
Kinetics of Plasma Cell-Free DNA under a Highly Standardized and Controlled Stress Induction

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
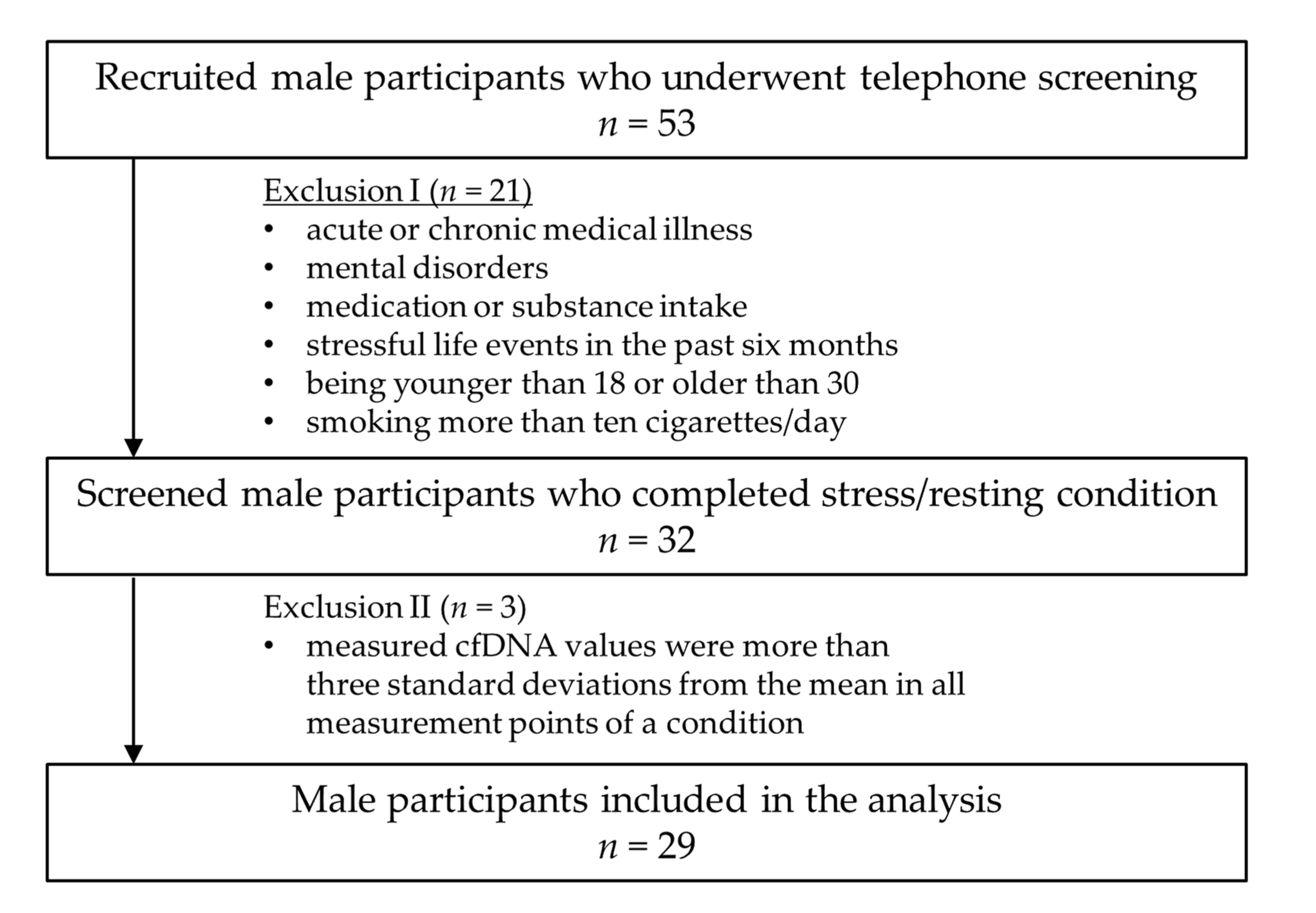
2.1. Study Participants
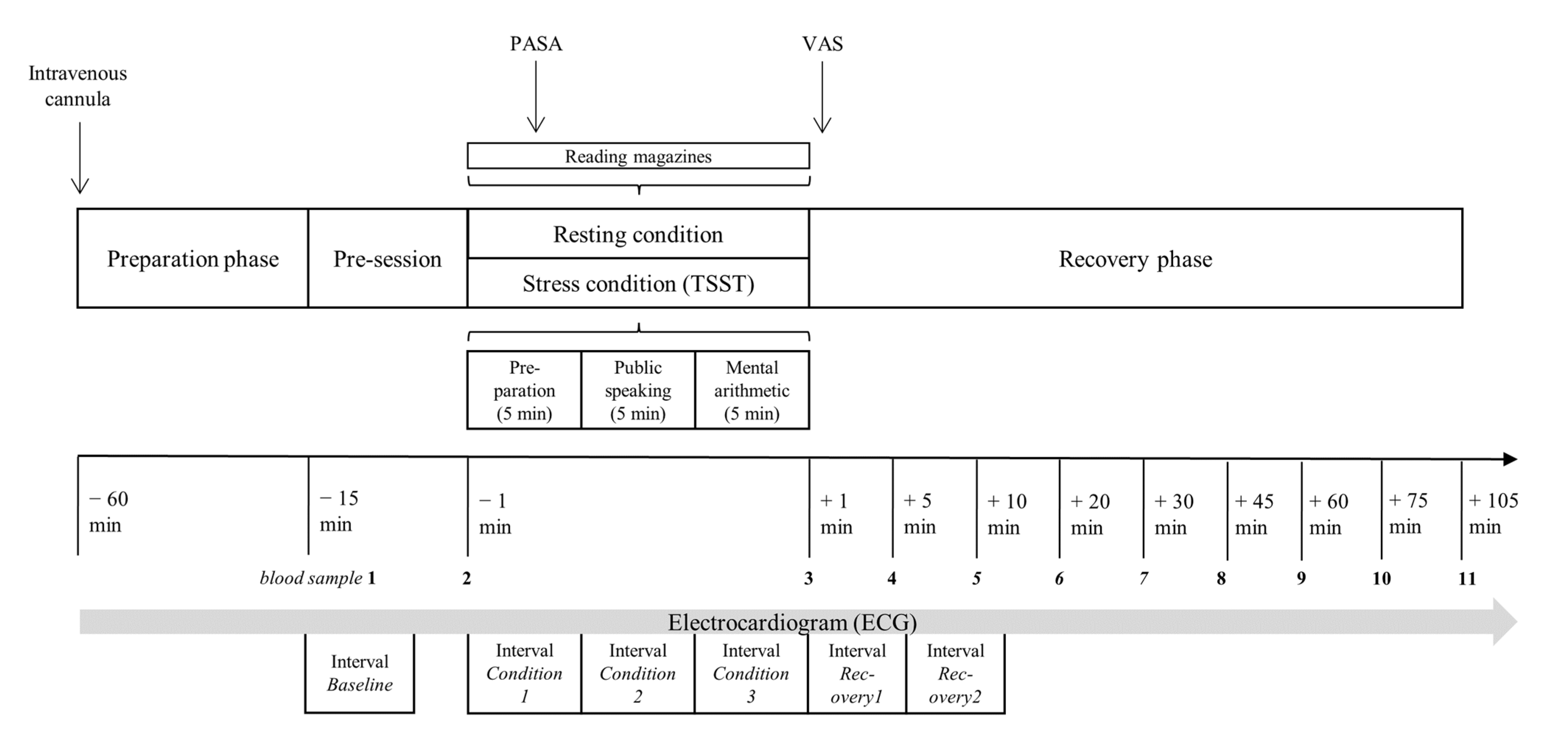
2.2. Procedures
2.3. Blood Analytics
2.4. Psychological and Clinical Measures
2.5. Heart Rate Measurement
2.6. Quantification of cfDNA
2.7. Statistical Analysis
3. Results
3.1. cfDNA Reactivity
3.2. Cortisol Reactivity
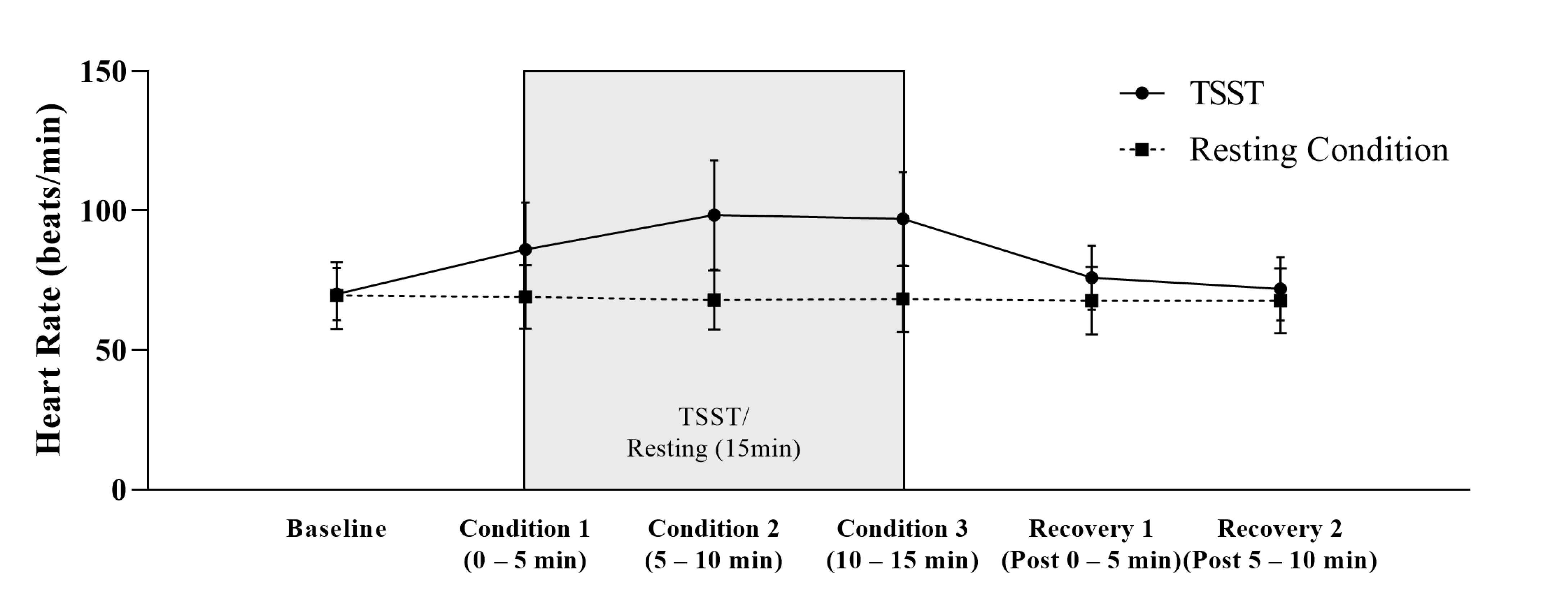
3.3. Heart Rate Reactivity
3.4. Stress Appraisal
3.5. Associations between Psychological and cfDNA and Cortisol Measures
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chrousos, G.P. Stress and Disorders of the Stress System. Nat. Rev. Endocrinol. 2009, 5, 374–381. [Google Scholar] [CrossRef]
- Chu, B.; Marwaha, K.; Sanvictores, T.; Ayers, D. Physiology, Stress Reaction; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar]
- Mifsud, K.R.; Reul, J.M.H.M. Mineralocorticoid and Glucocorticoid Receptor-Mediated Control of Genomic Responses to Stress in the Brain. Stress 2018, 21, 389–402. [Google Scholar] [CrossRef]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 247054701769232. [Google Scholar] [CrossRef]
- Wirtz, P.H.; von Känel, R. Psychological Stress, Inflammation, and Coronary Heart Disease. Curr. Cardiol. Rep. 2017, 19, 111. [Google Scholar] [CrossRef] [PubMed]
- BAuA Stressreport Deutschland 2019: Psychische Anforderungen, Ressourcen Und Befinden, 1st ed.; Druck & Verlag Kettler GmbH, Bönen: Dortmund, Germany, 2020.
