
Imidacloprid Induces Lysosomal Dysfunction and Cell Death in Human Astrocytes and Fibroblasts—Environmental Implication of a Clinical Case Report

 ,
,  , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Case Description
2.2. Cell Cultures and Experimental Conditions
2.3. Cell Number and DNA Content
2.4. Apoptosis and Necrosis
2.5. Reactive Oxygen Species (ROS)
2.6. Lysosomal Membrane Permeabilisation (LMP)
2.7. Immunocytochemistry of D384 Astrocytes
2.8. Immunocytochemistry of AG01518 Fibroblasts
2.9. Western Blot
2.10. Statistics
3. Results
3.1. Health Problems of the Patient
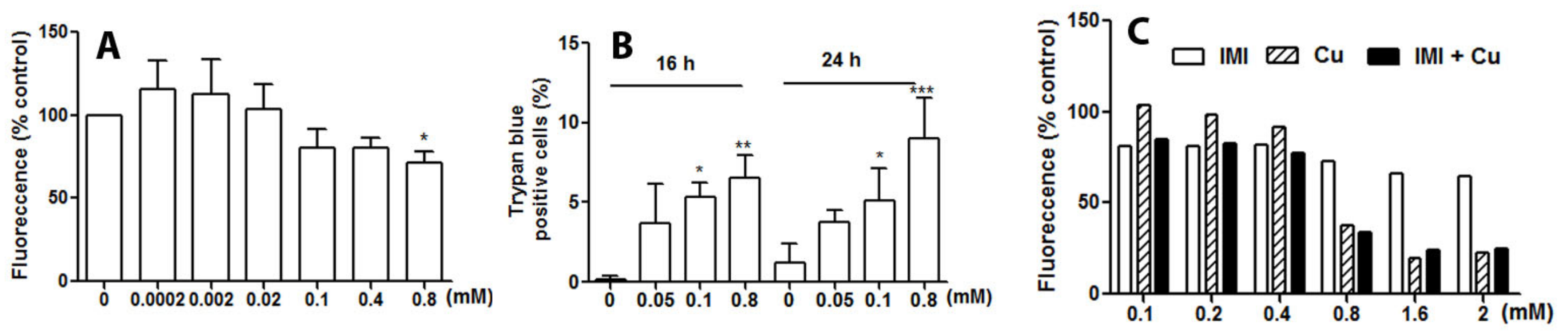
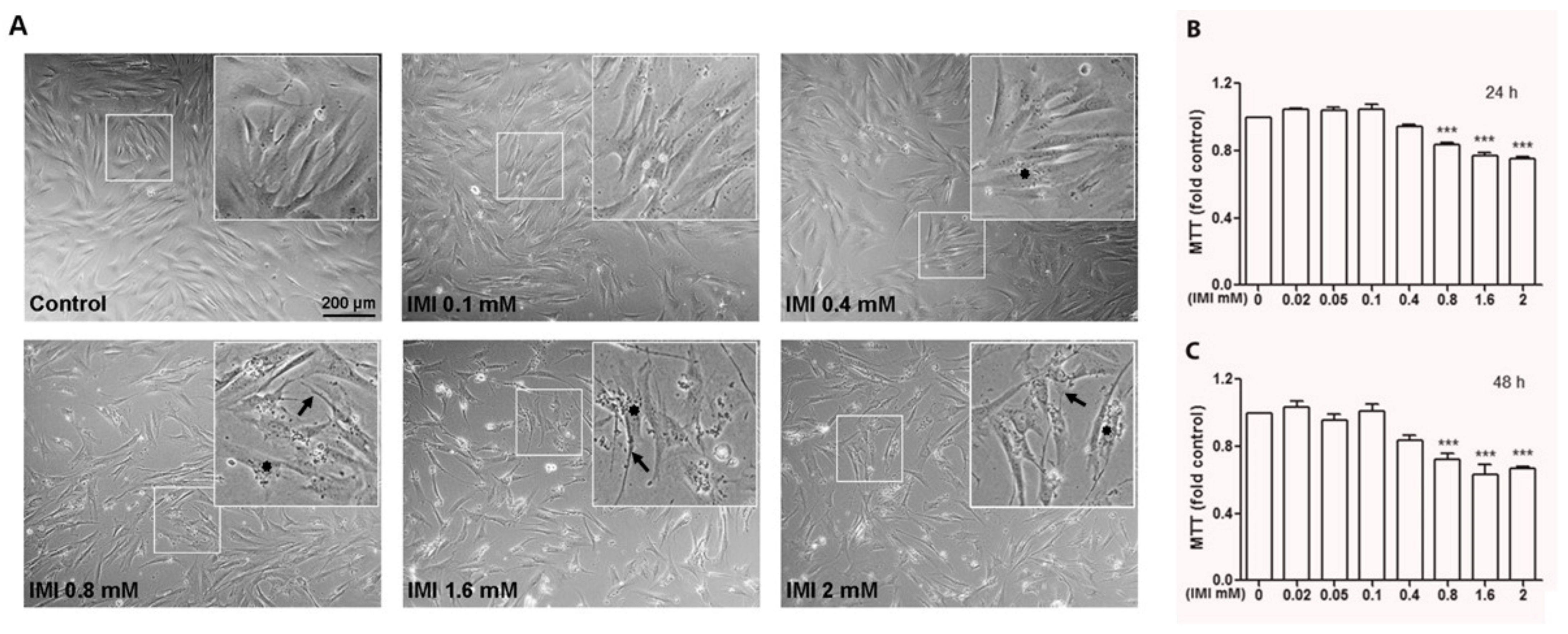
3.2. IMI Exposure Decreases Cell Proliferation and Induces Cell Death in Human D384 Astrocyte and Human Fibroblast Cells
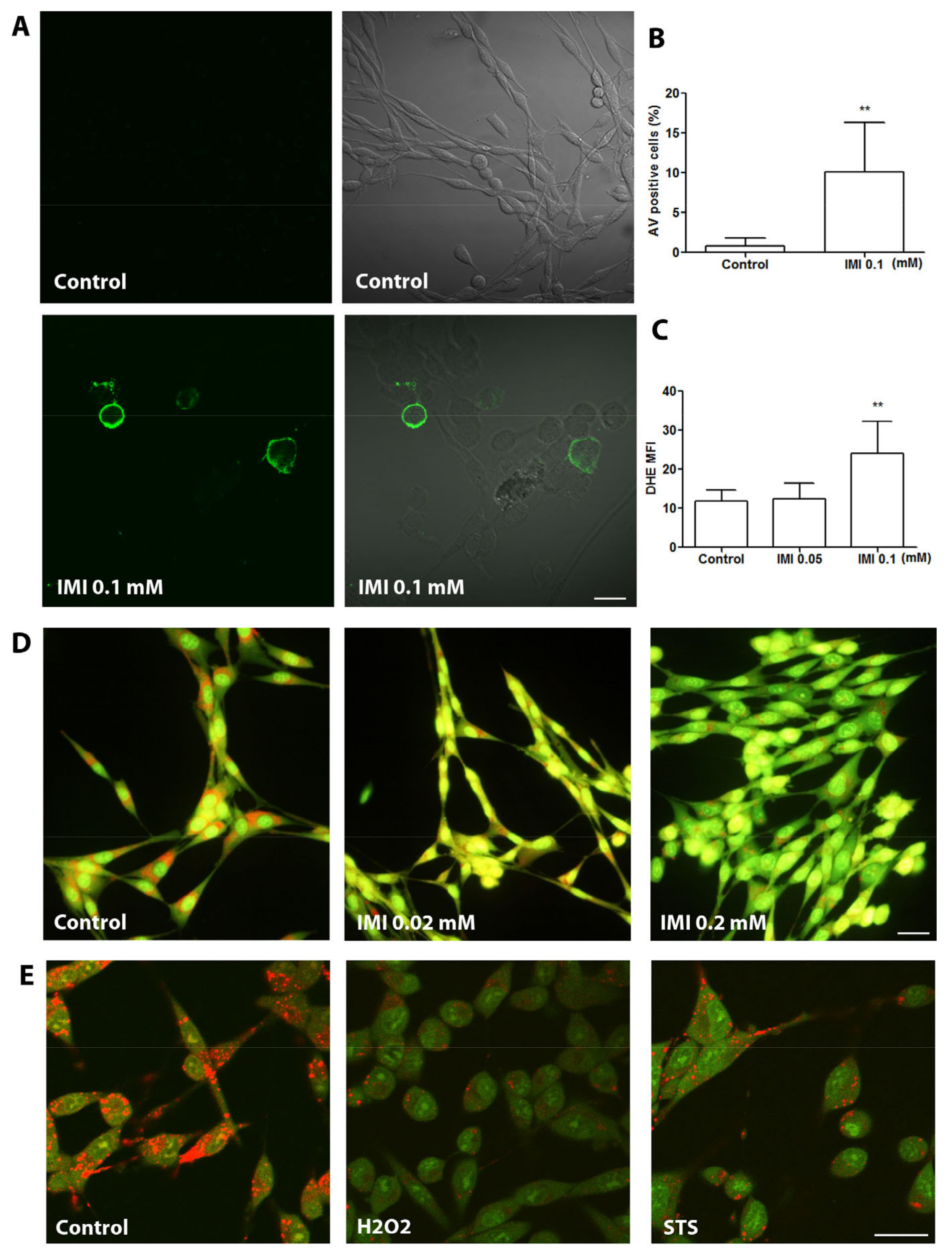
3.3. IMI Exposure Induces ROS Production, Apoptosis, and LMP in Human D384 Astrocytes
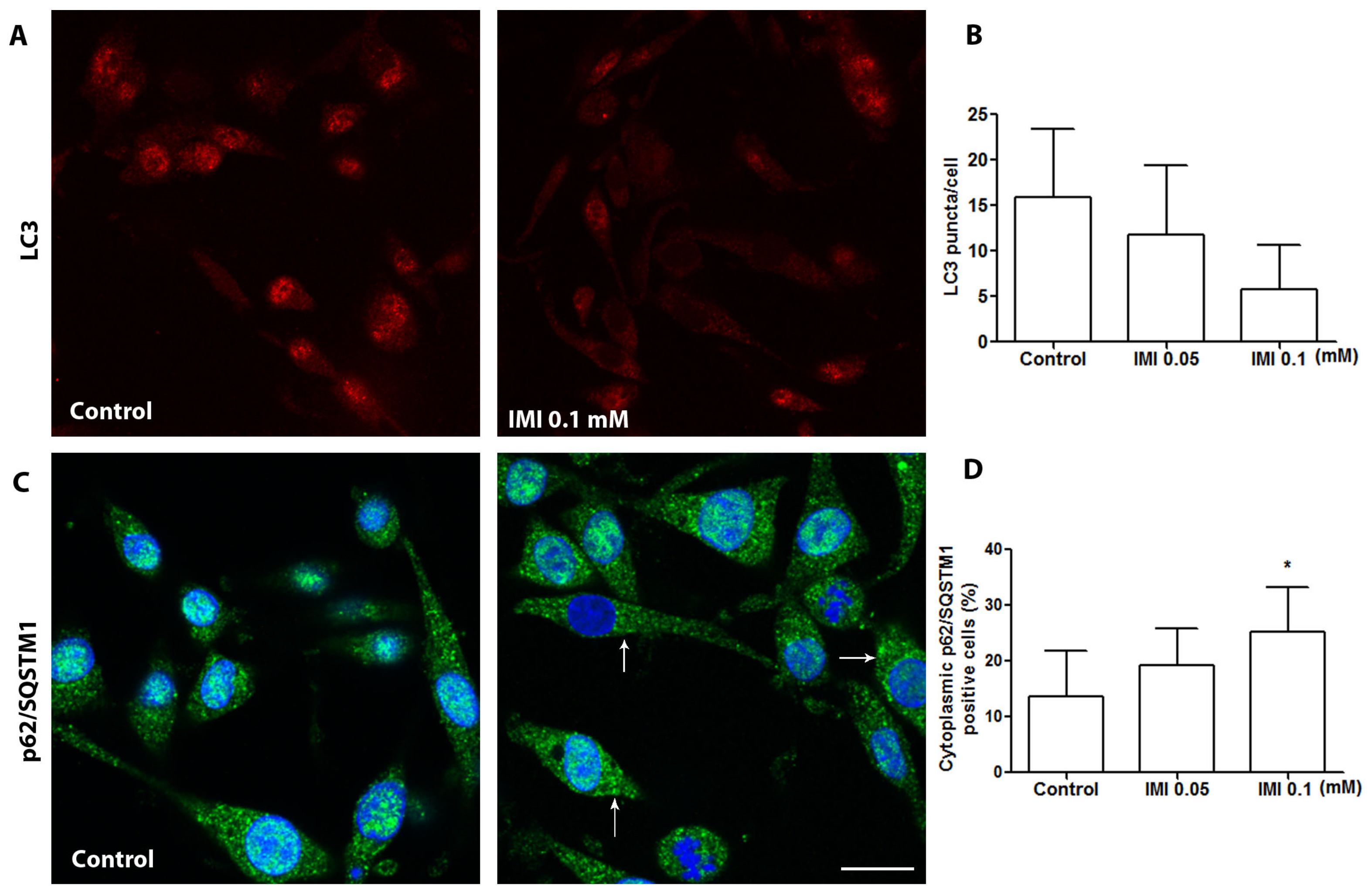
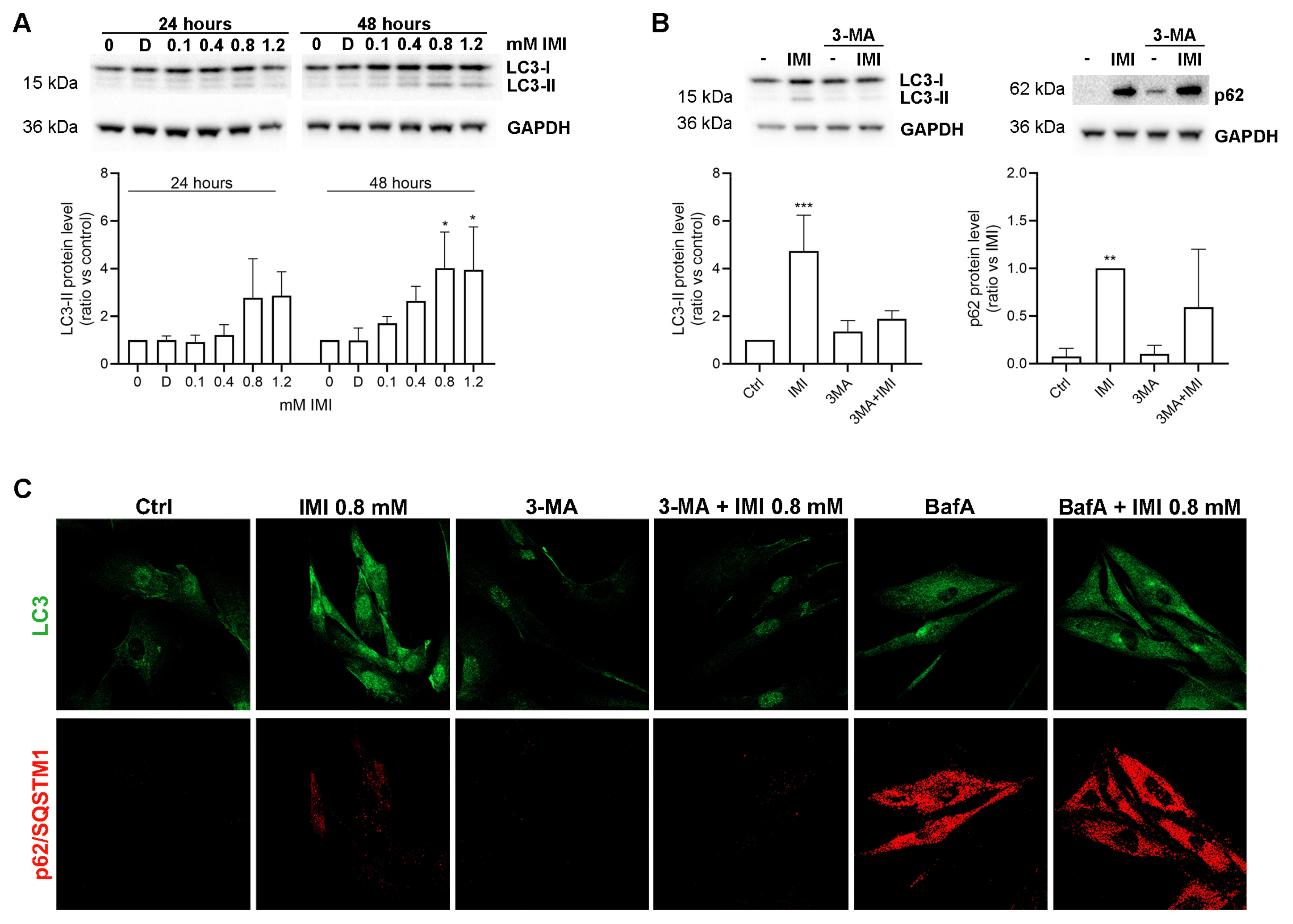
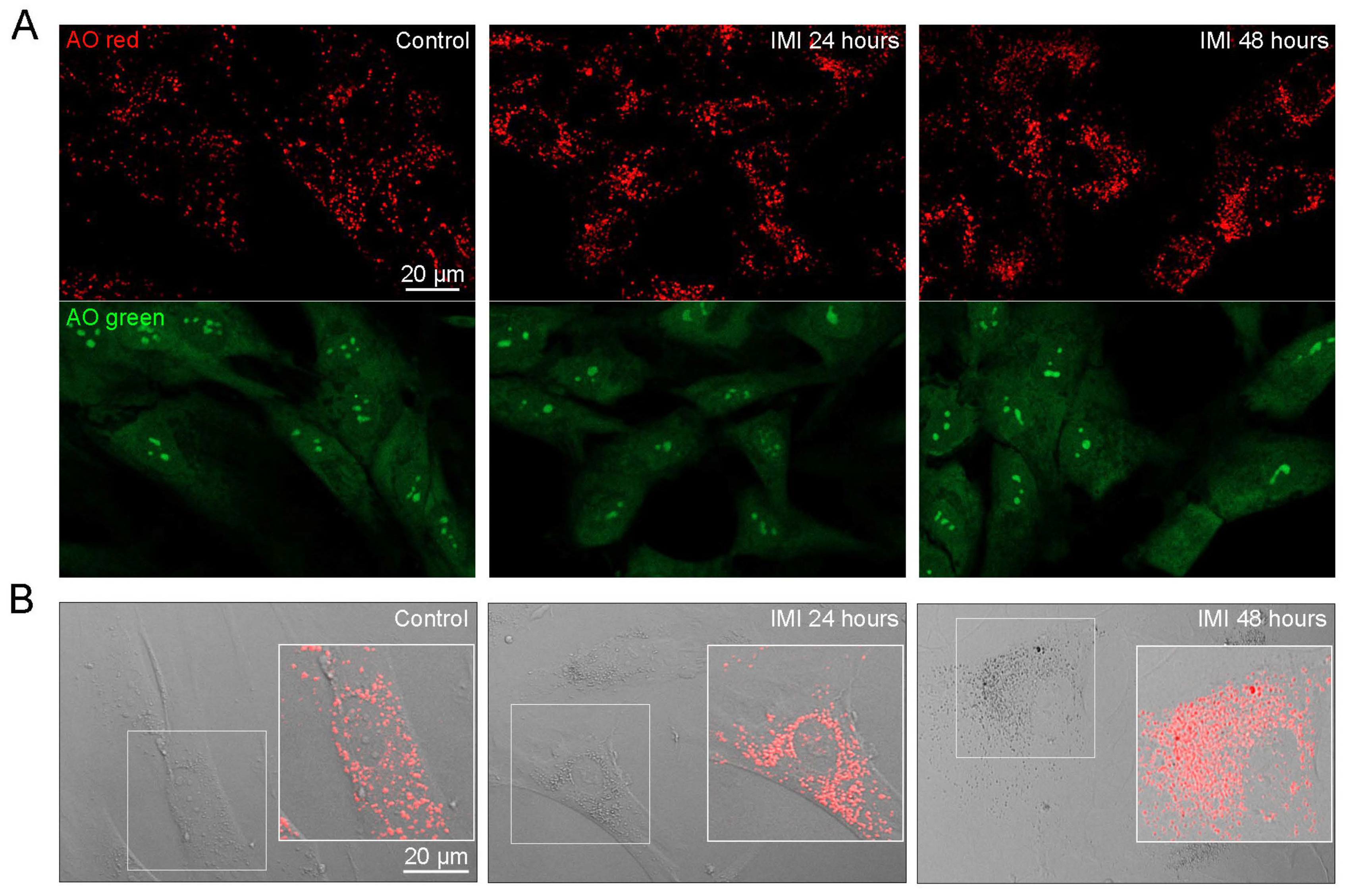
3.4. IMI Exposure Induces Autophagy Dysfunction in Human D384 Astrocytes
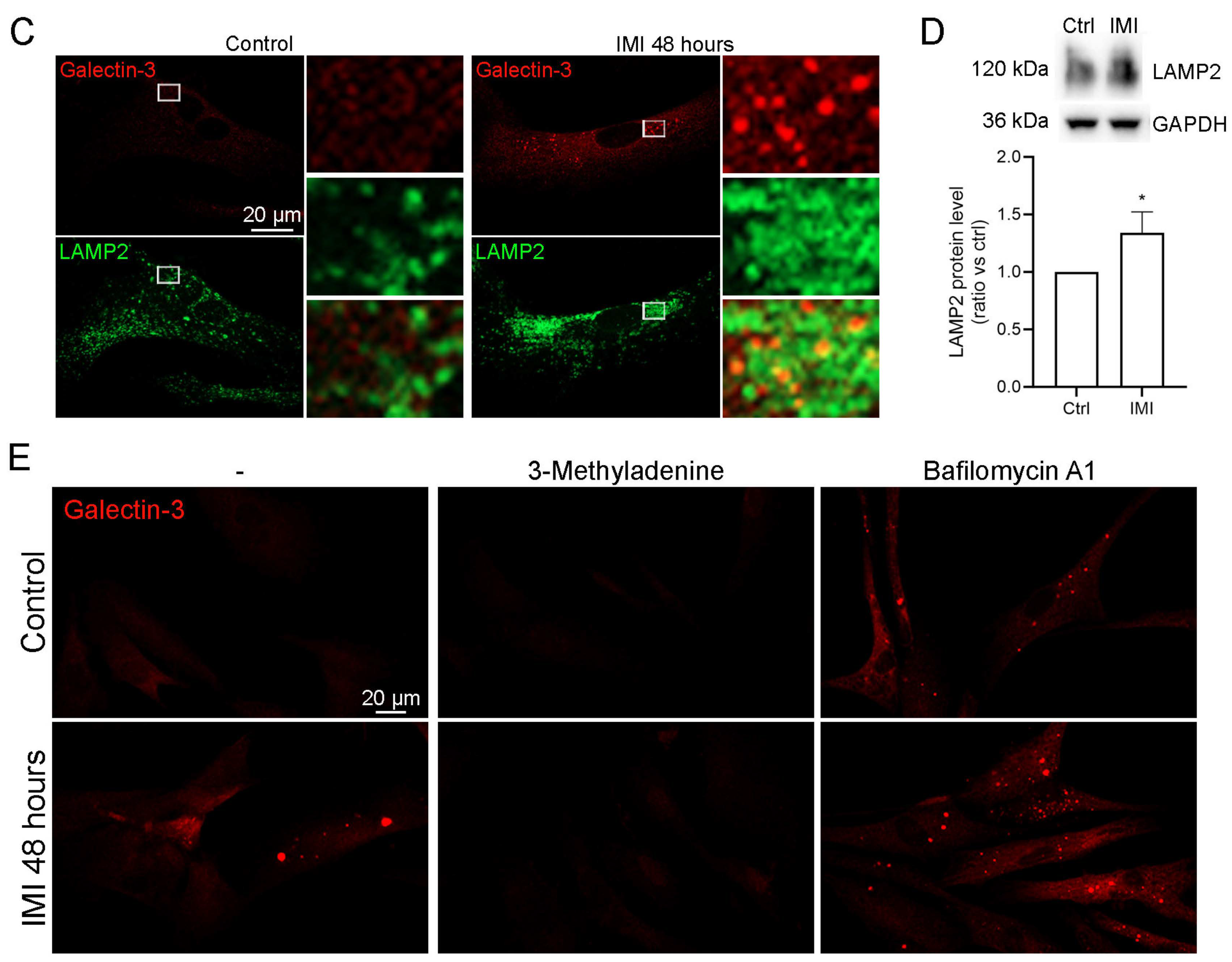
3.5. IMI-Induced Autophagy Dysfunction Is Inhibited by 3-Methyladenine but Is Not Affected by Bafilomycin A1 in Human Fibroblast Cells
3.6. IMI Accumulates in Lysosomes and Reduces Lysosomal Function, Causing Lysophagy and a Compensatory Upregulation of Lysosomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hladik, M.L.; Main, A.R.; Goulson, D. Environmental Risks and Challenges Associated with Neonicotinoid Insecticides. Environ. Sci. Technol. 2018, 52, 3329–3335. [Google Scholar] [CrossRef]
