
ELAC2 Functions as a Key Gene in the Early Development of Placental Formation Based on WGCNA


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Placental Target Genes
2.2. Animal Treatments
2.3. RNA Extraction and RT-qPCR
2.4. Cell Culture and Transfection
2.5. Western Blot Analysis
2.6. Wound Healing Assay
2.7. Transwell Cell Migration Assay
2.8. EdU Cell Proliferation Assay
2.9. Statistics
3. Results
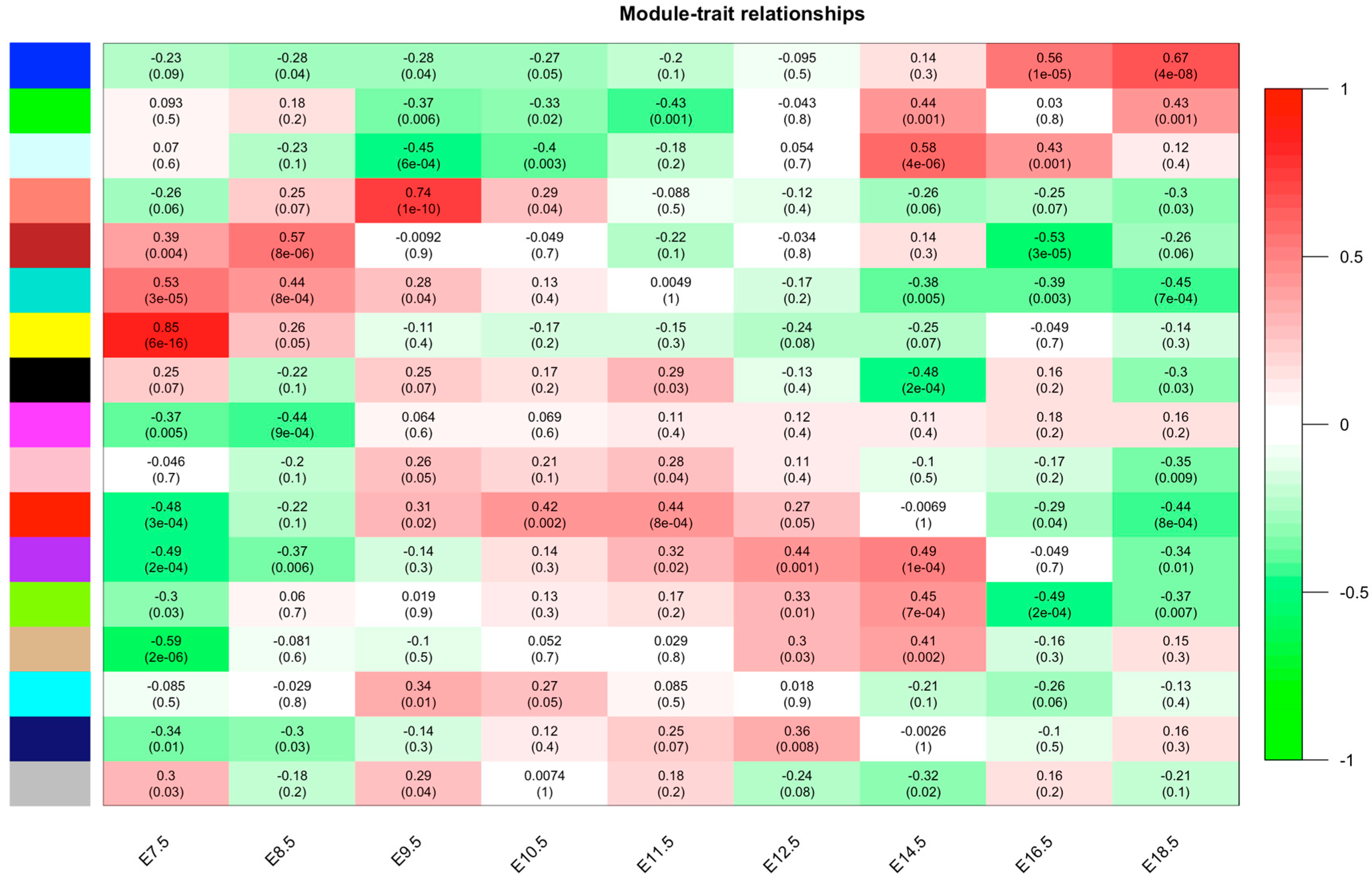
3.1. Construction of the Co-Expression Network
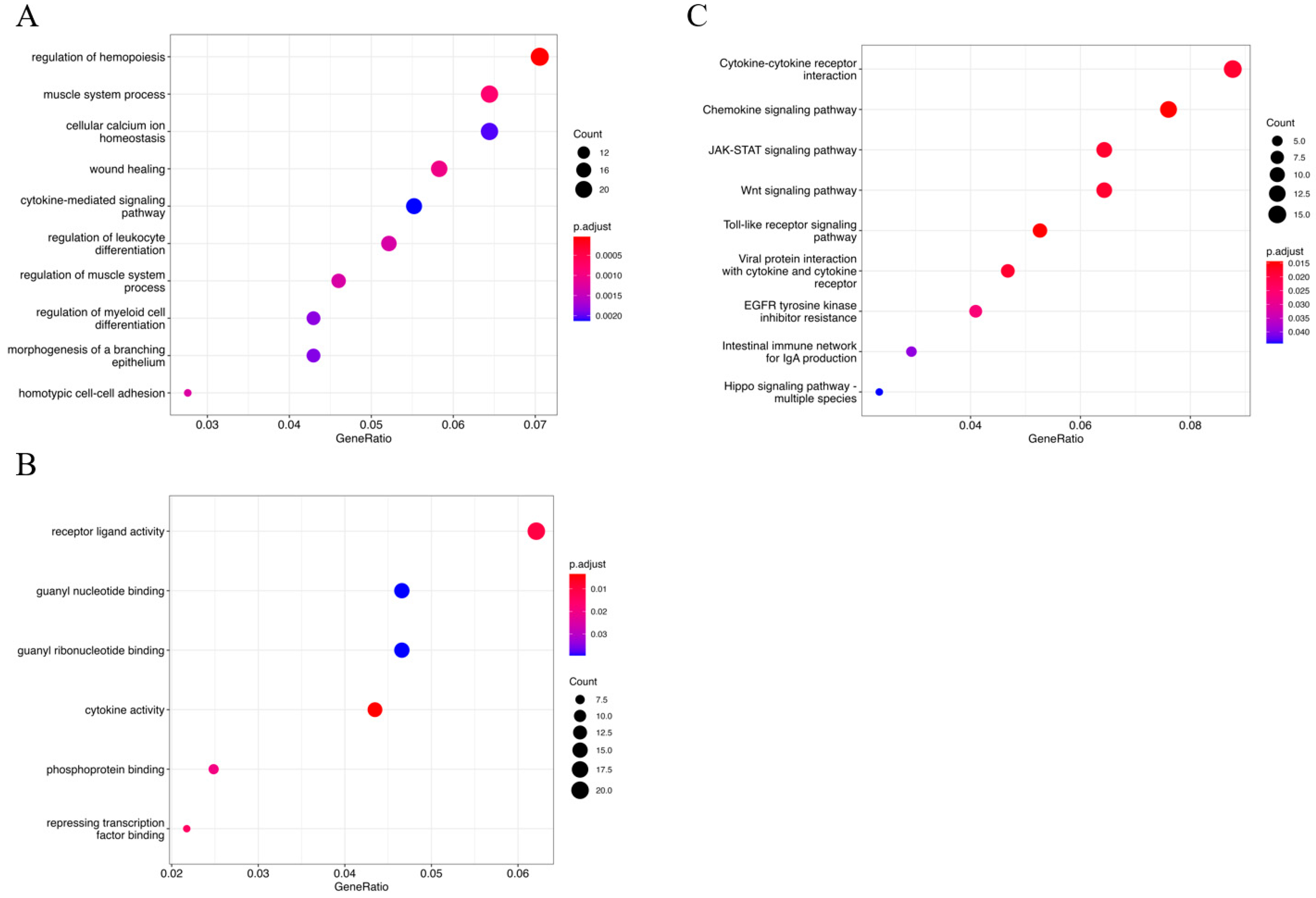
3.2. Functional Analysis of the Selected Genes
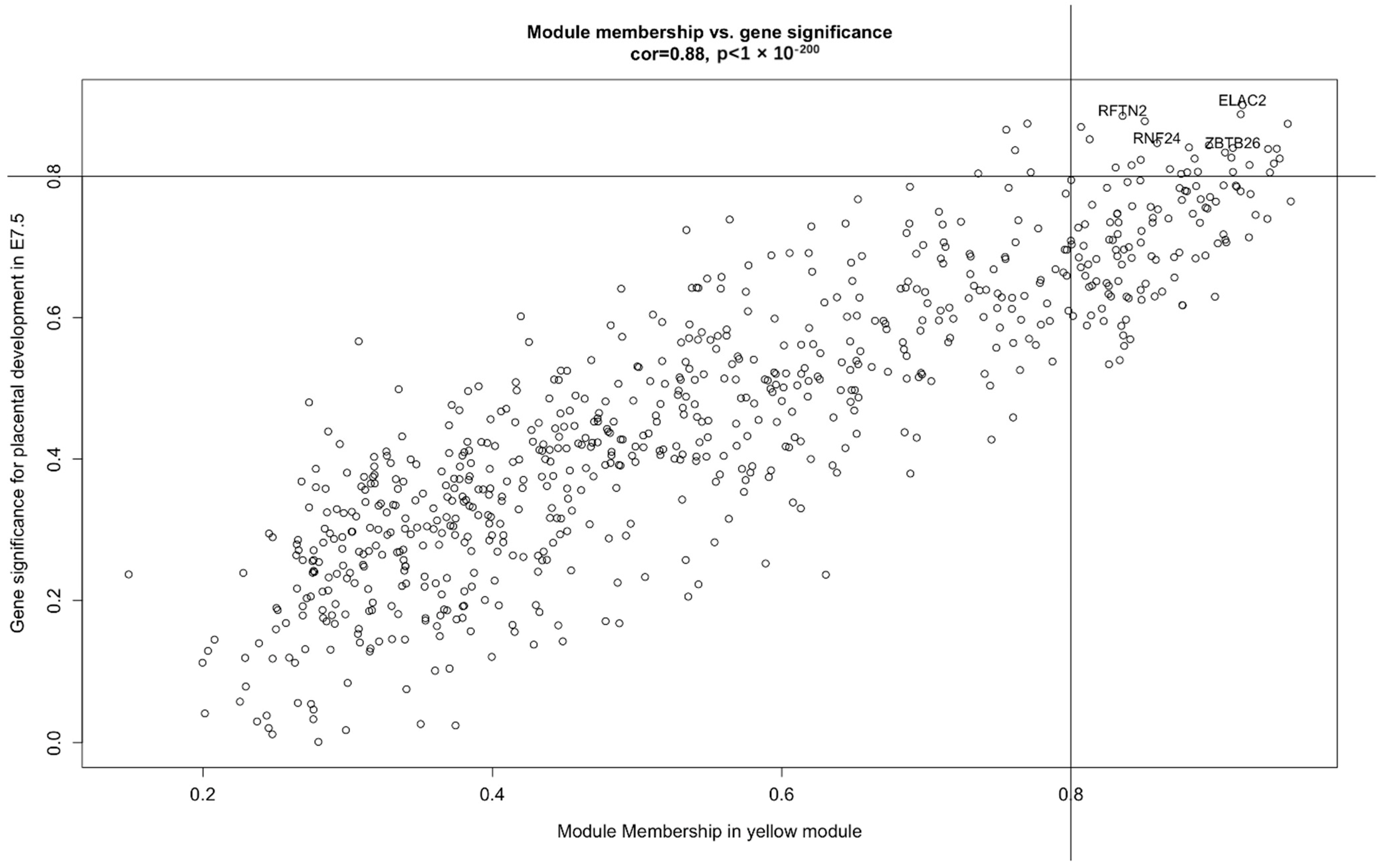
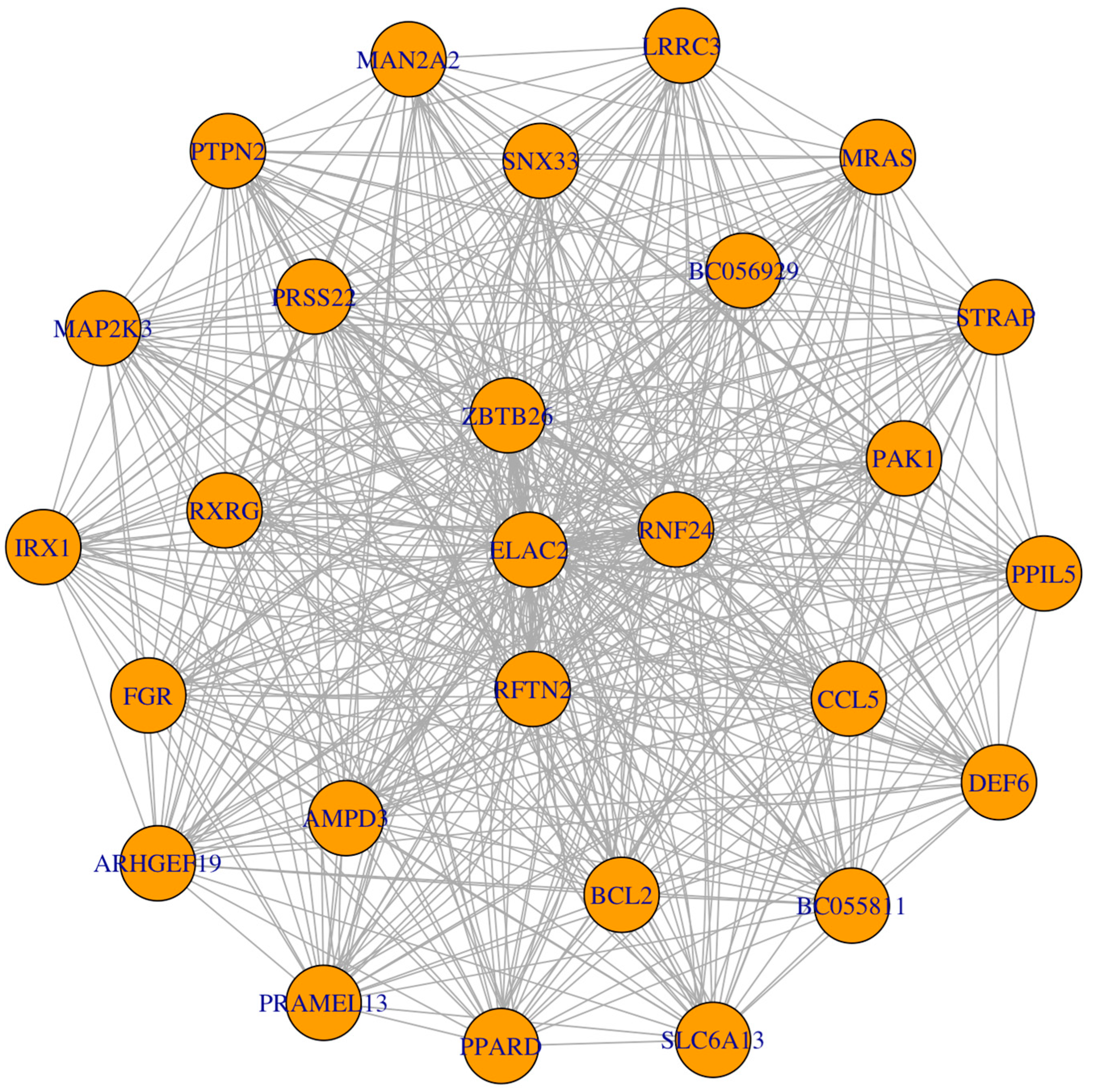
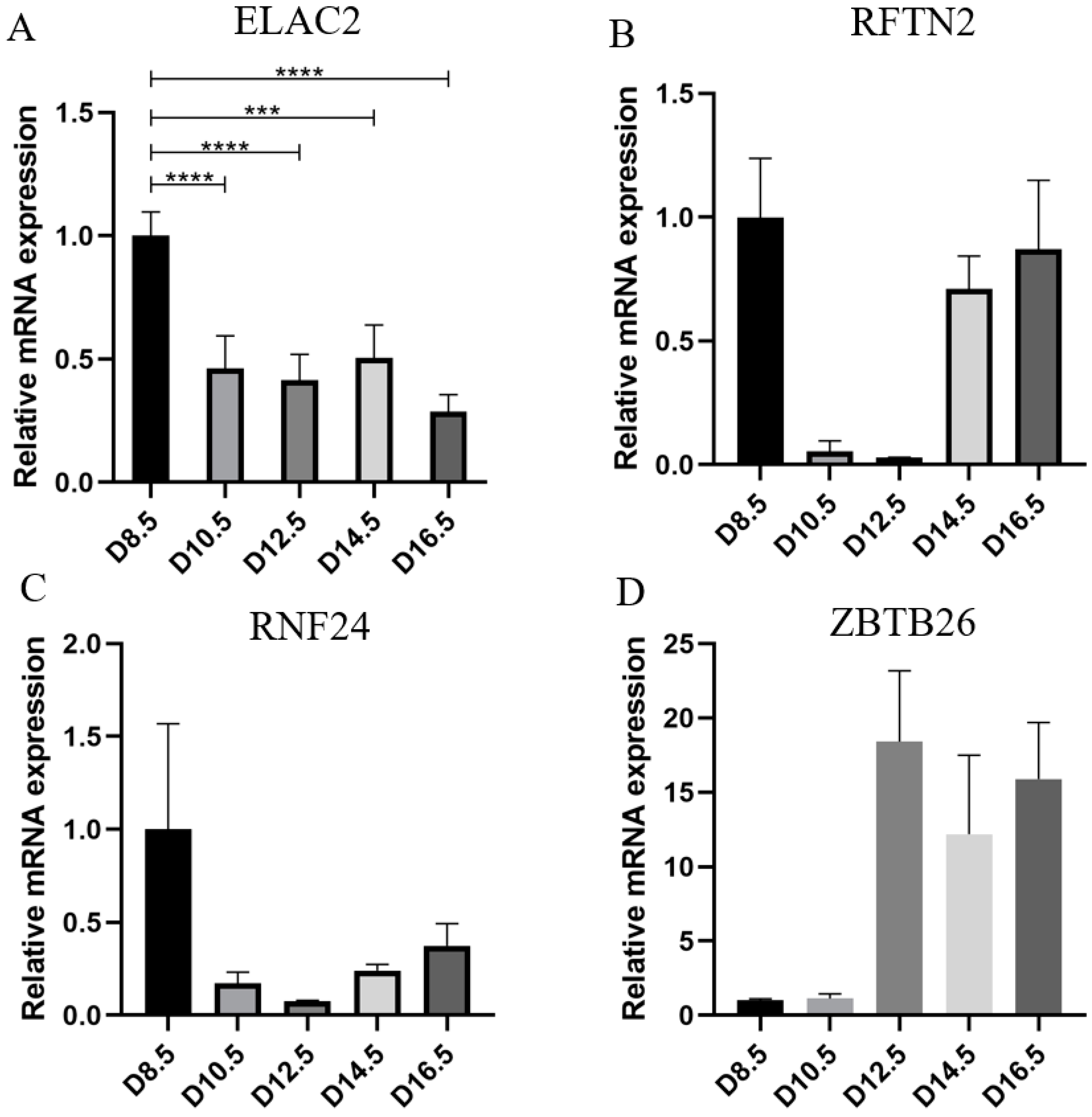
3.3. ELAC2 Was Screened as a Key Gene for Placental Development
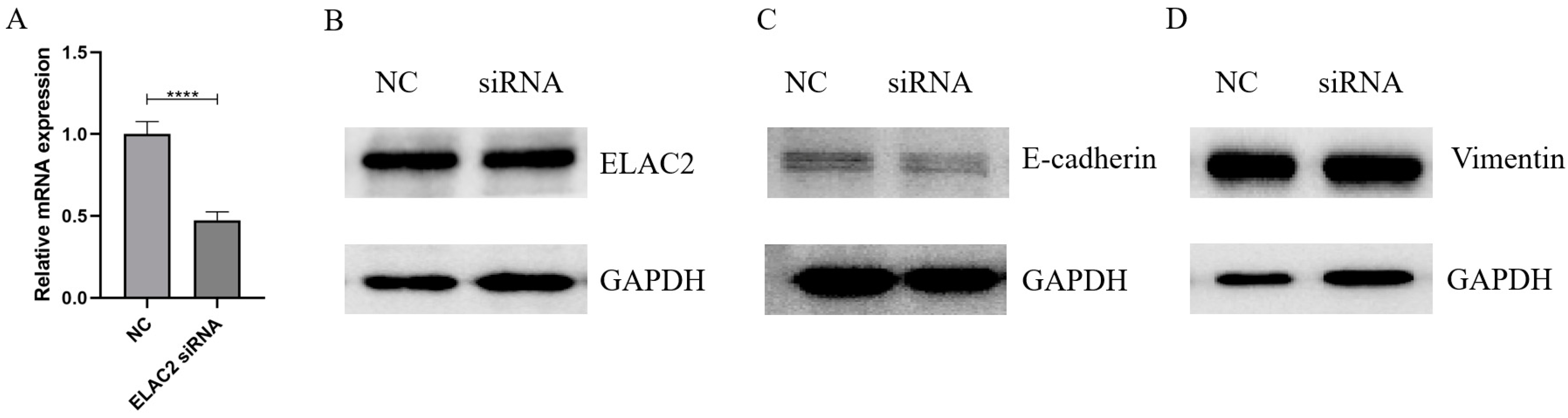
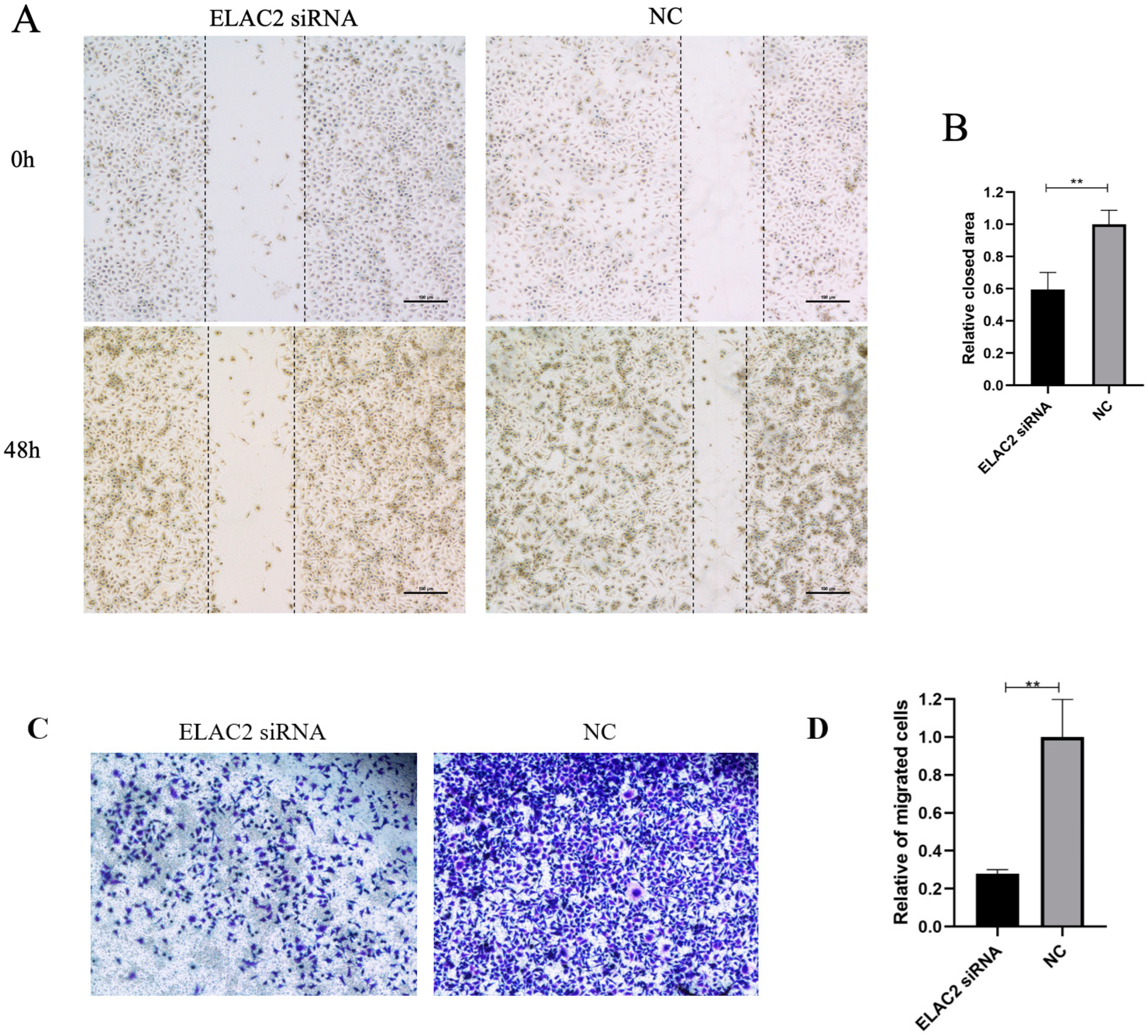
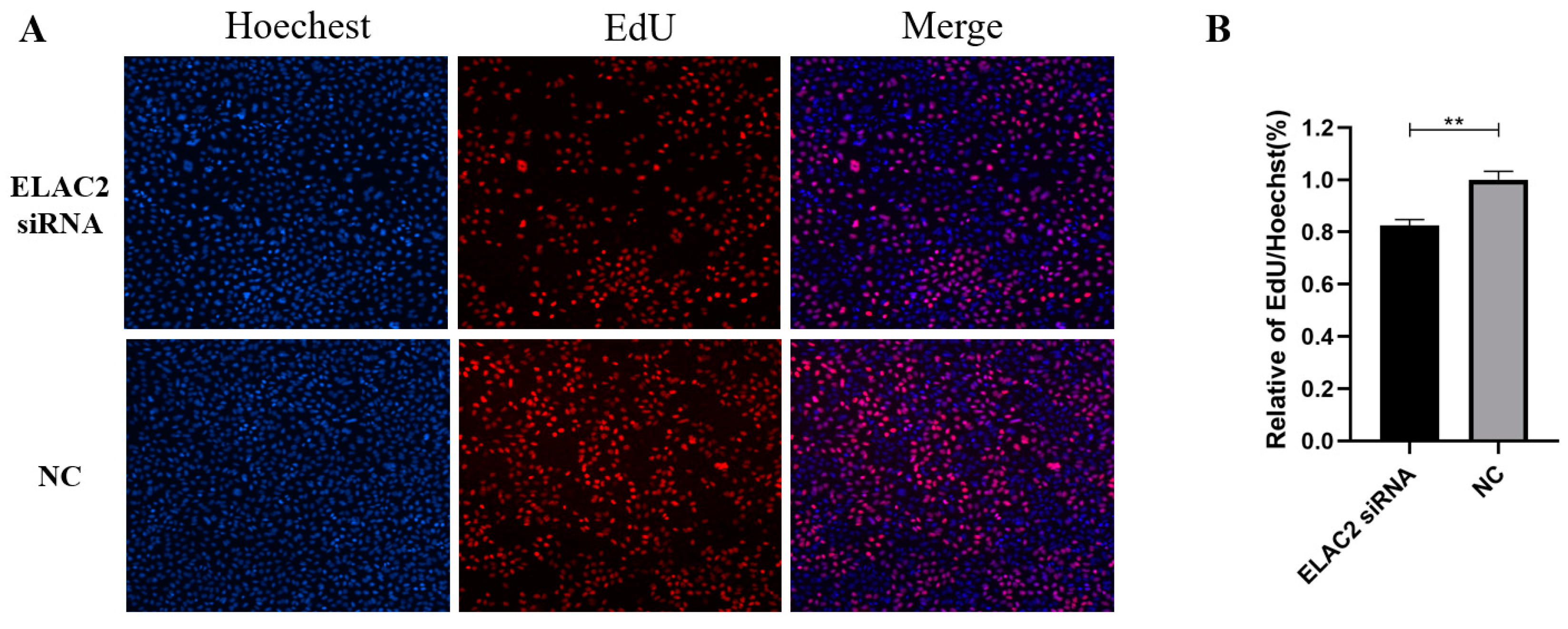
3.4. Cell Migration and Cell Proliferation Are Reduced When ELAC2 Was Downregulated
3.5. Interfering with ELAC2 Expression Possibly Promotes Epithelial Mesenchymal Transition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maltepe, E.; Bakardjiev, A.I.; Fisher, S.J. The Placenta: Transcriptional, Epigenetic, and Physiological Integration during Development. J. Clin. Investig. 2010, 120, 1016–1025. [Google Scholar] [CrossRef]
- Dilworth, M.R.; Sibley, C.P. Review: Transport across the Placenta of Mice and Women. Placenta 2013, 34, S34–S39. [Google Scholar] [CrossRef] [PubMed]