- Walker, F.R.; Pfingst, K.; Carnevali, L.; Sgoifo, A.; Nalivaiko, E. In the Search for Integrative Biomarker of Resilience to Psychological Stress. Neurosci. Biobehav. Rev. 2017, 74, 310–320. [Google Scholar] [CrossRef]
- Marsland, A.L.; Walsh, C.; Lockwood, K.; John-Henderson, N.A. The Effects of Acute Psychological Stress on Circulating and Stimulated Inflammatory Markers: A Systematic Review and Meta-Analysis. Brain Behav. Immun. 2017, 64, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Hummel, E.M.; Hessas, E.; Müller, S.; Beiter, T.; Fisch, M.; Eibl, A.; Wolf, O.T.; Giebel, B.; Platen, P.; Kumsta, R.; et al. Cell-Free DNA Release under Psychosocial and Physical Stress Conditions. Transl. Psychiatry 2018, 8, 236. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Chen, L.-Y.; Shen, X.-J.; Ju, S.-Q. Analytical Value of Cell-Free DNA Based on Alu in Psychiatric Disorders. Front. Psychiatry 2020, 10, 992. [Google Scholar] [CrossRef]
- Czamanski-Cohen, J.; Sarid, O.; Cwikel, J.; Levitas, E.; Lunenfeld, E.; Douvdevani, A.; Har-Vardi, I. Decrease in Cell Free DNA Levels Following Participation in Stress Reduction Techniques among Women Undergoing Infertility Treatment. Arch. Womens Ment. Health 2014, 17, 251–253. [Google Scholar] [CrossRef]
- Moss, J.; Magenheim, J.; Neiman, D.; Zemmour, H.; Loyfer, N.; Korach, A.; Samet, Y.; Maoz, M.; Druid, H.; Arner, P.; et al. Comprehensive Human Cell-Type Methylation Atlas Reveals Origins of Circulating Cell-Free DNA in Health and Disease. Nat. Commun. 2018, 9, 5068. [Google Scholar] [CrossRef] [Green Version]
- Lo, Y.M.D.; Han, D.S.C.; Jiang, P.; Chiu, R.W.K. Epigenetics, Fragmentomics, and Topology of Cell-Free DNA in Liquid Biopsies. Science 2021, 372, eaaw3616. [Google Scholar] [CrossRef]
- Clementi, A.; Virzì, G.M.; Brocca, A.; Pastori, S.; de Cal, M.; Marcante, S.; Granata, A.; Ronco, C. The Role of Cell-Free Plasma DNA in Critically Ill Patients with Sepsis. Blood Purif. 2016, 41, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lu, X.; Shu, X.; Tian, X.; Yang, H.; Yang, W.; Zhang, Y.; Wang, G. Elevated Plasma CfDNA May Be Associated with Active Lupus Nephritis and Partially Attributed to Abnormal Regulation of Neutrophil Extracellular Traps (NETs) in Patients with Systemic Lupus Erythematosus. Intern. Med. 2014, 53, 2763–2771. [Google Scholar] [CrossRef] [PubMed]
- Jylhävä, J.; Lehtimäki, T.; Jula, A.; Moilanen, L.; Kesäniemi, Y.A.; Nieminen, M.S.; Kähönen, M.; Hurme, M. Circulating Cell-Free DNA Is Associated with Cardiometabolic Risk Factors: The Health 2000 Survey. Atherosclerosis 2014, 233, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Bronkhorst, A.J.; Ungerer, V.; Holdenrieder, S. The Emerging Role of Cell-Free DNA as a Molecular Marker for Cancer Management. Biomol. Detect. Quantif. 2019, 17, 100087. [Google Scholar] [CrossRef]
- Kananen, L.; Hurme, M.; Jylhä, M.; Härkänen, T.; Koskinen, S.; Stenholm, S.; Kähönen, M.; Lehtimäki, T.; Ukkola, O.; Jylhävä, J. Circulating Cell-Free DNA Level Predicts All-Cause Mortality Independent of Other Predictors in the Health 2000 Survey. Sci. Rep. 2020, 10, 13809. [Google Scholar] [CrossRef]
- Andargie, T.E.; Tsuji, N.; Seifuddin, F.; Jang, M.K.; Yuen, P.S.T.; Kong, H.; Tunc, I.; Singh, K.; Charya, A.; Wilkins, K.; et al. Cell-Free DNA Maps COVID-19 Tissue Injury and Risk of Death and Can Cause Tissue Injury. JCI Insight 2021, 6, e147610. [Google Scholar] [CrossRef]
