

Kidney-Specific Membrane-Bound Serine Proteases CAP1/Prss8 and CAP3/St14 Affect ENaC Subunit Abundances but Not Its Activity

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Models
2.2. Protein Extraction and Western Blot Analysis
2.3. Metabolic Cage Studies
2.4. RNAscope Analysis
2.5. Quantitative Real-Time PCR
2.6. Diuretic Treatments
2.7. Statistical Analyses
3. Results
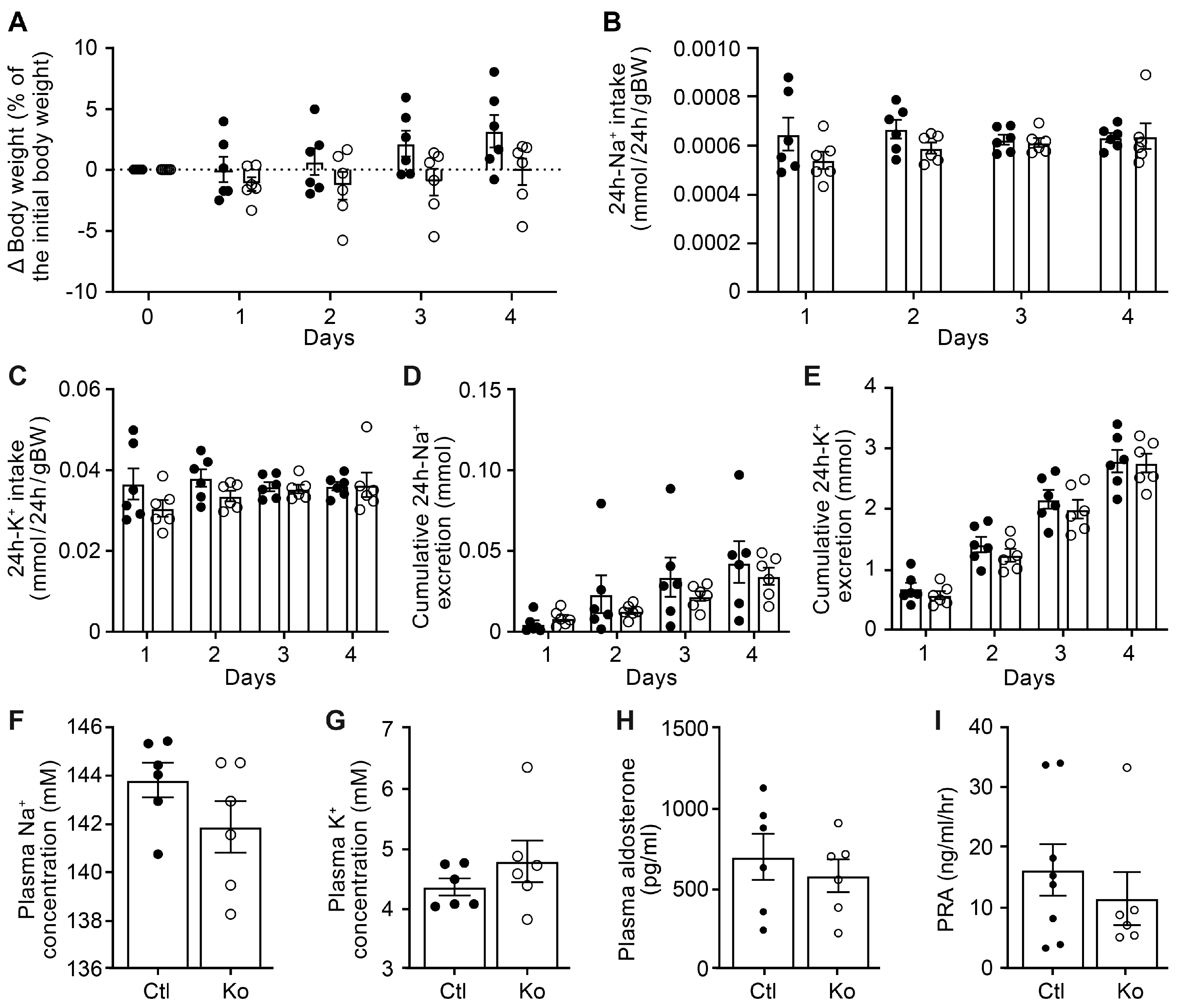
3.1. CAP3/St14 Tubule Specific Deficiency Changed Protein Abundances of ENaC Subunits but Did Not Impair Na+ Homeostasis
3.2. ENaC Is Highly Co-Expressed with CAP3/St14 and Less with CAP1/Prss8 in Distal Tubules
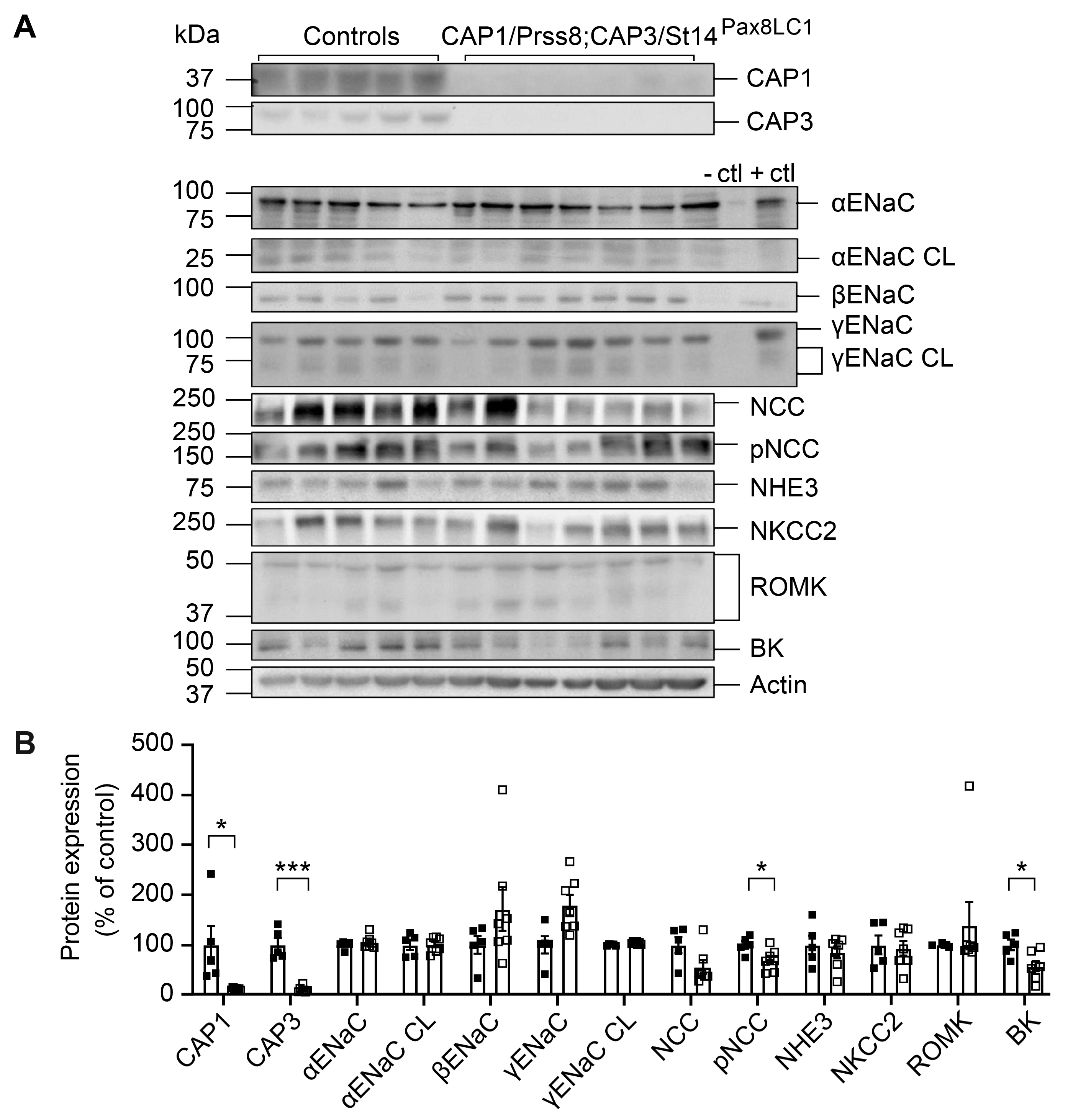
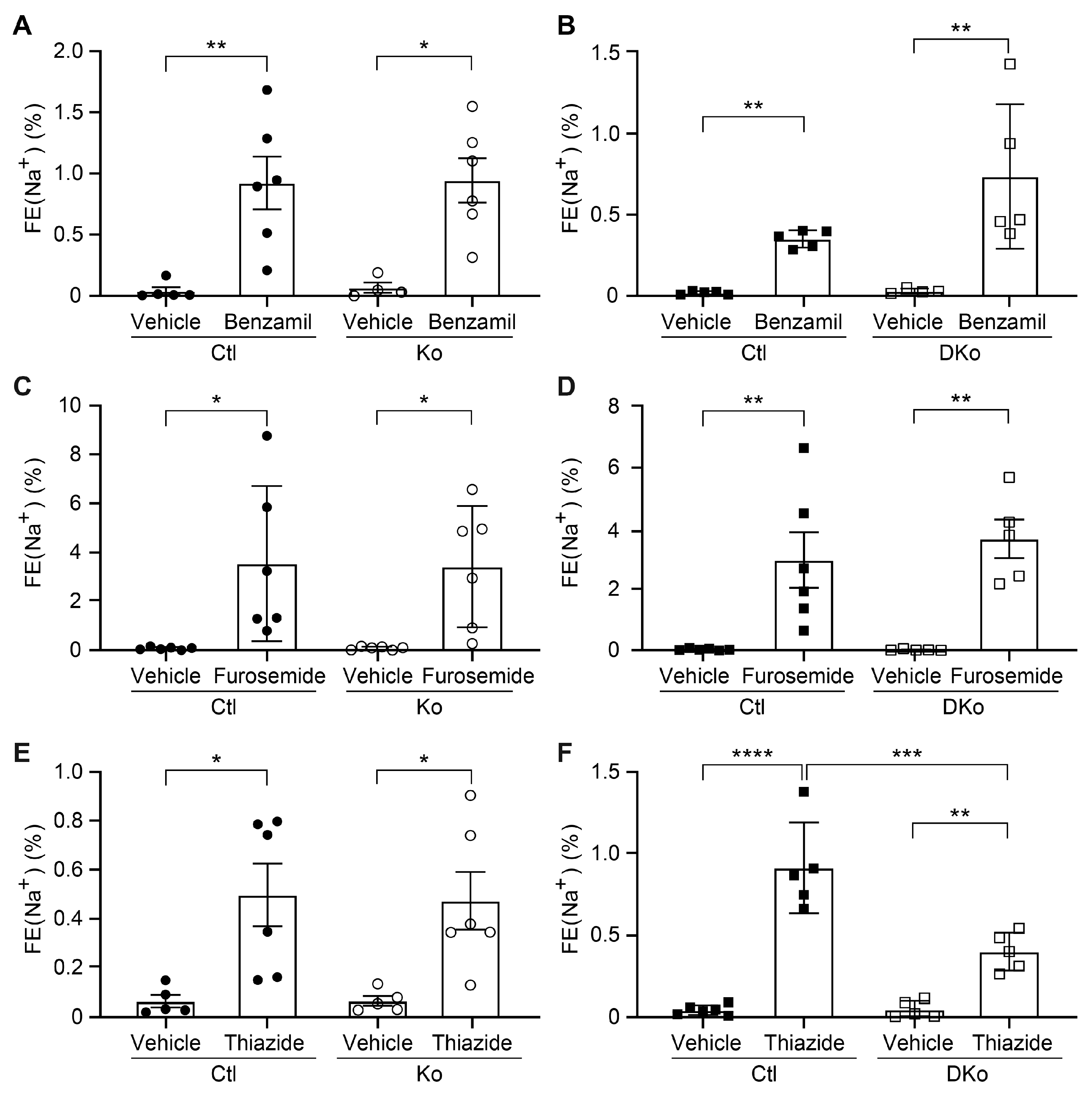
3.3. CAP1/Prss8; CAP3/St14 DKo Mice Restored ENaC Subunit Protein Abundances and Aldosterone Regulation of ENaC but Not of NCC
4. Discussion
4.1. CAP3/St14 Was Not Required for Proteolytic ENaC Activation, Albeit Affecting Its Protein Abundance
4.2. Aldosterone-Dependent ENaC Activation, but Not NCC Activity Was Restored in Na+-Deprived CAP1/Prss8;CAP3/St14 Dko Mice
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BK | Big potassium channel |
| CAP1/Prss8 | Channel-activating protease 1 |
| CAP3/St14 | Channel-activating protease 3 |
| ENaC | Epithelial sodium channel |
| HAI-1 | Hepatocyte growth factor activator inhibitor type 1 |
| HAI-2 | Hepatocyte growth factor activator inhibitor type 2 |
| HE4 | Human epididymis protein-4 |
| NCC | Sodium chloride cotransporter |
| NHE3 | Sodium hydrogen exchanger 3 |
| NKCC2 | Sodium potassium chloride cotransporter |
| PAI-1 | Plasminogen activator inhibitor-1 |
| ROMK | Renal outer medullary potassium channel |
References