- Rossant, J.; Cross, J.C. Placental Development: Lessons from Mouse Mutants. Nat. Rev. Genet. 2001, 2, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Perez-Garcia, V.; Fineberg, E.; Wilson, R.; Murray, A.; Mazzeo, C.I.; Tudor, C.; Sienerth, A.; White, J.K.; Tuck, E.; Ryder, E.J.; et al. Placentation Defects Are Highly Prevalent in Embryonic Lethal Mouse Mutants. Nature 2018, 555, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Brosens, I.; Pijnenborg, R.; Vercruysse, L.; Romero, R. The “Great Obstetrical Syndromes” Are Associated with Disorders of Deep Placentation. Am. J. Obstet. Gynecol. 2011, 204, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.; Brkić, J.; Liu, M.; Fu, G.; Peng, C.; Wang, Y.-L. Placental Trophoblast Cell Differentiation: Physiological Regulation and Pathological Relevance to Preeclampsia. Mol. Asp. Med. 2013, 34, 981–1023. [Google Scholar] [CrossRef]
- Odibo, A.O.; Zhong, Y.; Longtine, M.; Tuuli, M.; Odibo, L.; Cahill, A.G.; Macones, G.A.; Nelson, D.M. First-Trimester Serum Analytes, Biophysical Tests and the Association with Pathological Morphometry in the Placenta of Pregnancies with Preeclampsia and Fetal Growth Restriction. Placenta 2011, 32, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Massin, N.; Frendo, J.-L.; Guibourdenche, J.; Luton, D.; Giovangrandi, Y.; Muller, F.; Vidaud, M.; Evain-Brion, D. Defect of Syncytiotrophoblast Formation and Human Chorionic Gonadotropin Expression in Down’s Syndrome. Placenta 2001, 22, S93–S97. [Google Scholar] [CrossRef]