- Breitbach, S.; Tug, S.; Simon, P. Circulating Cell-Free DNA: An up-Coming Molecular Marker in Exercise Physiology. Sports Med. 2012, 42, 565–586. [Google Scholar] [CrossRef]
- Madsen, A.T.; Hojbjerg, J.A.; Sorensen, B.S.; Winther-Larsen, A. Day-to-Day and within-Day Biological Variation of Cell-Free DNA. EBioMedicine 2019, 49, 284–290. [Google Scholar] [CrossRef]
- Neuberger, E.W.I.; Brahmer, A.; Ehlert, T.; Kluge, K.; Philippi, K.F.A.; Boedecker, S.C.; Weinmann-Menke, J.; Simon, P. Validating Quantitative PCR Assays for CfDNA Detection without DNA Extraction in Exercising SLE Patients. Sci. Rep. 2021, 11, 13581. [Google Scholar] [CrossRef]
- Wittchen, H.U.; Zaudig, M.; Fydrich, T. Strukturiertes Klinisches Interview Für DSM-IV; Hogrefe: Göttingen, Germany, 1997. [Google Scholar]
- APA. Diagnostic and Statistical Manual of Mental Disorders, DSM-IV-TR, 4th ed.; American Psychiatric Association: Washington, DC, USA, 2000. [Google Scholar]
- Alghofaili, L.; Almubarak, H.; Gassem, K.; Islam, S.S.; Coskun, S.; Kaya, N.; Karakas, B. Cell-Free DNA Levels of Twins and Sibling Pairs Indicate Individuality and Possible Use as a Personalized Biomarker. PLoS ONE 2019, 14, e0223470. [Google Scholar] [CrossRef]
- Vasilyeva, I.N.; Bespalov, V.G.; Von, J.D.; Semenov, A.L.; Tochilnikov, G.V.; Romanov, V.A.; Alvovsky, I.K.; Baranenko, D.A. Cell-Free DNA Plasma Levels Differ in Age-Specific Pattern in Healthy Rats and Castrates with Testosterone-Induced Benign Prostatic Hyperplasia. Int. J. Genom. 2019, 2019, 8173630. [Google Scholar] [CrossRef]
- Kirschbaum, C.; Pirke, K.M.; Hellhammer, D.H. The ’Trier Social Stress Test’—A Tool for Investigating Psychobiological Stress Responses in a Laboratory Setting. Neuropsychobiology 1993, 28, 76–81. [Google Scholar] [CrossRef]
- Gaab, J. PASA—Primary Appraisal Secondary Appraisal. Ein Fragebogen Zur Erfassung von Situationsbezogenen Kognitiven Bewertungen. Verhaltenstherapie 2009, 19, 114–115. [Google Scholar] [CrossRef]
- Franke, G.H. Die Symptom-Checkliste von Derogatis (SCL-90-R)—Deutsche Version—Manual, 2nd ed.; Beltz Test: Göttingen, Germany, 2002. [Google Scholar]
- Schneider, E.E.; Schönfelder, S.; Domke-Wolf, M.; Wessa, M. Measuring Stress in Clinical and Nonclinical Subjects Using a German Adaptation of the Perceived Stress Scale. Int. J. Clin. Health Psychol. 2020, 20, 173–181. [Google Scholar] [CrossRef]
- Hautzinger, M.; Keller, F.; Kühner, C. Beck-Depressions-Inventar, BDI II.Revision; Harcourt Test Services: Frankfurt am Main, Germany, 2006. [Google Scholar]
- Schulz, P.; Schlotz, W.; Becker, P. Trierer Inventar Zum Chronischen Stress (TICS); Hogrefe: Göttingen, Germany, 2004. [Google Scholar]
- Frey, I.; Berg, A.; Grathwohl, D.; Keul, J. Freiburger Fragebogen Zur Körperlichen Aktivität-Entwicklung, Prüfung und Anwendung. Soz. Präventivmedizin SPM 1999, 44, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A Flexible Statistical Power Analysis Program for the Social, Behavioral, and Biomedical Sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Schlotz, W. Ambulatory Psychoneuroendocrinology: Assessing Salivary Cortisol and Other Hormones in Daily Life. In Handbook of Research Methods for Studying Daily Life; Mehl, M.R., Conner, T.S., Eds.; Guilford Press: New York, NY, USA, 2013; pp. 193–209. [Google Scholar]
