytes. J. Neurochem. 1997, 68, 2194–2200. [Google Scholar] [CrossRef]
- Matsuura, S.; Ikegaya, Y.; Yamada, M.K.; Nishiyama, N.; Matsuki, N. Endothelin downregulates the glutamate transporter GLAST in cAMP-differentiated astrocytes in vitro. Glia 2002, 37, 178–182. [Google Scholar] [CrossRef]
- Connaughton, V. Glutamate and Glutamate Receptors in Vertebrate Retina. In Webvision: The Organization of the Retina and Visual System; Kolb, H., Fernandez, E., Nelson, R., Eds.; University of Utah Health Sciences Center: Salt Lake City, UT, USA, 2005. [Google Scholar]
- Rangaswamy, N.V.; Hood, D.C.; Frishman, L.J. Regional variations in local contributions to the primate photopic flash ERG: Revealed using the slow-sequence mfERG. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3233–3247. [Google Scholar] [CrossRef]
- Harada, T.; Harada, C.; Watanabe, M.; Inoue, Y.; Sakagawa, T.; Nakayama, N.; Sasaki, S.; Okuyama, S.; Watase, K.; Wada, K.; et al. Functions of the two glutamate transporters GLAST and GLT-1 in the retina. Proc. Natl. Acad. Sci. USA 1998, 95, 4663–4666. [Google Scholar] [CrossRef]
- Hayreh, S.S.; Weingeist, T.A. Experimental occlusion of the central artery of the retina. IV: Retinal tolerance time to acute ischaemia. Br. J. Ophthalmol. 1980, 64, 818–825. [Google Scholar] [CrossRef] [Green Version]
- Barnett, N.L.; Osborne, N.N. Redistribution of GABA immunoreactivity following central retinal artery occlusion. Brain Res. 1995, 677, 337–340. [Google Scholar] [CrossRef]
- Holcombe, D.J.; Lengefeld, N.; Gole, G.A.; Barnett, N.L. Selective inner retinal dysfunction precedes ganglion cell loss in a mouse glaucoma model. Br. J. Ophthalmol. 2008, 92, 683–688. [Google Scholar] [CrossRef]
- Takei, K.; Sato, T.; Nonoyama, T.; Hommura, S.; Miyauchi, T.; Goto, K. Analysis of vasocontractile responses to endothelin-1 in rabbit retinal vessels using an ETA receptor antagonist and an ETB receptor agonist. Life Sci. 1993, 53, PL111–PL115. [Google Scholar] [CrossRef]
- Hein, T.W.; Ren, Y.; Yuan, Z.; Xu, W.; Somvanshi, S.; Nagaoka, T.; Yoshida, A.; Kuo, L. Functional and molecular characterization of the endothelin system in retinal arterioles. Investig. Ophthalmol. Vis. Sci. 2009, 50, 3329–3336. [Google Scholar] [CrossRef]
- Berthiaume, N.; Yanagisawa, M.; Labonte, J.; D’Orleans-Juste, P. Heterozygous knock-Out of ETB receptors induces BQ-123-sensitive hypertension in the mouse. Hypertension 2000, 36, 1002–1007. [Google Scholar] [CrossRef] [Green Version]
- Verhaar, M.C.; Strachan, F.E.; Newby, D.E.; Cruden, N.L.; Koomans, H.A.; Rabelink, T.J.; Webb, D.J. Endothelin-A receptor antagonist-mediated vasodilatation is attenuated by inhibition of nitric oxide synthesis and by endothelin-B receptor blockade. Circulation 1998, 97, 752–756. [Google Scholar] [CrossRef] [Green Version]
