
Insights on the Biomarker Potential of Exosomal Non-Coding RNAs in Colorectal Cancer: An In Silico Characterization of Related Exosomal lncRNA/circRNA–miRNA–Target Axis


 , ,
, , 
Abstract
:1. Introduction
2. Search Strategy
3. Exosomal Non-Coding RNAs as Potential Biomarkers in CRC
3.1. Diagnostic Biomarker Potential of Exosomal ncRNAs in CRC
3.2. Diagnostic and Prognostic Biomarker Potential of Exosomal ncRNAs in CRC
3.3. Prognostic/Predictive Biomarker Potential of Exosomal ncRNAs in CRC

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Coding RNA(s) | Non-Coding RNA Class | Number of CRC Patients | Source | Biomarker Potential | Status | Clinical Evidence | References |
|---|---|---|---|---|---|---|---|
| miR-208b | miRNA | N/A | Serum | Predictive | Upregulated in non-responders compared to responders | Yes, resistance to FOLFOX therapy in CRC patients | [57] |
| miR-934 | MiRNA | N/A | Serum | Prognostic | Upregulated in stage IV CRC compared to stage I CRC | Yes, promotes CRLM by regulating the crosstalk between CRC cells and TAMs | [44] |
| miR-25-3p | MiRNA | N/A | Serum | Prognostic | Upregulated in metastatic CRC tissues compared to those without metastasis | Yes, involved in pre-metastatic niche formation | [45] |
| miR-135a-5p | MiRNA | N/A | Serum | Prognostic | Upregulated in CRC tissues post metastasis compared to those premetastasis | Yes, correlates with the development, clinical severity, and prognosis of CRC liver metastases through the premetastatic niche | [50] |
| Circ-133 | CircRNA | N/A | Plasma | Prognostic | Upregulated in advanced CRC stages compared to early stage | Yes, promotes CRC metastasis by acting on miR-133a/GEF-H1/RhoA axis | [55] |
| miR-27b-3p | MiRNA | N/A | Serum | Prognostic | Upregulated in metastatic CRC patients compared to non-metastatic | Yes, promotes metastasis, increases blood vessel permeability and facilitate the generation of CTCs | [51] |
| miR-106b-5p | MiRNA | N/A | Plasma | Prognostic | Upregulated in CRC patients compared to normal ones | Yes, mediates CRC invasion, migration and metastasis | [52] |
| miR-6869-5p | miRNA | N/A | Serum | Prognostic | Downregulated in advanced CRC patients | Yes, inhibits cell proliferation and the production of inflammatory cytokines | [53] |
| RPPH1 | LncRNA | 52 | Plasma | Prognostic | Upregulated in treatment-naïve CRC patients compared to post tumor resection | Yes, promotes CRC metastasis | [38] |
| HOTTIP | LncRNA | 100 | Serum | Prognostic | Downregulated in CRC patients compared to healthy donors | Yes, associated with worse prognosis and poor overall survival | [39] |
| NNT-AS1 | LncRNA | 40 | Serum | Prognostic | Upregulated in preoperative compared to postoperative CRC samples and in advanced stage compared to early stage CRC patients | Yes, promotes CRC proliferation, migration, and invasion in CRC patients | [14] |
| circ0000338 | CircRNA | 17 | Serum | Predictive | Upregulated in chemoresistant CRC patients compared to chemosensitive | Yes, confers chemoresistance in CRC patients | [58] |
| hsa-circ-0005100 (circFMN2) | CircRNA | 35 | Serum | Prognostic | Upregulated in CRC patients compared to healthy ones | Yes, promotes tumor proliferation in CRC patients | [54] |
| miR-200c | miRNA | N/A | Serum | Prognostic/Predictive | Upregulated in advanced compared to early stage CRC patients | No | [61] |
| miR-6803-5p | miRNA | 168 | Serum | Prognostic | Upregulated in CRC patients compared to healthy controls | Yes, correlated with poor prognosis and low OS in CRC patients | [34] |
| miR-193a-5p | miRNA | 25 | Plasma | Prognostic | Upregulated in advanced CRC stage compared to early stage | Yes, promotes tumor progression in CRC patients | [46] |
| miR-146a-5p | MiRNA | 53 | Serum | Prognostic | Upregulated in CRC patients compared to normal | Yes, promotes tumor stemness expansion | [47] |
| miR-150-5p | MiRNA | 133 | Serum | Prognostic | Downregulated in preoperative CRC patient samples compared to postoperative ones | Yes, low levels correlate with poor prognosis, worse OS, and shorter survival | [32] |
| miR-199a/b-3p, miR-199a-5p and miR-199b-5p | MiRNA | 89 | Serum | Prognostic and Predictive | Downregulated in advanced CRC stage compared to early stage | Yes, upregulated levels are associated with superior RFS and OS | [60] |
| miR-874 | MiRNA | 125 | Serum | Prognostic | Downregulated in CRC patients compared to benign AD | Yes, low levels correlate with positive distant metastasis, lymph node metastasis, poor differentiation and advanced TNM stage | [30] |
| miR-221/222 | MiRNA | N/A | Serum | Prognostic | Upregulated in metastatic CRC patient samples compared to those without metastasis | Yes, promotes liver metastasis in CRC patients | [48] |
| miR-106b-3p | miRNA | 80 | Serum | Prognostic | Upregulated in patients with metastasis compared to those without metastasis | Yes, promotes CRC metastasis and high serum level correlate with poor prognosis | [49] |
| circPACRGL | circRNA | N/A | Serum | Prognostic | Upregulated in CRC patients | Yes, promotes CRC progression and metastasis | [56] |
| circ_0005963 | circRNA | 19 | Serum | Prognostic | Upregulated in oxaliplatin resistant patients compared to oxaliplatin sensitive patients | Yes, promotes glycolysis and oxaliplatin resistance | [59] |
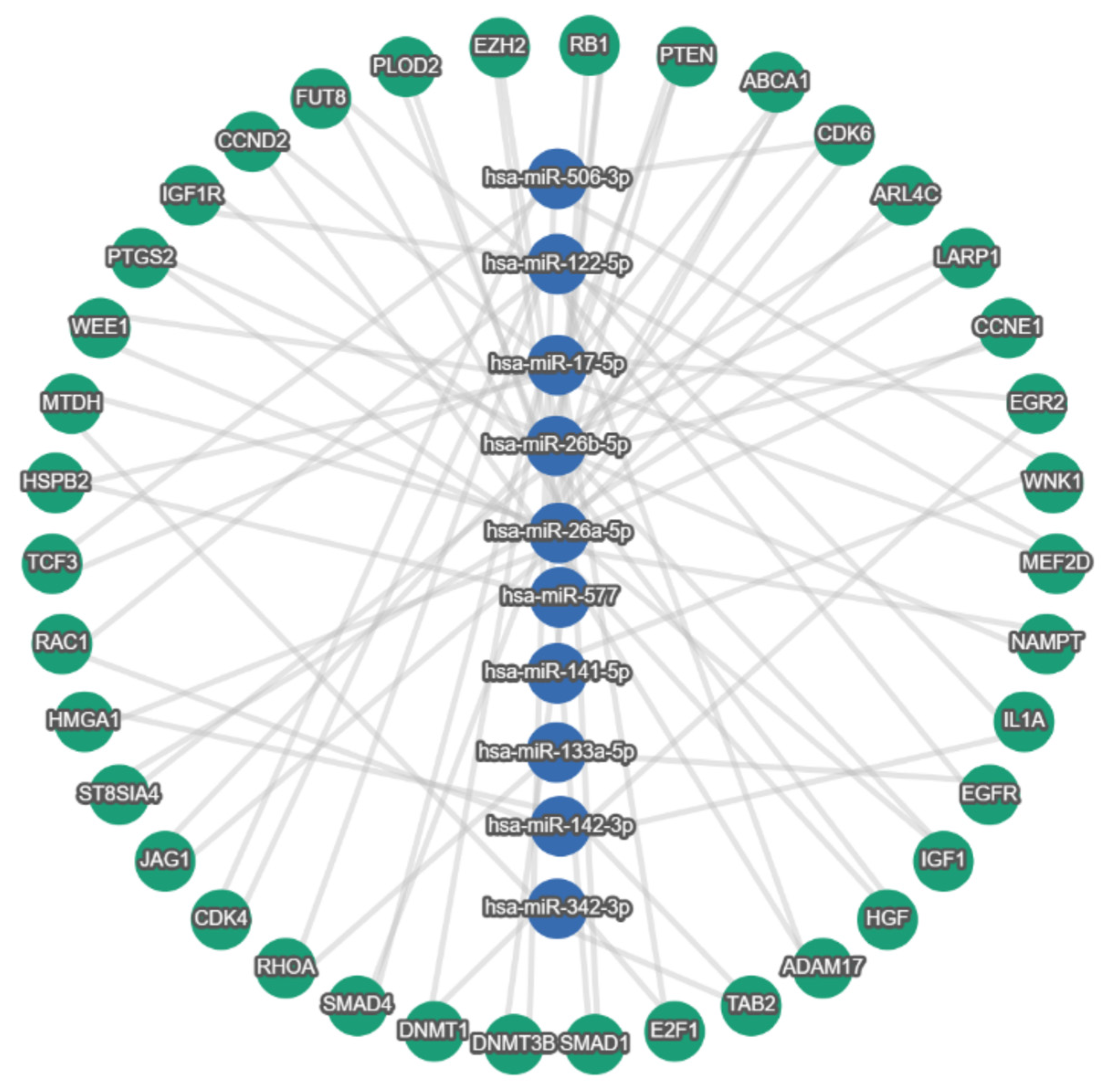
4. In Silico Analysis of CRC Exosomal lncRNA/circRNA–miRNA–Target Axis
4.1. Rationale
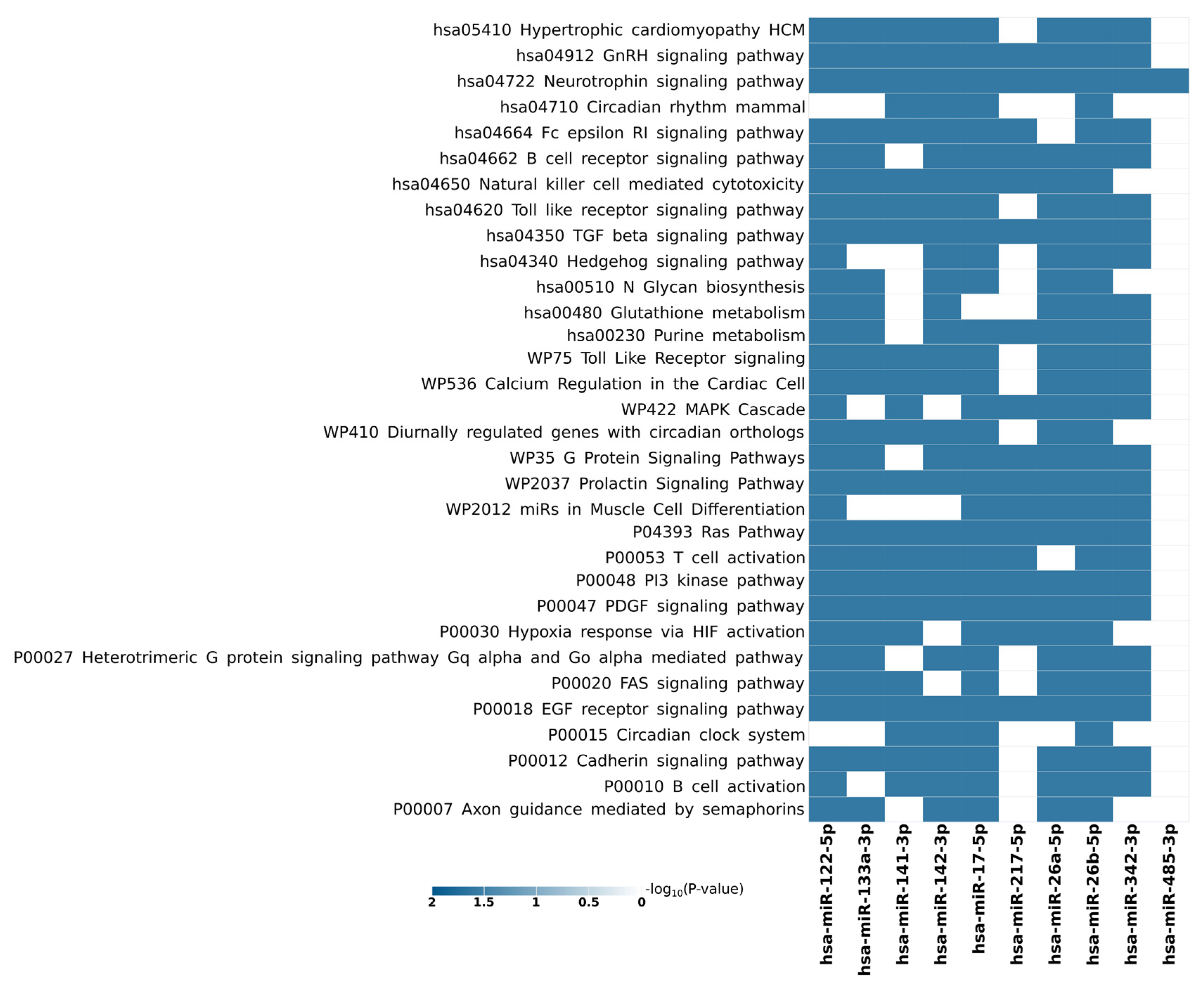
4.2. Shared Target Genes and Pathways
4.3. Gene Ontology and KEGG Pathways
5. Discussion
5.1. Advantages of Exosome-Derived Biomarkers
5.2. Limitations and Challenges of Exosome-Derived Biomarkers
