

Characterization of Non-Cholesterol Sterols in Microglia Cell Membranes Using Targeted Mass Spectrometry

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Human Samples
2.3. Sample Preparation
2.4. LC-MS/MS Analysis
2.5. Method Validation
2.6. Stability Assessment
2.7. Method Comparison
2.8. Cell Membrane Isolation
2.9. Western Blot
2.10. Lipid Extraction for LC-MS/MS Analysis
2.11. LC-MS/MS Analysis of PUFAs in Cell Membrane Fractions
2.12. Data Analysis
3. Results
3.1. Method Development and Validation
3.1.1. APCI-LC-MS/MS Sterol Method
3.1.2. Method Validation
3.1.3. Stability Assessment
3.1.4. Method Comparison
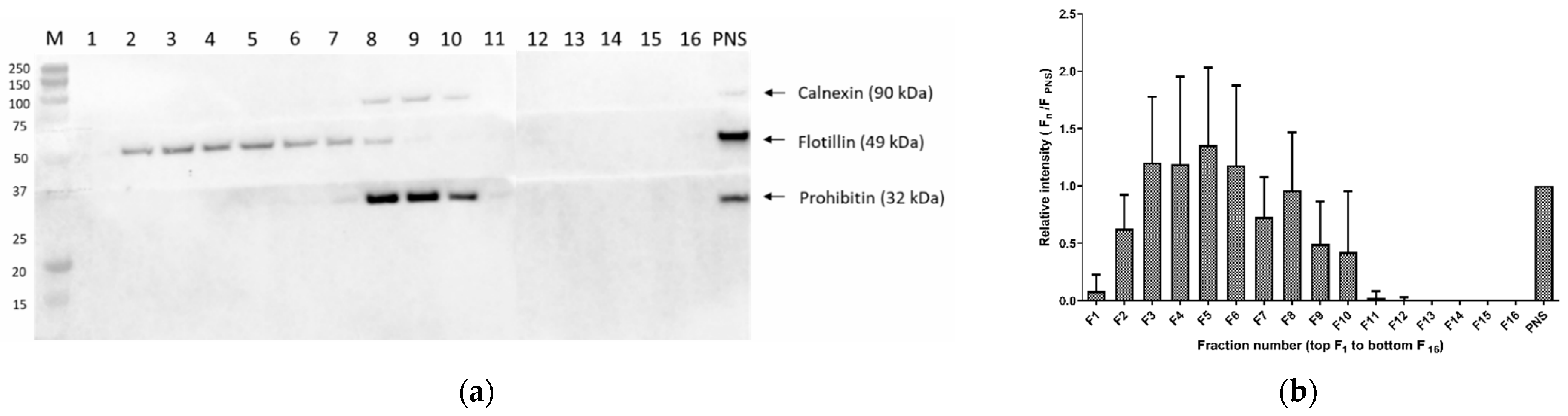
3.1.5. Optimization of Cell Membrane Isolation for Sterol Analysis
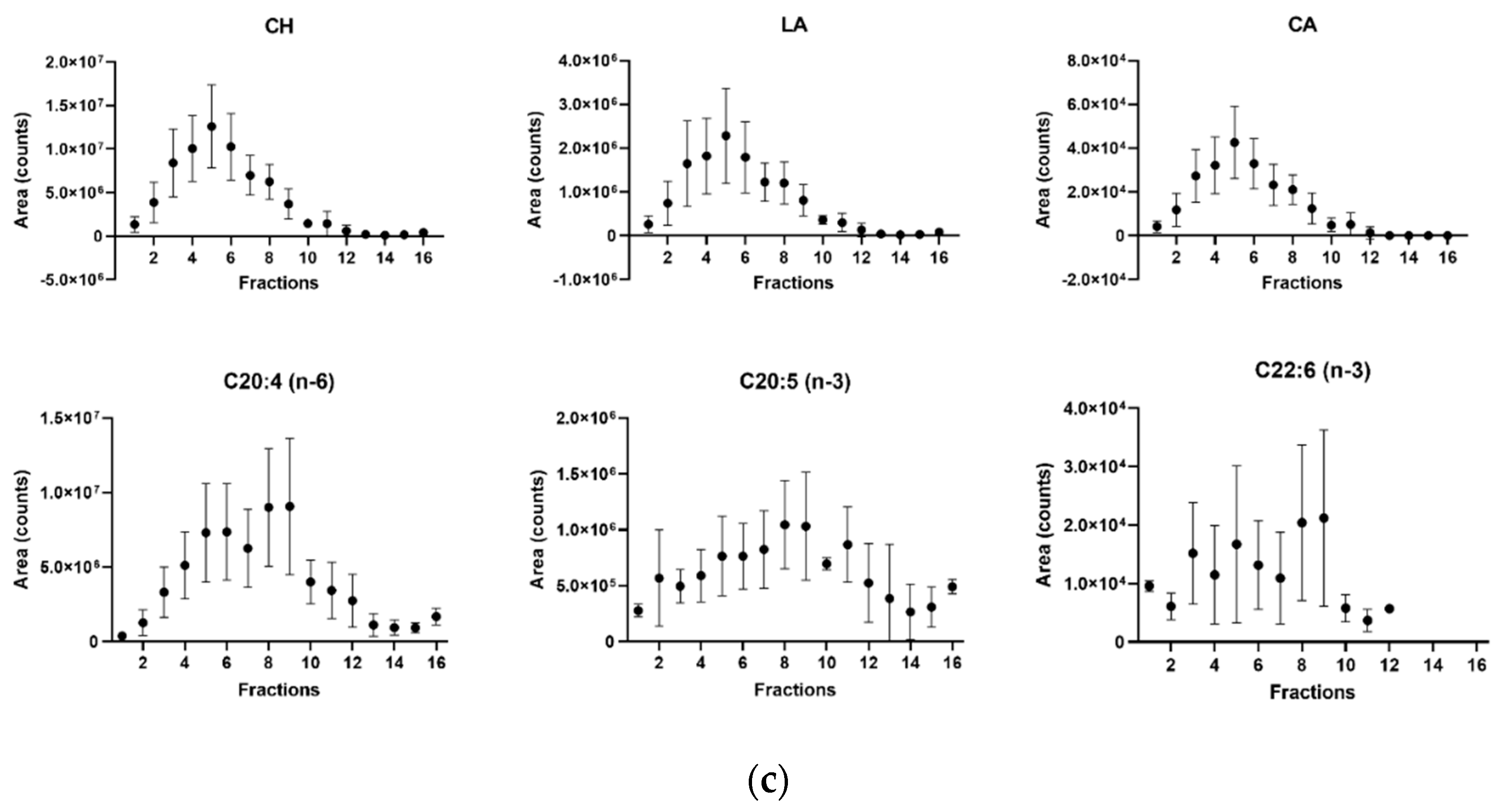
3.2. Sterol Composition of Microglia Cell Membranes
3.3. Sterol Quantification in Cell Membrane Fractions and Its Relation to Plasma
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, D.A.; London, E. Functions of lipid rafts in biological membranes. Annu. Rev. Cell Dev. Biol. 1998, 14, 111–136. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Gerl, M.J. Revitalizing membrane rafts: New tools and insights. Nat. Rev. Mol. Cell Biol. 2010, 11, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Kuksis, A. Plasma non-cholesterol sterols. J. Chromatogr. A 2001, 935, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Weingärtner, O.; Lütjohann, D.; Ji, S.; Weisshoff, N.; List, F.; Sudhop, T.; von Bergmann, K.; Gertz, K.; König, J.; Schäfers, H.-J.; et al. Vascular effects of diet supplementation with plant sterols. J. Am. Coll. Cardiol. 2008, 51, 1553–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, M.; Horn, K.; Pott, J.; Gross, A.; Kleber, M.E.; Delgado, G.E.; Mishra, P.P.; Kirsten, H.; Gieger, C.; Müller-Nurasyid, M.; et al. Genome-wide meta-analysis of phytosterols reveals five novel loci and a detrimental effect on coronary atherosclerosis. Nat. Commun. 2022, 13, 143. [Google Scholar] [CrossRef] [PubMed]
