
Low-Intensity Blue Light Exposure Reduces Melanopsin Expression in Intrinsically Photosensitive Retinal Ganglion Cells and Damages Mitochondria in Retinal Ganglion Cells in Wistar Rats


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Immunostaining
2.3. Counting of ipRGC Somas and Determination of Opn4-Positive Neural Process Lengths
2.4. Sholl Analysis
2.5. Electron Microscopy
2.6. TUNEL Assay
2.7. Statistical Analyses
3. Results
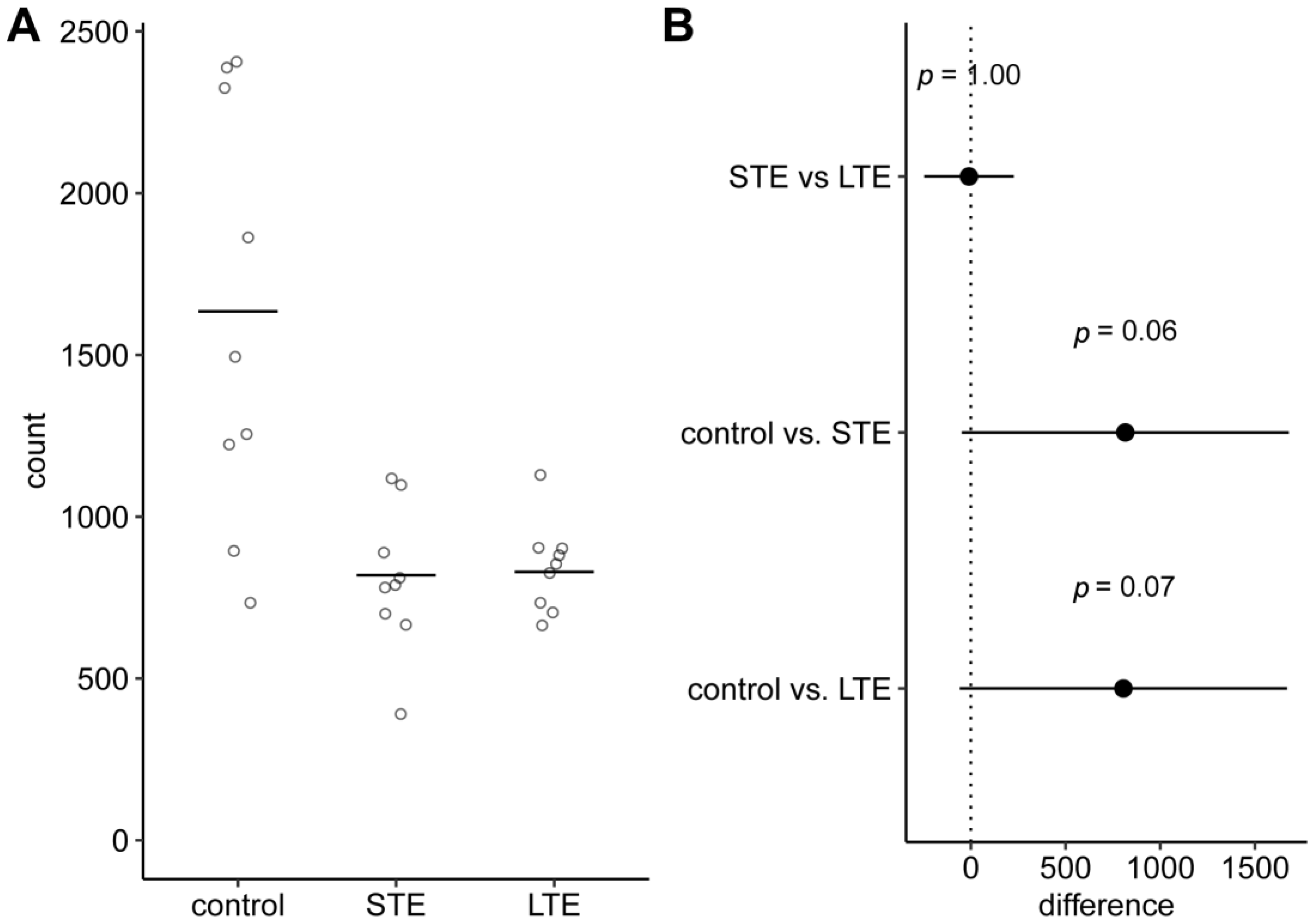
3.1. Number of Melanopsin Positive ipRGC Somas in Rat Retinas Was Lower after Blue Light Exposure
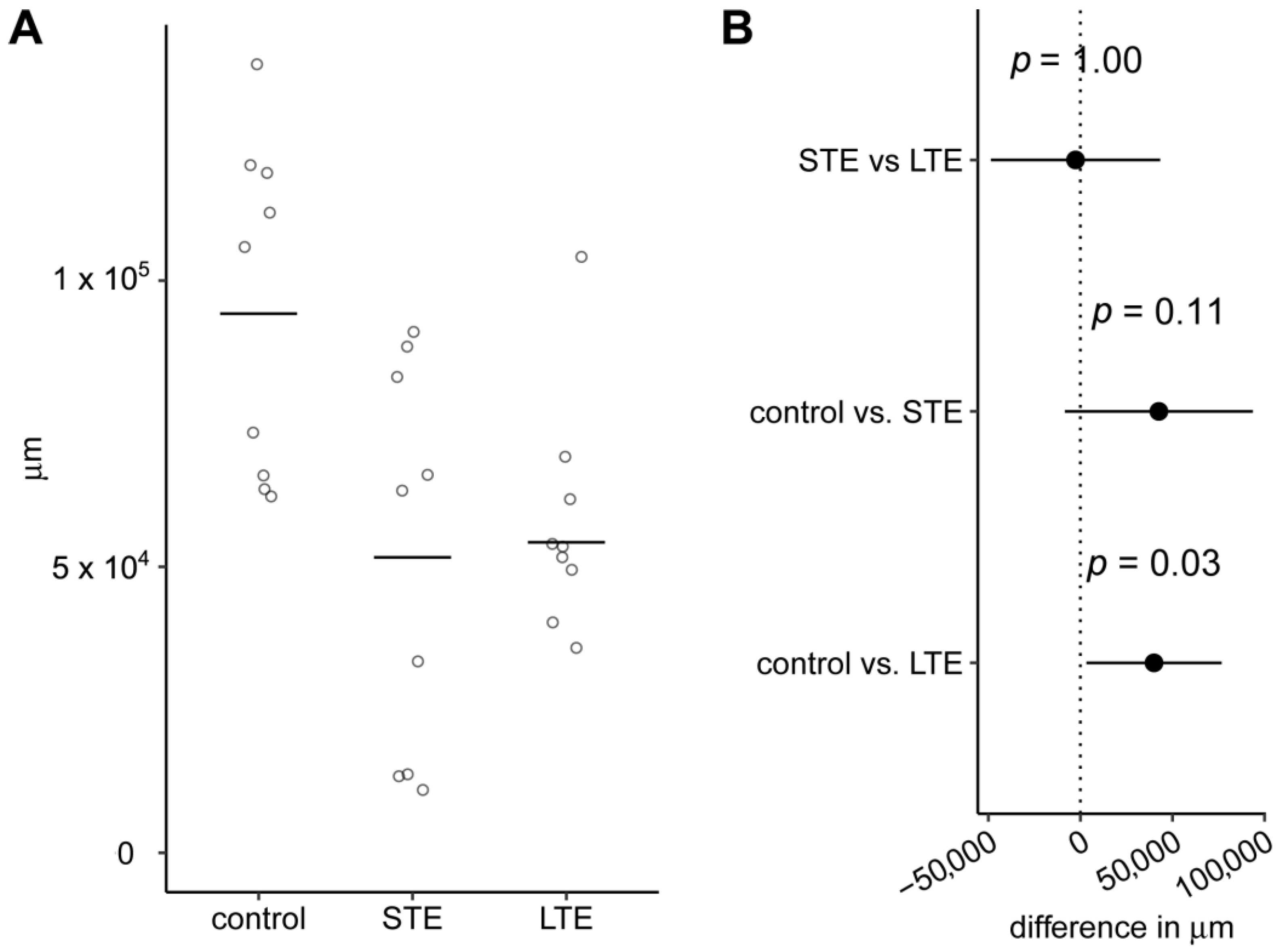
3.2. Length of Melanopsin Positive ipRGC Dendrites Was Shorter after Blue Light Exposure
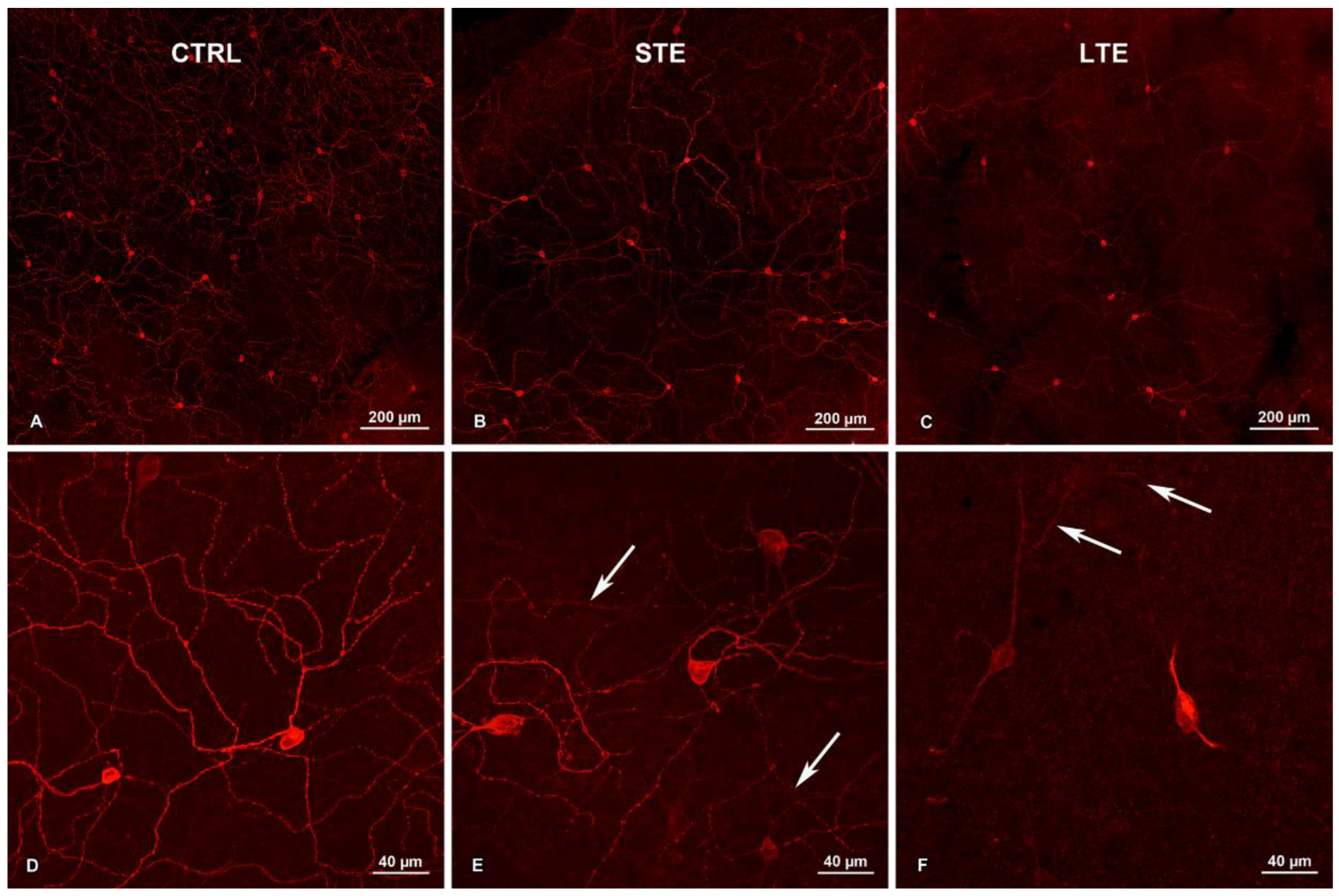
3.3. Exposure to Blue Light Changed Melanopsin Distribution in ipRGCs
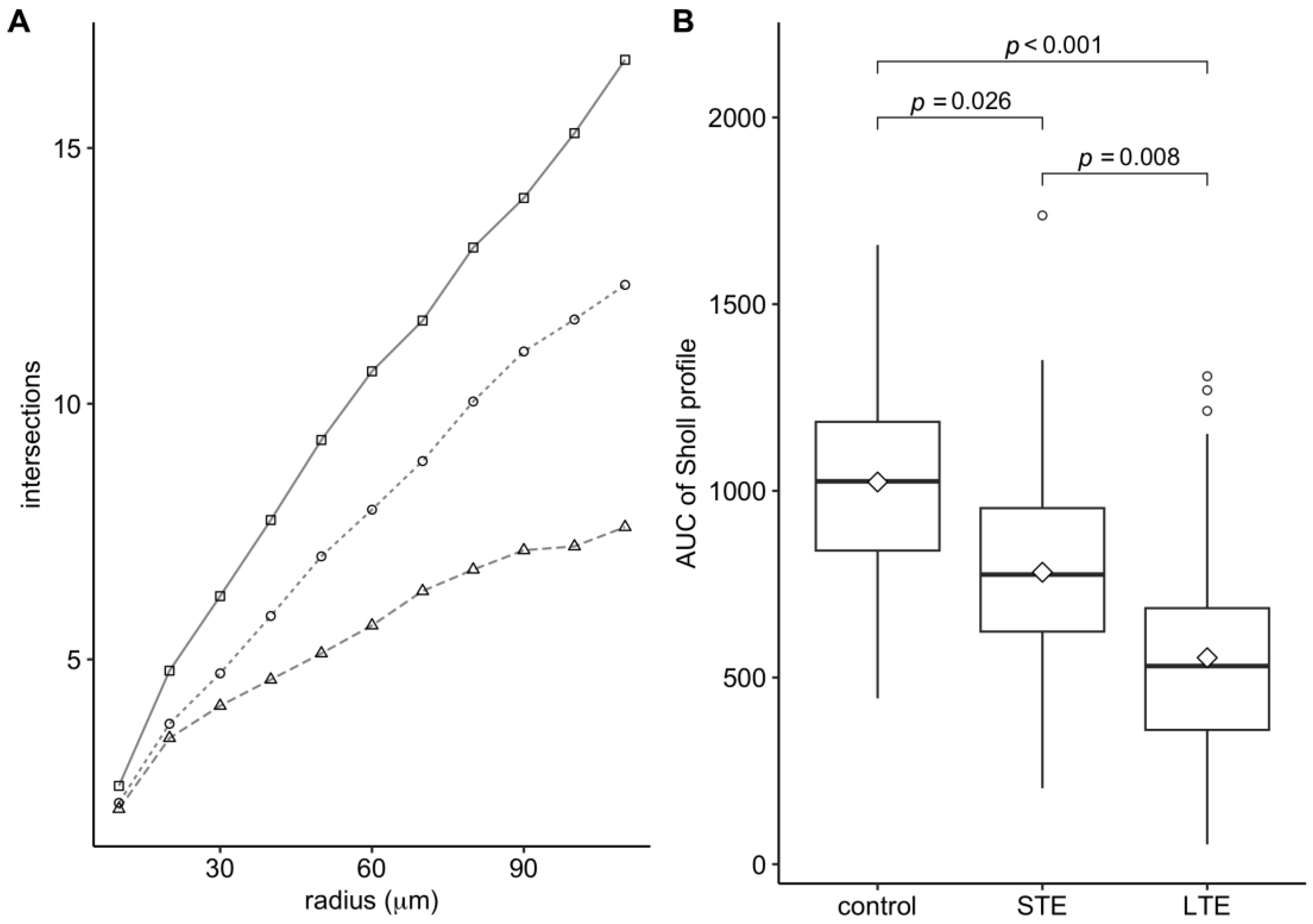
3.4. Blue-Light Exposure Reduced the Complexity of the Dendritic Arbor of ipRGCs
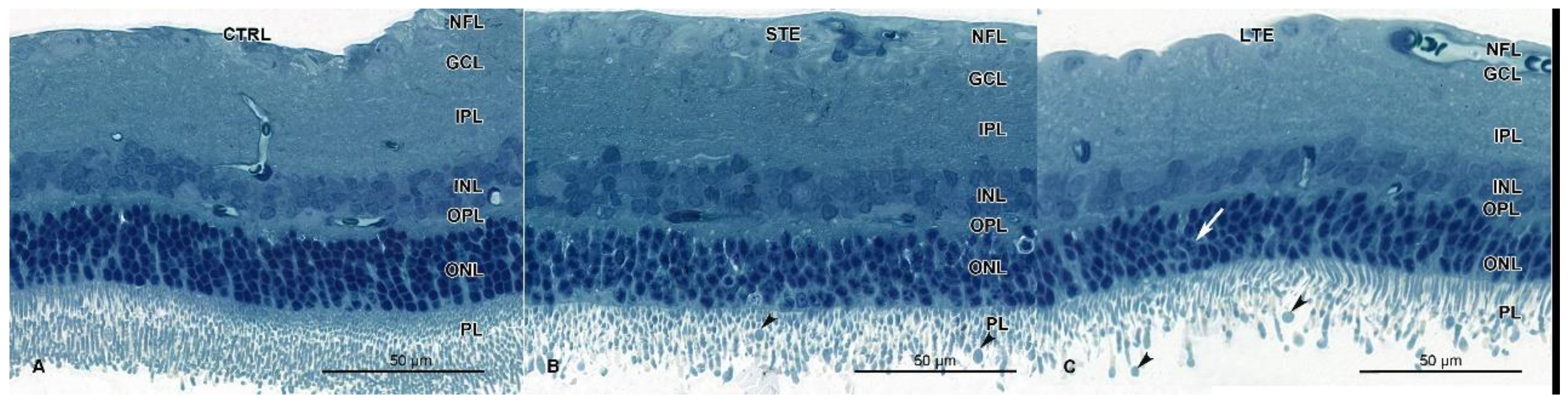
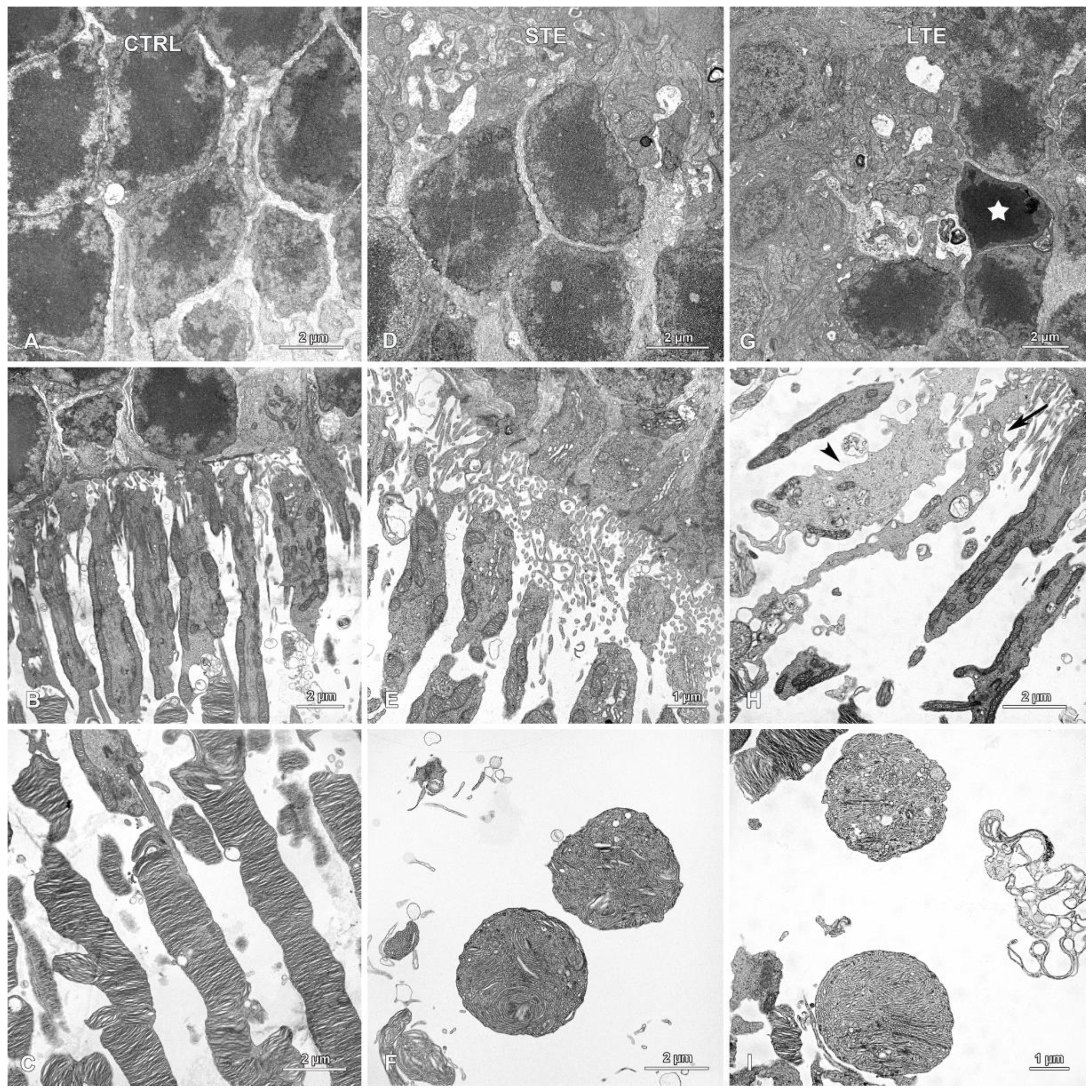
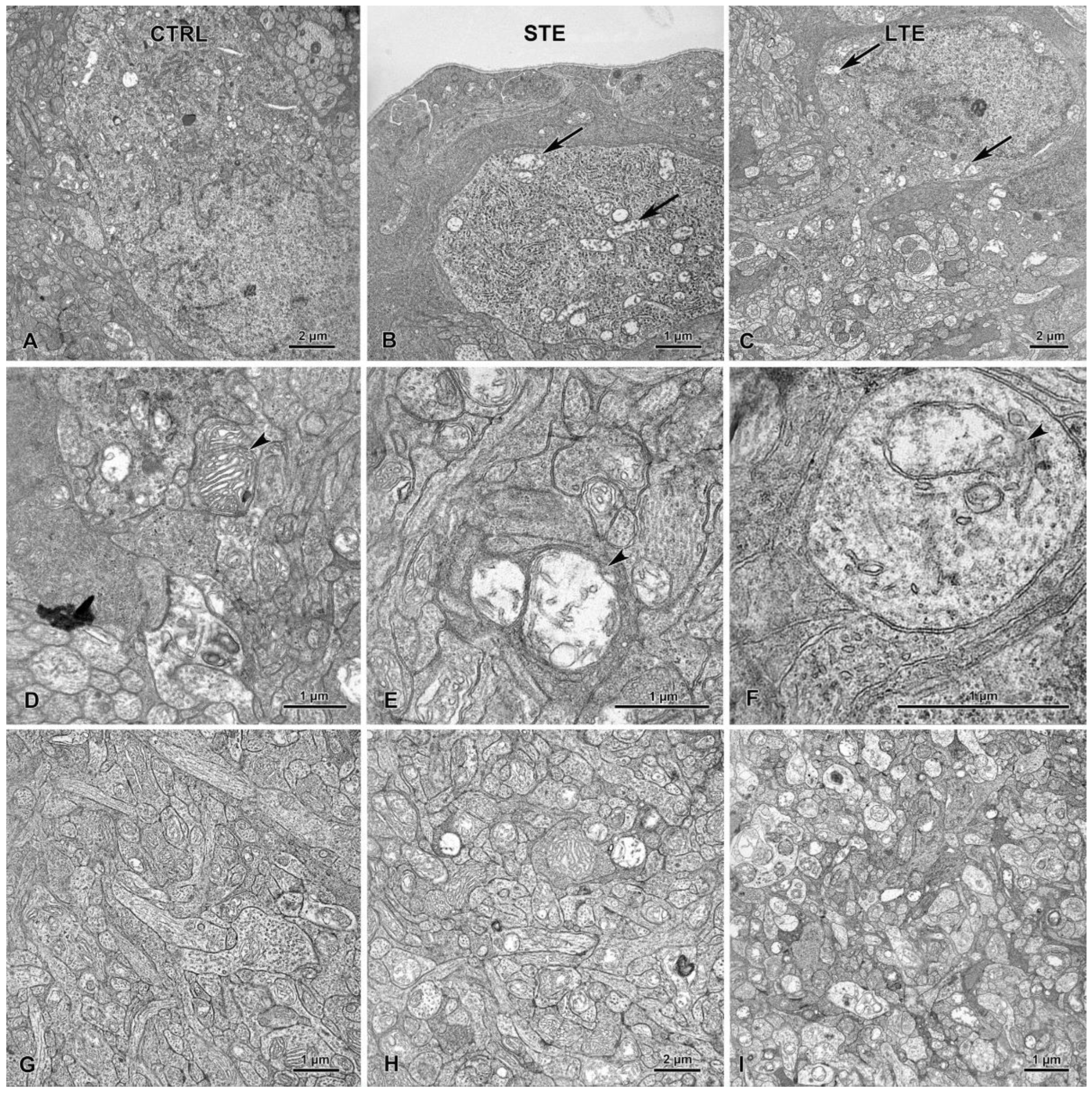
3.5. Retinal Damage after Exposure to Blue Light
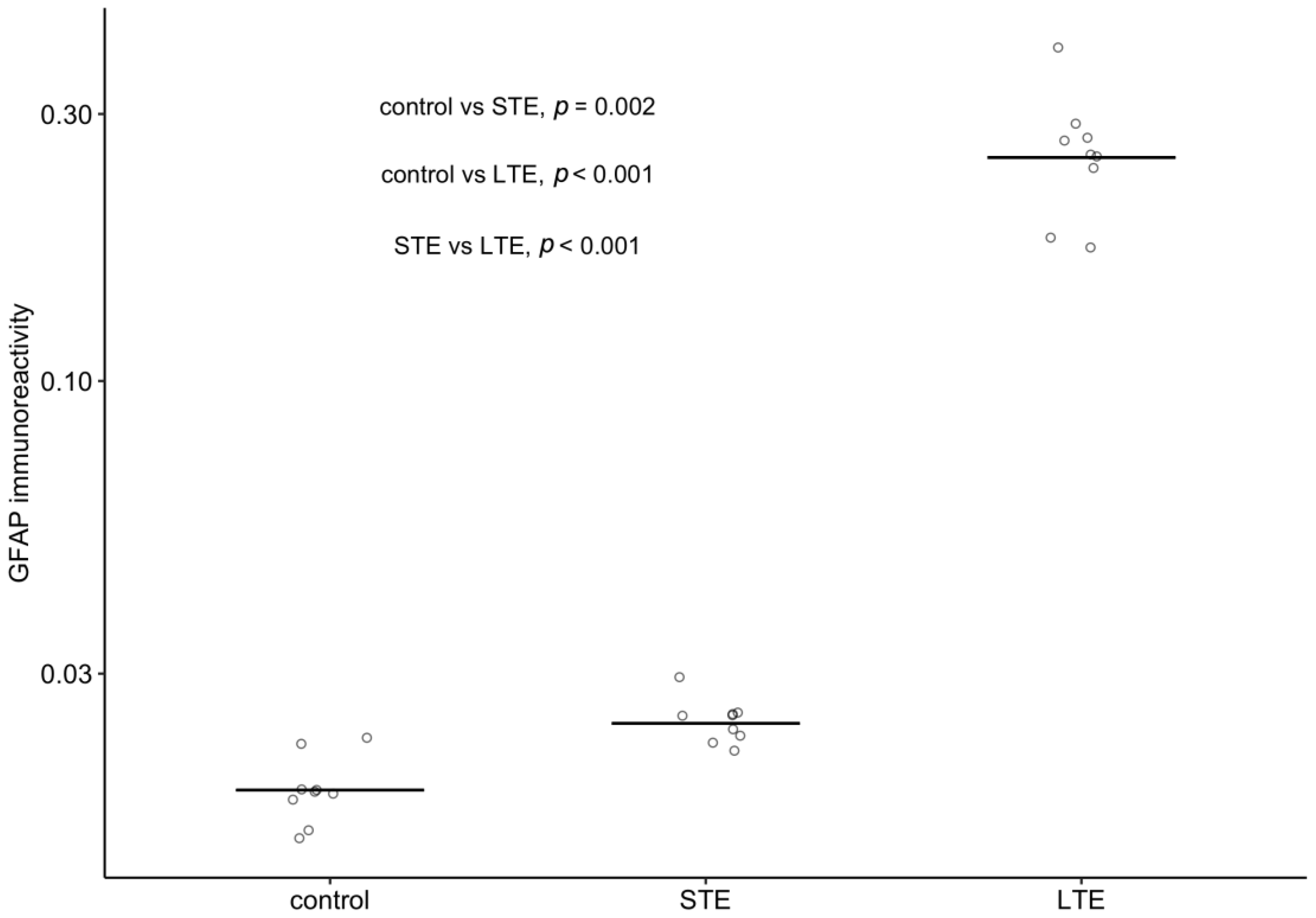
3.6. Blue-Light Exposure Increased GFAP Immunoreactivity and Changed Its Distribution in Rat Retinas
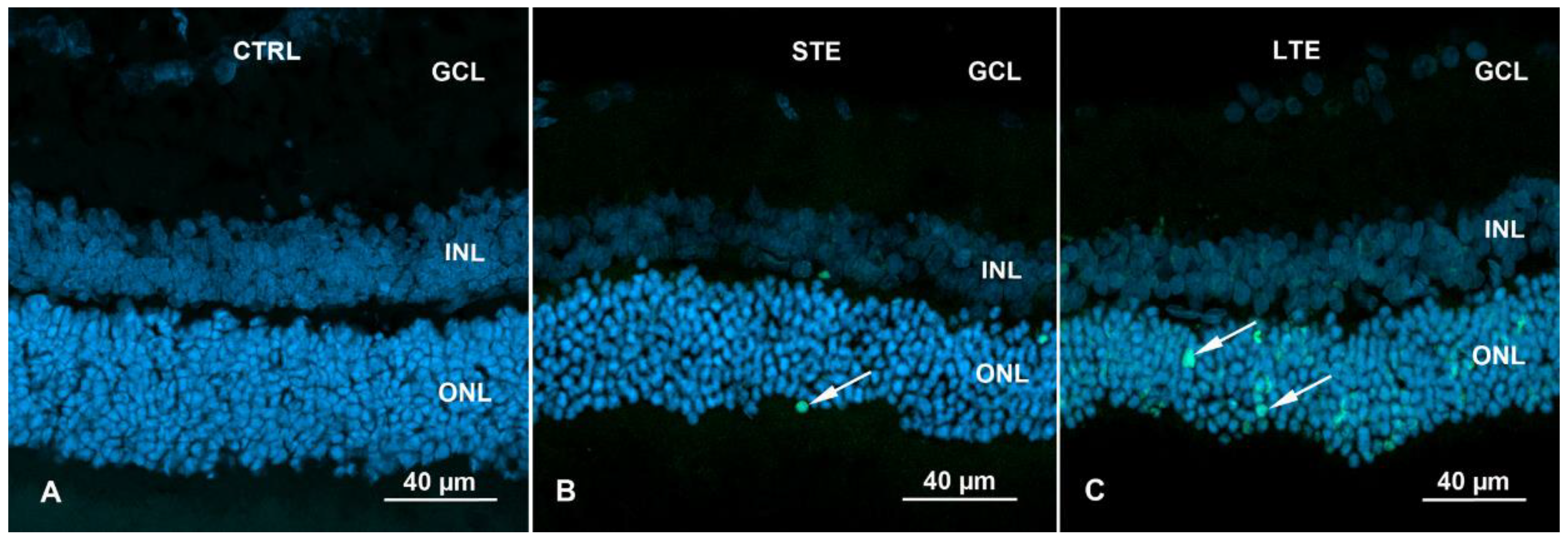
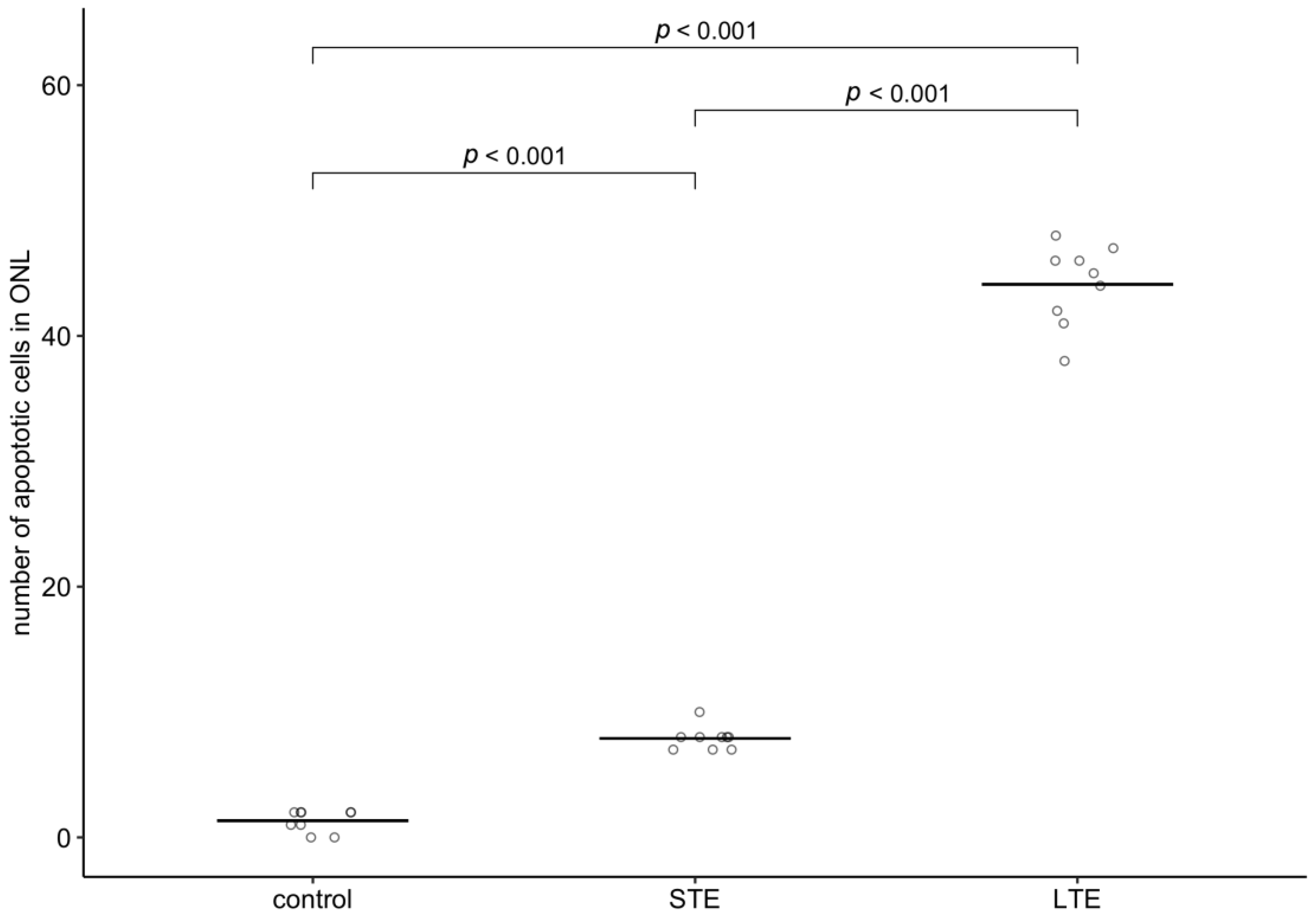
3.7. Increased Apoptosis in ONL of Rats Subjected to LTE and STE to Blue Light
3.8. Electron Microscopy Revealed Mitochondrial Swelling after LTE to Blue Light
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ham, W.T., Jr.; Mueller, H.A.; Sliney, D.H. Retinal sensitivity to damage from short wavelength light. Nature 1976, 260, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Grimm, C.; Wenzel, A.; Williams, T.; Rol, P.; Hafezi, F.; Reme, C. Rhodopsin-mediated blue-light damage to the rat retina: Effect of photoreversal of bleaching. Investig. Ophthalmol. Vis. Sci. 2001, 42, 497–505. [Google Scholar]
- Iandiev, I.; Wurm, A.; Hollborn, M.; Wiedemann, P.; Grimm, C.; Remé, C.E.; Reichenbach, A.; Pannicke, T.; Bringmann, A. Müller Cell Response to Blue Light Injury of the Rat Retina. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3559–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krigel, A.; Berdugo, M.; Picard, E.; Levy-Boukris, R.; Jaadane, I.; Jonet, L.; Dernigoghossian, M.; Andrieu-Soler, C.; Torriglia, A.; Behar-Cohen, F. Light-induced Retinal Damage Using Different Light Sources, Protocols and Rat Strains Reveals LED Phototoxicity. Neuroscience 2016, 339, 296–307. [Google Scholar] [CrossRef] [Green Version]
