
Intramuscular IL-10 Administration Enhances the Activity of Myogenic Precursor Cells and Improves Motor Function in ALS Mouse Model

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Behavioral Analysis
2.3. Dissection of Skeletal Muscles
2.4. Immunohistochemistry
2.5. Real-Time PCR
2.6. Western Blotting
2.7. Primary Satellite Cell Cultures and Immunofluorescence
2.8. Primary Mф Cultures
2.9. Mф Proliferation Assay
2.10. Sc/Mф Co-Culture for Migration Analysis
2.11. Statistics
3. Results
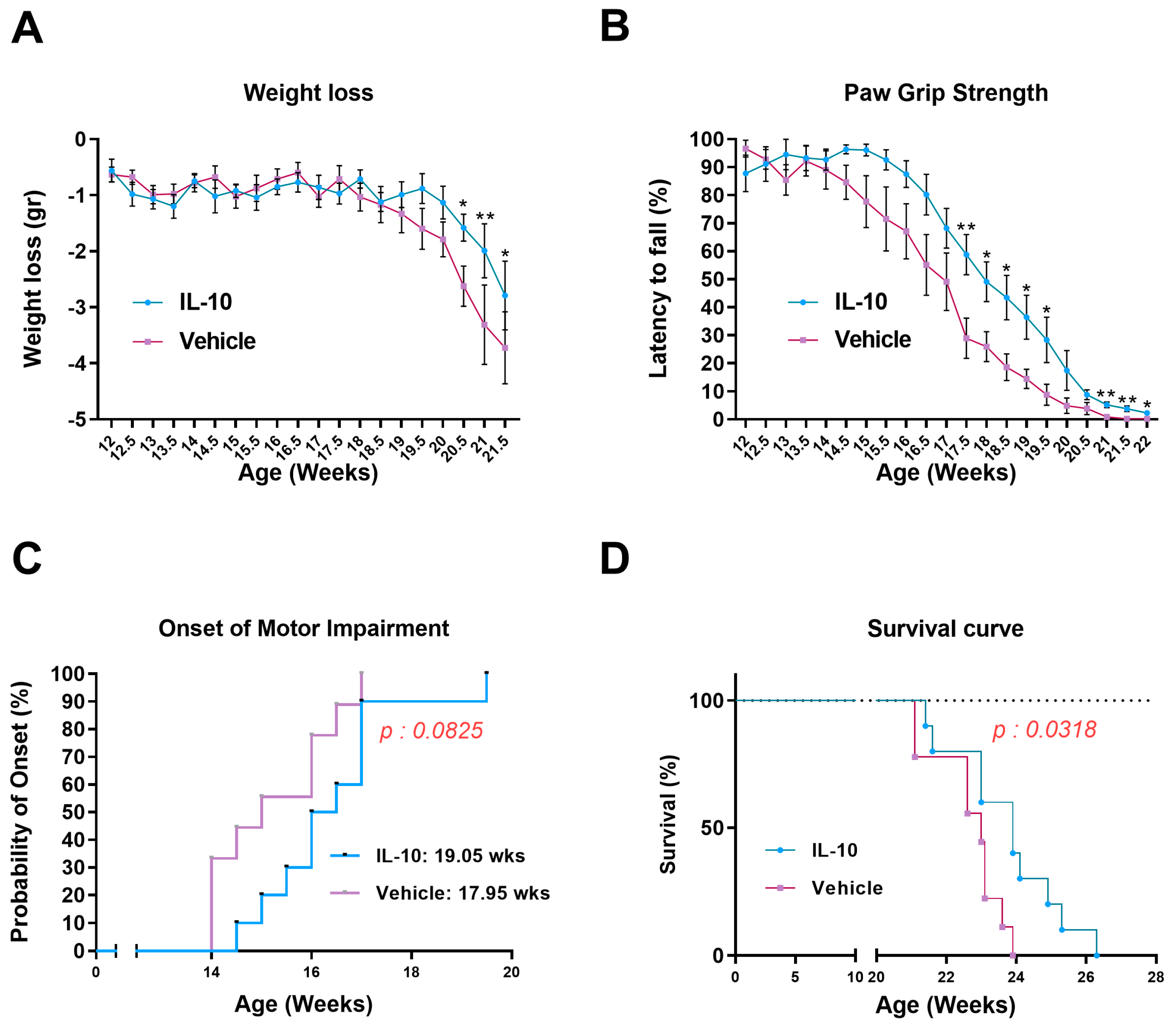
3.1. Intramuscular Il-10 Administration Ameliorates the Disease Progression and Extends the Survival of C57-SOD1G93A Mice
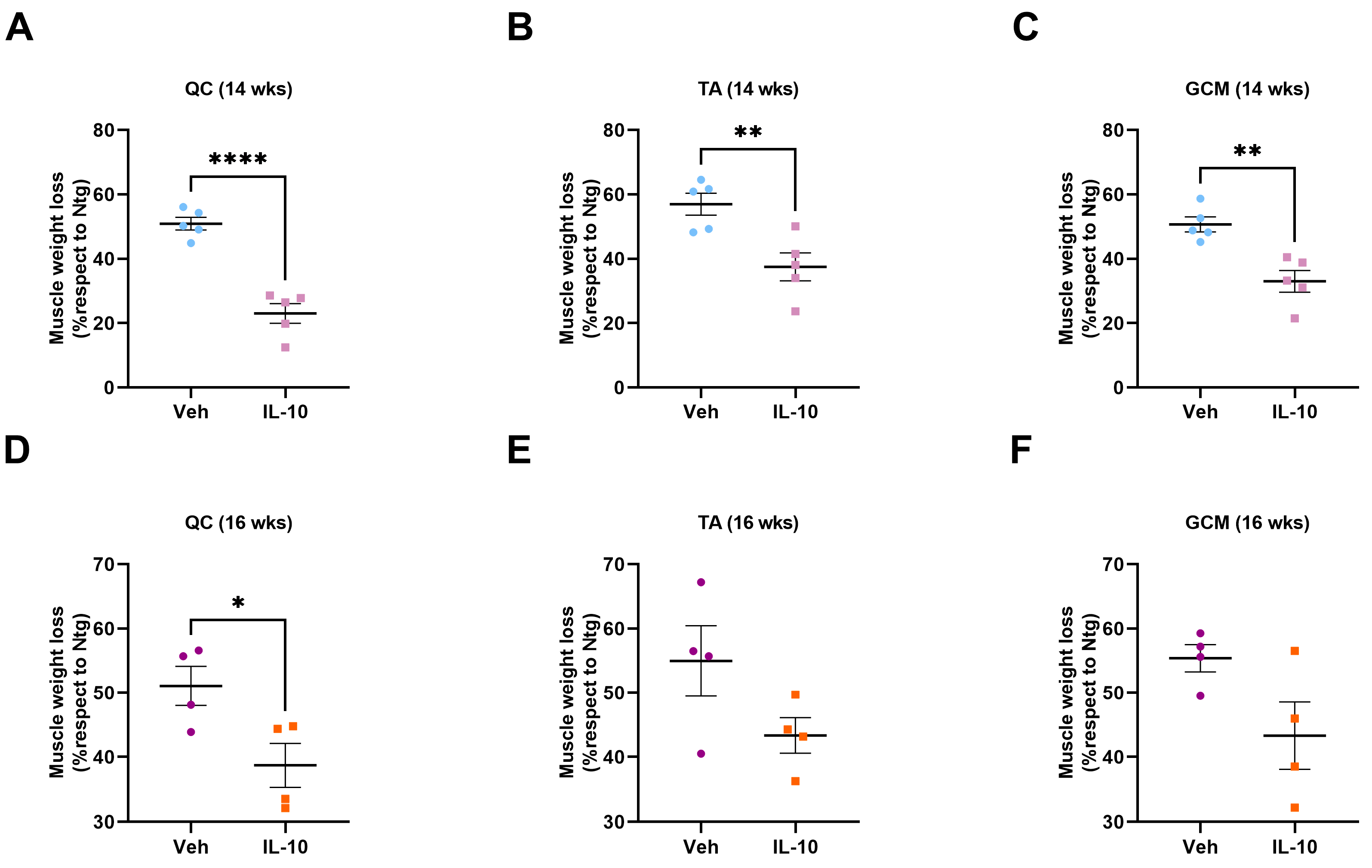
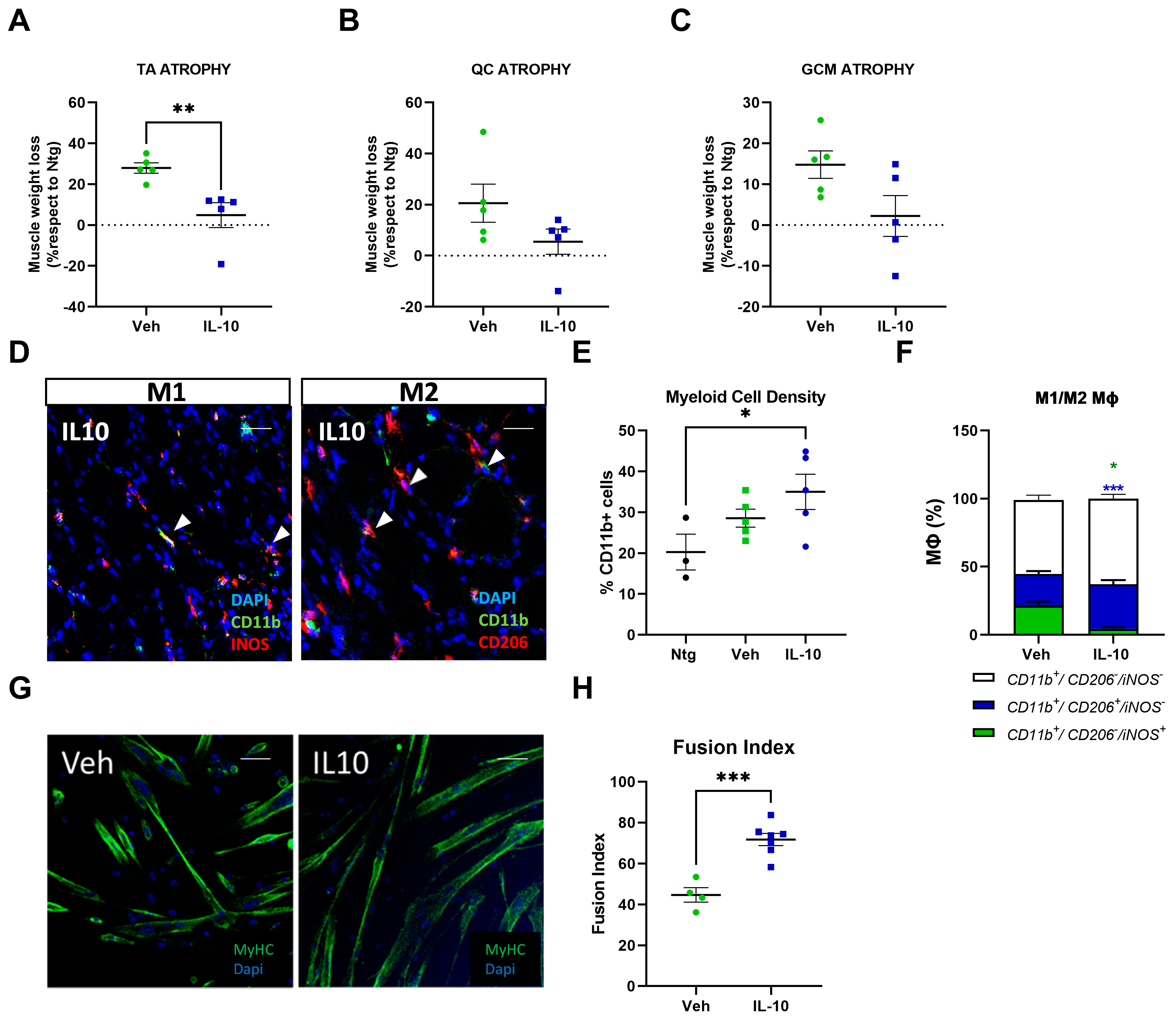
3.2. Intramuscular Il-10 Administration Delays Muscle Atrophy in C57-SOD1G93A Mice
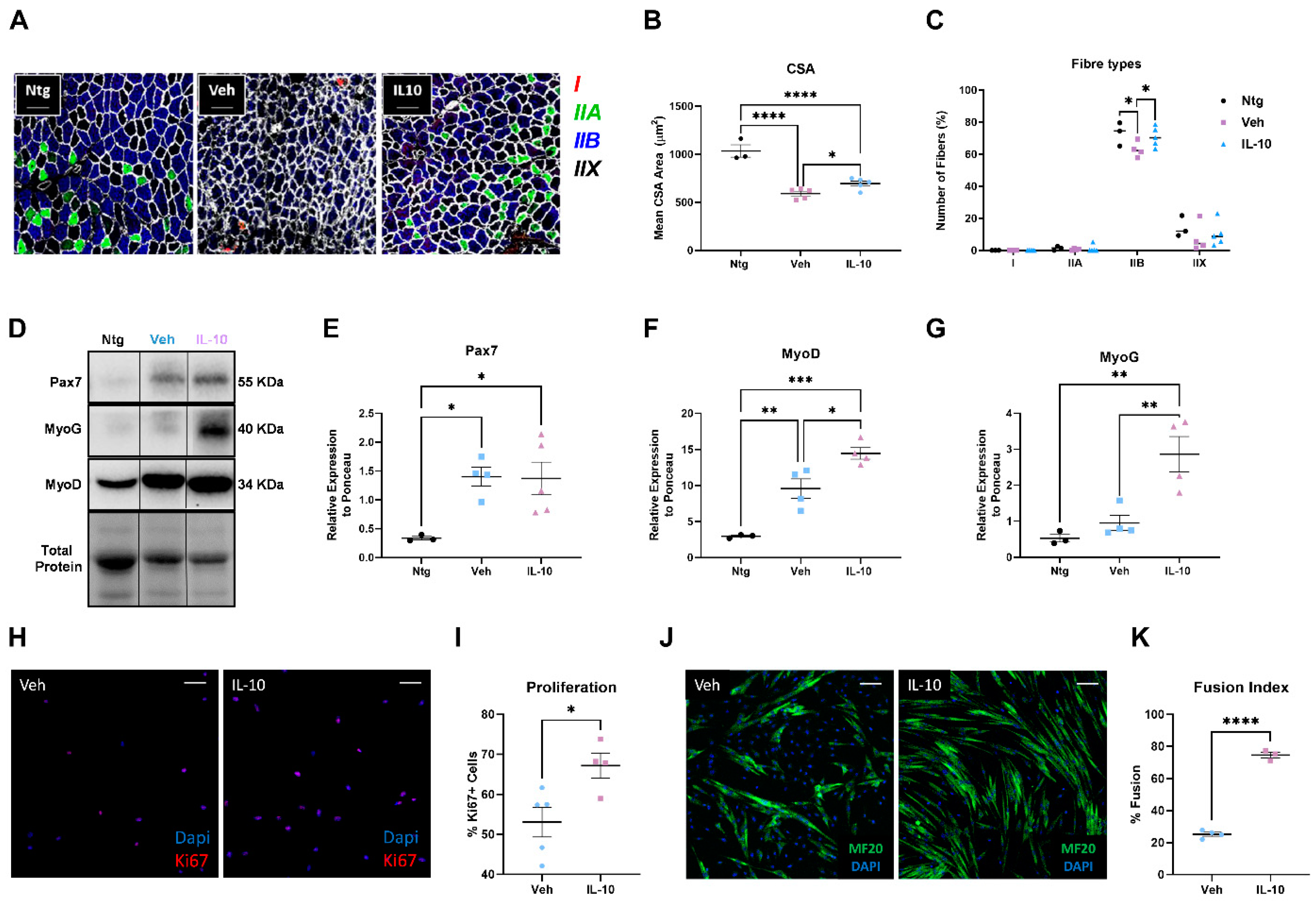
3.3. Intramuscular IL-10 Administration Preserves Muscle Architecture by Eliciting Satellite Cell Proliferation and Differentiation in C57-SOD1G93A Mice
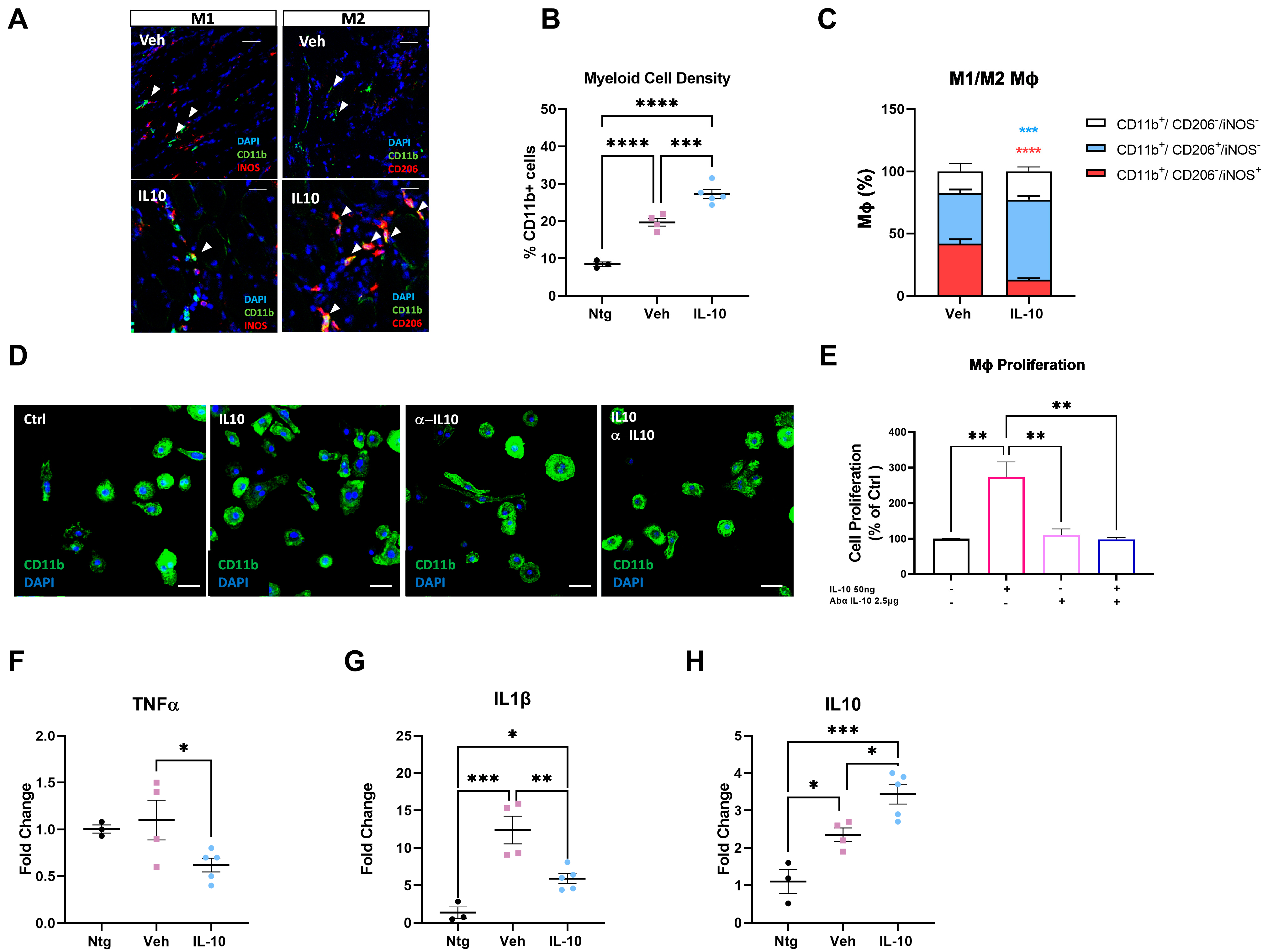
3.4. Intramuscular IL-10 Administration Promotes the Macrophage Polarization to M2 Pro-Regenerative Fingerprint in the Hind Limb Skeletal Muscles of C57-SOD1G93A Mice
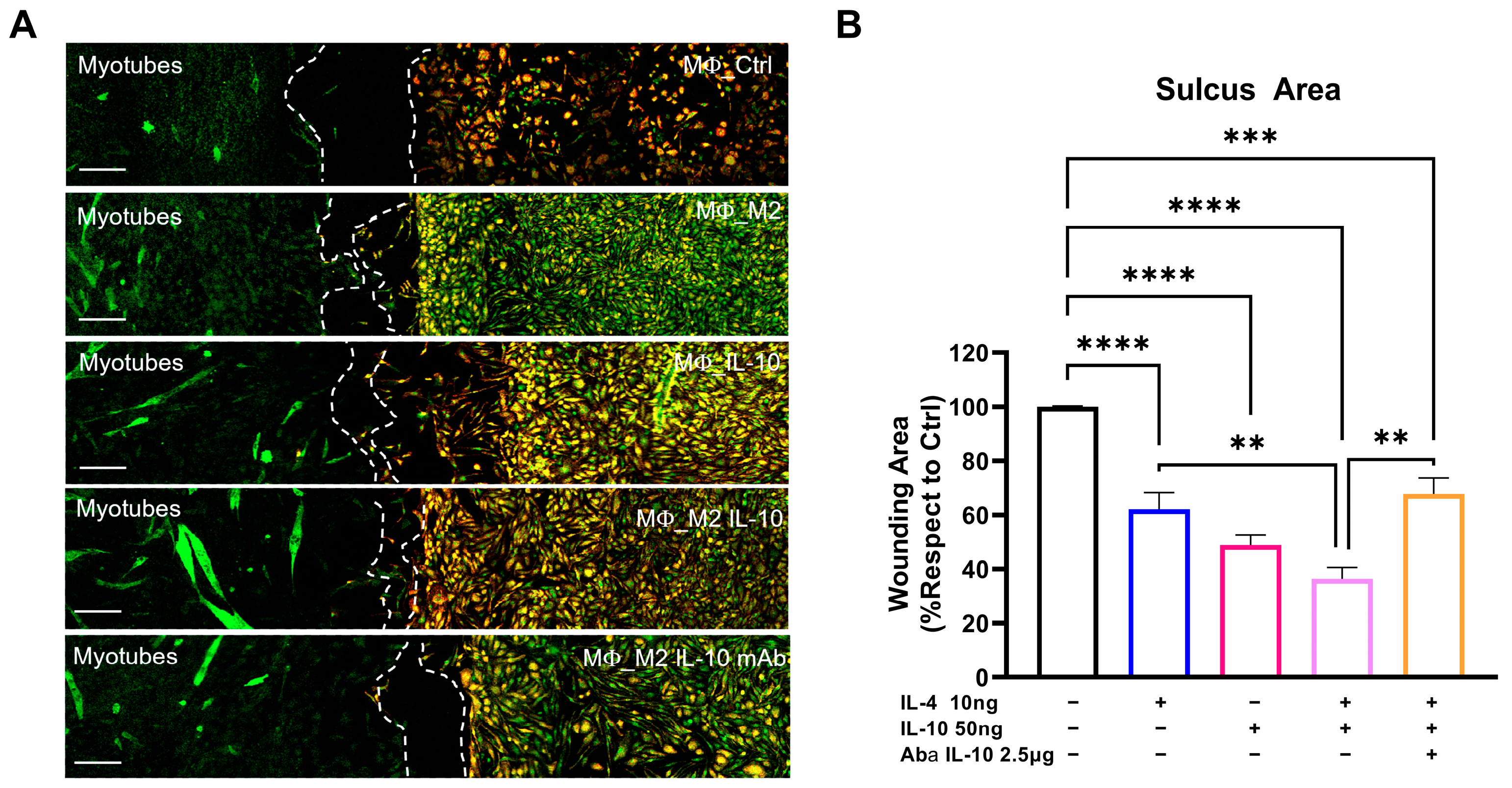
3.5. IL-10 In Vitro Administration Enhances the Macrophage–Satellite Cell Interplay
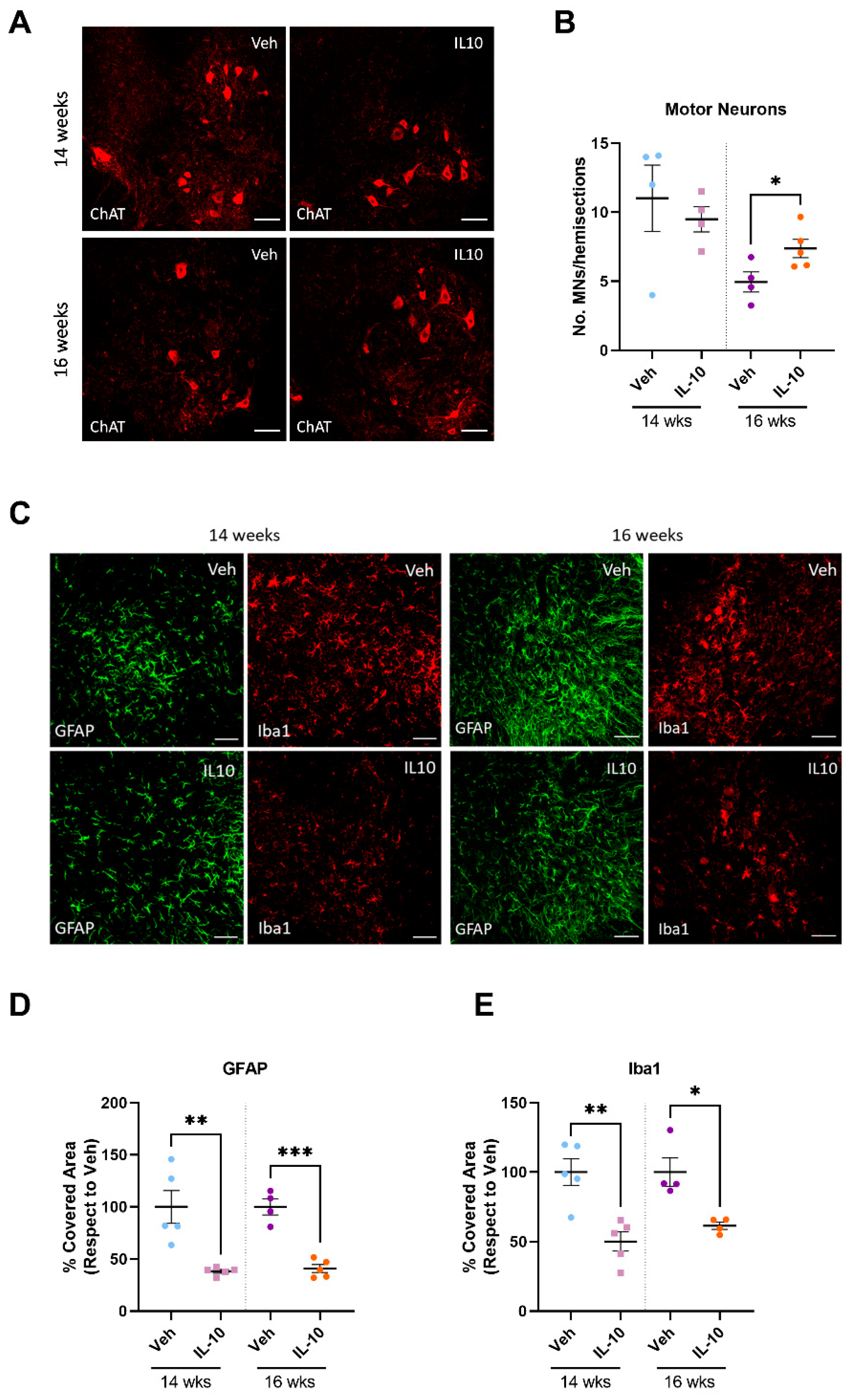
3.6. Intramuscular IL-10 Administration Promotes Spinal Motor Neuron Survival and Decreases Neuroinflammation in the Spinal Cord of C57-SOD1G93A Mice
3.7. Intramuscular IL-10 Administration Preserves Skeletal Muscle in Transgenic 129sv-SOD1G93A Mice Having Defective Immune Cell Recruitment
4. Discussion
4.1. The IL-10 Signaling Elicited the In Situ Proliferation and Polarization of MΦ toward an M2-Biased Phenotype in the Skeletal Muscle of C57-SOD1G93A Mice