- Hasselblatt, M.; Lewczuk, P.; Löffler, B.-M.; Kamrowski-Kruck, H.; Von Ahsen, N.; Sirén, A.-L.; Ehrenreich, H. Role of the astrocytic ETB receptor in the regulation of extracellular endothelin-1 during hypoxia. Glia 2001, 34, 18–26. [Google Scholar] [CrossRef]
- Fukuroda, T.; Fujikawa, T.; Ozaki, S.; Ishikawa, K.; Yano, M.; Nishikibe, M. Clearance of circulating endothelin-1 by ETB receptors in rats. Biochem. Biophys. Res. Commun. 1994, 199, 1461–1465. [Google Scholar] [CrossRef]
- Rogers, S.D.; Demaster, E.; Catton, M.; Ghilardi, J.R.; Levin, L.A.; Maggio, J.E.; Mantyh, P.W. Expression of endothelin-B receptors by glia in vivo is increased after CNS injury in rats, rabbits, and humans. Exp. Neurol. 1997, 145, 180–195. [Google Scholar] [CrossRef]
- Koyama, Y.; Takemura, M.; Fujiki, K.; Ishikawa, N.; Shigenaga, Y.; Baba, A. BQ788, an endothelin ETB receptor antagonist, attenuates stab wound injury-induced reactive astrocytes in rat brain. Glia 1999, 26, 268–271. [Google Scholar] [CrossRef]
- Minton, A.Z.; Phatak, N.R.; Stankowska, D.L.; He, S.; Ma, H.-Y.; Mueller, B.H.; Jiang, M.; Luedtke, R.; Yang, S.; Brownlee, C.; et al. Endothelin B receptors contribute to retinal ganglion cell loss in a rat model of glaucoma. PLoS ONE 2012, 7, e43199. [Google Scholar] [CrossRef]
- MacCumber, M.W.; D’Anna, S.A. Endothelin receptor-binding subtypes in the human retina and choroid. Arch. Ophthalmol. 1994, 112, 1231–1235. [Google Scholar] [CrossRef]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K.; Krause, D.N. CGRP as the target of new migraine therapies—Successful translation from bench to clinic. Nat. Rev. Neurol. 2018, 14, 338–350. [Google Scholar] [CrossRef]
- Tippins, J.R. CGRP: A novel neuropeptide from the calcitonin gene is the most potent vasodilator known. J. Hypertens. Suppl. 1986, 4, S102–S105. [Google Scholar] [PubMed]
- Meens, M.J.P.M.T.; Fazzi, G.E.; van Zandvoort, M.A.; De Mey, J.G.R. Calcitonin gene-related peptide selectively relaxes contractile responses to endothelin-1 in rat mesenteric resistance arteries. J. Pharmacol. Exp. Ther. 2009, 331, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Meens, M.J.P.M.T.; Compeer, M.G.; Hackeng, T.M.; van Zandvoort, M.A.; Janssen, B.J.A.; De Mey, J.G.R. Stimuli of sensory-motor nerves terminate arterial contractile effects of endothelin-1 by CGRP and dissociation of ET-1/ETA-receptor complexes. PLoS ONE 2010, 5, e10917. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, D.H. Prevention of endothelin-1-induced increases in blood pressure: Role of endogenous CGRP. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1868–H1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.-H.; Zhang, Y.-Q.; Guo, Z. Endogenous CGRP protects retinal cells against stress induced apoptosis in rats. Neurosci. Lett. 2011, 501, 83–85. [Google Scholar] [CrossRef] [PubMed]