- Strumpf, D.; Mao, C.-A.; Yamanaka, Y.; Ralston, A.; Chawengsaksophak, K.; Beck, F.; Rossant, J. Cdx2 Is Required for Correct Cell Fate Specification and Differentiation of Trophectoderm in the Mouse Blastocyst. Development 2005, 132, 2093–2102. [Google Scholar] [CrossRef]
- Saha, B.; Ganguly, A.; Home, P.; Bhattacharya, B.; Ray, S.; Ghosh, A.; Rumi, M.A.K.; Marsh, C.; French, V.A.; Gunewardena, S.; et al. TEAD4 Ensures Postimplantation Development by Promoting Trophoblast Self-Renewal: An Implication in Early Human Pregnancy Loss. Proc. Natl. Acad. Sci. USA 2020, 117, 17864–17875. [Google Scholar] [CrossRef] [PubMed]
- Knöfler, M.; Haider, S.; Saleh, L.; Pollheimer, J.; Gamage, T.K.J.B.; James, J. Human Placenta and Trophoblast Development: Key Molecular Mechanisms and Model Systems. Cell. Mol. Life Sci. 2019, 76, 3479–3496. [Google Scholar] [CrossRef] [PubMed]
- Latos, P.A.; Goncalves, A.; Oxley, D.; Mohammed, H.; Turro, E.; Hemberger, M. Fgf and Esrrb Integrate Epigenetic and Transcriptional Networks That Regulate Self-Renewal of Trophoblast Stem Cells. Nat. Commun. 2015, 6, 7776. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Nikaido, I.; Ohta, H.; Ohtsuka, S.; Ura, H.; Kadota, M.; Wakayama, T.; Ueda, H.R.; Niwa, H. Context-Dependent Wiring of Sox2 Regulatory Networks for Self-Renewal of Embryonic and Trophoblast Stem Cells. Mol. Cell 2013, 52, 380–392. [Google Scholar] [CrossRef]
