
The Potential of PSMA as a Vascular Target in TNBC


 , , and
, , and 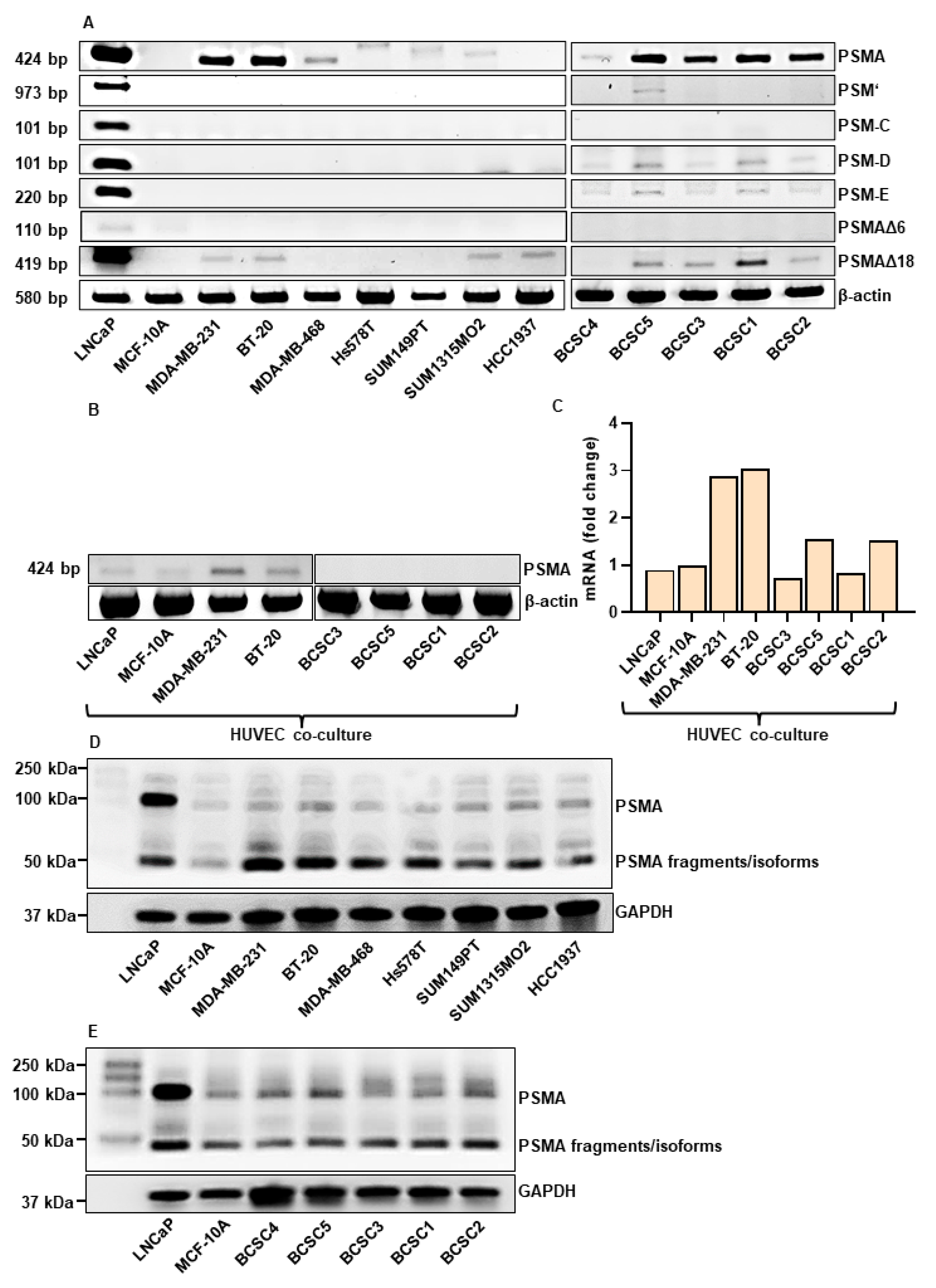{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture Conditions
2.2. Radiochemistry
2.3. RNA Isolation and Reverse Transcription
2.4. PCR
2.5. qPCR
2.6. Western Blot Analysis
2.7. Co-Cultures
2.8. Immunocytochemistry
2.9. Generation of Tumor Conditioned Media (TCM)
2.10. Tube Formation Assay
2.11. Spheroid Formation
2.12. Immunohistochemistry
2.13. Cell Uptake Experiments
2.14. Flow Cytometry
2.15. Quantification and Statistics
3. Results
3.1. PSMA is Expressed in TNBC Cells and BCSCs
3.2. TNBC Cells Promote Tube Formation In Vitro
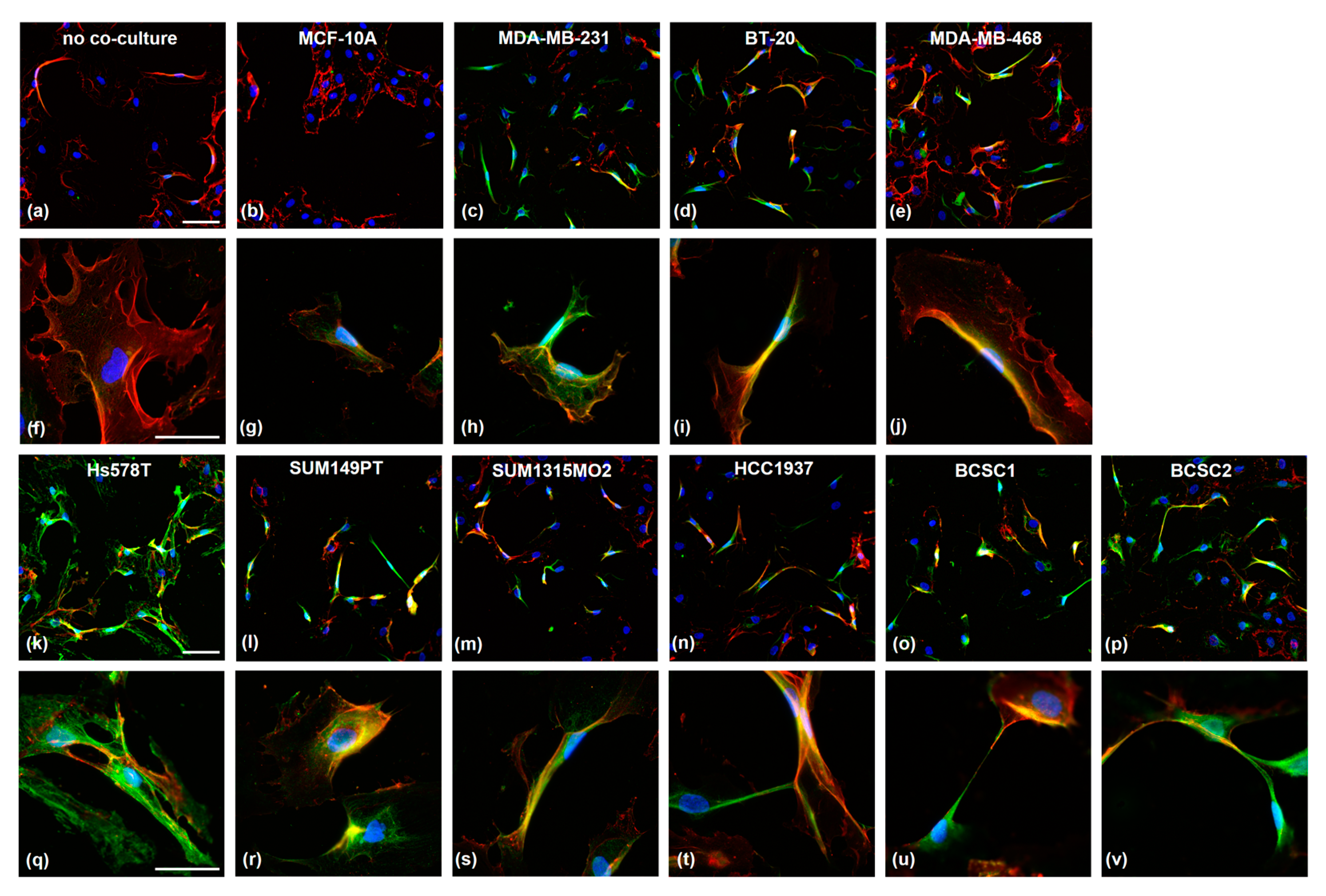
3.3. TNBC Cells Induce PSMA Expression in HUVECs and Affect Their Morphology
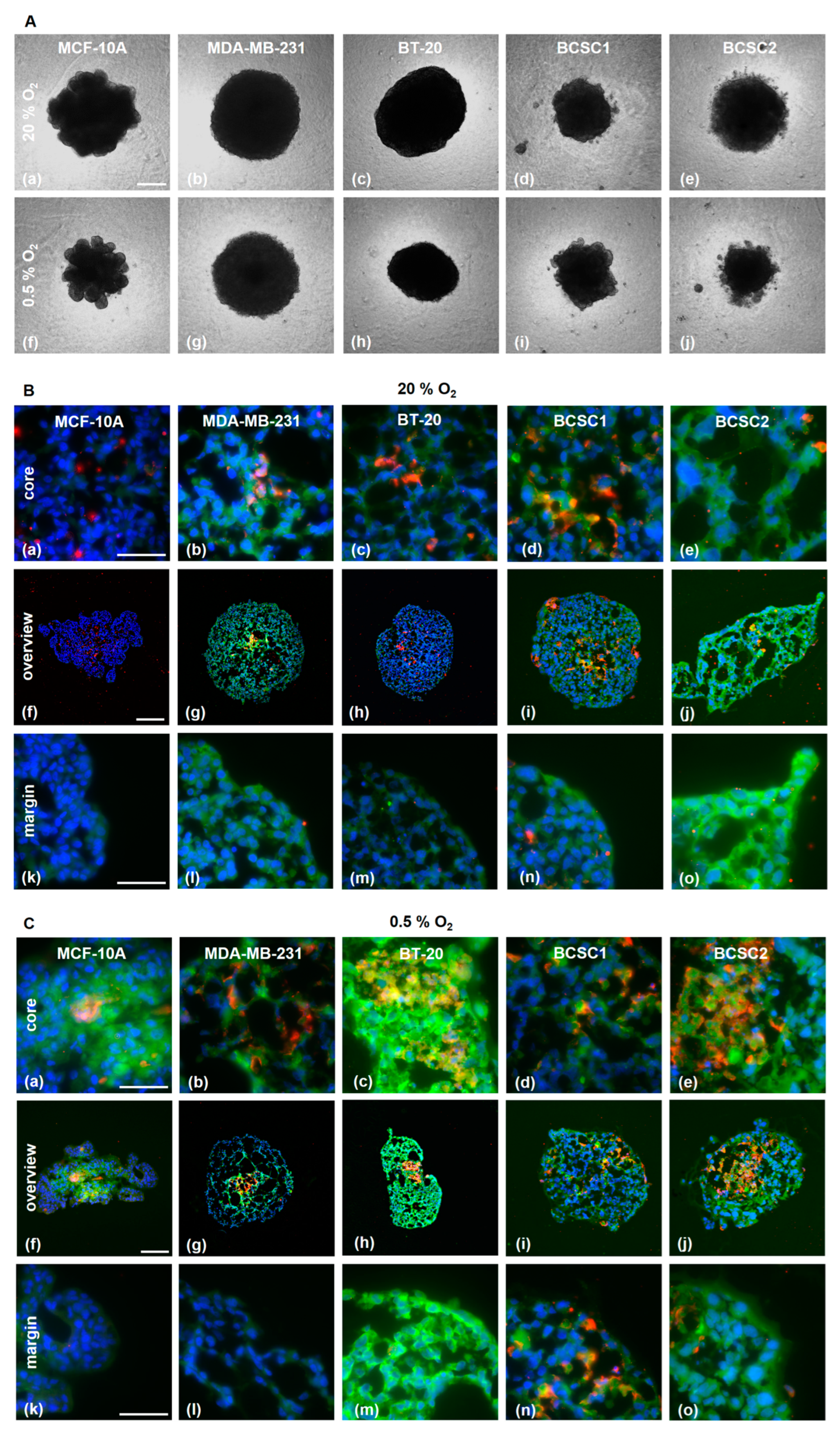
3.4. PSMA is Expressed in Endothelial Cells in TNBC/HUVEC Spheroids
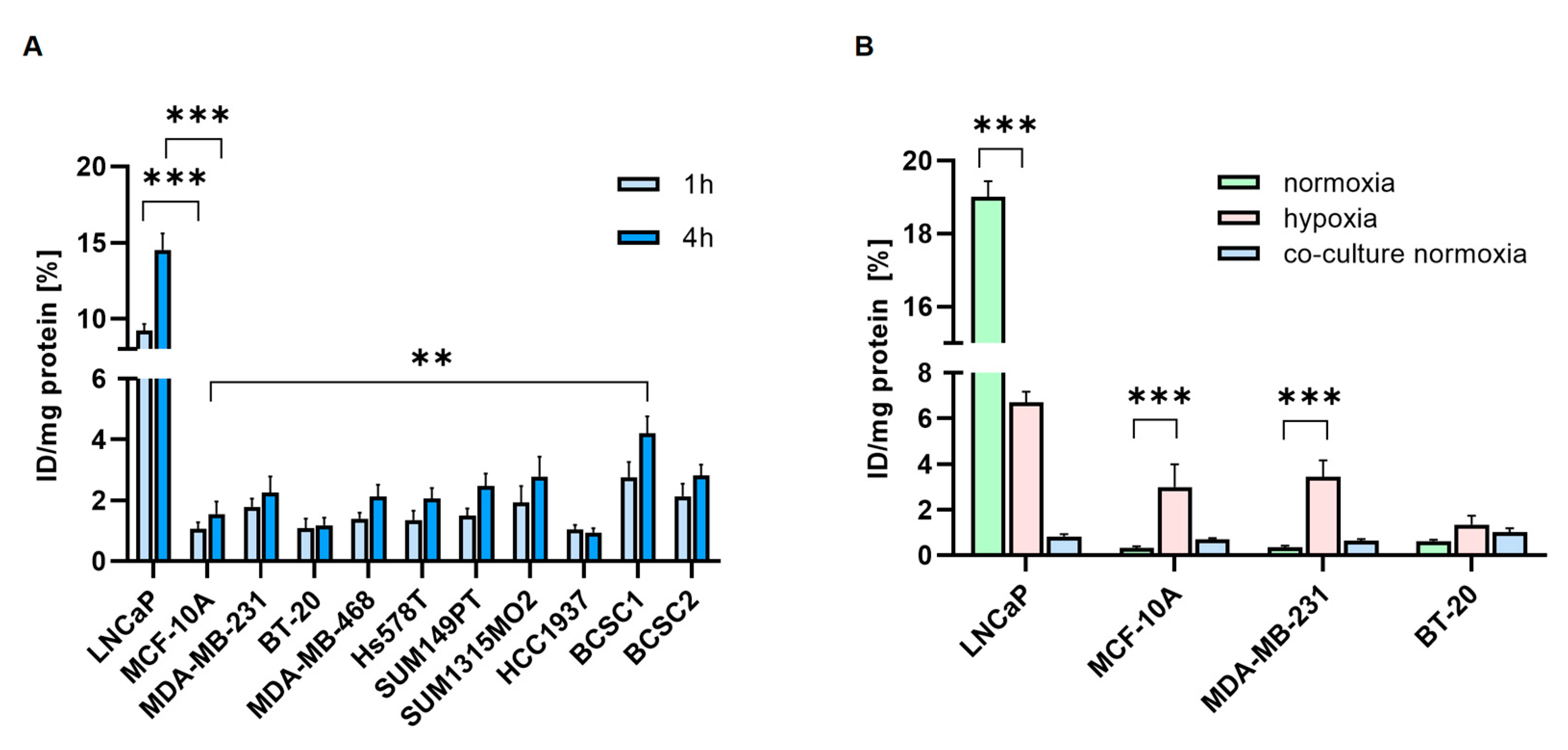
3.5. Radiolabeled PSMA-Ligands are Taken up by TNBC Cells
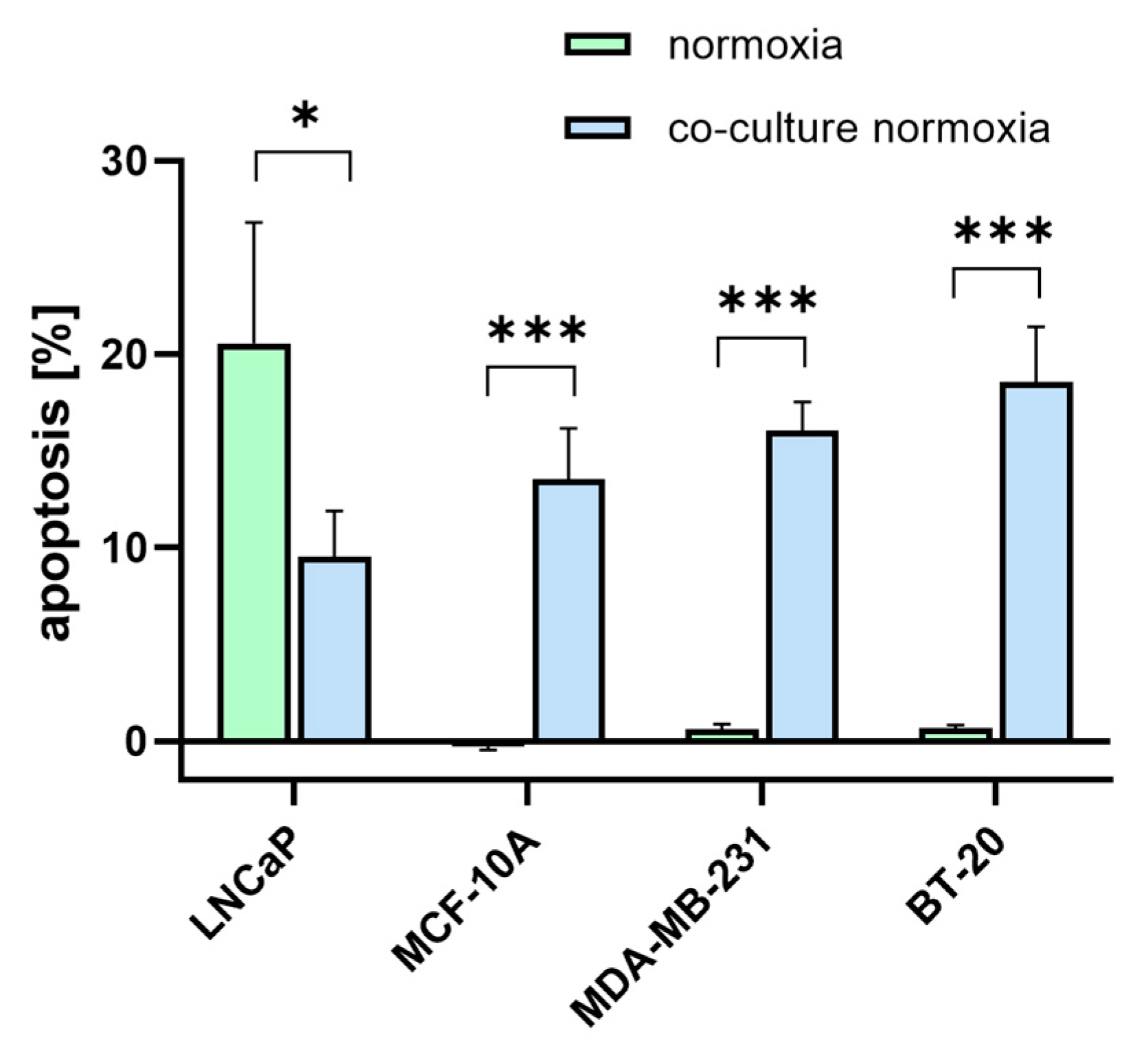
3.6. [177Lu]Lu-PSMA Induces Apoptosis in TNBC-Associated HUVECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tečić Vuger, A.; Šeparović, R.; Vazdar, L.; Pavlović, M.; Lepetić, P.; Šitić, S.; Bajić, Ž.; Šarčević, B.; Vrbanec, D. Characteristics and Prognosis of Triple-Negative Breast Cancer Patients: A Croatian Single Institution Retrospective Cohort Study. Acta Clin. Croat. 2020, 59, 97. [Google Scholar] [CrossRef]
- O’Reilly, E.A.; Gubbins, L.; Sharma, S.; Tully, R.; Guang, M.H.Z.; Weiner-Gorzel, K.; McCaffrey, J.; Harrison, M.; Furlong, F.; Kell, M.; et al. The fate of chemoresistance in triple negative breast cancer (TNBC). BBA Clin. 2015, 3, 257–275. [Google Scholar] [CrossRef]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef]
- He, J.; Lee, H.J.; Saha, S.; Ruan, D.; Guo, H.; Chan, C.H. Inhibition of USP2 eliminates cancer stem cells and enhances TNBC responsiveness to chemotherapy. Cell Death Dis. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Elbaiomy, M.A.; Akl, T.; Atwan, N.; Elsayed, A.A.; Elzaafarany, M.; Shamaa, S. Clinical Impact of Breast Cancer Stem Cells in Metastatic Breast Cancer Patients. J. Oncol. 2020, 2020, 2561726. [Google Scholar] [CrossRef]
- Mease, R.C.; Foss, C.A.; Pomper, M.G. PET Imaging in Prostate Cancer: Focus on Prostate-Specific Membrane Antigen. Curr. Top. Med. Chem. 2013, 13, 951–962. [Google Scholar] [CrossRef]