- Kuse, Y.; Ogawa, K.; Tsuruma, K.; Shimazawa, M.; Hara, H. Damage of Photoreceptor-Derived Cells in Culture Induced by Light Emitting Diode-Derived Blue Light. Sci. Rep. 2014, 4, 5223. [Google Scholar] [CrossRef] [Green Version]
- Ratnayake, K.; Payton, J.L.; Meger, M.E.; Godage, N.H.; Gionfriddo, E.; Karunarathne, A. Blue light-triggered photochemistry and cytotoxicity of retinal. Cell. Signal. 2020, 69, 109547. [Google Scholar] [CrossRef]
- Roehlecke, C.; Schumann, U.; Ader, M.; Knels, L.; Funk, R.H. Influence of blue light on photoreceptors in a live retinal explant system. Mol. Vis. 2011, 17, 876–884. Available online: http://www.molvis.org/molvis/v17/a98 (accessed on 8 April 2011).
- Shang, Y.M.; Wang, G.S.; Sliney, D.; Yang, C.H.; Lee, L.L. White light-emitting diodes (LEDs) at domestic lighting levels and retinal injury in a rat model. Environ. Health Perspect. 2014, 122, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Shang, Y.; Wang, G.; Sliney, D.H.; Yang, C.; Lee, L. Light-emitting diode induced retinal damage and its wavelength dependency in vivo. Int. J. Ophthalmol. 2017, 10, 191–202. [Google Scholar] [CrossRef]
- Jaadane, I.; Villalpando Rodriguez, G.; Boulenguez, P.; Carré, S.; Dassieni, I.; Lebon, C.; Chahory, S.; Behar-Cohen, F.; Martinsons, C.; Torriglia, A. Retinal phototoxicity and the evaluation of the blue light hazard of a new solid-state lighting technology. Sci. Rep. 2020, 10, 6733. [Google Scholar] [CrossRef] [Green Version]
- Lucas, R.J.; Douglas, R.H.; Foster, R.G. Characterization of an ocular photopigment capable of driving pupillary constriction in mice. Neuroscience 2001, 4, 621–626. [Google Scholar] [CrossRef]
- Güler, A.; Ecker, J.; Lall, G.; Haq, S.; Altimus, C.M.; Liao, H.W.; Barnard, A.R.; Cahill, H.; Badea, T.C.; Zhao, H.; et al. Melanopsin cells are the principal conduits for rod-cone input to non-image-forming vision. Nature 2008, 453, 102–105. [Google Scholar] [CrossRef] [Green Version]
