
Mast Cells as a Target—A Comprehensive Review of Recent Therapeutic Approaches


 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
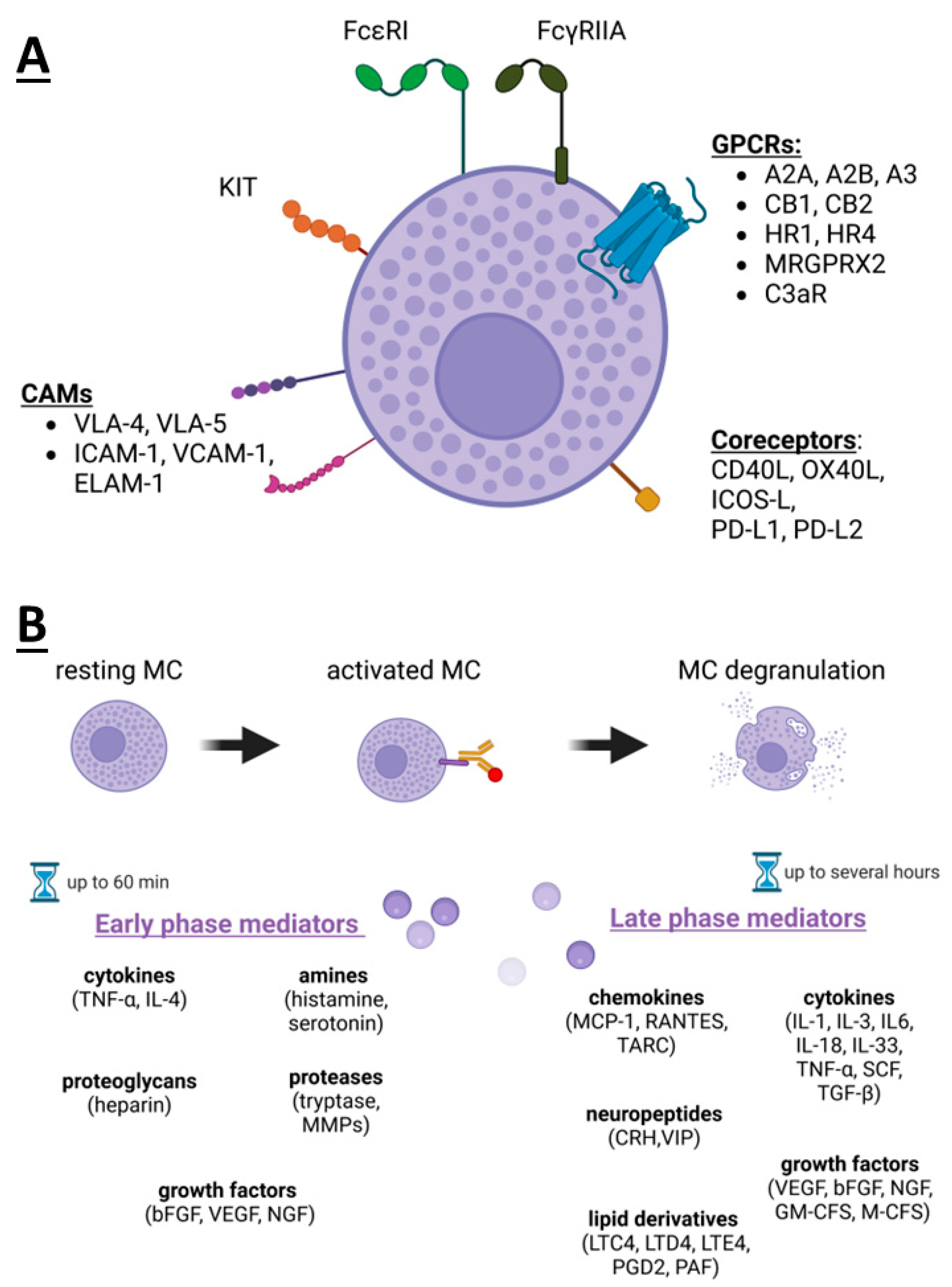
2. Mast Cells’ Receptors and Mediators
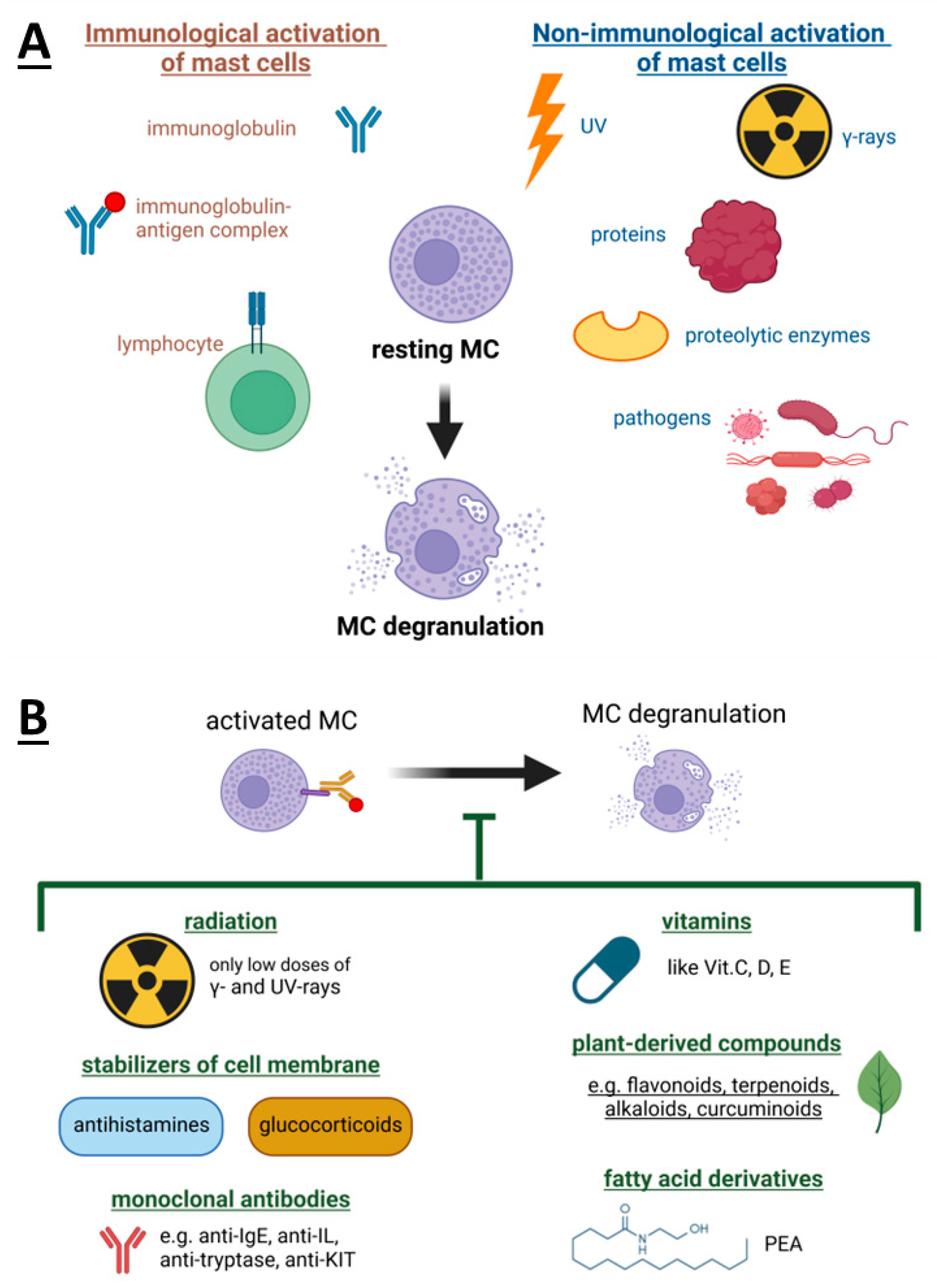
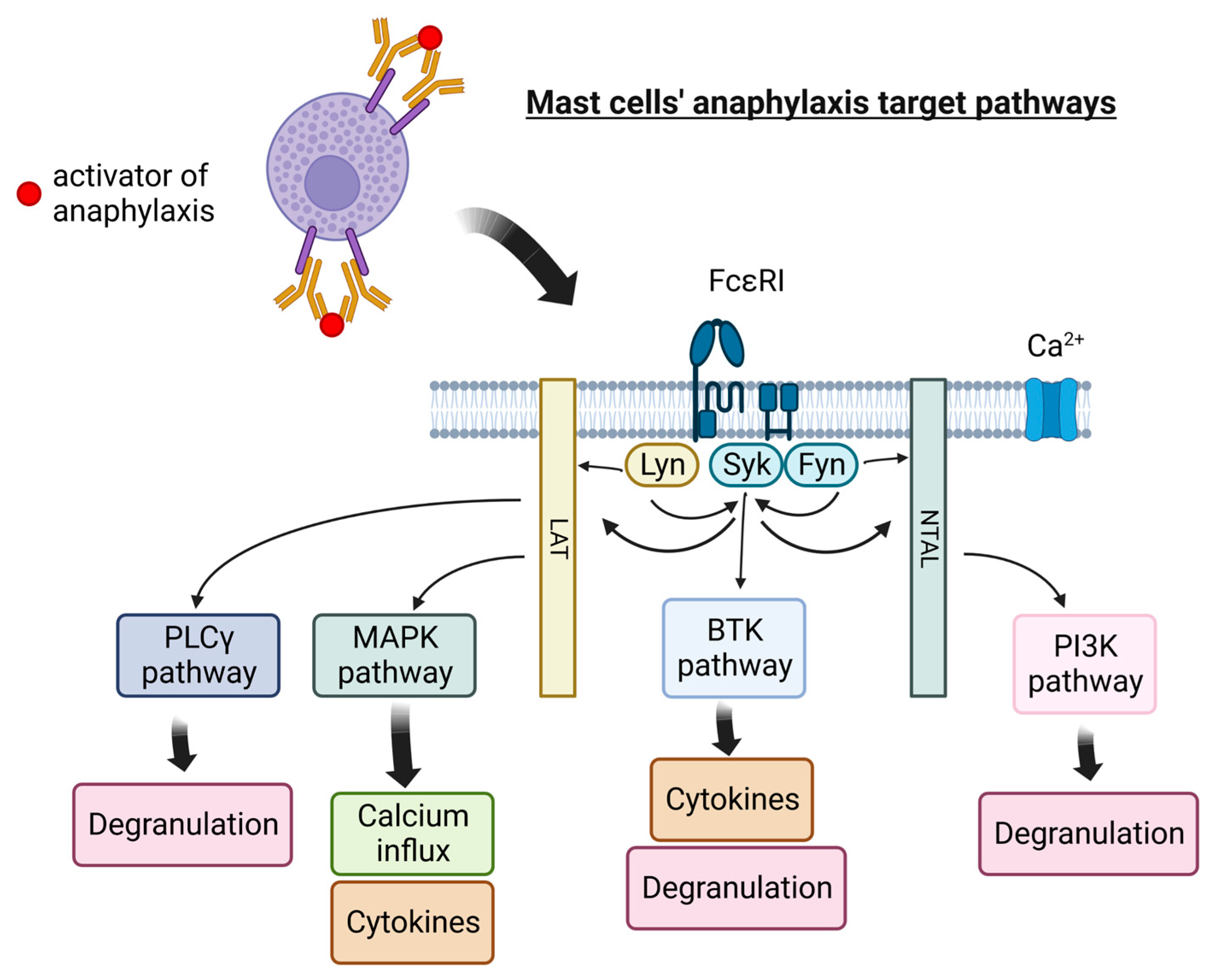
3. Mast Cells Degranulation and Its Inhibitors
4. Mast Cells as a Therapeutic Target in Allergic Inflammation
5. Mast Cell-Targeted Strategies in Cancer Therapy
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blank, U.; Falcone, F.H.; Nilsson, G. The history of mast cell and basophil research—Some lessons learnt from the last century. Allergy 2013, 68, 1093–1101. [Google Scholar] [CrossRef]
- Gentek, R.; Ghigo, C.; Hoeffel, G.; Bulle, M.J.; Msallam, R.; Gautier, G.; Launay, P.; Chen, J.; Ginhoux, F.; Bajénoff, M. Hemogenic Endothelial Fate Mapping Reveals Dual Developmental Origin of Mast Cells. Immunity 2018, 48, 1160–1171.e5. [Google Scholar] [CrossRef]
- Siraganian, R.P. Mast Cells. In Encyclopedia of Immunology, 2nd ed.; Delves, P.J., Ed.; Elsevier: Oxford, UK, 1998; pp. 1667–1671. ISBN 978-0-12-226765-9. [Google Scholar]
- Burks, A.W. (Ed.) Middleton’s Allergy: Principles and Practice, 9th ed.; Elsevier: Edinburgh, Scotland, 2020; ISBN 978-0-323-54424-5. [Google Scholar]
- Kiener, H.P.; Nigrovic, P.A.; Lee, D.M. Mast Cells. In Rheumatoid Arthritis; Elsevier: Philadelphia, PA, USA, 2009; pp. 124–135. ISBN 978-0-323-05475-1. [Google Scholar]
- Varricchi, G.; Galdiero, M.R.; Marone, G.; Granata, F.; Borriello, F.; Marone, G. Controversial role of mast cells in skin cancers. Exp. Dermatol. 2017, 26, 11–17. [Google Scholar] [CrossRef]