- Santulli, G.; Ciccarelli, M.; Trimarco, B.; Iaccarino, G. Physical activity ameliorates cardiovascular health in elderly subjects: The functional role of the β adrenergic system. Front. Physiol. 2013, 4, 209. [Google Scholar] [CrossRef]
- Weinberger, M.H. Salt Sensitivity of Blood Pressure in Humans. Hypertension 1996, 27, 481–490. [Google Scholar] [CrossRef]
- Pratt, J.H. Central Role for ENaC in Development of Hypertension. J. Am. Soc. Nephrol. 2005, 16, 3154–3159. [Google Scholar] [CrossRef]
- Vallet, V.; Chraibi, A.; Gaeggeler, H.-P.; Horisberger, J.-D.; Rossier, B.C. An epithelial serine protease activates the amiloride-sensitive sodium channel. Nature 1997, 389, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, D.; Vuagniaux, G.A.A.; Fowler-Jaeger, N.; Hummler, E.; Rossier, B.C. Activation of Epithelial Sodium Channels by Mouse Channel Activating Proteases (mCAP) Expressed in Xenopus Oocytes Requires Catalytic Activity of mCAP3 and mCAP2 but not mCAP1. J. Am. Soc. Nephrol. 2006, 17, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Vuagniaux, G.; Vallet, V.; Jaeger, N.F.; Hummler, E.; Rossier, B.C. Synergistic Activation of ENaC by Three Membrane-bound Channel-activating Serine Proteases (mCAP1, mCAP2, and mCAP3) and Serum- and Glucocorticoid-regulated Kinase (Sgk1) in Xenopus Oocytes. J. Gen. Physiol. 2002, 120, 191–201. [Google Scholar] [CrossRef]
- Anand, D.; Hummler, E.; Rickman, O.J. ENaC activation by proteases. Acta Physiol. 2022, 235, e13811. [Google Scholar] [CrossRef]
- Kleyman, T.R.; Eaton, D.C. Regulating ENaC’s gate. Am. J. Physiol. Physiol. 2020, 318, C150–C162. [Google Scholar] [CrossRef]
- Picard, N.; Eladari, D.; El Moghrabi, S.; Planès, C.; Bourgeois, S.; Houillier, P.; Wang, Q.; Burnier, M.; Deschenes, G.; Knepper, M.A.; et al. Defective ENaC Processing and Function in Tissue Kallikrein-deficient Mice. J. Biol. Chem. 2008, 283, 4602–4611. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhao, R.; Zhao, M.; Liang, X.; Bhattarai, D.; Dhiman, R.; Shetty, S.; Idell, S.; Ji, H.-L.; Bartoszewski, R.; et al. Regulation of epithelial sodium channels in urokinase plasminogen activator deficiency. Am. J. Physiol. Cell. Mol. Physiol. 2014, 307, L609–L617. [Google Scholar] [CrossRef] [PubMed]
- Keppner, A.; Maric, D.; Sergi, C.; Ansermet, C.; De Bellis, D.; Kratschmar, D.V.; Canonica, J.; Klusonova, P.; Fenton, R.A.; Odermatt, A.; et al. Deletion of the serine protease CAP2/Tmprss4 leads to dysregulated renal water handling upon dietary potassium depletion. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Lai, C.-H.; Lai, Y.-J.J.; Chou, F.-P.; Chang, H.-H.D.; Tseng, C.-C.; Johnson, M.D.; Wang, J.-K.; Lin, C.-Y. Matriptase Complexes and Prostasin Complexes with HAI-1 and HAI-2 in Human Milk: Significant Proteolysis in Lactation. PLOS ONE 2016, 11, e0152904. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Chao, J.; Chao, L. Adenovirus-mediated human prostasin gene delivery is linked to increased aldosterone production and hypertension in rats. Am. J. Physiol. Integr. Comp. Physiol. 2003, 284, R1031–R1036. [Google Scholar] [CrossRef]
- Zhu, H.; Guo, D.; Li, K.; Yan, W.; Tan, Y.; Wang, X.; Treiber, F.A.; Chao, J.; Snieder, H.; Dong, Y. Prostasin: A Possible Candidate Gene for Human Hypertension. Am. J. Hypertens. 2008, 21, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, O.; Castagna, A.; Guarini, P.; Chiecchi, L.; Sabaini, G.; Pizzolo, F.; Corrocher, R.; Righetti, P.G. Urinary Prostasin. Hypertension 2005, 46, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Essigke, D.; Bohnert, B.N.; Janessa, A.; Wörn, M.; Omage, K.; Kalbacher, H.; Birkenfeld, A.L.; Bugge, T.H.; Szabo, R.; Artunc, F. Sodium retention in nephrotic syndrome is independent of the activation of the membrane-anchored serine protease prostasin (CAP1/PRSS8) and its enzymatic activity. Pflügers Arch.-Eur. J. Physiol. 2022, 474, 613–624. [Google Scholar] [CrossRef]