- Toth, C.C.; Willis, D.; Twiss, J.L.; Walsh, S.; Martinez, J.A.; Liu, W.-Q.; Midha, R.; Zochodne, D.W. Locally synthesized calcitonin gene-related Peptide has a critical role in peripheral nerve regeneration. J. Neuropathol. Exp. Neurol. 2009, 68, 326–337. [Google Scholar] [CrossRef] [Green Version]
- Blixt, F.W.; Radziwon-Balicka, A.; Edvinsson, L.; Warfvinge, K. Distribution of CGRP and its receptor components CLR and RAMP1 in the rat retina. Exp. Eye Res. 2017, 161, 124–131. [Google Scholar] [CrossRef]
- Kerr, N.M.; Johnson, C.S.; Zhang, J.; Eady, E.K.; Green, C.R.; Danesh-Meyer, H.V. High pressure-induced retinal ischaemia reperfusion causes upregulation of gap junction protein connexin43 prior to retinal ganglion cell loss. Exp. Neurol. 2012, 234, 144–152. [Google Scholar] [CrossRef]
- Tang, X.; Tzekov, R.; Passaglia, C.L. Retinal cross talk in the mammalian visual system. J. Neurophysiol. 2016, 115, 3018–3029. [Google Scholar] [CrossRef] [Green Version]
- Nadal-Nicolás, F.M.; Valiente-Soriano, F.J.; Salinas-Navarro, M.; Jiménez-López, M.; Vidal-Sanz, M.; Agudo-Barriuso, M. Retino-retinal projection in juvenile and young adult rats and mice. Exp. Eye Res. 2015, 134, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Lucas-Ruiz, F.; Galindo-Romero, C.; Rodríguez-Ramírez, K.T.; Vidal-Sanz, M.; Agudo-Barriuso, M. Neuronal Death in the Contralateral Un-Injured Retina after Unilateral Axotomy: Role of Microglial Cells. Int. J. Mol. Sci. 2019, 20, 5733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kergoat, H.; Lovasik, J.V. Unilateral ocular vascular stress in man and retinal responsivity in the contralateral eye. Ophthalmic Physiol. Opt. 1994, 14, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Nicolás, F.M.; Jiménez-López, M.; Salinas-Navarro, M.; Sobrado-Calvo, P.; Vidal-Sanz, M.; Agudo-Barriuso, M. Microglial dynamics after axotomy-induced retinal ganglion cell death. J. Neuroinflamm. 2017, 14, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak Amplitudes. Right Eye (Mean µV ± SD) | ||||||
|---|---|---|---|---|---|---|
| Wave- form Light Intensity * | Day 3 | Day 22 | ||||
| ET-1-Treated | PBS-Treated | p-Value # | ET-1-Treated | PBS-Treated | p-Value # | |
| a-wave | ns | ns | ||||
| 0.1 | −101.7 ± 28.7 | −108.2 ± 25.3 | −102.7 ± 36.7 | −94.5 ± 22.9 | ||
| 1 | −139.1 ± 37.1 | −166.6 ± 37.3 | −147.7 ± 42.0 | −166.4 ± 36.2 | ||
| 3 | −146.7 ± 29.7 | −190.3 ± 35.7 | −158.0 ± 52.0 | −189.0 ± 46.6 | ||
| 5 | −159.7 ± 39.8 | −207.9 ± 46.2 | −166.9 ± 54.1 | −205.4 ± 49.8 | ||
| b-wave | <0.001 | 0.006 | ||||
| 0.1 | 152.8 ± 70.7 | 489.0 ± 153.1 | 241.1 ± 124.8 | 479.2 ± 114.0 | ||
| 1 | 201.2 ± 68.1 | 574.6 ± 153.6 | 286.0 ± 129.2 | 568.7 ± 128.7 | ||
| 3 | 217.8 ± 63.0 | 589.7 ± 158.4 | 294.4 ± 127.0 | 595.9 ± 141.0 | ||
| 5 | 228.2 ± 79.5 | 624.5 ± 162.1 | 299.1 ± 136.3 | 619.3 ± 147.2 | ||
| OPs | <0.0001 | 0.013 | ||||