- Morasso, M.I.; Grinberg, A.; Robinson, G.; Sargent, T.D.; Mahon, K.A. Placental Failure in Mice Lacking the Homeobox Gene Dlx 3. Proc. Natl. Acad. Sci. USA 1999, 96, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R Package for Weighted Correlation Network Analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, D.; Lei, C.; Li, Y.; Sun, X.; Chen, M.; Wu, X.; Zheng, Y. Identification of Crucial Genes in Abdominal Aortic Aneurysm by WGCNA. PeerJ 2019, 7, e7873. [Google Scholar] [CrossRef]
- Guo, Y.; Ma, J.; Xiao, L.; Fang, J.; Li, G.; Zhang, L.; Xu, L.; Lai, X.; Pan, G.; Chen, Z. Identification of Key Pathways and Genes in Different Types of Chronic Kidney Disease Based on WGCNA. Mol. Med. Rep. 2019, 20, 2245–2257. [Google Scholar] [CrossRef]
- Yin, L.; Cai, Z.; Zhu, B.; Xu, C. Identification of Key Pathways and Genes in the Dynamic Progression of HCC Based on WGCNA. Genes 2018, 9, 92. [Google Scholar] [CrossRef]
- Noda, D.; Itoh, S.; Watanabe, Y.; Inamitsu, M.; Dennler, S.; Itoh, F.; Koike, S.; Danielpour, D. ELAC2, a Putative Prostate Cancer Susceptibility Gene Product, Potentiates TGF-b/Smad-Induced Growth Arrest of Prostate Cells. Oncogene 2006, 25, 5591–5600. [Google Scholar] [CrossRef] [PubMed]