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Wong, M.C.; Ding, H.; Wang, J.; Chan, P.S.; Huang, J. Prevalence and risk factors of colorectal cancer in Asia. Intest. Res. 2019, 17, 317–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, T.; Zhang, Y.; Wang, X.; Liu, W.; Feng, W.; Song, W. Integrated analysis of the functions and clinical implications of exosome circRNAs in colorectal cancer. Front. Immunol. 2022, 13, 919014. [Google Scholar] [CrossRef] [PubMed]
- Rapado-González, Ó.; Álvarez-Castro, A.; López-López, R.; Iglesias-Canle, J.; Suárez-Cunqueiro, M.M.; Muinelo-Romay, L. Circulating microRNAs as Promising Biomarkers in Colorectal Cancer. Cancers 2019, 11, 898. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.R.; Kwon, H.N.; Nam, H.; Kim, J.J.; Park, S.; Kim, Y.-H. Urine-NMR metabolomics for screening of advanced colorectal adenoma and early stage colorectal cancer. Sci. Rep. 2019, 9, 4786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toiyama, Y.; Okugawa, Y.; Fleshman, J.; Boland, C.R.; Goel, A. MicroRNAs as potential liquid biopsy biomarkers in colorectal cancer: A systematic review. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2018, 1870, 274–282. [Google Scholar] [CrossRef]
- Baassiri, A.; Nassar, F.; Mukherji, D.; Shamseddine, A.; Nasr, R.; Temraz, S. Exosomal Non Coding RNA in LIQUID Biopsies as a Promising Biomarker for Colorectal Cancer. Int. J. Mol. Sci. 2020, 21, 1398. [Google Scholar] [CrossRef] [Green Version]
- Walsh, J.M.E.; Terdiman, J.P. Colorectal Cancer Screening. JAMA 2003, 289, 1297. [Google Scholar] [CrossRef] [Green Version]
- Hon, K.W.; Abu, N.; Ab Mutalib, N.-S.; Jamal, R. Exosomes as Potential Biomarkers and Targeted Therapy in Colorectal Cancer: A Mini-Review. Front. Pharmacol. 2017, 8, 583. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Han, B.; Gao, S.; Wang, X.; Wang, Z.; Wang, F.; Zhang, J.; Xu, D.; Sun, B. Exosome-encapsulated microRNAs as circulating biomarkers for colorectal cancer. Oncotarget 2017, 8, 60149–60158. [Google Scholar] [CrossRef] [Green Version]
- De Rubis, G.; Krishnan, S.R.; Bebawy, M. Liquid Biopsies in Cancer Diagnosis, Monitoring, and Prognosis. Trends Pharmacol. Sci. 2019, 40, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Aalami, A.H.; Abdeahad, H.; Mesgari, M. Circulating miR-21 as a potential biomarker in human digestive system carcinoma: A systematic review and diagnostic meta-analysis. Biomarkers 2021, 26, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Aalami, A.H.; Mesgari, M.; Sahebkar, A. Synthesis and Characterization of Green Zinc Oxide Nanoparticles with Antiproliferative Effects through Apoptosis Induction and MicroRNA Modulation in Breast Cancer Cells. Bioinorg. Chem. Appl. 2020, 2020, 8817110. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Hu, J.; Ye, Z.; Chen, S.; Chen, Y. Serum long non-coding RNA NNT–AS1 protected by exosome is a potential biomarker and functions as an oncogene via the miR-496/RAP2C axis in colorectal cancer. Mol. Med. Rep. 2021, 24, 585. [Google Scholar] [CrossRef]
- Han, D.; Wang, M.; Ma, N.; Xu, Y.; Jiang, Y.; Gao, X. Long noncoding RNAs: Novel players in colorectal cancer. Cancer Lett. 2015, 361, 13–21. [Google Scholar] [CrossRef]
- Li, C.; Li, X. Exosome-Derived Circ_0094343 Promotes Chemosensitivity of Colorectal Cancer Cells by Regulating Glycolysis via the miR-766-5p/TRIM67 Axis. Contrast Media Mol. Imaging 2022, 2022, 2878557. [Google Scholar] [CrossRef]
- Ashekyan, O.; Abdallah, S.; Shoukari, A.A.; Chamandi, G.; Choubassy, H.; Itani, A.R.S.; Nasr, R. Spotlight on Exosomal Non-Coding RNAs in Breast Cancer: An In Silico Analysis to Identify Potential lncRNA/circRNA-miRNA-Target Axis. Int. J. Mol. Sci. 2022, 23, 8351. [Google Scholar] [CrossRef]
- Su, K.; Wang, N.; Shao, Q.; Liu, H.; Zhao, B.; Ma, S. The role of a ceRNA regulatory network based on lncRNA MALAT1 site in cancer progression. Biomed. Pharmacother. 2021, 137, 111389. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yao, W.; Liu, R.; Gao, L.; Lu, Y.; Zhang, H.; Liang, X. Long non-coding RNA LINC00152 in cancer: Roles, mechanisms, and chemotherapy and radiotherapy resistance. Front. Oncol. 2022, 12, 193. [Google Scholar] [CrossRef]
- Ogata-Kawata, H.; Izumiya, M.; Kurioka, D.; Honma, Y.; Yamada, Y.; Furuta, K.; Gunji, T.; Ohta, H.; Okamoto, H.; Sonoda, H.; et al. Circulating Exosomal microRNAs as Biomarkers of Colon Cancer. PLoS ONE 2014, 9, e92921. [Google Scholar] [CrossRef] [PubMed]
- Dokhanchi, M.; Pakravan, K.; Zareian, S.; Hussen, B.M.; Farid, M.; Razmara, E.; Mossahebi-Mohammadi, M.; Cho, W.C.; Babashah, S. Colorectal cancer cell-derived extracellular vesicles transfer miR-221-3p to promote endothelial cell angiogenesis via targeting suppressor of cytokine signaling 3. Life Sci. 2021, 285, 119937. [Google Scholar] [CrossRef] [PubMed]
- Min, L.; Chen, L.; Liu, S.; Yu, Y.; Guo, Q.; Li, P.; Zhu, S. Loss of Circulating Exosomal miR-92b is a Novel Biomarker of Colorectal Cancer at Early Stage. Int. J. Med. Sci. 2019, 16, 1231–1237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Chen, Y.; Guo, X.; Zhou, L.; Jia, Z.; Peng, Z.; Ren, C. GPC 1 exosome and its regulatory mi RNA s are specific markers for the detection and target therapy of colorectal cancer. J. Cell. Mol. Med. 2017, 21, 838–847. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Li, Y.; Zheng, X.; Gan, J.; Wan, Z.; Liu, Y. Exosomal-miR-10a derived from colorectal cancer cells suppresses migration of human lung fibroblasts, and expression of IL-6, IL-8 and IL-1β. Mol. Med. Rep. 2021, 23, 1. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, X.; Yu, M.; Niu, L.; Zhao, Y.; Tang, Y.; Zheng, B.; Song, X.; Xie, L. Serum exosomal miR-377-3p and miR-381-3p as diagnostic biomarkers in colorectal cancer. Futur. Oncol. 2022, 18, 793–805. [Google Scholar] [CrossRef]
- Handa, T.; Kuroha, M.; Nagai, H.; Shimoyama, Y.; Naito, T.; Moroi, R.; Masamune, A. Liquid biopsy for colorectal adenoma: Is the exosomal miRNA derived from organoid a potential diagnostic biomarker? Clin. Transl. Gastroenterol. 2021, 12, e00356. [Google Scholar] [CrossRef]