- Hatori, M.; Le, H.; Vollmers, C.; Keding, S.R.; Tanaka, N.; Schmedt, C.; Jegla, T.; Panda, S. Inducible ablation of melanopsin-expressing retinal ganglion cells reveals their central role in non-image forming visual responses. PLoS ONE 2008, 3, e2451. [Google Scholar] [CrossRef]
- Jones, K.A.; Hatori, M.; Mure, L.S.; Bramley, J.R.; Artymyshyn, R.; Hong, S.P.; Marzabadi, M.; Zhong, H.; Sprouse, J.; Zhu, Q.; et al. Small-molecule antagonists of melanopsin-mediated phototransduction. Nat. Chem. Biol. 2013, 9, 630–635. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Provencio, I.; Tu, D.C.; Pires, S.S.; Rollag, M.D.; Castrucci, A.M.; Pletcher, M.T.; Sato, T.K.; Wiltshire, T.; Andahazy, M.; et al. Melanopsin is required for non-image-forming photic responses in blind mice. Science 2003, 301, 525–527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, A.E.; Storchi, R.; Martial, F.P.; Bedford, R.A.; Lucas, R.J. Melanopsin contributions to the representation of images in the early visual system. Curr. Biol. 2017, 27, 1623–1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnard, A.R.; Hattar, S.; Hankins, M.W.; Lucas, R.J. Melanopsin regulates visual processing in the mouse retina. Curr. Biol. 2006, 16, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.M.; Tsujimura, S.I.; Allen, A.E.; Wynne, J.; Bedford, R.; Vickery, G.; Vugler, A.; Lucas, R.J. Melanopsin-based brightness discrimination in mice and humans. Curr. Biol. 2012, 22, 1134–1141. [Google Scholar] [CrossRef] [Green Version]
- Berson, D.M. Strange vision: Ganglion cells as circadian photoreceptors. Trends Neurosci. 2003, 26, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Pickard, G.E.; Baver, S.B.; Ogilvie, M.D.; Sollars, P.J. Light-induced Fos expression in intrinsically photosensitive retinal ganglion cells in melanopsin knockout (Opn4−/−) mice. PLoS ONE 2009, 4, e4984. [Google Scholar] [CrossRef] [Green Version]
- García-Ayuso, D.; Galindo-Romero, C.; Di Pierdomenico, J.; Vidal-Sanz, M.; Agudo-Barriuso, M.; Villegas Pérez, M.P. Light-induced retinal degeneration causes a transient downregulation of melanopsin in the rat retina. Exp. Eye Res. 2017, 161, 10–16. [Google Scholar] [CrossRef]
- Hannibal, J.; Georg, B.; Hindersson, P.; Fahrenkrug, J. Light and darkness regulate melanopsin in the retinal ganglion cells of the albino Wistar rat. J. Mol. Neurosci. 2005, 27, 147–155. [Google Scholar] [CrossRef]