- Valerio, M.S.; Minderman, H.; Mace, T.; Awad, A.B. β-Sitosterol modulates TLR4 receptor expression and intracellular MyD88-dependent pathway activation in J774A.1 murine macrophages. Cell. Immunol. 2013, 285, 76–83. [Google Scholar] [CrossRef]
- Dierckx, T.; Bogie, J.F.J.; Hendriks, J.J.A. The Impact of Phytosterols on the Healthy and Diseased Brain. Curr. Med. Chem. 2019, 26, 6750–6765. [Google Scholar] [CrossRef]
- Björkhem, I.; Meaney, S. Brain cholesterol: Long secret life behind a barrier. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 806–815. [Google Scholar] [CrossRef] [Green Version]
- Vanmierlo, T.; Weingärtner, O.; van der Pol, S.; Husche, C.; Kerksiek, A.; Friedrichs, S.; Sijbrands, E.; Steinbusch, H.; Grimm, M.; Hartmann, T.; et al. Dietary intake of plant sterols stably increases plant sterol levels in the murine brain. J. Lipid Res. 2012, 53, 726–735. [Google Scholar] [CrossRef] [Green Version]
- Smiljanic, K.; Vanmierlo, T.; Mladenovic Djordjevic, A.; Perovic, M.; Ivkovic, S.; Lütjohann, D.; Kanazir, S. Cholesterol metabolism changes under long-term dietary restrictions while the cholesterol homeostasis remains unaffected in the cortex and hippocampus of aging rats. Age 2014, 36, 9654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, P.J.; Lütjohann, D.; Abildayeva, K.; Vanmierlo, T.; Plösch, T.; Plat, J.; von Bergmann, K.; Groen, A.K.; Ramaekers, F.C.S.; Kuipers, F.; et al. Dietary plant sterols accumulate in the brain. Biochim. Biophys. Acta 2006, 1761, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Reinicke, M.; Leyh, J.; Zimmermann, S.; Chey, S.; Brkovic, I.B.; Wassermann, C.; Landmann, J.; Lütjohann, D.; Isermann, B.; Bechmann, I.; et al. Plant Sterol-Poor Diet Is Associated with Pro-Inflammatory Lipid Mediators in the Murine Brain. Int. J. Mol. Sci. 2021, 22, 13207. [Google Scholar] [CrossRef]