- Pang, S.; Lin, Z.; Zhang, Y.; Zhang, W.; Alansary, N.; Mishra, S.; Bhatt, P.; Chen, S. Insights into the Toxicity and Degradation Mechanisms of Imidacloprid Via Physicochemical and Microbial Approaches. Toxics 2020, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Herbertsson, L.; Jonsson, O.; Kreuger, J.; Smith, H.G.; Rundlöf, M. Scientific Note: Imidacloprid Found in Wild Plants Downstream Permanent Greenhouses in Sweden. Apidologie 2021, 52, 946–949. [Google Scholar] [CrossRef]
- Boye, K.; Boström, G.; Jonsson, O.; Gönczi, M.; Löfkvist, K.; Kreuger, J. Greenhouse Production Contributes to Pesticide Occurrences in Swedish Streams. Sci. Total Environ. 2022, 809, 152215. [Google Scholar] [CrossRef] [PubMed]
- Boye, K.; Lindström, B.; Boström, G.; Kreuger, J. Long-term Data from the Swedish National Environmental Monitoring Program of Pesticides in Surface Waters. J. Environ. Qual. 2019, 48, 1109–1119. [Google Scholar] [CrossRef] [PubMed]
- Distefano, G.G.; Zangrando, R.; Basso, M.; Panzarin, L.; Gambaro, A.; Volpi Ghirardini, A.; Picone, M. The Ubiquity of Neonicotinoid Contamination: Residues in Seabirds with Different Trophic Habits. Environ. Res. 2022, 206, 112637. [Google Scholar] [CrossRef]
- Costa, C.; Silvari, V.; Melchini, A.; Catania, S.; Heffron, J.J.; Trovato, A.; de Pasquale, R. Genotoxicity of Imidacloprid in Relation to Metabolic Activation and Composition of the Commercial Product. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2009, 672, 40–44. [Google Scholar] [CrossRef]
- Feng, S.; Kong, Z.; Wang, X.; Peng, P.; Zeng, E.Y. Assessing the Genotoxicity of Imidacloprid and RH-5849 in Human Peripheral Blood Lymphocytes in vitro with Comet Assay and Cytogenetic Tests. Ecotoxicol. Environ. Saf. 2005, 61, 239–246. [Google Scholar] [CrossRef]
- Abdel-Halim, K.Y.; Osman, S.R. Cytotoxicity and Oxidative Stress Responses of Imidacloprid and Glyphosate in Human Prostate Epithelial WPM-Y.1 Cell Line. J. Toxicol. 2020, 2020, 4364650. [Google Scholar] [CrossRef]