- Soncin, F.; Khater, M.; To, C.; Pizzo, D.; Farah, O.; Wakeland, A.; Rajan, K.A.N.; Nelson, K.K.; Chang, C.-W.; Moretto-Zita, M.; et al. Comparative Analysis of Mouse and Human Placentae across Gestation Reveals Species-Specific Regulators of Placental Development. Development 2018, 146, 156273. [Google Scholar] [CrossRef] [Green Version]
- Hemberger, M.; Hanna, C.W.; Dean, W. Mechanisms of Early Placental Development in Mouse and Humans. Nat. Rev. Genet. 2020, 21, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Graham, C.H.; Hawley, T.S.; Hawley, R.G.; MacDougall, J.R.; Kerbel, R.S.; Khoo, N.; Lala, P.K. Establishment and characterization of first trimester human trophoblast cells with extended lifespan. Exp. Cell Res. 1993, 206, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Janssens, K.; Sung, H.-H.; Rørth, P. Direct Detection of Guidance Receptor Activity during Border Cell Migration. Proc. Natl. Acad. Sci. USA 2010, 107, 7323–7328. [Google Scholar] [CrossRef] [PubMed]
- Herrera, S.C.; Bach, E.A. JAK/STAT Signaling in Stem Cells and Regeneration: From Drosophila to Vertebrates. Development 2019, 146, dev167643. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Loh, K.M.; Nusse, R. An Integral Program for Tissue Renewal and Regeneration: Wnt Signaling and Stem Cell Control. Science 2014, 6205, 54–70. [Google Scholar] [CrossRef]
- Han, H. RNA Interference to Knock Down Gene Expression. In Disease Gene Identification; DiStefano, J.K., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; Volume 1706, pp. 293–302. [Google Scholar] [CrossRef]