4.2. The IL-10 Signaling Triggers Satellite Cell Differentiation in an Immune-Independent Manner
4.3. Preservation of Skeletal Muscle Influences Inflammation and Motor Neuron Survival within the Spinal Cord of C57-SOD1G93A Mice
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mejzini, R.; Flynn, L.L.; Pitout, I.L.; Fletcher, S.; Wilton, S.D.; Akkari, P.A. ALS Genetics, Mechanisms, and Therapeutics: Where Are We Now? Front. Neurosci. 2019, 13, 1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masrori, P.; van Damme, P. Amyotrophic Lateral Sclerosis: A Clinical Review. Eur. J. Neurol. 2020, 27, 1918–1929. [Google Scholar] [CrossRef] [PubMed]
- Moloney, E.B.; de Winter, F.; Verhaagen, J. ALS as a Distal Axonopathy: Molecular Mechanisms Affecting Neuromuscular Junction Stability in the Presymptomatic Stages of the Disease. Front. Neurosci. 2014, 8, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rocha, M.C.; Pousinha, P.A.; Correia, A.M.; Sebastião, A.M.; Ribeiro, J.A. Early Changes of Neuromuscular Transmission in the SOD1(G93A) Mice Model of ALS Start Long before Motor Symptoms Onset. PLoS ONE 2013, 8, e73846. [Google Scholar] [CrossRef] [Green Version]
- Fischer, L.R.; Glass, J.D. Axonal Degeneration in Motor Neuron Disease. Neurodegener. Dis. 2007, 4, 431–442. [Google Scholar] [CrossRef]
- Fischer, L.R.; Culver, D.G.; Tennant, P.; Davis, A.A.; Wang, M.; Castellano-Sanchez, A.; Khan, J.; Polak, M.A.; Glass, J.D. Amyotrophic Lateral Sclerosis Is a Distal Axonopathy: Evidence in Mice and Man. Exp. Neurol. 2004, 185, 232–240. [Google Scholar] [CrossRef]
- Scaricamazza, S.; Salvatori, I.; Ferri, A.; Valle, C. Skeletal Muscle in ALS: An Unappreciated Therapeutic Opportunity? Cells 2021, 10, 525. [Google Scholar] [CrossRef]
- Manzano, R.; Toivonen, J.M.; Moreno-Martínez, L.; Torre, M.; Moreno-García, L.; López-Royo, T.; Molina, N.; Zaragoza, P.; Calvo, A.C.; Osta, R. What Skeletal Muscle Has to Say in Amyotrophic Lateral Sclerosis: Implications for Therapy. Br. J. Pharmacol. 2021, 178, 1279–1297. [Google Scholar] [CrossRef]