- Rajasekaran, A.K.; Anilkumar, G.; Christiansen, J.J. Is prostate-specific membrane antigen a multifunctional protein? Am. J. Physiol.-Cell Physiol. 2005, 288, 975–981. [Google Scholar] [CrossRef]
- Uijen, M.J.M.; Derks, Y.H.W.; Merkx, R.I.J.; Schilham, M.G.M.; Roosen, J.; Privé, B.M.; van Lith, S.A.M.; van Herpen, C.M.L.; Gotthardt, M.; Heskamp, S.; et al. PSMA radioligand therapy for solid tumors other than prostate cancer: Background, opportunities, challenges, and first clinical reports. Eur. J. Nucl. Med. Mol. Imaging 2021, 48, 4350–4368. [Google Scholar] [CrossRef]
- Wernicke, A.G.; Varma, S.; Greenwood, E.A.; Christos, P.J.; Chao, K.S.C.; Liu, H.; Bander, N.H.; Shin, S.J. Prostate-specific membrane antigen expression in tumor-associated vasculature of breast cancers. APMIS 2014, 122, 482–489. [Google Scholar] [CrossRef]
- Kaittanis, C.; Andreou, C.; Hieronymus, H.; Mao, N.; Foss, C.A.; Eiber, M.; Weirich, G.; Panchal, P.; Gopalan, A.; Zurita, J.; et al. Prostate-specific membrane antigen cleavage of vitamin B9 stimulates oncogenic signaling through metabotropic glutamate receptors. J. Exp. Med. 2018, 215, 159. [Google Scholar] [CrossRef] [Green Version]
- Conway, R.E.; Petrovic, N.; Li, Z.; Heston, W.; Wu, D.; Shapiro, L.H. Prostate-Specific Membrane Antigen Regulates Angiogenesis by Modulating Integrin Signal Transduction. Mol. Cell. Biol. 2006, 26, 5310. [Google Scholar] [CrossRef]
- Conway, R.E.; Joiner, K.; Patterson, A.; Bourgeois, D.; Rampp, R.; Hannah, B.C.; McReynolds, S.; Elder, J.M.; Gilfilen, H.; Shapiro, L.H. Prostate specific membrane antigen produces pro-angiogenic laminin peptides downstream of matrix metalloprotease-2. Angiogenesis 2013, 16, 847–860. [Google Scholar] [CrossRef]
- Nguyen, D.P.; Xiong, P.L.; Liu, H.; Pan, S.; Leconet, W.; Navarro, V.; Guo, M.; Moy, J.; Kim, S.; Ramirez-Fort, M.K.; et al. Induction of PSMA and Internalization of an Anti-PSMA mAb in the Vascular Compartment. Mol. Cancer Res. 2016, 14, 1045–1053. [Google Scholar] [CrossRef]
- Morgenroth, A.; Tinkir, E.; Vogg, A.T.J.; Sankaranarayanan, R.A.; Baazaoui, F.; Mottaghy, F.M. Targeting of prostate-specific membrane antigen for radio-ligand therapy of triple-negative breast cancer. Breast Cancer Res. 2019, 21, 116. [Google Scholar] [CrossRef]