- Wang, L.; Duan, W.; Yan, S.; Xie, Y.; Wang, C. Circulating long non-coding RNA colon cancer-associated transcript 2 protected by exosome as a potential biomarker for colorectal cancer. Biomed. Pharmacother. 2019, 113, 108758. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhan, Y.; Zhu, K.; Bai, M.; Han, J.; Si, Y.; Zhang, H.; Kong, D. Plasma Exosomal Long Non-Coding RNAs Serve as Biomarkers for Early Detection of Colorectal Cancer. Cell. Physiol. Biochem. 2018, 51, 2704–2715. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, P.-P.; Huang, J.-J.; Wang, Z.-Y.; Zhang, Z.-H.; Yuan, J.-Z.; Ma, E.-M.; Liu, X.; Bai, J. Reduced serum exosomal miR-874 expression predicts poor prognosis in colorectal cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 664–672. [Google Scholar]
- Chen, H.-L.; Li, J.J.; Jiang, F.; Shi, W.J.; Chang, G.Y. MicroRNA-4461 derived from bone marrow mesenchymal stem cell exosomes inhibits tumorigenesis by downreg-ulating COPB2 expression in colorectal cancer. Biosci. Biotechnol. Biochem. 2020, 84, 338–346. [Google Scholar] [CrossRef]
- Zou, S.-L.; Chen, Y.-L.; Ge, Z.-Z.; Qu, Y.-Y.; Cao, Y.; Kang, Z.-X. Downregulation of serum exosomal miR-150-5p is associated with poor prognosis in patients with colorectal cancer. Cancer Biomark. 2019, 26, 69–77. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, S.; Lai, Q.; Fang, Y.; Wu, C.; Liu, Y.; Li, Q.; Wang, X.; Gu, C.; Chen, J.; et al. Cancer-associated fibroblasts-derived exosomal miR-17-5p promotes colorectal cancer aggressive phenotype by initiating a RUNX3/MYC/TGF-β1 positive feedback loop. Cancer Lett. 2020, 491, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Jiang, Y.; Liang, C.; Cheng, M.; Jin, C.; Duan, Q.; Jin, P. Exosomal miR-6803-5p as potential diagnostic and prognostic marker in colorectal cancer. J. Cell. Biochem. 2018, 119, 4113–4119. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yan, F.; Zhao, Q.; Zhan, F.; Wang, R.; Wang, L.; Zhang, Y.; Huang, X. Circulating exosomal miR-125a-3p as a novel biomarker for early-stage colon cancer. Sci. Rep. 2017, 7, 4150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.; Ji, X.; Liu, K.; Shi, Y.; Wang, C.; Li, Y.; Zhang, T.; He, Y.; Xiang, M.; Zhao, R. Exosomal miR-200c-3p negatively regulates the migration and invasion of lipopolysaccharide (LPS)-stimulated colo-rectal cancer (CRC). BMC Mol. Cell Biol. 2020, 21, 48. [Google Scholar] [CrossRef]
- Dai, W.; Zhou, J.; Wang, H.; Zhang, M.; Yang, X.; Song, W. miR-424-5p promotes the proliferation and metastasis of colorectal cancer by directly targeting SCN4B. Pathol. Res. Pract. 2020, 216, 152731. [Google Scholar] [CrossRef]
- Liang, Z.-X.; Liu, H.S.; Wang, F.W.; Xiong, L.; Zhou, C.; Hu, T.; He, X.W.; Wu, X.J.; Xie, D.; Wu, X.R.; et al. LncRNA RPPH1 promotes colorectal cancer metastasis by interacting with TUBB3 and by promoting exo-somes-mediated macrophage M2 polarization. Cell Death Dis. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Oehme, F.; Krahl, S.; Gyorffy, B.; Muessle, B.; Rao, V.; Greif, H.; Ziegler, N.; Lin, K.; Thepkaysone, M.-L.; Polster, H.; et al. Low level of exosomal long non-coding RNA HOTTIP is a prognostic biomarker in colorectal cancer. RNA Biol. 2019, 16, 1339–1345. [Google Scholar] [CrossRef]
- Li, N.; Li, J.; Mi, Q.; Xie, Y.; Li, P.; Wang, L.; Wang, C. Long non-coding RNA ADAMTS9-AS1 suppresses colorectal cancer by inhibiting the Wnt/β-catenin signalling pathway and is a potential diagnostic biomarker. J. Cell. Mol. Med. 2020, 24, 11318–11329. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, X.; Gao, S.; Jing, F.; Yang, Y.; Du, L.; Zheng, G.; Li, P.; Li, C.; Wang, C. Exosomal long noncoding RNA CRNDE-h as a novel serum-based biomarker for diagnosis and prognosis of colorectal cancer. Oncotarget 2016, 7, 85551–85563. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Meng, T.; Yang, X.-H.; Sayim, P.; Lei, C.; Jin, B.; Ge, L.; Wang, H.-J. Prognostic and predictive value of long non-coding RNA GAS5 and mircoRNA-221 in colorectal cancer and their effects on colorectal cancer cell proliferation, migration and invasion. Cancer Biomark. 2018, 22, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Zhang, K.; Tan, S.; Gao, F.; Zhang, Y.; Xu, W.; Wang, H.; Gu, D.; Zhu, L.; Li, S.; et al. Exosomal circLPAR1 functions in colorectal cancer diagnosis and tumorigenesis through suppressing BRD4 via METTL3–eIF3h interaction. Mol. Cancer 2022, 21, 49. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Mi, Y.; Guan, B.; Zheng, B.; Wei, P.; Gu, Y.; Zhang, Z.; Cai, S.; Xu, Y.; Li, X.; et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J. Hematol. Oncol. 2020, 13, 156. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Li, Y.; Pan, Y.; Lan, X.; Song, F.; Sun, J.; Zhou, K.; Liu, X.; Ren, X.; Wang, F.; et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat. Commun. 2018, 9, 5395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, Y.; Ren, Y.; Hu, X.; Mu, J.; Samykutty, A.; Zhuang, X.; Deng, Z.; Kumar, A.; Zhang, L.; Merchant, M.L.; et al. MVP-mediated exosomal sorting of miR-193a promotes colon cancer progression. Nat. Commun. 2017, 8, 14448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.C.; Liao, T.T.; Lin, C.C.; Yuan, L.T.E.; Lan, H.Y.; Lin, H.H.; Teng, H.W.; Chang, H.C.; Lin, C.H.; Yang, C.Y.; et al. RAB27B-activated secretion of stem-like tumor exosomes delivers the biomarker microRNA-146a-5p, which promotes tumorigenesis and associates with an immunosuppressive tumor microenvironment in colorectal cancer. Int. J. Cancer 2019, 145, 2209–2224. [Google Scholar] [CrossRef]