- Shi, C.; Wu, F.; Zhu, X.C.; Xu, J. Incorporation of beta-sitosterol into the membrane increases resistance to oxidative stress and lipid peroxidation via estrogen receptor-mediated PI3K/GSK3beta signaling. Biochim. Biophys. Acta 2013, 1830, 2538–2544. [Google Scholar] [CrossRef]
- Vanmierlo, T.; Bogie, J.F.J.; Mailleux, J.; Vanmol, J.; Lütjohann, D.; Mulder, M.; Hendriks, J.J.A. Plant sterols: Friend or foe in CNS disorders? Prog. Lipid Res. 2015, 58, 26–39. [Google Scholar] [CrossRef]
- Wang, J.; Wu, F.; Shi, C. Substitution of membrane cholesterol with β-sitosterol promotes nonamyloidogenic cleavage of endogenous amyloid precursor protein. Neuroscience 2013, 247, 227–233. [Google Scholar] [CrossRef]
- Valerio, M.; Liu, H.-B.; Heffner, R.; Zivadinov, R.; Ramanathan, M.; Weinstock-Guttman, B.; Awad, A.B. Phytosterols ameliorate clinical manifestations and inflammation in experimental autoimmune encephalomyelitis. Inflamm. Res. 2011, 60, 457–465. [Google Scholar] [CrossRef]
- Li, Q.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2018, 18, 225–242. [Google Scholar] [CrossRef]
- Loving, B.A.; Bruce, K.D. Lipid and Lipoprotein Metabolism in Microglia. Front. Physiol. 2020, 11, 393. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Dahlman-Wright, K. Liver X receptor in cholesterol metabolism. J. Endocrinol. 2010, 204, 233–240. [Google Scholar] [CrossRef] [Green Version]
- Lembcke, J.; Ceglarek, U.; Fiedler, G.M.; Baumann, S.; Leichtle, A.; Thiery, J. Rapid quantification of free and esterified phytosterols in human serum using APPI-LC-MS/MS. J. Lipid Res. 2005, 46, 21–26. [Google Scholar] [CrossRef] [Green Version]
- ISO 5725-6; Accuracy (Trueness and Precision) of Measurement Methods and Results—Part 6: Use In Practice of Accuracy Values. International Organization for Standardization: Brussels, Belgium, 1994.