- Hannibal, J.; Georg, B.; Fahrenkrug, J. Differential expression of melanopsin mRNA and protein in Brown Norwegian rats. Exp. Eye Res. 2013, 106, 55–63. [Google Scholar] [CrossRef]
- Benedetto, M.M.; Guido, M.E.; Contin, M.A. Non-Visual Photopigments Effects of Constant Light-Emitting Diode Light Exposure on the Inner Retina of Wistar Rats. Front. Neurol. 2017, 8, 417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziółkowska, N.; Chmielewska-Krzesińska, M.; Vyniarska, A.; Sienkiewicz, W. Exposure to Blue Light Reduces Melanopsin Expression in Intrinsically Photoreceptive Retinal Ganglion Cells and Damages the Inner Retina in Rats. Investig. Ophthalmol. Vis. Sci. 2022, 63, 26. [Google Scholar] [CrossRef]
- Wasowicz, M.; Morice, C.; Ferrari, P.; Callebert, J.; Versaux-Botteri, C. Long-term effects of light damage on the retina of albino and pigmented rats. Investig. Ophthalmol. Vis. Sci. 2002, 43, 813–820. [Google Scholar]
- del Olmo-Aguado, S.; Núñez-Álvarez, C.; Osborne, N.N. Blue light action on mitochondria leads to cell death by necroptosis. Neurochem. Res. 2016, 41, 2324–2335. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Soderberg, P.G.; Lindstrom, B. Cytochrome oxidase activity in rat retina after exposure to 404 nm blue light. Curr. Eye Res. 1992, 11, 825–831. [Google Scholar] [CrossRef]
- Chen, P.G.; Soderberg, W. Qian Inhibition of cytochrome oxidase by blue light (404 nm). A factor that causes retinal injury? Investig. Ophthalmol. Vis. Sci. 1992, 33, 919. [Google Scholar] [CrossRef]
- Egorov, S.Y.; Krasnovsky, A.A., Jr.; Bashtanov, M.Y.; Mironov, E.A.; Ludnikova, T.A.; Kritsky, M.S. Photosensitization of singlet oxygen formation by pterins and flavins. Time-resolved studies of oxygen phosphorescence under laser excitation. Biochemistry 1999, 64, 1117. [Google Scholar]
- Osborne, N.N.; Nunez-Alvarez, C.; del Olmo-Aguado, S. The effect of visual blue light on mitochondrial function associated with retinal ganglions cells. Exp. Eye Res. 2014, 128, 8–14. [Google Scholar] [CrossRef]
- Bristow, E.A.; Griffiths, P.G.; Andrews, R.M.; Johnson, M.A.; Turnbull, D.M. The distribution of mitochondrial activity in relation to optic nerve structure. Arch. Ophthalmol. 2002, 120, 791–796. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.N. Mitochondria: Their role in ganglion cell death and survival in primary open angle glaucoma. Exp. Eye Res. 2010, 90, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, R. Introduction to Robust Estimation and Hypothesis Testing, 3rd ed.; Academic Press: Waltham, MA, USA, 2012; pp. 317–322. [Google Scholar]