- Tian, F.; Wang, P.; Lin, D.; Dai, J.; Liu, Q.; Guan, Y.; Zhu, K. Exosome-delivered miR-221/222 exacerbates tumor liver metastasis by targeting SPINT1 in colorectal cancer. Cancer Sci. 2021, 112, 3744. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Sun, P.; Leng, K.; Xu, Y.; Mei, L.; Han, P.; Zhang, B.; Yao, K.; Li, C.; et al. Colorectal cancer-derived exosomal miR-106b-3p promotes metastasis by down-regulating DLC-1 expression. Clin. Sci. 2020, 134, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Meng, Q.; Shi, C.; Yang, H.; Li, X.; Wu, S.; Chen, R. Hypoxia-Inducible Exosomes Facilitate Liver-Tropic Premetastatic Niche in Colorectal Cancer. Hepatology 2021, 74, 2633–2651. [Google Scholar] [CrossRef]
- Dou, R.; Liu, K.; Yang, C.; Zheng, J.; Shi, D.; Lin, X.; Xiong, B. EMT-cancer cells-derived exosomal miR-27b-3p promotes circulating tumour cells-mediated metastasis by modulating vascular permeability in colorectal cancer. Clin. Transl. Med. 2021, 11, e595. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Dou, R.; Wei, C.; Liu, K.; Shi, D.; Zhang, C.; Liu, Q.; Wang, S.; Xiong, B. Tumor-derived exosomal microRNA-106b-5p activates EMT-cancer cell and M2-subtype TAM interaction to facilitate CRC metastasis. Mol. Ther. 2021, 29, 2088–2107. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Liu, G.; Jin, C.; Wang, Z.; Duan, Q.; Xu, J.; Xu, D. MicroRNA-6869-5p acts as a tumor suppressor via targeting TLR4/NF-κB signaling pathway in colorectal cancer. J. Cell. Physiol. 2018, 233, 6660–6668. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, C.; Xu, R.; Wang, Y.; Li, D.; Zhang, B. A novel circFMN2 promotes tumor proliferation in CRC by regulating the miR-1182/hTERT signaling pathways. Clin. Sci. 2019, 133, 2463–2479. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhang, H.; Yang, Y.; Wang, X.; Deng, T.; Liu, R.; Ba, Y. Hypoxia induced exosomal circRNA promotes metastasis of Colorectal Cancer via targeting GEF-H1/RhoA axis. Theranostics 2020, 10, 8211–8226. [Google Scholar] [CrossRef]
- Shang, A.; Gu, C.; Wang, W.; Wang, X.; Sun, J.; Zeng, B.; Chen, C.; Chang, W.; Ping, Y.; Ji, P.; et al. Exosomal circPACRGL promotes progression of colorectal cancer via the miR-142-3p/miR-506-3p- TGF-β1 axis. Mol. Cancer 2020, 19, 117. [Google Scholar] [CrossRef]
- Ning, T.; Li, J.; He, Y.; Zhang, H.; Wang, X.; Deng, T.; Liu, R.; Li, H.; Bai, M.; Fan, Q.; et al. Exosomal miR-208b related with oxaliplatin resistance promotes Treg expansion in colorectal cancer. Mol. Ther. 2021, 29, 2723–2736. [Google Scholar] [CrossRef]
- Hon, K.W.; Ab-Mutalib, N.S.; Abdullah, N.M.A.; Jamal, R.; Abu, N. Extracellular Vesicle-derived circular RNAs confers chemoresistance in Colorectal cancer. Sci. Rep. 2019, 9, 16497. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Ba, Y. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef] [Green Version]
- Baek, D.W.; Kim, G.; Kang, B.W.; Kim, H.J.; Park, S.Y.; Park, J.S.; Choi, G.-S.; Kang, M.K.; Hur, K.; Kim, J.G. High expression of microRNA-199a-5p is associated with superior clinical outcomes in patients with locally advanced rectal cancer. J. Cancer Res. Clin. Oncol. 2019, 146, 105–115. [Google Scholar] [CrossRef]
- Toiyama, Y.; Hur, K.; Tanaka, K.; Inoue, Y.; Kusunoki, M.; Boland, C.R.; Goel, A. Serum miR-200c Is a Novel Prognostic and Metastasis-Predictive Biomarker in Patients with Colorectal Cancer. Ann. Surg. 2014, 259, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kern, F.; Aparicio-Puerta, E.; Li, Y.; Fehlmann, T.; Kehl, T.; Wagner, V.; Keller, A. miRTargetLink 2.0-interactive miRNA target gene and target pathway networks. Nucleic Acids Res. 2021, 49, W409–W416. [Google Scholar] [CrossRef]
- Bai, L.; Gao, Z.; Jiang, A.; Ren, S.; Wang, B. Circular noncoding RNA circ_0007334 sequestrates miR-577 to derepress KLF12 and accelerate colorectal cancer progression. Anti-Cancer Drugs 2022, 33, e409–e422. [Google Scholar] [CrossRef] [PubMed]
- Han, K.; Wang, F.-W.; Cao, C.-H.; Ling, H.; Chen, J.-W.; Chen, R.-X.; Feng, Z.-H.; Luo, J.; Jin, X.-H.; Duan, J.-L.; et al. CircLONP2 enhances colorectal carcinoma invasion and metastasis through modulating the maturation and exosomal dissemination of microRNA-17. Mol. Cancer 2020, 19, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Ding, L.; Zhang, D.; Shi, G.; Xu, Q.; Shen, S.; Wang, Y.; Wang, T.; Hou, Y. Carcinoma-associated fibroblasts promote the stemness and chemoresistance of colorectal cancer by transferring exosomal lncRNA H19. Theranostics 2018, 8, 3932–3948. [Google Scholar] [CrossRef]
- Xu, J.; Xiao, Y.; Liu, B.; Pan, S.; Liu, Q.; Shan, Y.; Li, S.; Qi, Y.; Huang, Y.; Jia, L. Exosomal MALAT1 sponges miR-26a/26b to promote the invasion and metastasis of colorectal cancer via FUT4 enhanced fucosylation and PI3K/Akt pathway. J. Exp. Clin. Cancer Res. 2020, 39, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Cheng, X.; Ye, Z.; Li, Y.; Peng, W.; Wu, Y.; Xing, C. Exosome-Mediated Transfer of circ_0000338 Enhances 5-Fluorouracil Resistance in Colorectal Cancer through Regulating MicroRNA 217 (miR-217) and miR-485-3p. Mol. Cell Biol. 2021, 41, e00517-20. [Google Scholar] [CrossRef]
- Zhou, L.; Li, J.; Tang, Y.; Yang, M. Exosomal LncRNA LINC00659 transferred from cancer-associated fibroblasts promotes colorectal cancer cell progression via miR-342-3p/ANXA2 axis. J. Transl. Med. 2021, 19, 8. [Google Scholar] [CrossRef]
- Xu, M.; Chen, X.; Lin, K.; Zeng, K.; Liu, X.; Xu, X.; Pan, B.; Xu, T.; Sun, L.; He, B.; et al. lncRNA SNHG6 regulates EZH2 expression by sponging miR-26a/b and miR-214 in colorectal cancer. J. Hematol. Oncol. 2019, 12, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostovarpour, M.; Khalaj-Kondori, M.; Ghasemi, T. Correlation between expression levels of lncRNA FER1L4 and RB1 in patients with colorectal cancer. Mol. Biol. Rep. 2021, 48, 4581–4589. [Google Scholar] [CrossRef]