- Gao, Y.; Zheng, H.; Li, L.; Feng, M.; Chen, X.; Hao, B.; Lv, Z.; Zhou, X.; Cao, Y. Prostate-Specific Membrane Antigen (PSMA) Promotes Angiogenesis of Glioblastoma Through Interacting with ITGB4 and Regulating NF-κB Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 462. [Google Scholar] [CrossRef]
- Namba, T.; Koike, H.; Murakami, K.; Aoki, M.; Makino, H.; Hashiya, N.; Ogihara, T.; Kaneda, Y.; Kohno, M.; Morishita, R. Angiogenesis Induced by Endothelial Nitric Oxide Synthase Gene Through Vascular Endothelial Growth Factor Expression in a Rat Hindlimb Ischemia Model. Circulation 2003, 108, 2250–2257. [Google Scholar] [CrossRef]
- Unger, C.; Bronsert, P.; Michalski, K.; Bicker, A.; Juhasz-Böss, I. Expression of Prostate Specific Membrane Antigen (PSMA) in Breast Cancer. Geburtshilfe Frauenheilkd. 2022, 82, 50. [Google Scholar] [CrossRef]
- Watanabe, R.; Maekawa, M.; Kiyoi, T.; Kurata, M.; Miura, N.; Kikugawa, T.; Higashiyama, S.; Saika, T. PSMA-positive membranes secreted from prostate cancer cells have potency to transform vascular endothelial cells into an angiogenic state. Prostate 2021, 81, 1390–1401. [Google Scholar] [CrossRef]
- Apte, R.S.; Chen, D.S.; Ferrara, N. VEGF in Signaling and Disease: Beyond Discovery and Development. Cell 2019, 176, 1248. [Google Scholar] [CrossRef]
- Diéras, V.; Campone, M.; Yardley, D.A.; Romieu, G.; Valero, V.; Isakoff, S.J.; Koeppen, H.; Wilson, T.R.; Xiao, Y.; Shames, D.S.; et al. Randomized, phase II, placebo-controlled trial of onartuzumab and/or bevacizumab in combination with weekly paclitaxel in patients with metastatic triple-negative breast cancer. Ann. Oncol. 2015, 26, 1904–1910. [Google Scholar] [CrossRef]
- Curigliano, G.; Pivot, X.; Cortés, J.; Elias, A.; Cesari, R.; Khosravan, R.; Collier, M.; Huang, X.; Cataruozolo, P.E.; Kern, K.A.; et al. Randomized phase II study of sunitinib versus standard of care forpatients with previously treated advanced triple-negative breastcancer. Breast 2013, 22, 650–656. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, D.; Yao, Z.; Lin, X.; Liu, J.; Gu, Q.; Dong, X.; Liu, F.; Wang, Y.; Yao, N.; et al. Anti-angiogenic treatment promotes triple-negative breast cancer invasion via vasculogenic mimicry. Cancer Biol. Ther. 2017, 18, 205. [Google Scholar] [CrossRef]
- van Kalmthout, L.W.M.; van der Sar, E.C.A.; Braat, A.J.A.T.; de Keizer, B.; Lam, M.G.E.H. Lutetium-177-PSMA therapy for prostate cancer patients—A brief overview of the literature. Tijdschr. voor Urol. 2020, 10, 141–146. [Google Scholar] [CrossRef]