- Dobrowolny, G.; Aucello, M.; Rizzuto, E.; Beccafico, S.; Mammucari, C.; Bonconpagni, S.; Belia, S.; Wannenes, F.; Nicoletti, C.; del Prete, Z.; et al. Skeletal Muscle Is a Primary Target of SOD1G93A-Mediated Toxicity. Cell Metab. 2008, 8, 425–436. [Google Scholar] [CrossRef]
- Wong, M.; Martin, L.J. Skeletal Muscle-Restricted Expression of Human SOD1 Causes Motor Neuron Degeneration in Transgenic Mice. Hum. Mol. Genet. 2010, 19, 2284–2302. [Google Scholar] [CrossRef] [Green Version]
- Rios, R.; Jablonka-Shariff, A.; Broberg, C.; Snyder-Warwick, A.K. Macrophage Roles in Peripheral Nervous System Injury and Pathology: Allies in Neuromuscular Junction Recovery. Mol. Cell. Neurosci. 2021, 111, 103590. [Google Scholar] [CrossRef]
- Nardo, G.; Trolese, M.C.; de Vito, G.; Cecchi, R.; Riva, N.; Dina, G.; Heath, P.R.; Quattrini, A.; Shaw, P.J.; Piazza, V.; et al. Immune Response in Peripheral Axons Delays Disease Progression in SOD1G93A Mice. J. Neuroinflammation 2016, 13, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiraishi, W.; Yamasaki, R.; Hashimoto, Y.; Ko, S.; Kobayakawa, Y.; Isobe, N.; Matsushita, T.; Kira, J.-I. Clearance of Peripheral Nerve Misfolded Mutant Protein by Infiltrated Macrophages Correlates with Motor Neuron Disease Progression. Sci. Rep. 2021, 11, 16438. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, S.; Schreiber, F.; Garz, C.; Debska-Vielhaber, G.; Assmann, A.; Perosa, V.; Petri, S.; Dengler, R.; Nestor, P.; Vielhaber, S. Toward in Vivo Determination of Peripheral Nervous System Immune Activity in Amyotrophic Lateral Sclerosis. Muscle Nerve 2019, 59, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Trolese, M.C.; Scarpa, C.; Melfi, V.; Fabbrizio, P.; Sironi, F.; Rossi, M.; Bendotti, C.; Nardo, G. Boosting the Peripheral Immune Response in the Skeletal Muscles Improved Motor Function in ALS Transgenic Mice. Mol. Ther. 2022, 30, 2760–2784. [Google Scholar] [CrossRef] [PubMed]