- Skandrani, D.; Gaubin, Y.; Beau, B.; Murat, J.C.; Vincent, C.; Croute, F. Effect of Selected Insecticides on Growth Rate and Stress Protein Expression in Cultured Human A549 and SH-SY5Y Cells. Toxicol. Vitr. 2006, 20, 1378–1386. [Google Scholar] [CrossRef]
- Şenyildiz, M.; Kilinc, A.; Ozden, S. Investigation of the Genotoxic and Cytotoxic Effects of Widely Used Neonicotinoid Insecticides in HepG2 and SH-SY5Y Cells. Toxicol. Ind. Health 2018, 34, 375–383. [Google Scholar] [CrossRef]
- Loser, D.; Hinojosa, M.G.; Blum, J.; Schaefer, J.; Brüll, M.; Johansson, Y.; Suciu, I.; Grillberger, K.; Danker, T.; Möller, C.; et al. Functional Alterations by a Subgroup of Neonicotinoid Pesticides in Human Dopaminergic Neurons. Arch. Toxikol. 2021, 95, 2081–2107. [Google Scholar] [CrossRef]
- Nakayama, A.; Yoshida, M.; Kagawa, N.; Nagao, T. The Neonicotinoids Acetamiprid and Imidacloprid Impair Neurogenesis and Alter the Microglial Profile in the Hippocampal Dentate Gyrus of Mouse Neonates. J. Appl. Toxicol. 2019, 39, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Reinwald, H.; Alvincz, J.; Salinas, G.; Schäfers, C.; Hollert, H.; Eilebrecht, S. Toxicogenomic Profiling after Sublethal Exposure to Nerve- and Muscle-Targeting Insecticides Reveals Cardiac and Neuronal Developmental Effects in Zebrafish Embryos. Chemosphere 2022, 291, 132746. [Google Scholar] [CrossRef]
- Janner, D.E.; Gomes, N.S.; Poetini, M.R.; Poleto, K.H.; Musachio, E.A.S.; de Almeida, F.P.; de Matos Amador, E.C.; Reginaldo, J.C.; Ramborger, B.P.; Roehrs, R.; et al. Oxidative stress and decreased dopamine levels induced by imidacloprid exposure cause behavioral changes in a neurodevelopmental disorder model in Drosophila melanogaster. Neurotoxicology 2021, 85, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Baysal, M.; Atlı-Eklioğlu, Ö. Comparison of the Toxicity of Pure Compounds and Commercial Formulations of Imidacloprid and Acetamiprid on HT-29 Cells: Single and Mixture Exposure. Food Chem. Toxicol. 