- Schneider, C.A.; Täger, P.; Hammes, J.; Fischer, T.; Drzezga, A.; Pfister, D.; Heidenreich, A.; Schmidt, M. Treatment outcome and identification of factors influencing overall survival after Lu-177-PSMA-617 radioligand therapy in metastatic prostate cancer. Nuklearmedizin 2022, 61, 25–32. [Google Scholar] [CrossRef]
- Stuparu, A.D.; Capri, J.R.; Meyer, C.A.L.; Le, T.M.; Evans-Axelsson, S.L.; Current, K.; Lennox, M.; Mona, C.E.; Fendler, W.P.; Calais, J.; et al. Mechanisms of Resistance to Prostate-Specific Membrane Antigen–Targeted Radioligand Therapy in a Mouse Model of Prostate Cancer. J. Nucl. Med. 2021, 62, 989–995. [Google Scholar] [CrossRef]
- Su, S.L.; Huang, I.-P.; Fair, W.R.; Powell, C.T.; Heston, W.D.W. Alternatively Spliced Variants of Prostate-specific Membrane Antigen RNA: Ratio of Expression as a Potential Measurement of Progression. Cancer Res. 1995, 55, 1441–1443. [Google Scholar]
- Schmittgen, T.D.; Teske, S.; Vessella, R.L.; True, L.D.; Zakrajsek, B.A. Expression of prostate specific membrane antigen and three alternatively spliced variants of PSMA in prostate cancer patients. Int. J. cancer 2003, 107, 323–329. [Google Scholar] [CrossRef]
- Williams, T.; Kole, R. Analysis of Prostate-Specific Membrane Antigen Splice Variants in LNCap Cells. Oligonucleotides 2006, 16, 186–195. [Google Scholar] [CrossRef]
- Cao, K.Y.; Mao, X.P.; Wang, D.H.; Xu, L.; Yuan, G.Q.; Dai, S.Q.; Zheng, B.J.; Qiu, S.P. High expression of PSM-E correlated with tumor grade in prostate cancer: A new alternatively spliced variant of prostate-specific membrane antigen. Prostate 2007, 67, 1791–1800. [Google Scholar] [CrossRef]
- Metzger, E.; Stepputtis, S.S.; Strietz, J.; Preca, B.T.; Urban, S.; Willmann, D.; Allen, A.; Zenk, F.; Iovino, N.; Bronsert, P.; et al. KDM4 inhibition targets breast cancer stem–like cells. Cancer Res. 2017, 77, 5900–5912. [Google Scholar] [CrossRef]
- Strietz, J.; Stepputtis, S.S.; Follo, M.; Bronsert, P.; Stickeler, E.; Maurer, J. Human Primary Breast Cancer Stem Cells Are Characterized by Epithelial-Mesenchymal Plasticity. Int. J. Mol. Sci. 2021, 22, 1–19. [Google Scholar] [CrossRef]
- Li, X.; Strietz, J.; Bleilevens, A.; Stickeler, E.; Maurer, J. Chemotherapeutic Stress Influences Epithelial–Mesenchymal Transition and Stemness in Cancer Stem Cells of Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2020, 21, 404. [Google Scholar] [CrossRef]