- Mair, P.; Wilcox, R. Robust statistical methods in R using the WRS2 package. Behav. Res. Methods 2020, 52, 464–488. [Google Scholar] [CrossRef] [PubMed]
- Lenth, R. emmeans: Estimated Marginal Means, aka Least-Squares Means. Available online: https://github.com/rvlenth/emmeans (accessed on 8 March 2023).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2022. Available online: https://www.R-project.org/ (accessed on 15 March 2023).
- García-Ayuso, D.; Di Pierdomenico, J.; Hadj-Said, W.; Marie, M.; Agudo-Barriuso, M.; Vidal-Sanz, M.; Villegas-Pérez, M.P. Taurine depletion causes ipRGC loss and increases light-induced photoreceptor degeneration. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1396–1409. [Google Scholar] [CrossRef]
- Galindo-Romero, C.; Jiménez-López, M.; Garcia-Ayuso, D.; Salinas-Navarro, M.; Nadal-Nicolás, F.M.; Agudo-Barriuso, M.; Vidal-Sanz, M. Number and spatial distribution of intrinsically photosensitive retinal ganglion cells in the adult albino rat. Exp. Eye Res. 2013, 108, 84–93. [Google Scholar] [CrossRef]
- Belenky, M.A.; Smeraski, C.A.; Provencio, I.; Sollars, P.J.; Pickard, G.E. Melanopsin retinal ganglion cells receive bipolar and amacrine cell synapses. J. Comp. Neurol. 2003, 460, 380–393. [Google Scholar] [CrossRef]
- Dacey, D.M.; Liao, H.W.; Peterson, B.B.; Robinson, F.R.; Smith, V.C.; Pokorny, J.; Yau, K.W.; Gamlin, P.D. Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN. Nature 2005, 433, 749–754. [Google Scholar] [CrossRef]
- Castrucci, A.D.L.; Ihara, N.; Doyle, S.E.; Rollag, M.D.; Provencio, I.; Menaker, M. Light regulation of melanopsin-positive retinal ganglion cells in the albino hamster. ARVO Annual Meeting Abstract. Investig. Ophthalmol. Vis. Sci. 2004, 45, 4645. [Google Scholar]
- Gonzalez-Menendez, I.; Contreras, F.; Cernuda-Cernuda, R.; Provencio, I.; Garcia- Fernandez, J.M. Postnatal development and functional adaptations of the melanopsin photoreceptive system in the albino mouse retina. Investig. Ophthalmol. Vis. Sci. 2010, 51, 4840–4847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannibal, J.; Georg, B.; Fahrenkrug, J. Melanopsin changes in neonatal albino rat independent of rods and cones. Neuroreport 2007, 18, 81–85. [Google Scholar] [CrossRef]