- Varricchi, G.; Rossi, F.W.; Galdiero, M.R.; Granata, F.; Criscuolo, G.; Spadaro, G.; de Paulis, A.; Marone, G. Physiological Roles of Mast Cells: Collegium Internationale Allergologicum Update 2019. Int. Arch. Allergy Immunol. 2019, 179, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Elst, J.; van der Poorten, M.M.; Van Gasse, A.L.; De Puysseleyr, L.; Hagendorens, M.M.; Faber, M.A.; Van Houdt, M.; Passante, E.; Bahri, R.; Walschot, M.; et al. Mast cell activation tests by flow cytometry: A new diagnostic asset? Clin. Exp. Allergy 2021, 51, 1482–1500. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Pi, J.; Wu, J.; Zhou, H.; Cai, J.; Li, T.; Liu, L. A rapid and sensitive assay based on particle analysis for cell degranulation detection in basophils and mast cells. Pharmacol. Res. 2016, 111, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Baran, J. Practical applications of peripheral blood mononuclear cells (PBMCs) in immunotherapy preclinical research. J. Curr. Sci. Technol. 2022, 12, 592604. [Google Scholar] [CrossRef]
- Piasek, A.M.; Sobiepanek, A. Medicinal potential of anandamide as a representative endocannabinoid and its effect at the cellular level in skin, in book. In Cannabis, Cannabinoids and Endocannabinoids; Academic Press: Cambridge, MA, USA, 2023. [Google Scholar]
- Sobiepanek, A.; Kuryk, Ł.; Garofalo, M.; Kumar, S.; Baran, J.; Musolf, P.; Siebenhaar, F.; Fluhr, J.W.; Kobiela, T.; Plasenzotti, R.; et al. The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers. Int. J. Mol. Sci. 2022, 23, 2249. [Google Scholar] [CrossRef]
- Musolf, P.; Baran, J.; Ścieżyńska, A.; Staniszewska, M.; Sobiepanek, A. Rola mastocytów w nadzorze odpornościowym procesów fizjologicznych i patologicznych skóry. In Zagadnienia Aktualnie Poruszane Przez Młodych Naukowców; Creativetime: Kraków, Poland, 2021; Volume 19, pp. 78–83. ISBN 978-83-66772-06-9. [Google Scholar]