- Silbernagel, G.; Steiner, L.K.; Hollstein, T.; Fauler, G.; Scharnagl, H.; Stojakovic, T.; Schumann, F.; Bölükbasi, B.; März, W.; Steinhagen-Thiessen, E.; et al. The interrelations between PCSK9 metabolism and cholesterol synthesis and absorption. J. Lipid Res. 2019, 60, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, C.; Pan, M.; Han, X. A Detergent-Free Method for Preparation of Lipid Rafts for the Shotgun Lipidomics Study. Methods Mol. Biol. 2021, 2187, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.L.; Pike, L.J. A simplified method for the preparation of detergent-free lipid rafts. J. Lipid Res. 2005, 46, 1061–1067. [Google Scholar] [CrossRef] [Green Version]
- Kortz, L.; Dorow, J.; Becker, S.; Thiery, J.; Ceglarek, U. Fast liquid chromatography-quadrupole linear ion trap-mass spectrometry analysis of polyunsaturated fatty acids and eicosanoids in human plasma. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 927, 209–213. [Google Scholar] [CrossRef]
- Reinicke, M.; Dorow, J.; Bischof, K.; Leyh, J.; Bechmann, I.; Ceglarek, U. Tissue pretreatment for LC-MS/MS analysis of PUFA and eicosanoid distribution in mouse brain and liver. Anal. Bioanal. Chem. 2020, 412, 2211–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.; Röhnike, S.; Empting, S.; Haas, D.; Mohnike, K.; Beblo, S.; Mütze, U.; Husain, R.A.; Thiery, J.; Ceglarek, U. LC–MS/MS-based quantification of cholesterol and related metabolites in dried blood for the screening of inborn errors of sterol metabolism. Anal. Bioanal. Chem. 2015, 407, 5227–5233. [Google Scholar] [CrossRef]
- Luo, J.; Yang, H.; Song, B.-L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef]
- Baila-Rueda, L.; Cenarro, A.; Cofán, M.; Orera, I.; Barcelo-Batllori, S.; Pocoví, M.; Ros, E.; Civeira, F.; Nerín, C.; Domeño, C. Simultaneous determination of oxysterols, phytosterols and cholesterol precursors by high performance liquid chromatography tandem mass spectrometry in human serum. Anal. Methods 2013, 5, 2249–2257. [Google Scholar] [CrossRef]
- Kim, D.; Park, J.B.; Choi, W.K.; Lee, S.J.; Lim, I.; Bae, S.K. Simultaneous determination of β-sitosterol, campesterol, and stigmasterol in rat plasma by using LC–APCI-MS/MS: Application in a pharmacokinetic study of a titrated extract of the unsaponifiable fraction of Zea mays L. J. Sep. Sci. 2016, 39, 4060–4070. [Google Scholar] [CrossRef] [PubMed]