- Red-Horse, K.; Zhou, Y.; Genbacev, O.; Prakobphol, A.; Foulk, R.; McMaster, M.; Fisher, S.J. Trophoblast Differentiation during Embryo Implantation and Formation of the Maternal-Fetal Interface. J. Clin. Investig. 2004, 114, 744–754. [Google Scholar] [CrossRef]
- Burton, G.J.; Charnock-Jones, D.S.; Jauniaux, E. Regulation of Vascular Growth and Function in the Human Placenta. Reproduction 2009, 138, 895–902. [Google Scholar] [CrossRef]
- Kalluri, R.; Weinberg, R.A. The Basics of Epithelial-Mesenchymal Transition. J. Clin. Invest. 2009, 119, 1420–1428. [Google Scholar] [CrossRef]
- Wang, Z.; Divanyan, A.; Jourd’heuil, F.L.; Goldman, R.D.; Ridge, K.M.; Jourd’heuil, D.; Lopez-Soler, R.I. Vimentin Expression Is Required for the Development of EMT-Related Renal Fibrosis Following Unilateral Ureteral Obstruction in Mice. Am. J. Physiol.-Ren. Physiol. 2018, 315, F769–F780. [Google Scholar] [CrossRef] [Green Version]
- van Roy, F.; Berx, G. The Cell-Cell Adhesion Molecule E-Cadherin. Cell. Mol. Life Sci. 2008, 65, 3756–3788. [Google Scholar] [CrossRef]
- Cheng, F.; Eriksson, J.E. Intermediate Filaments and the Regulation of Cell Motility during Regeneration and Wound Healing. Cold Spring Harb. Perspect. Biol. 2017, 9, a022046. [Google Scholar] [CrossRef]
- Battaglia, R.A.; Delic, S.; Herrmann, H.; Snider, N.T. Vimentin on the Move: New Developments in Cell Migration. F1000Research 2018, 7, 1796. [Google Scholar] [CrossRef] [PubMed]