- Pinto, J.T.; Suffoletto, B.P.; Berzin, T.M.; Hong Qiao, C.; Lin, S.; Tong, W.P.; May, F.; Mukherjee, B.; Heston, W.D.W.; Tj, W.P.; et al. Prostate-specific Membrane Antigen: A Novel Folate Hydrolase in Human Prostatic Carcinoma Cells’. Clin. Cancer Res. 1996, 2, 1445–1451. [Google Scholar]
- Luthi-Carter, R.; Barczak, A.; Speno, H.; Coyle, J. Molecular Characterization of Human Brain N-Acetylated a-Linked Acidic Dipeptidase (NAALADase) 1. J. Pharmacol. Exp. Ther. 1998, 286, 1020–1025. [Google Scholar]
- Seifert, R.; Kessel, K.; Boegemann, M.; Köhler, M.; Roll, W.; Stegger, L.; Weckesser, M.; Rahbar, K. Additional Local Therapy for Liver Metastases in Patients with Metastatic Castration-Resistant Prostate Cancer Receiving Systemic PSMA-Targeted Therapy. J. Nucl. Med. 2020, 61, 723–728. [Google Scholar] [CrossRef]
- Devlin, A.M.; Ling, E.H.; Peerson, J.M.; Fernando, S.; Clarke, R.; Smith, A.D.; Halsted, C.H. Glutamate carboxypeptidase II: A polymorphism associated with lower levels of serum folate and hyperhomocysteinemia. Hum. Mol. Genet. 2000, 9, 2837–2844. [Google Scholar] [CrossRef]
- Fang, A.P.; Liu, Z.Y.; Liao, G.C.; Chen, P.Y.; Wang, X.Y.; Zhang, D.M.; Luo, Y.; Long, J.A.; Zhong, R.H.; Zhou, Z.G.; et al. Serum folate concentrations at diagnosis are associated with hepatocellular carcinoma survival in the Guangdong Liver Cancer Cohort study. Br. J. Nutr. 2019, 121, 1376–1388. [Google Scholar] [CrossRef]
- Yao, V.; Berkman, C.E.; Choi, J.K.; O’Keefe, D.S.; Bacich, D.J. Expression of prostate-specific membrane antigen (PSMA), increases cell folate uptake and proliferation and suggests a novel role for PSMA in the uptake of the non-polyglutamated folate, folic acid. Prostate 2010, 70, 305–316. [Google Scholar] [CrossRef]
- Wu, K.; Xu, H.; Wu, K.; Tian, Y.; Liu, Q.; Han, N.; Yuan, X.; Zhang, L.; Wu, G.S. CD44 correlates with clinicopathological characteristics and is upregulated by EGFR in breast cancer. Int. J. Oncol. 2016, 49, 1343. [Google Scholar] [CrossRef]
- Kim, H.; Woo, J.; Dan, K.; Lee, K.M.; Jin, M.S.; Park, I.A.; Ryu, H.S.; Han, D. Quantitative Proteomics Reveals Knockdown of CD44 Promotes Proliferation and Migration in Claudin-Low MDA-MB-231 and Hs 578T Breast Cancer Cell Lines. J. Proteome Res. 2021, 20, 3720–3733. [Google Scholar] [CrossRef]