- Honda, A.; Yamashita, K.; Miyazaki, H.; Shirai, M.; Ikegami, T.; Xu, G.; Numazawa, M.; Hara, T.; Matsuzaki, Y. Highly sensitive analysis of sterol profiles in human serum by LC-ESI-MS/MS. J. Lipid Res. 2008, 49, 2063–2073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, J.G.; Smith, D.D.; Stiles, A.R.; Russell, D.W. A comprehensive method for extraction and quantitative analysis of sterols and secosteroids from human plasma. J. Lipid Res. 2012, 53, 1399–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sezgin, E.; Levental, I.; Mayor, S.; Eggeling, C. The mystery of membrane organization: Composition, regulation and roles of lipid rafts. Nat. Rev. Mol. Cell Biol. 2017, 18, 361–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catani, M.V.; Gasperi, V.; Bisogno, T.; Maccarrone, M. Essential Dietary Bioactive Lipids in Neuroinflammatory Diseases. Antioxid. Redox Signal. 2018, 29, 37–60. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Jia, L.; Gao, P.; Li, Q.; Lu, X.; Li, J.; Xu, G. Study on the effect of eicosapentaenoic acid on phospholipids composition in membrane microdomains of tight junctions of epithelial cells by liquid chromatography/electrospray mass spectrometry. J. Pharm. Biomed. Anal. 2008, 47, 343–350. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, H.; Pu, H.; Wang, G.; Li, W.; Leak, R.K.; Chen, J.; Liou, A.K.; Hu, X. n-3 PUFA supplementation benefits microglial responses to myelin pathology. Sci. Rep. 2014, 4, 7458. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Sterol | LOD (mg/L) | LLOQ (mg/L) | Linear Range (mg/L) | R2 | Calibration Range (mg/L) | R2 |
|---|---|---|---|---|---|---|
| Brassicasterol | 0.05 | 0.1 | 0.1–10 | 0.9984 | 0.1–10 | 0.9984 |
| Campesterol | 0.005 | 0.05 | 0.05–10 | 0.9986 | 0.1–10 | 0.9986 |
| Stigmasterol | 0.05 | 0.1 | 0.1–10 | 0.9988 | 0.1–10 | 0.9988 |
| Sitosterol | 0.005 | 0.05 | 0.05–10 | 0.9990 | 0.1–10 | 0.9990 |
| Lanosterol | 0.005 | 0.05 | 0.05–10 | 0.9984 | 0.1–10 | 0.9984 |
| Desmosterol | 0.01 | 0.05 | 0.05–10 | 0.9988 | 0.1–10 | 0.9986 |
| 7-Dehydrocholesterol | 0.05 | 0.05 | 0.05–10 | 0.9982 | 0.1–10 | 0.9980 |
| Cholesterol | 0.0125 | 2.5 | 2.5–1000 | 0.9992 | 10–1,000 | 0.9992 |
| Sterol Compound | Concentration (mg/L) | Within-Day CV (%) 3 | Between-Day 4 CV (%) | Recovery (%) | |||
|---|---|---|---|---|---|---|---|
| Native | Spiked | Native | Spiked | Native | Spiked | ||
| Brassicasterol | NA | 1.16 ± 0.061 | NA | 5.4 | NA | 5.2 | NA |
| Campesterol | 0.723 ± 0.031 1 | 5.48 ± 0.292 | 4.0 | 5.7 | 4.2 | 5.3 | 97 |
| Stigmasterol | 0.147 ± 0.014 | 0.991 ± 0.053 | 8.9 | 5.0 | 9.6 | 5.4 | 87 |
| Sitosterol | 0.680 ± 0.021 | 5.82 ± 0.360 | 3.1 | 5.8 | 3.1 | 6.2 | 94 |
| Lanosterol | 0.130 ± 0.004 | 1.226 ± 0.053 | 3.2 | 4.1 | 3.3 | 4.3 | 114 |
| Desmosterol | 0.167 ± 0.011 | 0.960 ± 0.043 | 5.7 | 4.7 | 6.6 | 4.5 | 85 |