2021, 155, 112430. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.M.; Martins-Gomes, C.; Ferreira, S.S.; Souto, E.B.; Andreani, T. Molecular Physicochemical Properties of Selected Pesticides as Predictive Factors for Oxidative Stress and Apoptosis-Dependent Cell Death in Caco-2 and HepG2 Cells. Int. J. Mol. Sci. 2022, 23, 8107. [Google Scholar] [CrossRef]
- Quent, V.M.C.; Loessner, D.; Friis, T.; Reichert, J.C.; Hutmacher, D.W. Discrepancies between Metabolic Activity and DNA Content as Tool to Assess Cell Proliferation in Cancer Research. J. Cell Mol. Med. 2010, 14, 1003–1013. [Google Scholar] [CrossRef]
- Yuan, X.M.; Li, W.; Olsson, A.G.; Brunk, U.T. The Toxicity to Macrophages of Oxidized Low-Density Lipoprotein Is Mediated through Lysosomal Damage. Atherosclerosis 1997, 133, 153–161. [Google Scholar] [CrossRef]
- Ladoire, S.; Chaba, K.; Martins, I.; Sukkurwala, A.Q.; Adjemian, S.; Michaud, M.; Poirier-Colame, V.; Andreiuolo, F.; Galluzzi, L.; White, E.; et al. Immunohistochemical Detection of Cytoplasmic LC3 Puncta in Human Cancer Specimens. Autophagy 2012, 8, 1175. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 Years of Image Analysis. Nat. Methods 2012, 9, 671. [Google Scholar] [CrossRef]
- Eriksson, I.; Wäster, P.; Öllinger, K. Restoration of lysosomal function after damage is accompanied by recycling of lysosomal membrane proteins. Cell Death Dis. 2020, 11, 370. [Google Scholar] [CrossRef]
- Abd-Elhakim, Y.M.; Mohammed, H.H.; Mohamed, W.A.M. Imidacloprid Impacts on Neurobehavioral Performance, Oxidative Stress, and Apoptotic Events in the Brain of Adolescent and Adult Rats. J. Agric. Food Chem. 2018, 66, 13513–13524. [Google Scholar] [CrossRef]