- Ehret, E.; Jäger, Y.; Sergi, C.; Mérillat, A.-M.; Peyrollaz, T.; Anand, D.; Wang, Q.; Ino, F.; Maillard, M.; Kellenberger, S.; et al. Kidney-Specific CAP1/Prss8-Deficient Mice Maintain ENaC-Mediated Sodium Balance through an Aldosterone Independent Pathway. Int. J. Mol. Sci. 2022, 23, 6745. [Google Scholar] [CrossRef]
- Friis, S.; Sales, K.U.; Godiksen, S.; Peters, D.E.; Lin, C.-Y.; Vogel, L.K.; Bugge, T.H. A Matriptase-Prostasin Reciprocal Zymogen Activation Complex with Unique Features. J. Biol. Chem. 2013, 288, 19028–19039. [Google Scholar] [CrossRef]
- Holt-Danborg, L.; Skovbjerg, S.; Goderum, K.W.; Nonboe, A.W.; Stankevic, E.; Frost, K.; Vitved, L.; Jensen, J.K.; Vogel, L.K. Insights into the regulation of the matriptase-prostasin proteolytic system. Biochem. J. 2020, 477, 4349–4365. [Google Scholar] [CrossRef]
- Su, H.C.; Liang, Y.A.; Lai, Y.-J.J.; Chiu, Y.-L.; Barndt, R.B.; Shiao, F.; Chang, H.-H.D.; Lu, D.D.; Huang, N.; Tseng, C.-C.; et al. Natural Endogenous Human Matriptase and Prostasin Undergo Zymogen Activation via Independent Mechanisms in an Uncoupled Manner. PLoS ONE 2016, 11, e0167894. [Google Scholar] [CrossRef]
- Bohnert, B.N.; Menacher, M.; Janessa, A.; Wörn, M.; Schork, A.; Daiminger, S.; Kalbacher, H.; Häring, H.-U.; Daniel, C.; Amann, K.; et al. Aprotinin prevents proteolytic epithelial sodium channel (ENaC) activation and volume retention in nephrotic syndrome. Kidney Int. 2017, 93, 159–172. [Google Scholar] [CrossRef] [PubMed]
- List, K.; Kosa, P.; Szabo, R.; Bey, A.L.; Wang, C.B.; Molinolo, A.; Bugge, T.H. Epithelial Integrity Is Maintained by a Matriptase-Dependent Proteolytic Pathway. Am. J. Pathol. 2009, 175, 1453–1463. [Google Scholar] [CrossRef] [PubMed]