- Meddeb, R.; Dache, Z.A.A.; Thezenas, S.; Otandault, A.; Tanos, R.; Pastor, B.; Sanchez, C.; Azzi, J.; Tousch, G.; Azan, S.; et al. Quantifying Circulating Cell-Free DNA in Humans. Sci. Rep. 2019, 9, 5220. [Google Scholar] [CrossRef] [PubMed]
- Trumpff, C.; Marsland, A.L.; Basualto-Alarcón, C.; Martin, J.L.; Carroll, J.E.; Sturm, G.; Vincent, A.E.; Mosharov, E.V.; Gu, Z.; Kaufman, B.A.; et al. Acute Psychological Stress Increases Serum Circulating Cell-Free Mitochondrial DNA. Psychoneuroendocrinology 2019, 106, 268–276. [Google Scholar] [CrossRef]
- Frattini, M.; Gallino, G.; Signoroni, S.; Balestra, D.; Battaglia, L.; Sozzi, G.; Leo, E.; Pilotti, S.; Pierotti, M.A. Quantitative Analysis of Plasma DNA in Colorectal Cancer Patients: A Novel Prognostic Tool. Ann. N. Y. Acad. Sci. 2006, 1075, 185–190. [Google Scholar] [CrossRef]
- al Amir Dache, Z.; Otandault, A.; Tanos, R.; Pastor, B.; Meddeb, R.; Sanchez, C.; Arena, G.; Lasorsa, L.; Bennett, A.; Grange, T.; et al. Blood Contains Circulating Cell-free Respiratory Competent Mitochondria. FASEB J. 2020, 34, 3616–3630. [Google Scholar] [CrossRef] [PubMed]
- Hellhammer, D.H.; Wüst, S.; Kudielka, B.M. Salivary Cortisol as a Biomarker in Stress Research. Psychoneuroendocrinology 2009, 34, 163–171. [Google Scholar] [CrossRef]
- Schneider, M.; Kraemmer, M.M.; Weber, B.; Schwerdtfeger, A.R. Life Events Are Associated with Elevated Heart Rate and Reduced Heart Complexity to Acute Psychological Stress. Biol. Psychol. 2021, 163, 108116. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.D. Neuroendocrine Interactions in the Immune System. Cell Immunol. 2008, 252, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bucsek, M.J.; Giridharan, T.; MacDonald, C.R.; Hylander, B.L.; Repasky, E.A. An Overview of the Role of Sympathetic Regulation of Immune Responses in Infectious Disease and Autoimmunity. Int. J. Hyperth. 2018, 34, 135–143. [Google Scholar] [CrossRef]
- Korabecna, M.; Zinkova, A.; Brynychova, I.; Chylikova, B.; Prikryl, P.; Sedova, L.; Neuzil, P.; Seda, O. Cell-Free DNA in Plasma as an Essential Immune System Regulator. Sci. Rep. 2020, 10, 17478. [Google Scholar] [CrossRef]
- Zorn, J.V.; Schür, R.R.; Boks, M.P.; Kahn, R.S.; Joëls, M.; Vinkers, C.H. Cortisol Stress Reactivity across Psychiatric Disorders: A Systematic Review and Meta-Analysis. Psychoneuroendocrinology 2017, 77, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Petrowski, K.; Wintermann, G.-B.; Schaarschmidt, M.; Bornstein, S.R.; Kirschbaum, C. Blunted Salivary and Plasma Cortisol Response in Patients with Panic Disorder under Psychosocial Stress. Int. J. Psychophysiol. 2013, 88, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Beck, C.R.; Collier, P.; Macfarlane, C.; Malig, M.; Kidd, J.M.; Eichler, E.E.; Badge, R.M.; Moran, J.v. LINE-1 Retrotransposition Activity in Human Genomes. Cell 2010, 141, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Sultana, T.; Van Essen, D.; Siol, O.; Bailly-Bechet, M.; Philippe, C.; Zine El Aabidine, A.; Pioger, L.; Nigumann, P.; Saccani, S.; Andrau, J.-C.; et al. The Landscape of L1 Retrotransposons in the Human Genome Is Shaped by Pre-Insertion Sequence Biases and Post-Insertion Selection. Mol. Cell 2019, 74, 555–570.e7. [Google Scholar] [CrossRef] [PubMed]
- Cuarenta, A.; Kigar, S.L.; Henion, I.C.; Chang, L.; Bakshi, V.P.; Auger, A.P. Early Life Stress during the Neonatal Period Alters Social Play and Line1 during the Juvenile Stage of Development. Sci. Rep. 2021, 11, 3549. [Google Scholar] [CrossRef]