- Carneiro, L.S.; Martinez, L.C.; de Oliveira, A.H.; Cossolin, J.F.S.; de Resende, M.T.C.S.; Gonçalves, W.G.; Medeiros-Santana, L.; Serrão, J.E. Acute Oral Exposure to Imidacloprid Induces Apoptosis and Autophagy in the Midgut of Honey Bee Apis Mellifera Workers. Sci. Total Environ. 2022, 815, 152847. [Google Scholar] [CrossRef]
- Wu, Y.Y.; Zhou, T.; Wang, Q.; Dai, P.L.; Xu, S.F.; Jia, H.R.; Wang, X. Programmed Cell Death in the Honey Bee (Apis mellifera) (Hymenoptera: Apidae) Worker Brain Induced by Imidacloprid. J. Econ. Entomol. 2015, 108, 1486–1494. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yao, Y.; Wang, J.; Shen, Z.; Jiang, Z.; Xu, S. Eucalyptol Relieves Imidacloprid-Induced Autophagy through the MiR-451/Cab39/AMPK Axis in Ctenopharyngodon Idellus Kidney Cells. Aquat. Toxicol. 2022, 249, 106204. [Google Scholar] [CrossRef] [PubMed]
- Seglen, P.O.; Gordon, P.B. 3-Methyladenine: Specific inhibitor of autophagic/lysosomal protein degradation in isolated rat hepatocytes. Proc. Natl. Acad. Sci. USA 1982, 79, 1889–1892. [Google Scholar] [CrossRef] [PubMed]
- Mauvezin, C.; Neufeld, T.P. Bafilomycin A1 disrupts autophagic flux by inhibiting both V-ATPase-dependent acidification and Ca-P60A/SERCA-dependent autophagosome-lysosome fusion. Autophagy 2015, 11, 1437–1438. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.; Kumar, S.; Jain, A.; Ponpuak, M.; Mudd, M.H.; Kimura, T.; Choi, S.W.; Peters, R.; Mandell, M.; Bruun, J.A.; et al. TRIMs and galectins globally cooperate and TRIM16 and galectin-3 co-direct autophagy in endomembrane damage homeostasis. Dev. Cell 2016, 39, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Maejima, I.; Takahashi, A.; Omori, H.; Kimura, T.; Takabatake, Y.; Saitoh, T.; Yamamoto, A.; Hamasaki, M.; Noda, T.; Isaka, Y.; et al. Autophagy sequesters damaged lysosomes to control lysosomal biogenesis and kidney injury. EMBO J. 2013, 32, 2336–2347. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Y.; Jing, L.; Wang, J.; Yan, Y.; Feng, Y.; Zhang, Y.; Liu, Z.; Ma, L.; Diao, A. Lysosome Inhibitors Enhance the Chemotherapeutic Activity of Doxorubicin in HepG2 Cells. Chemotherapy 2017, 62, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.; Nandy, S.K.; Cantí, C.; Lavandero, S. Bafilomycin-A1 and ML9 Exert Different Lysosomal Actions to Induce Cell Death. Curr. Mol. Pharmacol. 2019, 12, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, F.; Gawarammana, I.; Robertson, T.A.; Roberts, M.S.; Palangasinghe, C.; Zawahir, S.; Jayamanne, S.; Kandasamy, J.; Eddleston, M.; Buckley, N.A.; et al. Acute Human Self-Poisoning with Imidacloprid Compound: A Neonicotinoid Insecticide. PLoS ONE 2009, 4, e5127. [Google Scholar] [CrossRef]
- Sriapha, C.; Trakulsrichai, S.; Tongpoo, A.; Pradoo, A.; Rittilert, P.; Wananukul, W. Acute Imidacloprid Poisoning in Thailand. Ther. Clin. Risk Manag. 2020, 16, 1081. [Google Scholar] [CrossRef] [PubMed]
- Bonmatin, J.M.; Giorio, C.; Girolami, V.; Goulson, D.; Kreutzweiser, D.P.; Krupke, C.; Liess, M.; Long, E.; Marzaro, M.; Mitchell, E.A.; et al. Environ-mental fate and exposure neonicotinoids and fipronil. Environ. Sci. Pollut. Res. Int. 2015, 22, 35–67. [Google Scholar] [CrossRef] [PubMed]
- Suchail, S.; Guez, D.; Belzunces, L.P. Discrepancy between Acute and Chronic Toxicity Induced by Imidacloprid and Its Metabolites in Apis Mellifera. Environ. Toxicol. Chem. 2001, 20, 2482–2486. [Google Scholar] [CrossRef] [PubMed]