- Bailes, H.J.; Lucas, R.J. Human melanopsin forms a pigment maximally sensitive to blue light (λmax ≈ 479 nm) supporting activation of Gq/11 and Gi/o signalling cascades. Proc. R. Soc. B Biol. Sci. 2013, 280, 20122987. [Google Scholar] [CrossRef] [Green Version]
- Miralles de Imperial-Ollero, J.A.; Gallego-Ortega, A.; Norte-Muñoz, M.; Di Pierdomenico, J.; Bernal-Garro, J.M.; Valiente-Soriano, F.J.; Vidal-Sanz, M. Short-and Long-Term Study of the Impact of Focal Blue Light-Emitting Diode-Induced Phototoxicity in Adult Albino Rats. Int. J. Mol. Sci. 2021, 22, 9742. [Google Scholar] [CrossRef]
- Ziółkowska, N.; Lewczuk, B. Profiles of Rho, Opn4, c-Fos, and Birc5 mRNA expression in Wistar rat retinas exposed to white or monochromatic light. Front. Neuroanat. 2022, 16, 956000. [Google Scholar] [CrossRef] [PubMed]
- Kuwabara, T.; Gorn, R.A. Retinal damage by visible light: An electron microscopic study. Arch. Ophthalmol. 1968, 79, 69–78. [Google Scholar] [CrossRef]
- de Raad, S.; Szczesny, P.J.; Munz, K.; Remé, C.E. Light damage in the rat retina: Glial fibrillary acidic protein accumulates in Müller cells in correlation with photoreceptor damage. Ophthalmic Res. 1996, 28, 99–107. [Google Scholar] [CrossRef]
- Bringmann, A.; Pannicke, T.; Grosche, J.; Francke, M.; Wiedemann, P.; Skatchkov, S.N.; Osborne, N.N.; Reichenbach, A. Müller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 2006, 25, 397–424. [Google Scholar] [CrossRef]
- Burns, M.S.; Robles, M. Müller cell GFAP expression exhibits gradient from focus of photoreceptor light damage. Curr. Eye Res. 1990, 9, 479–486. [Google Scholar] [CrossRef]
- Jaadane, I.; Boulenguez, P.; Chahory, S.; Carré, S.; Savoldelli, M.; Jonet, L.; Behar-Cohen, F.; Martinsons, C.; Torriglia, A. Retinal Damage Induced by Commercial Light Emitting Diodes (LEDs). Free Radic. Biol. Med. 2015, 84, 373–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Lai, Z.; Wu, Y.; Xu, L.; Cai, X.; Qiu, J.; Zhuang, J. Retinal neuron is more sensitive to blue light-induced damage than glia cell due to DNA double-strand breaks. Cells 2019, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Fietz, A.; Hurst, J.; Schnichels, S. Out of the Shadow: Blue Light Exposure Induces Apoptosis in Müller Cells. Int. J. Mol. Sci. 2022, 23, 14540. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, P.; Wang, W.; Xu, Y.; Wang, M.; Chen, X.; Dong, X. Long-term blue light exposure induces RGC-5 cell death in vitro: Involvement of mitochondria-dependent apoptosis, oxidative stress, and MAPK signaling pathways. Apoptosis 2014, 19, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Álvarez, C.; Osborne, N.N. Blue light exacerbates and red light counteracts negative insults to retinal ganglion cells in situ and R28 cells in vitro. Neurochem. Int. 2019, 125, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Ren, C.; Sollars, P.J.; Pickard, G.E.; So, K.F. The injury resistant ability of melanopsin-expressing intrinsically photosensitive retinal ganglion cells. Neuroscience 2015, 284, 845–853. [Google Scholar] [CrossRef] [Green Version]
- Li, R.S.; Chen, B.Y.; Tay, D.K.; Chan, H.H.; Pu, M.L.; So, K.F. Melanopsin-expressing retinal ganglion cells are more injury-resistant in a chronic ocular hypertension model. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2951–2958. [Google Scholar] [CrossRef] [PubMed]
- Ba-Ali, S.; Lund-Andersen, H. Pupillometric evaluation of the melanopsin containing retinal ganglion cells in mitochondrial and non-mitochondrial optic neuropathies. Mitochondrion 2017, 36, 124–129. [Google Scholar] [CrossRef]
- Hannibal, J. Roles of PACAP-containing retinal ganglion cells in circadian timing. Int. Rev. Cytol. 2006, 251, 1–39. [Google Scholar]
- Osborne, N.N.; Li, G.Y.; Ji, D.; Mortiboys, H.J.; Jackson, S. Light affects mitochondria to cause apoptosis to cultured cells: Possible relevance to ganglion cell death in certain optic neuropathies. J. Neurochem. 2008, 105, 2013–2028. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Quintanilha, A.T.; Cammack, R.; Packer, L. Damage to mitochondrial electron transport and energy coupling by visible light. Biochim. Biophys. Acta Bioenergy 2014, 502, 367–382. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Number | Whole Mount Immunocytochemistry a | TUNEL b and ICH c | Transmission Electron Microscopy d | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Short Term | Long Term | Control | Short Term | Long Term | Control | Short Term | Long Term | Control | ||||||||||