- Traykova-Brauch, M.; Schönig, K.; Greiner, O.; Miloud, T.; Jauch, A.; Bode, M.; Felsher, D.W.; Glick, A.B.; Kwiatkowski, D.J.; Bujard, H.; et al. An efficient and versatile system for acute and chronic modulation of renal tubular function in transgenic mice. Nat. Med. 2008, 14, 979–984. [Google Scholar] [CrossRef]
- Sorensen, M.V.; Grossmann, S.; Roesinger, M.; Gresko, N.; Todkar, A.P.; Barmettler, G.; Ziegler, U.; Odermatt, A.; Loffing-Cueni, D.; Loffing, J. Rapid dephosphorylation of the renal sodium chloride cotransporter in response to oral potassium intake in mice. Kidney Int. 2013, 83, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.R.; Plotkin, M.D.; Lee, W.-S.; Xu, Z.-C.; Lytton, J.; Hebert, S.C. Apical localization of the Na-K-Cl cotransporter, rBSC1, on rat thick ascending limbs. Kidney Int. 1996, 49, 40–47. [Google Scholar] [CrossRef]
- Wu, P.; Gao, Z.; Zhang, D.; Duan, X.; Terker, A.S.; Lin, D.; Ellison, D.H.; Wang, W. Effect of Angiotensin II on ENaC in the Distal Convoluted Tubule and in the Cortical Collecting Duct of Mineralocorticoid Receptor Deficient Mice. J. Am. Hear. Assoc. 2020, 9, e014996. [Google Scholar] [CrossRef]
- Sassi, A.; Wang, Y.; Chassot, A.; Komarynets, O.; Roth, I.; Olivier, V.; Crambert, G.; Dizin, E.; Boscardin, E.; Hummler, E.; et al. Interaction between Epithelial Sodium Channel γ-Subunit and Claudin-8 Modulates Paracellular Sodium Permeability in Renal Collecting Duct. J. Am. Soc. Nephrol. 2020, 31, 1009–1023. [Google Scholar] [CrossRef]
- Kota, P.; García-Caballero, A.; Dang, H.; Gentzsch, M.; Stutts, M.J.; Dokholyan, N.V. Energetic and Structural Basis for Activation of the Epithelial Sodium Channel by Matriptase. Biochemistry 2012, 51, 3460–3469. [Google Scholar] [CrossRef]