- Musarò, A. The Basis of Muscle Regeneration. Adv. Biol. 2014, 2014, 612471. [Google Scholar] [CrossRef] [Green Version]
- Tidball, J.G. Regulation of Muscle Growth and Regeneration by the Immune System. Nat. Rev. Immunol. 2017, 17, 165–178. [Google Scholar] [CrossRef]
- Oishi, Y.; Manabe, I. Macrophages in Inflammation, Repair and Regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Dort, J.; Fabre, P.; Molina, T.; Dumont, N.A. Macrophages Are Key Regulators of Stem Cells during Skeletal Muscle Regeneration and Diseases. Stem Cells Int. 2019, 2019, 4761427. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M.; Zhang, X. Interleukin-10: New Perspectives on an Old Cytokine. Immunol. Rev. 2008, 226, 205–218. [Google Scholar] [CrossRef]
- Villalta, S.A.; Rinaldi, C.; Deng, B.; Liu, G.; Fedor, B.; Tidball, J.G. Interleukin-10 Reduces the Pathology of Mdx Muscular Dystrophy by Deactivating M1 Macrophages and Modulating Macrophage Phenotype. Hum. Mol. Genet. 2011, 20, 790–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, M.; Papa, S.; Crippa, V.; Nardo, G.; Peviani, M.; Cheroni, C.; Trolese, M.C.; Lauranzano, E.; Bonetto, V.; Poletti, A.; et al. Differences in Protein Quality Control Correlate with Phenotype Variability in 2 Mouse Models of Familial Amyotrophic Lateral Sclerosis. Neurobiol. Aging 2015, 36, 492–504. [Google Scholar] [CrossRef] [PubMed]
- Nardo, G.; Iennaco, R.; Fusi, N.; Heath, P.R.; Marino, M.; Trolese, M.C.; Ferraiuolo, L.; Lawrence, N.; Shaw, P.J.; Bendotti, C. Transcriptomic Indices of Fast and Slow Disease Progression in Two Mouse Models of Amyotrophic Lateral Sclerosis. Brain 2013, 136, 3305–3332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madaro, L.; Torcinaro, A.; de