| Right | Left | Right | Left | Right | Left | Right | Left | Right | Left | Right | Left | Right | Left | Right | Left | Right | Left | |
| Rat 1 ♀ | + | − | + | − | + | − | − | + | − | + | − | + | − | + | − | + | − | + |
| Rat 2 ♂ | − | + | − | + | − | + | + | − | + | − | + | − | + | − | + | − | + | − |
| Rat 3 ♀ | + | − | + | − | + | − | − | + | − | + | − | + | − | + | − | + | − | + |
| Rat 4 ♂ | − | + | − | + | − | + | + | − | + | − | + | − | + | − | + | − | + | − |
| Rat 5 ♀ | + | − | + | − | + | − | − | + | − | + | − | + | − | + | − | + | − | + |
| Rat 6 ♂ | − | + | − | + | − | + | + | − | + | − | + | − | + | − | + | − | + | − |
| Rat 7 ♀ | + | − | + | − | + | − | − | + | − | + | − | + | − | − | − | − | − | − |
| Rat 8 ♂ | − | + | − | + | − | + | + | − | + | − | + | − | − | − | − | − | − | − |
| Rat 9 ♀ | + | − | + | − | + | − | − | + | − | + | − | + | − | − | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ziółkowska, N.; Lewczuk, B.; Szyryńska, N.; Rawicka, A.; Vyniarska, A. Low-Intensity Blue Light Exposure Reduces Melanopsin Expression in Intrinsically Photosensitive Retinal Ganglion Cells and Damages Mitochondria in Retinal Ganglion Cells in Wistar Rats. Cells 2023, 12, 1014. https://doi.org/10.3390/cells12071014
Ziółkowska N, Lewczuk B, Szyryńska N, Rawicka A, Vyniarska A. Low-Intensity Blue Light Exposure Reduces Melanopsin Expression in Intrinsically Photosensitive Retinal Ganglion Cells and Damages Mitochondria in Retinal Ganglion Cells in Wistar Rats. Cells. 2023; 12(7):1014. https://doi.org/10.3390/cells12071014
Chicago/Turabian StyleZiółkowska, Natalia, Bogdan Lewczuk, Natalia Szyryńska, Aleksandra Rawicka, and Alla Vyniarska. 2023. "Low-Intensity Blue Light Exposure Reduces Melanopsin Expression in Intrinsically Photosensitive Retinal Ganglion Cells and Damages Mitochondria in Retinal Ganglion Cells in Wistar Rats" Cells 12, no. 7: 1014. https://doi.org/10.3390/cells12071014
APA StyleZiółkowska, N., Lewczuk, B., Szyryńska, N., Rawicka, A., & Vyniarska, A. (2023). Low-Intensity Blue Light Exposure Reduces Melanopsin Expression in Intrinsically Photosensitive Retinal Ganglion Cells and Damages Mitochondria in Retinal Ganglion Cells in Wistar Rats. Cells, 12(7), 1014. https://doi.org/10.3390/cells12071014