- Desai, R.A.; Gao, L.; Raghavan, S.; Liu, W.F.; Chen, C.S. Cell Polarity Triggered by Cell-Cell Adhesion via E-Cadherin. J. Cell Sci. 2009, 122, 905–911. [Google Scholar] [CrossRef]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a Knowledge-Based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef] [PubMed]
- Takaku, H. A Candidate Prostate Cancer Susceptibility Gene Encodes TRNA 3’ Processing Endoribonuclease. Nucleic Acids Res. 2003, 31, 2272–2278. [Google Scholar] [CrossRef]
- Korver, W.; Guevara, C.; Chen, Y.; Neuteboom, S.; Bookstein, R.; Tavtigian, S.; Lees, E. The Product of the Candidate Prostate Cancer Susceptibility GeneELAC2 Interacts with the -Tubulin Complex. Int. J. Cancer 2003, 104, 283–288. [Google Scholar] [CrossRef]
- Nashimoto, M. Distribution of Both Lengths and 5’ Terminal Nucleotides of Mammalian Pre-TRNA 3’ Trailers Reflects Properties of 3’ Processing Endoribonuclease. Nucleic Acids Res. 1997, 25, 1148–1154. [Google Scholar] [CrossRef]
- Lee, Y.S.; Shibata, Y.; Malhotra, A.; Dutta, A. A Novel Class of Small RNAs: TRNA-Derived RNA Fragments (TRFs). Genes Dev. 2009, 23, 2639–2649. [Google Scholar] [CrossRef]
- Morikawa, M.; Derynck, R.; Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harb. Perspect. Biol. 2016, 8, a021873. [Google Scholar] [CrossRef] [Green Version]
- Woods, L.; Perez-Garcia, V.; Hemberger, M. Regulation of Placental Development and Its Impact on Fetal Growth—New Insights from Mouse Models. Front. Endocrinol. 2018, 9, 570. [Google Scholar] [CrossRef]