- Seifrtova, M.; Halesova, T.; Sulcova, K.; Riddellova, K.; Erban, T. Distributions of Imidacloprid, Imidacloprid-Olefin and Imidacloprid-Urea in Green Plant Tissues and Roots of Rapeseed (Brassica napus) from Artificially Contaminated Potting Soil. Pest. Manag. Sci. 2017, 73, 1010–1016. [Google Scholar] [CrossRef]
- Erban, T.; Sopko, B.; Talacko, P.; Harant, K.; Kadlikova, K.; Halesova, T.; Riddellova, K.; Pekas, A. Chronic Exposure of Bumblebees to Neonicotinoid Imidacloprid Suppresses the Entire Mevalonate Pathway and Fatty Acid Synthesis. J. Proteomics 2019, 196, 69–80. [Google Scholar] [CrossRef]
- Guimarães, A.R.D.J.S.; Bizerra, P.F.V.; Miranda, C.A.; Mingatto, F.E. Effects of Imidacloprid on Viability and Increase of Reactive Oxygen and Nitrogen Species in HepG2 Cell Line. Toxicol. Mech. Methods 2022, 32, 204–212. [Google Scholar] [CrossRef]
- Wolfe, D.M.; Lee, J.H.; Kumar, A.; Lee, S.; Orenstein, S.J.; Nixon, R.A. Autophagy failure in Alzheimer’s disease and the role of defective lysosomal acidification. Eur. J. Neurosci. 2013, 37, 1949–1961. [Google Scholar] [CrossRef]
- Vidyadhara, D.J.; Lee, J.E.; Chandra, S.S. Role of the endolysosomal system in Parkinson’s disease. J. Neurochem. 2019, 150, 487–506. [Google Scholar] [CrossRef] [PubMed]
- Flavin, W.P.; Bousset, L.; Green, Z.C.; Chu, Y.; Skarpathiotis, S.; Chaney, M.J.; Kordower, J.H.; Melki, R.; Campbell, E.M. Endocytic vesicle rupture is a conserved mechanism of cellular invasion by amyloid proteins. Acta. Neuropathol. 2017, 134, 629–653. [Google Scholar] [CrossRef] [PubMed]
- Dehay, B.; Bove, J.; Rodriguez-Muela, N.; Perier, C.; Recasens, A.; Boya, P.; Vila, M. Pathogenic lysosomal depletion in Parkinson’s disease. J. Neurosci. 2010, 30, 12535–12544. [Google Scholar] [CrossRef] [PubMed]