- Liu, S.; Du, T.; Liu, Z.; Shen, Y.; Xiu, J.; Xu, Q. Inverse Changes in L1 Retrotransposons between Blood and Brain in Major Depressive Disorder. Sci. Rep. 2016, 6, 37530. [Google Scholar] [CrossRef]
- Terry, D.M.; Devine, S.E. Aberrantly High Levels of Somatic LINE-1 Expression and Retrotransposition in Human Neurological Disorders. Front. Genet. 2020, 10, 1244. [Google Scholar] [CrossRef] [PubMed]
- Simon, M.; van Meter, M.; Ablaeva, J.; Ke, Z.; Gonzalez, R.S.; Taguchi, T.; De Cecco, M.; Leonova, K.I.; Kogan, V.; Helfand, S.L.; et al. LINE1 Derepression in Aged Wild-Type and SIRT6-Deficient Mice Drives Inflammation. Cell Metab. 2019, 29, 871–885.e5. [Google Scholar] [CrossRef]
- Neuberger, E.W.I.; Sontag, S.; Brahmer, A.; Philippi, K.F.A.; Radsak, M.P.; Wagner, W.; Simon, P. Physical Activity Specifically Evokes Release of Cell-Free DNA from Granulocytes Thereby Affecting Liquid Biopsy. Clin. Epigenetics 2022, 14, 29. [Google Scholar] [CrossRef]
- Beiter, T.; Fragasso, A.; Hartl, D.; Nieß, A.M. Neutrophil Extracellular Traps: A Walk on the Wild Side of Exercise Immunology. Sports Med. 2015, 45, 625–640. [Google Scholar] [CrossRef] [PubMed]
- Neuberger, E.W.I.; Hillen, B.; Mayr, K.; Simon, P.; Krämer-Albers, E.M.; Brahmer, A. Kinetics and Topology of DNA Associated with Circulating Extracellular Vesicles Released during Exercise. Genes 2021, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Helmig, S.; Frühbeis, C.; Krämer-Albers, E.M.; Simon, P.; Tug, S. Release of Bulk Cell Free DNA during Physical Exercise Occurs Independent of Extracellular Vesicles. Eur. J. Appl. Physiol. 2015, 115, 2271–2280. [Google Scholar] [CrossRef]
- van der Meer, A.J.; Kroeze, A.; Hoogendijk, A.J.; Soussan, A.A.; van der Schoot, C.E.; Wuillemin, W.A.; Voermans, C.; Van der Poll, T.; Zeerleder, S. Systemic Inflammation Induces Release of Cell-Free DNA from Hematopoietic and Parenchymal Cells in Mice and Humans. Blood Adv. 2019, 3, 724–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individuals (n = 29) | |
|---|---|
| Demographic data | |
| Age (years), M (SD), [Min–Max] | 24.34 (4.08), [19–34] |
| Body mass index, M (SD), [Min–Max] | 22.94 (1.61), [19.94–26.32] |
| Smoking, n (%) | 3 (10.4) |
| FFKA—Total activity (min/week), M (SD), [Min–Max] | 1534 (1047), [246–4230] |
| FFKA—Sport activity (min/week), M (SD), [Min–Max] | 473 (583), [0–2700] |
| Psychological Assessment | |
| BDI, M (SD), [Min–Max] | 5.62 (4.67), [0–15] |
| SCL Global Severity Index, M (SD), [Min–Max] | 0.36 (0.29), [0.01–1.17] |
| PSS, M (SD), [Min–Max] | 22.24 (6.78), [7–36] |
| TICS-SCSS, M (SD), [Min–Max] | 12.10 (6.26), [3–28] |
| Healthy Men (n = 29) | |||||||
|---|---|---|---|---|---|---|---|
| Resting Condition | TSST | Dependent t-Test | |||||
| Derived cfDNA Parameters | M | SD | M | SD | t | p | |
| Peak | 14.25 | 4.58 | 17.92 | 7.08 | −2.914 | ≤0.01 ** (d = −0.54) | |
| Delta Peak-Baseline | 4.75 | 2.92 | 9.23 | 5.10 | −4.192 | ≤0.001 *** (d = −0.78) | |
| AUCI | 102.17 | 260.75 | 306.70 | 198.08 | −3.712 | ≤0.001 ** (d = −0.69) | |
| Derived cortisol parameters | |||||||
| Peak a) | 63.09 | 17.90 | 94.34 | 23.26 | −7.046 | ≤0.001 *** (d = −1.33) | |
| Delta Peak-Baseline a) | 10.74 | 8.69 | 42.35 | 25.87 | −6.471 | ≤0.001 *** (d = −1.22) | |
| AUCI a) | −1168.22 | 1299.72 | 930.13 | 2457.85 | −3.927 | ≤0.001 *** (d = −0.74) | |
| Derived heart rate parameters | |||||||