- Nagano, O.; Okazaki, S.; Saya, H. Redox regulation in stem-like cancer cells by CD44 variant isoforms. Oncogene 2013, 32, 5191–5198. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.L.; Klimova, T.A.; Eisenbart, J.; Schumacker, P.T.; Chandel, N.S. Mitochondrial reactive oxygen species trigger hypoxia-inducible factor-dependent extension of the replicative life span during hypoxia. Mol. Cell. Biol. 2007, 27, 5737–5745. [Google Scholar] [CrossRef] [PubMed]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Saha, T.; Lukong, K.E. Breast Cancer Stem-Like Cells in Drug Resistance: A Review of Mechanisms and Novel Therapeutic Strategies to Overcome Drug Resistance. Front. Oncol. 2022, 12, 1. [Google Scholar] [CrossRef]
- Zuazo-Gaztelu, I.; Casanovas, O. Unraveling the role of angiogenesis in cancer ecosystems. Front. Oncol. 2018, 8, 248. [Google Scholar] [CrossRef]
- Dai, C.; Heemers, H.; Sharifi, N. Androgen Signaling in Prostate Cancer. Cold Spring Harb. Perspect. Med. 2017, 7, a030452. [Google Scholar] [CrossRef]
- Karagiannis, G.S.; Saraon, P.; Jarvi, K.A.; Diamandis, E.P. Proteomic signatures of angiogenesis in androgen-independent prostate cancer. Prostate 2014, 74, 260–272. [Google Scholar] [CrossRef]
- Liu, C.Y.; Lau, K.Y.; Hsu, C.C.; Chen, J.L.; Lee, C.H.; Huang, T.T.; Chen, Y.T.; Huang, C.T.; Lin, P.H.; Tseng, L.M. Combination of palbociclib with enzalutamide shows in vitro activity in RB proficient and androgen receptor positive triple negative breast cancer cells. PLoS ONE 2017, 12, e0189007. [Google Scholar] [CrossRef]
- Bogina, G.S.; Lunardi, G.; Marcolini, L.; Brunelli, M.; Bortesi, L.; Marconi, M.; Coati, F.; Valerio, M.; Guerriero, M.; Massocco, A.; et al. P16 but not retinoblastoma expression is related to clinical outcome in no-special-type triple-negative breast carcinomas. Mod. Pathol. 2014, 27, 204–213. [Google Scholar] [CrossRef]
- Harada, H.; Nakagavva, K.; Iwata, S.; Saito, M.; Kumon, Y.; Sakaki, S.; Sato, K.; Hamada, K. Restoration of wild-type p16 down-regulates vascular endothelial growth factor expression and inhibits angiogenesis in human gliomas. Cancer Res. 1999, 59, 3783–3789. [Google Scholar]
- Chintala, S.K.; Fueyo, J.; Gomez-Manzano, C.; Venkaiah, B.; Bjerkvig, R.; Yung, W.K.A.; Sawaya, R.; Kyritsis, A.P.; Rao, J.S. Adenovirus-mediated p16/CDKN2 gene transfer suppresses glioma invasion in vitro. Oncogene 1997, 15, 2049–2057. [Google Scholar] [CrossRef]