- Verouti, S.N.; Boscardin, E.; Hummler, E.; Frateschi, S. Regulation of blood pressure and renal function by NCC and ENaC: Lessons from genetically engineered mice. Curr. Opin. Pharmacol. 2015, 21, 60–72. [Google Scholar] [CrossRef]
- Higashi, T.; Saito, A.C.; Fukazawa, Y.; Furuse, M.; Higashi, A.Y.; Ono, M.; Chiba, H. EpCAM proteolysis and release of complexed claudin-7 repair and maintain the tight junction barrier. J. Cell Biol. 2022, 222. [Google Scholar] [CrossRef]
- Frateschi, S.; Camerer, E.; Crisante, G.; Rieser, S.; Membrez, M.; Charles, R.-P.; Beermann, F.; Stehle, J.-C.; Breiden, B.; Sandhoff, K.; et al. PAR2 absence completely rescues inflammation and ichthyosis caused by altered CAP1/Prss8 expression in mouse skin. Nat. Commun. 2011, 2, 161. [Google Scholar] [CrossRef] [PubMed]
- Szabo, R.; Sales, K.U.; Kosa, P.; Shylo, N.A.; Godiksen, S.; Hansen, K.K.; Friis, S.; Gutkind, J.S.; Vogel, L.K.; Hummler, E.; et al. Reduced Prostasin (CAP1/PRSS8) Activity Eliminates HAI-1 and HAI-2 Deficiency–Associated Developmental Defects by Preventing Matriptase Activation. PLOS Genet. 2012, 8, e1002937. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, M.; Yamamoto, K.; Takeda, N.; Fukushima, T.; Yamashita, F.; Sato, K.; Kitamura, K.; Hippo, Y.; Janetka, J.W.; Kataoka, H. Hepatocyte growth factor activator inhibitor-2 stabilizes Epcam and maintains epithelial organization in the mouse intestine. Commun. Biol. 2019, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Szabo, R.; Callies, L.K.; Bugge, T.H. Matriptase drives early-onset intestinal failure in a mouse model of congenital tufting enteropathy. Development 2019, 146. [Google Scholar] [CrossRef] [PubMed]