| 7-Dehydrocholesterol | 0.470 ± 0.034 | 5.64 ± 0.306 | 3.8 | 4.7 | 7.2 | 5.4 | 99 |
| Cholesterol | 509 ± 12.3 | NA | 2.6 | NA | 2.5 | 3.5 | NA |
| 4-cholestenone | NA | 1.33 ± 0.073 2 | NA | 4.2 | NA | 5.5 | NA |
| Sterol Compound | ||||||
|---|---|---|---|---|---|---|
| mg/L | Cholesterol | Lanosterol | Desmosterol | Sitosterol | Campesterol | Stigmasterol |
| LF2 * | 17.1 ± 11.5 (30.2) | 3.7 ± 2.6 (6.9) | 7.3 ± 4.3 (11.2) | 0.070 ± 0.032 (0.085) | 0.041 ± 0.017 (0.044) | 0.057 ± 0.033 (0.080) |
| LF3 | 38.4 ± 20.7 (54.1) | 7.8 ± 4.8 (12.6) | 15.7 ± 7.6 (18.5) | 0.120 ± 0.057 (0.149) | 0.070 ± 0.031 (0.080) | 0.100 ± 0.031 (0.086) |
| LF4 | 44.1 ± 18.4 (42.9) | 8.6 ± 4.8 (11.6) | 18.0 ± 6.4 (13.4) | 0.135 ± 0.050 (0.111) | 0.079 ± 0.030 (0.063) | 0.111 ± 0.024 (0.066) |
| LF5 | 55.4 ± 20.8 (49.2) | 11.0 ± 6.0 (15.4) | 22.9 ± 7.3 (16.0) | 0.175 ± 0.060 (0.143) | 0.098 ± 0.031 (0.075) | 0.137 ± 0.023 (0.058) |
| LF6 | 45.0 ± 17.7 (45.5) | 9.1 ± 5.2 (13.2) | 19.1 ± 6.3 (14.8) | 0.143 ± 0.050 (0.127) | 0.084 ± 0.028 (0.071) | 0.133 ± 0.022 (0.052) |
| LF7 | 31.0 ± 9.6 (25.4) | 5.9 ± 1.9 (4.9) | 13.2 ± 4.3 (10.7) | 0.105 ± 0.031 (0.077) | 0.062 ± 0.014 (0.037) | 0.098 ± 0.037 (0.096) |
| Plasma | 471.7 ± 35.0 (62) | 0.137 ± 0.006 (0.010) | 0.177 ± 0.011 (0.022) | 0.632 ± 0.048 (0.092) | 0.642 ± 0.015 (0.030) | 0.164 ± 0.0032 (0.006) |
| (FR2-7/CH) × 1000 | NA | 200 ± 45 (130) | 440 ± 72 (340) | 3.8 ± 2.3 (13) | 2.3 ± 1.8 (10.1) | 2.9 ± 1.2 (6.5) |
| (Sterol/CH) × 1000 plasma | NA | 0.29 ± 0.016 (0.020) | 0.38 ± 0.021 (0.040) | 1.3 ± 0.030 (0.060) | 1.4 ± 0.08 (0.15) | 0.35 ± 0.026 (0.050) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Begcevic Brkovic, I.; Reinicke, M.; Chey, S.; Bechmann, I.; Ceglarek, U. Characterization of Non-Cholesterol Sterols in Microglia Cell Membranes Using Targeted Mass Spectrometry. Cells 2023, 12, 974. https://doi.org/10.3390/cells12070974
Begcevic Brkovic I, Reinicke M, Chey S, Bechmann I, Ceglarek U. Characterization of Non-Cholesterol Sterols in Microglia Cell Membranes Using Targeted Mass Spectrometry. Cells. 2023; 12(7):974. https://doi.org/10.3390/cells12070974
Chicago/Turabian StyleBegcevic Brkovic, Ilijana, Madlen Reinicke, Soroth Chey, Ingo Bechmann, and Uta Ceglarek. 2023. "Characterization of Non-Cholesterol Sterols in Microglia Cell Membranes Using Targeted Mass Spectrometry" Cells 12, no. 7: 974. https://doi.org/10.3390/cells12070974
APA StyleBegcevic Brkovic, I., Reinicke, M., Chey, S., Bechmann, I., & Ceglarek, U. (2023). Characterization of Non-Cholesterol Sterols in Microglia Cell Membranes Using Targeted Mass Spectrometry. Cells, 12(7), 974. https://doi.org/10.3390/cells12070974