- DeTurk, S.; Reddy, S.; Ng Pellegrino, A.; Wilson, J. Anaphylactic Shock. In Clinical Management of Shock—The Science and Art of Physiological Restoration; Stawicki, S.P., Swaroop, M., Eds.; IntechOpen: London, UK, 2020; ISBN 978-1-83881-170-9. [Google Scholar]
- Joo, H.M.; Nam, S.Y.; Yang, K.H.; Kim, C.S.; Jin, Y.W.; Kim, J.Y. The Effects of Low-dose Ionizing Radiation in the Activated Rat Basophilic Leukemia (RBL-2H3) Mast Cells. J. Biol. Chem. 2012, 287, 27789–27795. [Google Scholar] [CrossRef]
- Siiskonen, H.; Smorodchenko, A.; Krause, K.; Maurer, M. Ultraviolet radiation and skin mast cells: Effects, mechanisms and relevance for skin diseases. Exp. Dermatol. 2018, 27, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Sobiepanek, A.; Kowalska, P.D.; Szota, M.; Grzywa, T.M.; Nowak, J.; Włodarski, P.K.; Galus, R.; Jachimska, B.; Kobiela, T. Novel diagnostic and prognostic factors for the advanced melanoma based on the glycosylation-related changes studied by biophysical profiling methods. Biosens. Bioelectron. 2022, 203, 114046. [Google Scholar] [CrossRef]
- Sobiepanek, A.; Milner-Krawczyk, M.; Bobecka-Wesołowska, K.; Kobiela, T. The effect of delphinidin on the mechanical properties of keratinocytes exposed to UVB radiation. J. Photochem. Photobiol. B 2016, 164, 264–270. [Google Scholar] [CrossRef]
- Chadwick, S.J. Principles of Allergy Management. In Managing the Allergic Patient; Elsevier: Philadelphia, PA, USA, 2008; pp. 19–72. ISBN 978-1-4160-3677-7. [Google Scholar]