- Xu, L.; Wang, Z.; Li, X.F.; He, X.; Guan, L.L.; Tuo, J.L.; Wang, Y.; Luo, Y.; Zhong, H.L.; Qiu, S.P.; et al. Screening and identification of significant genes related to tumor metastasis and PSMA in prostate cancer using microarray analysis. Oncol. Rep. 2013, 30, 1920–1928. [Google Scholar] [CrossRef] [PubMed]
- Caromile, L.A.; Dortche, K.; Rahman, M.M.; Grant, C.L.; Stoddard, C.; Ferrer, F.A.; Shapiro, L.H. PSMA redirects cell survival signaling from the MAPK to the PI3K-AKT pathways to promote the progression of prostate cancer. Sci. Signal. 2017, 10, 3326. [Google Scholar] [CrossRef]
- Narasimhan, H.; Ferraro, F.; Bleilevens, A.; Weiskirchen, R.; Stickeler, E.; Maurer, J. Tumor Necrosis Factor-α (TNFα) Stimulate Triple-Negative Breast Cancer Stem Cells to Promote Intratumoral Invasion and Neovasculogenesis in the Liver of a Xenograft Model. Biology 2022, 11, 1481. [Google Scholar] [CrossRef] [PubMed]
- McHenry, P.R.; Prosperi, J.R. Proteins Found in the Triple-Negative Breast Cancer Secretome and Their Therapeutic Potential. Int. J. Mol. Sci. 2023, 24, 2100. [Google Scholar] [CrossRef]
- Cheng, H.W.; Chen, Y.F.; Wong, J.M.; Weng, C.W.; Chen, H.Y.; Yu, S.L.; Chen, H.W.; Yuan, A.; Chen, J.J.W. Cancer cells increase endothelial cell tube formation and survival by activating the PI3K/Akt signalling pathway. J. Exp. Clin. Cancer Res. 2017, 36, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Gericke, B.; Fracasso, G.; Ramarli, D.; Colombatti, M.; Naim, H.Y. Discriminatory Role of Detergent-Resistant Membranes in the Dimerization and Endocytosis of Prostate-Specific Membrane Antigen. PLoS ONE 2013, 8, e66193. [Google Scholar] [CrossRef]
- Anilkumar, G.; Rajasekaran, S.; Wang, S.; Hankinson, O.; Bander, N.; Rajasekaran, A. Prostate-specific membrane antigen association with filamin A modulates its internalization and NAALADase activity. Cancer Res. 2003, 63, 2645–2648. [Google Scholar]
- Uzunalli, G.; Dieterly, A.M.; Kemet, C.M.; Weng, H.Y.; Soepriatna, A.H.; Goergen, C.J.; Shinde, A.B.; Wendt, M.K.; Lyle, L.T. Dynamic transition of the blood-brain barrier in the development of non-small cell lung cancer brain metastases. Oncotarget 2019, 10, 6334–6348. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, G.; Ni, P.; Jiang, X.; Huang, H.; Wu, J.; Shi, X.; Jiang, X.; Liu, J. CD31 and D2-40 Contribute to Peritoneal Metastasis of Colorectal Cancer by Promoting Epithelial-Mesenchymal Transition. Gut Liver 2021, 15, 273. [Google Scholar] [CrossRef]