- Yu, J.; Li, S.; Xu, Z.; Guo, J.; Li, X.; Wu, Y.; Zheng, J.; Sun, X. CDX2 inhibits epithelial–mesenchymal transition in colorectal cancer by modulation of Snail expression and β-catenin stabilisation via transactivation of PTEN expression. Br. J. Cancer 2020, 124, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Aguirre-Portolés, C.; Feliu, J.; Reglero, G.; Ramírez de Molina, A. ABCA1 overexpression worsens colorectal cancer prognosis by facilitating tumour growth and caveo-lin-1-dependent invasiveness, and these effects can be ameliorated using the BET inhibitor apabetalone. Mol. Oncol. 2018, 12, 1735–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; He, C.; Guo, X.; Fang, Y.; Lai, Q.; Wang, X.; Li, Q. DDX39B contributes to the proliferation of colorectal cancer through direct binding to CDK6/CCND1. Cell Death Discov. 2022, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Fujii, S.; Matsumoto, S.; Nojima, S.; Morii, E.; Kikuchi, A. Arl4c expression in colorectal and lung cancers promotes tumorigenesis and may represent a novel therapeutic target. Oncogene 2014, 34, 4834–4844. [Google Scholar] [CrossRef] [Green Version]
- Ye, L.; Lin, S.-T.; Mi, Y.-S.; Liu, Y.; Ma, Y.; Sun, H.-M.; Peng, Z.-H.; Fan, J.-W. Overexpression of LARP1 predicts poor prognosis of colorectal cancer and is expected to be a potential therapeutic target. Tumor Biol. 2016, 37, 14585–14594. [Google Scholar] [CrossRef] [Green Version]
- Huang, F.K.; Zheng, C.Y.; Huang, L.K.; Lin, C.Q.; Zhou, J.F.; Wang, J.X. Long non-coding RNA MCF2L-AS1 promotes the aggressiveness of colorectal cancer by sponging miR-874-3p and thereby up-regulating CCNE1. J. Gene Med. 2021, 23, e3285. [Google Scholar] [CrossRef]
- Jiang, H.; Cheng, X.; Liang, Y.; Wang, Y.; Li, Y.; Li, Y. Aberrant expression of WNK lysine deficient protein kinase 1 is associated with poor prognosis of colon adenocarcinoma. Ir. J. Med. Sci. 2022, 192, 57–64. [Google Scholar] [CrossRef]
- Su, L.; Luo, Y.; Yang, Z.; Yang, J.; Yao, C.; Cheng, F.; Shan, J.; Chen, J.; Li, F.; Liu, L.; et al. MEF2D Transduces Microenvironment Stimuli to ZEB1 to Promote Epithelial–Mesenchymal Transition and Metastasis in Colorectal Cancer. Cancer Res. 2016, 76, 5054–5067. [Google Scholar] [CrossRef] [Green Version]
- Lv, X.; Zhang, J.; Zhang, J.; Guan, W.; Ren, W.; Liu, Y.; Xu, G. A Negative Feedback Loop Between NAMPT and TGF-β Signaling Pathway in Colorectal Cancer Cells. OncoTargets Ther. 2021, 14, 187–198. [Google Scholar] [CrossRef]
- Ji, H.; Lu, L.; Huang, J.; Liu, Y.; Zhang, B.; Tang, H.; Sun, D.; Zhang, Y.; Shang, H.; Li, Y.; et al. IL1A polymorphisms is a risk factor for colorectal cancer in Chinese Han population: A case control study. BMC Cancer 2019, 19, 181. [Google Scholar] [CrossRef]
- Siravegna, G.; Mussolin, B.; Buscarino, M.; Corti, G.; Cassingena, A.; Crisafulli, G.; Ponzetti, A.; Cremolini, C.; Amatu, A.; Lauricella, C.; et al. Clonal evolution and resistance to EGFR blockade in the blood of colorectal cancer patients. Nat. Med. 2015, 21, 795–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, C.; Huang, W.; Chen, J.; Feng, W.; Zhang, T.; Wang, Y.; Liu, D.; Ji, X.; Xie, M.; Sun, M.; et al. IGF1-mediated HOXA13 overexpression promotes colorectal cancer metastasis through upregulating ACLY and IGF1R. Cell Death Dis. 2021, 12, 564. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, M.H.; Shaik, A.S.; Vaali-Mohammed, M.A.; Al Khayal, K.A.; Traiki, T.B.; Zubaidi, A.M.; Al-Obeed, O.A. Expression of VEGF, EGF and HGF in early- and late-stage colorectal cancer. Mol. Clin. Oncol. 2021, 15, 251. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Lu, Z.; Fu, W.; Lu, K.; Gu, X.; Xu, F.; Dai, J.; Yang, Y.; Jiang, J. Exosome-Derived ADAM17 Promotes Liver Metastasis in Colorectal Cancer. Front. Pharmacol. 2021, 12, 734351. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, Z.; Pan, T.; Hu, H.; He, R.; Yi, M.; Wang, C. RNF186/EPHB2 Axis Is Essential in Regulating TNF Signaling for Colorectal Tumorigenesis in Colorectal Epithelial Cells. J. Immunol. 2022, 209, 1796–1805. [Google Scholar] [CrossRef]
- Zhao, J.P.; Chen, L.L. Circular RNA MAT2B Induces Colorectal Cancer Proliferation via Sponging miR-610, Resulting in an Increased E2F1 Expression. Cancer Manag. Res. 2020, 12, 7107–7116. [Google Scholar] [CrossRef]
- Yang, D.; Hou, T.; Li, L.; Chu, Y.; Zhou, F.; Xu, Y.; Hou, X.; Song, H.; Zhu, K.; Hou, Z.; et al. Smad1 promotes colorectal cancer cell migration through Ajuba transactivation. Oncotarget 2017, 8, 110415–110425. [Google Scholar] [CrossRef] [Green Version]
- Nosho, K.; Shima, K.; Irahara, N.; Kure, S.; Baba, Y.; Kirkner, G.J.; Chen, L.; Gokhale, S.; Hazra, A.; Spiegelman, D.; et al. DNMT3B Expression Might Contribute to CpG Island Methylator Phenotype in Colorectal Cancer. Clin. Cancer Res. 2009, 15, 3663–3671. [Google Scholar] [CrossRef] [Green Version]
- Mao, G.; Zhou, B.; Xu, W.; Jiao, N.; Wu, Z.; Li, J.; Liu, Y. Hsa_circ_0040809 regulates colorectal cancer development by upregulating methyltransferase DNMT1 via targeting miR-515-5p. J. Gene Med. 2021, 23, e3388. [Google Scholar] [CrossRef]
- Fleming, N.I.; Jorissen, R.N.; Mouradov, D.; Christie, M.; Sakthianandeswaren, A.; Palmieri, M.; Sieber, O.M. SMAD2, SMAD3 and SMAD4 mutations in colorectal cancer. Cancer Res. 2013, 73, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Wang, D.; Wang, X.; Sun, S.; Zhang, Y.; Wang, S.; Qu, X. CXCL12/CXCR4 promotes inflammation-driven colorectal cancer progression through activation of RhoA signaling by sponging miR-133a-3p. J. Exp. Clin. Cancer Res. 2019, 38, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Ma, S.-P.; Yang, D.; Liu, Y.; Wang, Y.-P.; Lin, T.; Li, Y.-X.; Yang, S.-H.; Zhang, W.-C.; Wang, X.-L. miR-142-3p Suppresses Cell Growth by Targeting CDK4 in Colorectal Cancer. Cell. Physiol. Biochem. 2018, 51, 1969–1981. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, H.; Chen, Y.; Qiao, G.; Jiang, W.; Ni, P.; Liu, X.; Ma, L. miR-598 inhibits metastasis in colorectal cancer by suppressing JAG1/Notch2 pathway stimulating EMT. Exp. Cell Res. 2017, 352, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Zhang, T.; Deng, S.-C.; Wei, J.-C.; Yang, P.; Wang, Q.; Chen, Z.-P.; Li, W.