- Ohtsu, H.; Seike, M. Histamine and Histamine Receptors in Allergic Dermatitis. In Histamine and Histamine Receptors in Health and Disease; Hattori, Y., Seifert, R., Eds.; Handbook of Experimental Pharmacology; Springer International Publishing: Cham, Switzerland, 2016; Volume 241, pp. 333–345. ISBN 978-3-319-58192-7. [Google Scholar]
- Kopeć-Szlęzak, J. Mast cells and their role in immunity and cancer. J. Transfus. Med. 2015, 8, 49–59. [Google Scholar]
- Day, M.J. Glucocorticosteroids and antihistamines. In Small Animal Clinical Pharmacology; Elsevier: Amsterdam, The Netherlands, 2008; pp. 261–269. ISBN 978-0-7020-2858-8. [Google Scholar]
- Belvisi, M.G. Regulation of Inflammatory Cell Function by Corticosteroids. Proc. Am. Thorac. Soc. 2004, 1, 207–214. [Google Scholar] [CrossRef]
- Burchett, J.R.; Dailey, J.M.; Kee, S.A.; Pryor, D.T.; Kotha, A.; Kankaria, R.A.; Straus, D.B.; Ryan, J.J. Targeting Mast Cells in Allergic Disease: Current Therapies and Drug Repurposing. Cells 2022, 11, 3031. [Google Scholar] [CrossRef] [PubMed]
- Molderings, G.J.; Haenisch, B.; Brettner, S.; Homann, J.; Menzen, M.; Dumoulin, F.L.; Panse, J.; Butterfield, J.; Afrin, L.B. Pharmacological treatment options for mast cell activation disease. Naunyn. Schmiedeberg’s Arch. Pharmacol. 2016, 389, 671–694. [Google Scholar] [CrossRef]
- Conti, P.; Kempuraj, D. Impact of Vitamin D on Mast Cell Activity, Immunity and Inflammation. J. Food Nutr. Res. 2016, 4, 33–39. [Google Scholar]
- Liu, Z.-Q.; Li, X.-X.; Qiu, S.-Q.; Yu, Y.; Li, M.-G.; Yang, L.-T.; Li, L.-J.; Wang, S.; Zheng, P.-Y.; Liu, Z.-G.; et al. Vitamin D contributes to mast cell stabilization. Allergy 2017, 72, 1184–1192. [Google Scholar] [CrossRef] [PubMed]
- Kakavas, S.; Karayiannis, D.; Mastora, Z. The Complex Interplay between Immunonutrition, Mast Cells, and Histamine Signaling in COVID-19. Nutrients 2021, 13, 3458. [Google Scholar] [CrossRef] [PubMed]