- Wong, C.W.Y.; Wiedle, G.; Ballestrem, C.; Wehrle-Haller, B.; Etteldorf, S.; Bruckner, M.; Engelhardt, B.; Gisler, R.H.; Imhof, B.A. PECAM-1/CD31 Trans-homophilic Binding at the Intercellular Junctions Is Independent of Its Cytoplasmic Domain; Evidence for Heterophilic Interaction with Integrin αvβ3 in Cis. Mol. Biol. Cell 2000, 11, 3109. [Google Scholar] [CrossRef]
- MacPherson, M.; Fagerholm, S.C. Filamin and filamin-binding proteins in integrin-regulation and adhesion. Focus on: “FilaminA is required for vimentin-mediated cell adhesion and spreading”. Am. J. Physiol.-Cell Physiol. 2010, 298, C206–C208. [Google Scholar] [CrossRef]
- Butterworth, K.T.; McCarthy, H.O.; Devlin, A.; Ming, L.; Robson, T.; McKeown, S.R.; Worthington, J. Hypoxia selects for androgen independent LNCaP cells with a more malignant geno- and phenotype. Int. J. Cancer 2008, 123, 760–768. [Google Scholar] [CrossRef]
- Xie, J.; Xiao, Y.; Zhu, X.Y.; Ning, Z.Y.; Xu, H.F.; Wu, H.M. Hypoxia regulates stemness of breast cancer MDA-MB-231 cells. Med. Oncol. 2016, 33, 42. [Google Scholar] [CrossRef]
- Tolkach, Y.; Gevensleben, H.; Bundschuh, R.; Koyun, A.; Huber, D.; Kehrer, C.; Hecking, T.; Keyver-Paik, M.D.; Kaiser, C.; Ahmadzadehfar, H.; et al. Prostate-specific membrane antigen in breast cancer: A comprehensive evaluation of expression and a case report of radionuclide therapy. Breast Cancer Res. Treat. 2018, 169, 447–455. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heesch, A.; Ortmanns, L.; Maurer, J.; Stickeler, E.; Sahnoun, S.E.M.; Mottaghy, F.M.; Morgenroth, A. The Potential of PSMA as a Vascular Target in TNBC. Cells 2023, 12, 551. https://doi.org/10.3390/cells12040551
Heesch A, Ortmanns L, Maurer J, Stickeler E, Sahnoun SEM, Mottaghy FM, Morgenroth A. The Potential of PSMA as a Vascular Target in TNBC. Cells. 2023; 12(4):551. https://doi.org/10.3390/cells12040551
Chicago/Turabian StyleHeesch, Amelie, Lars Ortmanns, Jochen Maurer, Elmar Stickeler, Sabri E. M. Sahnoun, Felix M. Mottaghy, and Agnieszka Morgenroth. 2023. "The Potential of PSMA as a Vascular Target in TNBC" Cells 12, no. 4: 551. https://doi.org/10.3390/cells12040551
APA StyleHeesch, A., Ortmanns, L., Maurer, J., Stickeler, E., Sahnoun, S. E. M., Mottaghy, F. M., & Morgenroth, A. (2023). The Potential of PSMA as a Vascular Target in TNBC. Cells, 12(4), 551. https://doi.org/10.3390/cells12040551