-L.; Chen, H.-C.; Hu, H.; et al. PD-L1 promotes colorectal cancer stem cell expansion by activating HMGA1-dependent signaling pathways. Cancer Lett. 2019, 450, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Long, X.; Duan, S.; Liu, X.; Chen, J.; Lan, J.; Zhou, J. CSRP2 suppresses colorectal cancer progression via p130Cas/Rac1 axis-meditated ERK, PAK, and HIPPO signaling pathways. Theranostics 2020, 10, 11063–11079. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Cai, S.; Wang, X.; Jiang, Z. Hypomethylation-Associated Up-Regulation of TCF3 Expression and Recurrence in Stage II and III Colorectal Cancer. PLoS ONE 2014, 9, e112005. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Wang, J.; Li, C.; Xuan, M.; Han, S.; Zhang, Y.; Liu, P.; Zhao, Z. miR-17-5p promotes the invasion and migration of colorectal cancer by regulating HSPB2. J. Cancer 2022, 13, 918–931. [Google Scholar] [CrossRef]
- Shen, M.; Xie, S.; Rowicki, M.; Michel, S.; Wei, Y.; Hang, X.; Wan, L.; Lu, X.; Yuan, M.; Jin, J.F.; et al. Therapeutic Targeting of Metadherin Suppresses Colorectal and Lung Cancer Progression and Metastasis. Cancer Res. 2021, 81, 1014–1025. [Google Scholar] [CrossRef]
- Ma, L.; Lin, Y.; Sun, S.-W.; Xu, J.; Yu, T.; Chen, W.-L.; Zhang, L.-H.; Guo, Y.-C.; Wang, Y.-W.; Chen, T.; et al. KIAA1429 is a potential prognostic marker in colorectal cancer by promoting the proliferation via downregulating WEE1 expression in an m6A-independent manner. Oncogene 2021, 41, 692–703. [Google Scholar] [CrossRef]
- Mima, K.; Nishihara, R.; Yang, J.; Dou, R.; Masugi, Y.; Shi, Y.; da Silva, A.; Cao, Y.; Song, M.; Nowak, J.; et al. MicroRNA MIR21 (miR-21) and PTGS2 Expression in Colorectal Cancer and Patient Survival. Clin. Cancer Res. 2016, 22, 3841–3848. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Zhang, Y.; Segar, N.; Huang, C.; Zeng, P.; Tan, X.; Mao, L.; Chen, Z.; Haglund, F.; Larsson, O.; et al. Nuclear IGF1R interacts with NuMA and regulates 53BP1-dependent DNA double-strand break repair in colorectal cancer. Oncol. Rep. 2021, 46, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Lee, C.J.; Choi, J.H.; Kim, J.H.; Kim, J.W.; Kim, J.Y.; Nam, J.S. The JAK2/STAT3/CCND2 Axis promotes colorectal Cancer stem cell persistence and radioresistance. J. Exp. Clin. Cancer Res. 2019, 38, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noda, M.; Okayama, H.; Kofunato, Y.; Chida, S.; Saito, K.; Tada, T.; Ashizawa, M.; Nakajima, T.; Aoto, K.; Kikuchi, T.; et al. Prognostic role of FUT8 expression in relation to p53 status in stage II and III colorectal cancer. PLoS ONE 2018, 13, e0200315. [Google Scholar] [CrossRef] [Green Version]
- Du, W.; Liu, N.; Zhang, Y.; Liu, X.; Yang, Y.; Chen, W.; He, Y. PLOD2 promotes aerobic glycolysis and cell progression in colorectal cancer by upregulating HK2. Biochem. Cell Biol. 2020, 98, 386–395. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-H.; Ro, E.J.; Yoon, J.-S.; Mizutani, T.; Kang, D.-W.; Park, J.-C.; Kim, T.I.; Clevers, H.; Choi, K.-Y. 5-FU promotes stemness of colorectal cancer via p53-mediated WNT/β-catenin pathway activation. Nat. Commun. 2020, 11, 5321. [Google Scholar] [CrossRef]
- Ma, Z.; Lou, S.; Jiang, Z. PHLDA2 regulates EMT and autophagy in colorectal cancer via the PI3K/AKT signaling pathway. Aging 2020, 12, 7985–8000. [Google Scholar] [CrossRef]
- Wang, X.; Lai, Q.; He, J.; Li, Q.; Ding, J.; Lan, Z.; Liu, S. LncRNA SNHG6 promotes proliferation, invasion and migration in colorectal cancer cells by activating TGF-β/Smad signaling pathway via targeting UPF1 and inducing EMT via regulation of ZEB1. Int. J. Med. Sci. 2019, 16, 51–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Xu, X.; Chen, D.; Zhao, F.; Wang, W. Therapeutic potential of targeting the Wnt/β-catenin signaling pathway in colorectal cancer. Biomed. Pharmacother. 2018, 110, 473–481. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Ogata, H.; Goto, S.; Sato, K.; Fujibuchi, W.; Bono, H.; Kanehisa, M. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Kern, F.; Fehlmann, T.; Solomon, J.; Schwed, L.; Grammes, N.; Backes, C.; Van Keuren-Jensen, K.; Craig, D.W.; Meese, E.; Keller, A. miEAA 2.0: Integrating multi-species microRNA enrichment analysis and workflow management systems. Nucleic Acids Res. 2020, 48, W521–W528. [Google Scholar] [CrossRef]
- Dweep, H.; Gretz, N. miRWalk2.0: A comprehensive atlas of microRNA-target interactions. Nat. Methods 2015, 12, 697. [Google Scholar] [CrossRef]
- Xia, R.M.; Liu, T.; Li, W.G.; Xu, X.Q. RNA-binding protein RBM24 represses colorectal tumourigenesis by stabilising PTEN mRNA. Clin. Transl. Med. 2021, 11, e383. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Dang, J.; Song, A.; Cui, X.; Ma, Z.; Zhang, Z. NEAT1 promotes colon cancer progression through sponging miR-495-3p and activating CDK6 in vitro and in vivo. J. Cell. Physiol. 2019, 234, 19582–19591. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Tang, C.; Tang, M.; Zhang, Q.; Gao, Y.; Ge, Z. TRIM47 is up-regulated in colorectal cancer, promoting ubiquitination and degradation of SMAD4. J. Exp. Clin. Cancer Res. 2019, 38, 159. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, A.V.; Bitner, L.; Syed, M.; Woods, M.; Manyam, G.; Kwong, L.N.; Kopetz, S. Targeting RAS Mutant Colorectal Cancer with Dual Inhibition of MEK and CDK4/6. Cancer Res. 2022, 82, 3335–3344. [Google Scholar] [CrossRef]
- Li, F.; Zhou, Y.D.; Liu, J.; Cai, J.D.; Liao, Z.M.; Chen, G.Q. RBP-J promotes cell growth and metastasis through regulating miR-182-5p-mediated Tiam1/Rac1/p38 MAPK axis in colorectal cancer. Cell. Signal. 2021, 87, 110103. [Google Scholar] [CrossRef]
- Chahdi, A.; Raufman, J.-P. The Cdc42/Rac nucleotide exchange factor protein β1Pix (Pak-interacting exchange factor) modulates β-catenin transcriptional activity in colon cancer cells: Evidence for direct interaction of β1PIX with β-catenin. J. Biol. Chem. 2013, 288, 34019–34029. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Leone, G.W.; Wang, H. Cyclin D-CDK4/6 functions in cancer. Adv. Cancer Res. 2020, 148, 147–169. [Google Scholar] [PubMed]