Bardi, M.; Contino, F.F.; Pelizzola, M.; Diaferia, G.R.; Imeneo, G.; Bouchè, M.; Puri, P.L.; de Santa, F. Macrophages Fine Tune Satellite Cell Fate in Dystrophic Skeletal Muscle of Mdx Mice. PLoS Genet 2019, 15, e1008408. [Google Scholar] [CrossRef] [PubMed]
- Fabbrizio, P.; D’Agostino, J.; Margotta, C.; Mella, G.; Panini, N.; Pasetto, L.; Sammali, E.; Raggi, F.; Sorarù, G.; Bonetto, V.; et al. Contingent Intramuscular Boosting of P2XR7 Axis Improves Motor Function in Transgenic ALS Mice. Cell. Mol. Life Sci. 2022, 79, 7. [Google Scholar] [CrossRef] [PubMed]
- Jin, R.M.; Warunek, J.; Wohlfert, E.A. Therapeutic Administration of IL-10 and Amphiregulin Alleviates Chronic Skeletal Muscle Inflammation and Damage Induced by Infection. Immunohorizons 2018, 2, 142–154. [Google Scholar] [CrossRef] [Green Version]
- Deng, B.; Wehling-Henricks, M.; Villalta, S.A.; Wang, Y.; Tidball, J.G. IL-10 Triggers Changes in Macrophage Phenotype That Promote Muscle Growth and Regeneration. J. Immunol. 2012, 189, 3669–3680. [Google Scholar] [CrossRef] [Green Version]
- Chazaud, B.; Sonnet, C.; Lafuste, P.; Bassez, G.; Rimaniol, A.C.; Poron, F.; Authier, F.J.; Dreyfus, P.A.; Gherardi, R.K. Satellite Cells Attract Monocytes and Use Macrophages as a Support to Escape Apoptosis and Enhance Muscle Growth. J. Cell Biol. 2003, 163, 1133–1143. [Google Scholar] [CrossRef] [Green Version]
- Friese, A.; Kaltschmidt, J.A.; Ladle, D.R.; Sigrist, M.; Jessell, T.M.; Arber, S. Gamma and Alpha Motor Neurons Distinguished by Expression of Transcription Factor Err3. Proc. Natl. Acad. Sci. USA 2009, 106, 13588–13593. [Google Scholar] [CrossRef] [Green Version]
- White, P.; Liebhaber, S.A.; Cooke, N.E. 129X1/SvJ Mouse Strain Has a Novel Defect in Inflammatory Cell Recruitment. J. Immunol. 2002, 168, 869–874. [Google Scholar] [CrossRef] [Green Version]