- Hook, G.; Hook, V.; Kindy, M. The cysteine protease inhibitor, E64d, reduces brain amyloid-beta and improves memory deficits in Alzheimer’s disease animal models by inhibiting cathepsin B, but not BACE1, beta-secretase activity. J. Alzheimers. Dis. 2011, 26, 387–408. [Google Scholar] [CrossRef]
- Fu, Z.; Han, F.; Huang, K.; Zhang, J.; Qin, J.G.; Chen, L.; Li, E. Impact of Imidacloprid Exposure on the Biochemical Responses, Transcriptome, Gut Microbiota and Growth Performance of the Pacific White Shrimp Litopenaeus Vannamei. J. Hazard Mater. 2022, 424, 127513. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eriksson, I.; Ward, L.J.; Vainikka, L.; Sultana, N.; Leanderson, P.; Flodin, U.; Li, W.; Yuan, X.-M. Imidacloprid Induces Lysosomal Dysfunction and Cell Death in Human Astrocytes and Fibroblasts—Environmental Implication of a Clinical Case Report. Cells 2023, 12, 2772. https://doi.org/10.3390/cells12242772
Eriksson I, Ward LJ, Vainikka L, Sultana N, Leanderson P, Flodin U, Li W, Yuan X-M. Imidacloprid Induces Lysosomal Dysfunction and Cell Death in Human Astrocytes and Fibroblasts—Environmental Implication of a Clinical Case Report. Cells. 2023; 12(24):2772. https://doi.org/10.3390/cells12242772
Chicago/Turabian StyleEriksson, Ida, Liam J. Ward, Linda Vainikka, Nargis Sultana, Per Leanderson, Ulf Flodin, Wei Li, and Xi-Ming Yuan. 2023. "Imidacloprid Induces Lysosomal Dysfunction and Cell Death in Human Astrocytes and Fibroblasts—Environmental Implication of a Clinical Case Report" Cells 12, no. 24: 2772. https://doi.org/10.3390/cells12242772
APA StyleEriksson, I., Ward, L. J., Vainikka, L., Sultana, N., Leanderson, P., Flodin, U., Li, W., & Yuan, X. -M. (2023). Imidacloprid Induces Lysosomal Dysfunction and Cell Death in Human Astrocytes and Fibroblasts—Environmental Implication of a Clinical Case Report. Cells, 12(24), 2772. https://doi.org/10.3390/cells12242772