- Huang, L.; Ren, F.; Tang, R.; Feng, Z.; Chen, G. Prognostic value of expression of cyclin E in gastrointestinal cancer: A systematic review and meta-analysis. Technol. Cancer Res. Treat. 2016, 15, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Clemo, N.; Arhel, N.; Barnes, J.; Baker, J.; Moorghen, M.; Packham, G.; Paraskeva, C.; Williams, A. The role of the retinoblastoma protein (Rb) in the nuclear localization of BAG-1: Implications for colorectal tumour cell survival. Biochem. Soc. Trans. 2005, 33, 676–678. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-B.; Lim, H.-J.; Lee, H.-J.; Park, J.H.; Park, S.-G. Evaluation and Clinical Significance of Jagged-1-activated Notch Signaling by APEX1 in Colorectal Cancer. Anticancer Res. 2019, 39, 6097–6105. [Google Scholar] [CrossRef]
- Vaish, V.; Kim, J.; Shim, M. Jagged-2 (JAG2) enhances tumorigenicity and chemoresistance of colorectal cancer cells. Oncotarget 2017, 8, 53262–53275. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Liang, S.; Cheng, Z.; Zhang, X.; Luo, L.; Li, L.; Zhang, W.; Li, S.; Xu, Q.; Zhong, M.; et al. ROS/PI3K/Akt and Wnt/β-catenin signalings activate HIF-1α-induced metabolic reprogramming to impart 5-fluorouracil resistance in colorectal cancer. J. Exp. Clin. Cancer Res. 2022, 41, 15. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, H.; Liu, L.; Song, H.; Zhang, Y.; Wang, J.; Song, J. CircIL4R activates the PI3K/AKT signaling pathway via the miR-761/TRIM29/PHLPP1 axis and promotes proliferation and metastasis in colorectal cancer. Mol. Cancer 2021, 20, 167. [Google Scholar] [CrossRef]
- Stefani, C.; Miricescu, D.; Stanescu-Spinu, I.I.; Nica, R.I.; Greabu, M.; Totan, A.R.; Jinga, M. Growth factors, PI3K/AKT/mTOR and MAPK signaling pathways in colorectal cancer pathogenesis: Where are we now? Int. J. Mol. Sci. 2021, 22, 10260. [Google Scholar] [CrossRef]
- Tovar-Camargo, O.A.; Toden, S.; Goel, A. Exosomal microRNA Biomarkers: Emerging Frontiers in Colorectal and Other Human Cancers. Expert Rev. Mol. Diagn. 2016, 16, 553–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toden, S.; Goel, A. Non-coding RNAs as liquid biopsy biomarkers in cancer. Br. J. Cancer 2022, 126, 351–360. [Google Scholar] [CrossRef]
- Bracht, J.W.P.; Mayo-De-Las-Casas, C.; Berenguer, J.; Karachaliou, N.; Rosell, R. The Present and Future of Liquid Biopsies in Non-Small Cell Lung Cancer: Combining Four Biosources for Diagnosis, Prognosis, Prediction, and Disease Monitoring. Curr. Oncol. Rep. 2018, 20, 70. [Google Scholar] [CrossRef]
- Minciacchi, V.; Zijlstra, A.; Rubin, M.A.; Di Vizio, D. Extracellular vesicles for liquid biopsy in prostate cancer: Where are we and where are we headed? Prostate Cancer Prostatic Dis. 2017, 20, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uratani, R.; Toiyama, Y.; Kitajima, T.; Kawamura, M.; Hiro, J.; Kobayashi, M.; Tanaka, K.; Inoue, Y.; Mohri, Y.; Mori, T.; et al. Diagnostic Potential of Cell-Free and Exosomal MicroRNAs in the Identification of Patients with High-Risk Colorectal Adenomas. PLoS ONE 2016, 11, e0160722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azuaje, F.; Devaux, Y.; Wagner, D. Challenges and Standards in Reporting Diagnostic and Prognostic Biomarker Studies. Clin. Transl. Sci. 2009, 2, 156–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]





| Non-Coding RNA(s) | Non-Coding RNA Class | Number of CRC Patients | Source | Biomarker Potential | Status | Clinical Evidence | References |
|---|---|---|---|---|---|---|---|
| circLPAR1 | circRNA | 112 | Plasma | Diagnostic | Downregulated | Yes | [43] |
| Let-7a, miR-1229, miR-1246, miR-150, miR-21, miR-223, and miR-23a | miRNA | 99 | Serum | Diagnostic | Upregulated | Yes | [21] |
| CCAT2 | LncRNA | 100 | Serum | Diagnostic | Upregulated | Yes | [28] |
| RPPH1 | LncRNA | 52 | Plasma | Diagnostic | Upregulated | Yes | [38] |
| HOTTIP | LncRNA | 100 | Serum | Diagnostic | Downregulated | Yes | [39] |
| miR-92b | miRNA | 40 | Plasma | Diagnostic | Downregulated | Yes | [23] |
| miR-6803-5p | miRNA | 168 | Serum | Diagnostic | Upregulated | Yes | [34] |
| miR-96-5p and miR-149 | miRNA | 102 | Plasma | Diagnostic | Downregulated | Yes | [24] |
| miR-125a, miR-320c | miRNA | 50 | Plasma | Diagnostic | Upregulated | Yes | [35] |
| MiR-150-5p | miRNA | 133 | Serum | Diagnostic | Downregulated | Yes | [34] |
| MiR-4461 | miRNA | 15 | BMSC | Diagnostic | Downregulated | Yes | [31] |
| MiR-10A | miRNA | 40 | Primary CRC cells | Diagnostic | Downregulated | Yes | [25] |
| MiR-874 | miRNA | 125 | Serum | Diagnostic | Downregulated | Yes | [30] |
| MiR-19a-3p, miR-203-3p, miR-221-3p, let-7f-5p | miRNA | 36 | Serum | Diagnostic | Upregulated | Yes | [22] |
| MiR-377-3p, miR-381-3p | miRNA | 175 | Serum | Diagnostic | Downregulated | Yes | [26] |
| LNCV6_116109, LNCV6_98390, LNCV6_38772, LNCV_108266, LNCV6_84003, and LNCV6_98602 | LncRNA | 50 | Plasma | Diagnostic | Upregulated | Yes | [29] |
| CRNDE-h | LncRNA | 148 | Serum | Diagnostic | Upregulated | Yes | [41] |
| GAS5 | LncRNA | 158 | Serum | Diagnostic | Downregulated | Yes | [42] |
| MiR-17-5p | miRNA | N/A | Serum | Diagnostic | Upregulated | No | [33] |
| ADAMTS9-AS1 | LncRNA | 886 | Serum | Diagnostic | Downregulated | Yes | [40] |
| miR-4323, miR-4284, miR-1268a, miR-1290, miR-6766-3p, miR-21-5p, and miR-1246 | miRNA | 26 | Serum | Diagnostic | Upregulated | Yes | [27] |
| miR-200c-3p | miRNA | N/A | Serum | Diagnostic | Downregulated | No | [36] |
| miR-424-5p | miRNA | N/A | Serum | Diagnostic | Upregulated | No | [37] |
| Biological Process | ||
| GO Term | Gene Count | FDR |
| positive regulation of transcription from RNA polymerase II promoter | 14 | 1.30 × 10−4 |