- Breiderhoff, T.; Himmerkus, N.; Drewell, H.; Plain, A.; Günzel, D.; Mutig, K.; Willnow, T.E.; Müller, D.; Bleich, M. Deletion of claudin-10 rescues claudin-16–deficient mice from hypomagnesemia and hypercalciuria. Kidney Int. 2018, 93, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Oakley, R.H.; Cruz-Topete, D.; He, B.; Foley, J.F.; Myers, P.H.; Xu, X.; Gomez-Sanchez, C.E.; Chambon, P.; Willis, M.S.; Cidlowski, J.A. Cardiomyocyte glucocorticoid and mineralocorticoid receptors directly and antagonistically regulate heart disease in mice. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ehret, E.; Stroh, S.; Auberson, M.; Ino, F.; Jäger, Y.; Maillard, M.; Szabo, R.; Bugge, T.H.; Frateschi, S.; Hummler, E. Kidney-Specific Membrane-Bound Serine Proteases CAP1/Prss8 and CAP3/St14 Affect ENaC Subunit Abundances but Not Its Activity. Cells 2023, 12, 2342. https://doi.org/10.3390/cells12192342
Ehret E, Stroh S, Auberson M, Ino F, Jäger Y, Maillard M, Szabo R, Bugge TH, Frateschi S, Hummler E. Kidney-Specific Membrane-Bound Serine Proteases CAP1/Prss8 and CAP3/St14 Affect ENaC Subunit Abundances but Not Its Activity. Cells. 2023; 12(19):2342. https://doi.org/10.3390/cells12192342
Chicago/Turabian StyleEhret, Elodie, Sévan Stroh, Muriel Auberson, Frédérique Ino, Yannick Jäger, Marc Maillard, Roman Szabo, Thomas H. Bugge, Simona Frateschi, and Edith Hummler. 2023. "Kidney-Specific Membrane-Bound Serine Proteases CAP1/Prss8 and CAP3/St14 Affect ENaC Subunit Abundances but Not Its Activity" Cells 12, no. 19: 2342. https://doi.org/10.3390/cells12192342
APA StyleEhret, E., Stroh, S., Auberson, M., Ino, F., Jäger, Y., Maillard, M., Szabo, R., Bugge, T. H., Frateschi, S., & Hummler, E. (2023). Kidney-Specific Membrane-Bound Serine Proteases CAP1/Prss8 and CAP3/St14 Affect ENaC Subunit Abundances but Not Its Activity. Cells, 12(19), 2342. https://doi.org/10.3390/cells12192342