- Tsitkanou, S.; della Gatta, P.A.; Russell, A.P. Skeletal Muscle Satellite Cells, Mitochondria, and MicroRNAs: Their Involvement in the Pathogenesis of ALS. Front. Physiol. 2016, 7, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chazaud, B. Inflammation and Skeletal Muscle Regeneration: Leave It to the Macrophages! Trends Immunol. 2020, 41, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, R.P.; Sheikh, F.; Kotenko, S.V.; Dickensheets, H. The Expanded Family of Class II Cytokines That Share the IL-10 Receptor-2 (IL-10R2) Chain. J. Leukoc. Biol. 2004, 76, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Prasse, A.; Germann, M.; Pechkovsky, D.V.; Markert, A.; Verres, T.; Stahl, M.; Melchers, I.; Luttmann, W.; Müller-Quernheim, J.; Zissel, G. IL-10-Producing Monocytes Differentiate to Alternatively Activated Macrophages and Are Increased in Atopic Patients. J. Allergy Clin. Immunol. 2007, 119, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, T.; Matsui, N.; Fujita, K.; Nodera, H.; Shimizu, F.; Miyamoto, K.; Takahashi, Y.; Kanda, T.; Kusunoki, S.; Izumi, Y.; et al. CSF Cytokine Profile Distinguishes Multifocal Motor Neuropathy from Progressive Muscular Atrophy. Neurol. Neuroimmunol. Neuroinflammation 2015, 2, e138. [Google Scholar] [CrossRef] [Green Version]
- Ayers, J.I.; Fromholt, S.; Sinyavskaya, O.; Siemienski, Z.; Rosario, A.M.; Li, A.; Crosby, K.W.; Cruz, P.E.; Dinunno, N.M.; Janus, C.; et al. Widespread and Efficient Transduction of Spinal Cord and Brain Following Neonatal AAV Injection and Potential Disease Modifying Effect in ALS Mice. Mol. Ther. 2015, 23, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Strickland, M.R.; Ibanez, K.R.; Yaroshenko, M.; Diaz, C.C.; Borchelt, D.R.; Chakrabarty, P. IL-10 Based Immunomodulation Initiated at Birth Extends Lifespan in a Familial Mouse Model of Amyotrophic Lateral Sclerosis. Sci. Rep. 2020, 10, 20862. [Google Scholar] [CrossRef]