| negative regulation of transcription from RNA polymerase II promoter | 10 | 7.80 × 10−3 |
| positive regulation of cell proliferation | 9 | 1.80 × 10−3 |
| regulation of transcription from RNA polymerase II promoter | 9 | 2.00 × 10−1 |
| positive regulation of gene expression | 8 | 6.30 × 10−3 |
| positive regulation of transcription, DNA-templated | 8 | 2.80 × 10−2 |
| negative regulation of cell proliferation | 7 | 1.80 × 10−2 |
| negative regulation of apoptotic process | 7 | 2.80 × 10−2 |
| regulation of transcription, DNA-templated | 7 | 1.80 × 10−1 |
| signal transduction | 7 | 3.30 × 10−1 |
| G1/S transition of mitotic cell cycle | 6 | 1.30 × 10−4 |
| response to hypoxia | 6 | 3.70 × 10−3 |
| positive regulation of cell migration | 6 | 1.30 × 10−2 |
| response to drug | 6 | 1.80 × 10−2 |
| cell division | 6 | 3.70 × 10−2 |
| positive regulation of fibroblast proliferation | 5 | 1.30 × 10−3 |
| positive regulation of smooth muscle cell proliferation | 5 | 1.80 × 10−3 |
| Aging | 5 | 2.90 × 10−2 |
| positive regulation of protein phosphorylation | 5 | 3.50 × 10−2 |
| regulation of gene expression | 5 | 5.10 × 10−2 |
| regulation of cell cycle | 5 | 9.70 × 10−2 |
| nervous system development | 5 | 1.50 × 10−1 |
| protein phosphorylation | 5 | 2.00 × 10−1 |
| negative regulation of transcription, DNA-templated | 5 | 3.00 × 10−1 |
| cell differentiation | 5 | 3.80 × 10−1 |
| Molecular Function | ||
| GO Term | Gene Count | FDR |
| protein binding | 36 | 3.40 × 10−3 |
| protein kinase binding | 9 | 1.00 × 10−3 |
| RNA polymerase II core promoter proximal region sequence-specific DNA binding | 8 | 1.10 × 10−1 |
| identical protein binding | 8 | 3.00 × 10−1 |
| chromatin binding | 7 | 1.30 × 10−2 |
| DNA binding | 7 | 3.00 × 10−1 |
| RNA polymerase II sequence-specific DNA binding transcription factor binding | 6 | 3.20 × 10−3 |
| enzyme binding | 6 | 3.20 × 10−2 |
| transcription factor activity, sequence-specific DNA binding | 6 | 9.50 × 10−2 |
| kinase activity | 5 | 3.20 × 10−2 |
| transcriptional activator activity, RNA polymerase II transcription regulatory region sequence-specific binding | 5 | 1.60 × 10−1 |
| Cellular Component | ||
| GO Term | Gene Count | FDR |
| Nucleus | 24 | 1.30 × 10−3 |
| Cytoplasm | 23 | 1.30 × 10−3 |
| Cytosol | 19 | 4.50 × 10−2 |
| Nucleoplasm | 18 | 3.80 × 10−3 |
| Chromatin | 11 | 1.30 × 10−3 |
| Membrane | 10 | 1.60 × 10−1 |
| intracellular membrane-bounded organelle | 7 | 5.70 × 10−2 |
| endoplasmic reticulum membrane | 7 | 6.60 × 10−2 |
| transcription factor complex | 6 | 1.30 × 10−3 |
| macromolecular complex | 6 | 5.70 × 10−2 |
| apical plasma membrane | 5 | 5.00 × 10−2 |
| focal adhesion | 5 | 5.70 × 10−2 |
| Golgi membrane | 5 | 1.50 × 10−1 |
| KEGG Pathway | Gene Count | FDR |
|---|---|---|
| Pathways in cancer | 16 | 1.00 × 10−8 |
| MicroRNAs in cancer | 11 | 2.90 × 10−6 |
| Human papillomavirus infection | 10 | 2.10 × 10−5 |
| PI3K-Akt signaling pathway | 10 | 3.40 × 10−5 |
| Melanoma | 9 | 1.00 × 10−8 |
| Breast cancer | 9 | 1.50 × 10−6 |
| Hepatocellular carcinoma | 9 | 2.90 × 10−6 |
| Human T-cell leukemia virus 1 infection | 9 | 1.40 × 10−5 |
| Glioma | 8 | 3.70 × 10−7 |
| Cell cycle | 8 | 5.20 × 10−6 |
| Cellular senescence | 8 | 1.40 × 10−5 |
| Focal adhesion | 8 | 5.20 × 10−5 |
| Epstein-Barr virus infection | 8 | 5.20 × 10−5 |
| Viral carcinogenesis | 8 | 5.30 × 10−5 |
| Human cytomegalovirus infection | 8 | 9.60 × 10−5 |
| Pancreatic cancer | 7 | 5.20 × 10−6 |
| Small cell lung cancer | 7 | 1.40 × 10−5 |
| Prostate cancer | 7 | 1.40 × 10−5 |
| Endocrine resistance | 7 | 1.40 × 10−5 |
| Proteoglycans in cancer | 7 | 5.60 × 10−4 |
| MAPK signaling pathway | 7 | 2.70 × 10−3 |
| Non-small cell lung cancer | 6 | 5.00 × 10−5 |
| p53 signaling pathway | 6 | 5.10 × 10−5 |
| FoxO signaling pathway | 6 | 6.40 × 10−4 |
| Measles | 6 | 7.70 × 10−4 |
| Gastric cancer | 6 | 9.60 × 10−4 |
| Cushing syndrome | 6 | 1.10 × 10−3 |
| Hepatitis B | 6 | 1.30 × 10−3 |
| Transcriptional misregulation in cancer | 6 | 2.70 × 10−3 |
| Kaposi sarcoma-associated herpesvirus infection | 6 | 2.70 × 10−3 |
| Rap1 signaling pathway | 6 | 3.70 × 10−3 |
| Ras signaling pathway | 6 | 6.00 × 10−3 |
| Adherens junction | 5 | 7.00 × 10−4 |
| Chronic myeloid leukemia | 5 | 8.50 × 10−4 |
| EGFR tyrosine kinase inhibitor resistance | 5 | 9.50 × 10−4 |
| Signaling pathways regulating pluripotency of stem cells | 5 | 6.50 × 10−3 |
| Hepatitis C | 5 | 8.90 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mezher, M.; Abdallah, S.; Ashekyan, O.; Shoukari, A.A.; Choubassy, H.; Kurdi, A.; Temraz, S.; Nasr, R. Insights on the Biomarker Potential of Exosomal Non-Coding RNAs in Colorectal Cancer: An In Silico Characterization of Related Exosomal lncRNA/circRNA–miRNA–Target Axis. Cells 2023, 12, 1081. https://doi.org/10.3390/cells12071081
Mezher M, Abdallah S, Ashekyan O, Shoukari AA, Choubassy H, Kurdi A, Temraz S, Nasr R. Insights on the Biomarker Potential of Exosomal Non-Coding RNAs in Colorectal Cancer: An In Silico Characterization of Related Exosomal lncRNA/circRNA–miRNA–Target Axis. Cells. 2023; 12(7):1081. https://doi.org/10.3390/cells12071081
Chicago/Turabian StyleMezher, Maria, Samira Abdallah, Ohanes Ashekyan, Ayman Al Shoukari, Hayat Choubassy, Abdallah Kurdi, Sally Temraz, and Rihab Nasr. 2023. "Insights on the Biomarker Potential of Exosomal Non-Coding RNAs in Colorectal Cancer: An In Silico Characterization of Related Exosomal lncRNA/circRNA–miRNA–Target Axis" Cells 12, no. 7: 1081. https://doi.org/10.3390/cells12071081
APA StyleMezher, M., Abdallah, S., Ashekyan, O., Shoukari, A. A., Choubassy, H., Kurdi, A., Temraz, S., & Nasr, R. (2023). Insights on the Biomarker Potential of Exosomal Non-Coding RNAs in Colorectal Cancer: An In Silico Characterization of Related Exosomal lncRNA/circRNA–miRNA–Target Axis. Cells, 12(7), 1081. https://doi.org/10.3390/cells12071081