- Bird, S.D. Artificial Placenta: Analysis of Recent Progress. Eur. J. Obstet. Gynecol. Reprod. Biol. 2017, 208, 61–70. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Primer Sequence (5′-3′) |
|---|---|
| GAPDH | F: AGGTCGGTGTGAACGGATTTG |
| R: TGTAGACCATGTAGTTGAGGTCA | |
| RFTN2 | F: ACCTAAAGGGGACCAGTTACC |
| R: ACAATGGCGTCGTTTCCTCTC | |
| RNF24 | F: TCCAGAATCTGCCTCTCAACA |
| R: CCAGTTCATCTCTAGGCTTGAAG | |
| ELAC2 | F: GAGAAGGCGTCCAACGACTTA |
| R: AGAAAGATGTTGTCCAAGCGAG | |
| ZBTB26 | F: TTGACTGCTGCGAGTTTTCTT |
| R: CTGCTGTTCTTTCGACTGGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, J.; Liang, J.; Tan, Q.; Wang, Z. ELAC2 Functions as a Key Gene in the Early Development of Placental Formation Based on WGCNA. Cells 2023, 12, 613. https://doi.org/10.3390/cells12040613
Liang J, Liang J, Tan Q, Wang Z. ELAC2 Functions as a Key Gene in the Early Development of Placental Formation Based on WGCNA. Cells. 2023; 12(4):613. https://doi.org/10.3390/cells12040613
Chicago/Turabian StyleLiang, Junyong, Jingjie Liang, Qiang Tan, and Zhengguang Wang. 2023. "ELAC2 Functions as a Key Gene in the Early Development of Placental Formation Based on WGCNA" Cells 12, no. 4: 613. https://doi.org/10.3390/cells12040613
APA StyleLiang, J., Liang, J., Tan, Q., & Wang, Z. (2023). ELAC2 Functions as a Key Gene in the Early Development of Placental Formation Based on WGCNA. Cells, 12(4), 613. https://doi.org/10.3390/cells12040613