- Yang, W.; Hu, P. Skeletal Muscle Regeneration Is Modulated by Inflammation. J. Orthop. Translat. 2018, 13, 25–32. [Google Scholar] [CrossRef]
- Sato, Y.; Ohshima, T.; Kondo, T. Regulatory Role of Endogenous Interleukin-10 in Cutaneous Inflammatory Response of Murine Wound Healing. Biochem. Biophys. Res. Commun. 1999, 265, 194–199. [Google Scholar] [CrossRef]
- Jenkins, S.J.; Ruckerl, D.; Cook, P.C.; Jones, L.H.; Finkelman, F.D.; van Rooijen, N.; MacDonald, A.S.; Allen, J.E. Local Macrophage Proliferation, Rather than Recruitment from the Blood, Is a Signature of TH2 Inflammation. Science 2011, 332, 1284–1288. [Google Scholar] [CrossRef] [Green Version]
- Welc, S.S.; Wehling-Henricks, M.; Antoun, J.; Ha, T.T.; Tous, I.; Tidball, J.G. Differential Effects of Myeloid Cell PPARδ and IL-10 in Regulating Macrophage Recruitment, Phenotype, and Regeneration Following Acute Muscle Injury. J. Immunol. 2020, 205, 1664–1677. [Google Scholar] [CrossRef] [PubMed]
- Pikatza-Menoio, O.; Elicegui, A.; Bengoetxea, X.; Naldaiz-Gastesi, N.; López de Munain, A.; Gerenu, G.; Gil-Bea, F.J.; Alonso-Martín, S. The Skeletal Muscle Emerges as a New Disease Target in Amyotrophic Lateral Sclerosis. J. Pers. Med. 2021, 11, 671. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, J.T.; King, K.Y.; Brett, J.O.; Cromie, M.J.; Charville, G.W.; Maguire, K.K.; Brunson, C.; Mastey, N.; Liu, L.; Tsai, C.-R.; et al. MTORC1 Controls the Adaptive Transition of Quiescent Stem Cells from G0 to GAlert. Nature 2014, 510, 393–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, J.T.; Schroeder, M.D.; Ma, C.; Rando, T.A. HGFA Is an Injury-Regulated Systemic Factor That Induces the Transition of Stem Cells into G Alert. Cell. Rep. 2017, 19, 479–486. [Google Scholar] [CrossRef]