| 1 | 40.0 ± 8.9 | 157.0 ± 47.8 | 117.2 ± 74.2 | 199.1 ± 33.7 | ||
| 3 | 55.6 ± 9.9 | 217.0 ± 58.7 | 131.8 ± 67.1 | 263.2 ± 54.9 | ||
| 5 | 61.1 ± 11.1 | 207.4 ± 49.9 | 137.6 ± 75.7 | 264.0 ± 44.6 | ||
| Relative Functional Change. Inter-Eye Difference (Mean% ± SD) * | |||||||
|---|---|---|---|---|---|---|---|
| Waveform | Baseline | Day 3 | Day 22 | ||||
| PBS-Treated | ET-1-Treated (p-Value) # | PBS-Treated (p-Value) # | p-Value † | ET-1-Treated (p-Value) # | PBS-Treated (p-Value) # | p-Value † | |
| a-wave | −10.2 ± 2.3 | −37.6 ± 3.9 (0.021) | −11.8 ± 2.2 (ns) | 0.010 | −25.4 ± 3.3 (ns) | −12.8 ± 1.1 (ns) | ns |
| b-wave | −9.9 ± 1.0 | −70.1 ± 0.7 (<0.001) | −11.1 ± 2.7 (ns) | <0.0001 | −58.3 ± 0.6 (<0.001) | −14.1 ± 0.5 (ns) | 0.001 |
| OPs | −12.8 ± 5.6 | −81.0 ± 1.7 (<0.001) | −21.4 ± 5.8 (ns) | <0.0001 | −58.0 ± 1.0 (0.007) | −14.7 ± 1.9 (ns) | 0.001 |
| Relative Functional Change. Inter-Eye Difference (Mean% ± SD) * | ||||||
|---|---|---|---|---|---|---|
| Waveform | AAV-ET-1 Treatment | Vehicle Treatment | ||||
| Day −6 Baseline | Day 3 (p-Value) # | Day 8 (p-Value) # | Day 22 (p-Value) # | Day 50 (p-Value) # | All Days Combined (p-Value) † | |
| a-wave | −4.0 ± 1.3 | −11.1 ± 0.6 (ns) | −15.2 ± 0.5 (0.002) | −10.4 ± 1.4 (ns) | 6.0 ± 2.6 (ns) | −7.9 ± 0.4 (ns) |
| b-wave | −4.6 ± 1.2 | −16.7 ± 2.2 (0.003) | −26.2 ± 0.6 (<0.0001) | −17.9 ± 1.1 (0.001) | −17.3 ± 1.1 (0.010) | −8.8 ± 0.5 (ns) |
| OPs | 3.1 ± 3.6 | −3.6 ± 1.3 (ns) | −20.2 ± 2.6 (<0.001) | −4.0 ± 2.3 (ns) | −11.5 ± 3.1 (ns) | 4.8 ± 3.3 (ns) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nordahl, K.M.L.; Fedulov, V.; Holm, A.; Haanes, K.A. Intraocular Adeno-Associated Virus-Mediated Transgene Endothelin-1 Delivery to the Rat Eye Induces Functional Changes Indicative of Retinal Ischemia—A Potential Chronic Glaucoma Model. Cells 2023, 12, 1987. https://doi.org/10.3390/cells12151987
Nordahl KML, Fedulov V, Holm A, Haanes KA. Intraocular Adeno-Associated Virus-Mediated Transgene Endothelin-1 Delivery to the Rat Eye Induces Functional Changes Indicative of Retinal Ischemia—A Potential Chronic Glaucoma Model. Cells. 2023; 12(15):1987. https://doi.org/10.3390/cells12151987
Chicago/Turabian StyleNordahl, Karin M. L., Vadim Fedulov, Anja Holm, and Kristian A. Haanes. 2023. "Intraocular Adeno-Associated Virus-Mediated Transgene Endothelin-1 Delivery to the Rat Eye Induces Functional Changes Indicative of Retinal Ischemia—A Potential Chronic Glaucoma Model" Cells 12, no. 15: 1987. https://doi.org/10.3390/cells12151987
APA StyleNordahl, K. M. L., Fedulov, V., Holm, A., & Haanes, K. A. (2023). Intraocular Adeno-Associated Virus-Mediated Transgene Endothelin-1 Delivery to the Rat Eye Induces Functional Changes Indicative of Retinal Ischemia—A Potential Chronic Glaucoma Model. Cells, 12(15), 1987. https://doi.org/10.3390/cells12151987