| Peak b) | 70.44 | 11.64 | 100.59 | 18.85 | −7.584 | ≤0.001 *** (d = −1.58) | |
| Delta Peak-Baseline b) | 0.88 | 3.20 | 31.04 | 20.20 | −7.584 | ≤0.001 *** (d = −1.58) | |
| Subjective Appraisal | |||||||
| PASA | Threat, (1–6) | 1.47 | 0.83 | 3.09 | 1.08 | −7.855 | ≤0.001 *** (d = −1.46) |
| Challenge, (1–6) | 2.35 | 0.96 | 4.07 | 0.79 | −8.100 | ≤0.001 *** (d = −1.50) | |
| Self concept, (1–6) | 4.55 | 0.89 | 3.91 | 1.02 | 2.660 | ≤0.05 * (d = 0.49) | |
| Control expectancy, (1–6) | 4.63 | 0.73 | 4.31 | 0.84 | 2.002 | 0.055 (d = 0.49) | |
| Primary appraisal, (1–6) | 1.91 | 0.79 | 3.58 | 0.82 | −9.529 | ≤0.001 *** (d = −1.77) | |
| Secondary appraisal, (1–6) | 4.59 | 0.64 | 4.11 | 0.79 | 3.143 | ≤0.001 *** (d = 0.58) | |
| PASA—Stress index | −2.68 | 1.28 | −0.54 | 1.39 | −7.663 | ≤0.001 *** (d = −1.41) | |
| VAS | 41.35 | 11.23 | 58.51 | 9.42 | −8.903 | ≤0.001 *** (d = −1.65) | |
| ln Baseline cfDNA | cfDNA AUCI | ln Baseline Cortisol a) | Cortisol AUCI a) | Psychological Measures | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Resting Condition | Stress Condition | Stress Condition | Resting Condition | Stress Condition | Stress Condition | BDI | SCL-GSI | PSS | TICS-SCSS | ||
| ln baseline cfDNA | Resting condition | 1.00 | |||||||||
| Stress condition | 0.63, p ≤ 0.001 | 1.00 | |||||||||
| cfDNA AUCI | Stress condition | 0.23, p = 0.23 | 0.41, p ≤ 0.05 | 1.00 | |||||||
| ln baseline cortisol a) | Resting condition | 0.13, p = 0.52 | 0.09, p = 0.66 | −0.04, p = 0.83 | 1.00 | ||||||
| Stress condition | 0.07, p = 0.74 | 0.15, p = 0.44 | 0.11, p = 0.58 | 0.28, p = 0.15 | 1.00 | ||||||
| cortisol AUCI a) | Stress condition | −0.01, p = 0.95 | 0.06, p = 0.75 | 0.18, p = 0.35 | 0.11, p = 0.57 | 0.59, p ≤ 0.01 | 1.00 | ||||
| Psycho-logical measures | BDI | −0.01, p = 0.95 | −0.21, p = 0.28 | −0.16, p = 0.41 | 0.24, p = 0.23 | 0.00, p = 0.99 | −0.26, p = 0.18 | 1.00 | |||
| SCL-GSI | 0.01, p = 0.95 | −0.15, p = 0.43 | −0.10, p = 0.60 | 0.19, p = 0.33 | 0.06, p = 0.76 | −0.22, p = 0.26 | 0.74, p ≤ 0.001 | 1.00 | |||
| PSS | 0.07, p = 0.73 | 0.27, p = 0.15 | 0.04, p = 0.85 | −0.04, p = 0.85 | 0.19, p = 0.33 | −0.34, p = 0.08 | 0.47, p ≤ 0.01 | 0.46, ≤ 0.05 | 1.00 | ||
| TICS-SCSS | −0.18, p = 0.35 | −0.01, p = 0.97 | 0.16, p = 0.40 | −0.23, p = 0.25 | 0.12, p = 0.54 | −0.21, p = 0.28 | 0.27, p = 0.17 | 0.49, p ≤ 0.01 | 0.65, p ≤ 0.001 | 1.00 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herhaus, B.; Neuberger, E.; Juškevičiūtė, E.; Simon, P.; Petrowski, K. Kinetics of Plasma Cell-Free DNA under a Highly Standardized and Controlled Stress Induction. Cells 2023, 12, 564. https://doi.org/10.3390/cells12040564
Herhaus B, Neuberger E, Juškevičiūtė E, Simon P, Petrowski K. Kinetics of Plasma Cell-Free DNA under a Highly Standardized and Controlled Stress Induction. Cells. 2023; 12(4):564. https://doi.org/10.3390/cells12040564
Chicago/Turabian StyleHerhaus, Benedict, Elmo Neuberger, Ema Juškevičiūtė, Perikles Simon, and Katja Petrowski. 2023. "Kinetics of Plasma Cell-Free DNA under a Highly Standardized and Controlled Stress Induction" Cells 12, no. 4: 564. https://doi.org/10.3390/cells12040564
APA StyleHerhaus, B., Neuberger, E., Juškevičiūtė, E., Simon, P., & Petrowski, K. (2023). Kinetics of Plasma Cell-Free DNA under a Highly Standardized and Controlled Stress Induction. Cells, 12(4), 564. https://doi.org/10.3390/cells12040564