- Tatsumi, R.; Sankoda, Y.; Anderson, J.E.; Sato, Y.; Mizunoya, W.; Shimizu, N.; Suzuki, T.; Yamada, M.; Rhoads, R.P.; Ikeuchi, Y.; et al. Possible Implication of Satellite Cells in Regenerative Motoneuritogenesis: HGF Upregulates Neural Chemorepellent Sema3A during Myogenic Differentiation. Am. J. Physiol. Cell Physiol. 2009, 297, C238–C252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCombe, P.A.; Henderson, R.D. The Role of Immune and Inflammatory Mechanisms in ALS. Curr. Mol. Med. 2011, 11, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Wosiski-Kuhn, M.; Lyon, M.S.; Caress, J.; Milligan, C. Inflammation, Immunity, and Amyotrophic Lateral Sclerosis: II. Immune-Modulating Therapies. Muscle Nerve 2019, 59, 23–33. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fabbrizio, P.; Margotta, C.; D’Agostino, J.; Suanno, G.; Quetti, L.; Bendotti, C.; Nardo, G. Intramuscular IL-10 Administration Enhances the Activity of Myogenic Precursor Cells and Improves Motor Function in ALS Mouse Model. Cells 2023, 12, 1016. https://doi.org/10.3390/cells12071016
Fabbrizio P, Margotta C, D’Agostino J, Suanno G, Quetti L, Bendotti C, Nardo G. Intramuscular IL-10 Administration Enhances the Activity of Myogenic Precursor Cells and Improves Motor Function in ALS Mouse Model. Cells. 2023; 12(7):1016. https://doi.org/10.3390/cells12071016
Chicago/Turabian StyleFabbrizio, Paola, Cassandra Margotta, Jessica D’Agostino, Giuseppe Suanno, Lorenzo Quetti, Caterina Bendotti, and Giovanni Nardo. 2023. "Intramuscular IL-10 Administration Enhances the Activity of Myogenic Precursor Cells and Improves Motor Function in ALS Mouse Model" Cells 12, no. 7: 1016. https://doi.org/10.3390/cells12071016
APA StyleFabbrizio, P., Margotta, C., D’Agostino, J., Suanno, G., Quetti, L., Bendotti, C., & Nardo, G. (2023). Intramuscular IL-10 Administration Enhances the Activity of Myogenic Precursor Cells and Improves Motor Function in ALS Mouse Model. Cells, 12(7), 1016. https://doi.org/10.3390/cells12071016