- Anogeianaki, A.; Castellani, M.L.; Tripodi, D.; Toniato, E.; De Lutiis, M.A.; Conti, F.; Felaco, P.; Fulcheri, M.; Theoharides, T.C.; Galzio, R.; et al. Vitamins and Mast Cells. Int. J. Immunopathol. Pharmacol. 2010, 23, 991–996. [Google Scholar] [CrossRef]
- Matsuda, H.; Nakamura, S.; Yoshikawa, M. Degranulation Inhibitors from Medicinal Plants in Antigen-Stimulated Rat Basophilic Leukemia (RBL-2H3) Cells. Chem. Pharm. Bull. 2016, 64, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-H.; Lee, S.; Son, H.-Y.; Park, S.-B.; Kim, M.-S.; Choi, E.-J.; Singh, T.S.K.; Ha, J.-H.; Lee, M.-G.; Kim, J.-E.; et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharmacal Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef]
- Han, S.-Y.; Bae, J.-Y.; Park, S.-H.; Kim, Y.-H.; Park, J.H.Y.; Kang, Y.-H. Resveratrol Inhibits IgE-Mediated Basophilic Mast Cell Degranulation and Passive Cutaneous Anaphylaxis in Mice. J. Nutr. 2013, 143, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Shou, Q.; Tan, T.; Xu, F. Salvinorin A inhibits ovalbumin-stimulated allergic rhinitis and RBL-2H3 cells degranulation. FEBS Open Bio 2021, 11, 2166–2173. [Google Scholar] [CrossRef]
- Fu, S.; Ni, S.; Wang, D.; Hong, T. Coptisine Suppresses Mast Cell Degranulation and Ovalbumin-Induced Allergic Rhinitis. Molecules 2018, 23, 3039. [Google Scholar] [CrossRef]
- Buyanravjikh, S.; Han, S.; Lee, S.; Jeong, A.; Ka, H.; Park, J.; Boldbaatar, A.; Lim, J.; Lee, M.; Yang, Y. Cryptotanshinone inhibits IgE-mediated degranulation through inhibition of spleen tyrosine kinase and tyrosine-protein kinase phosphorylation in mast cells. Mol. Med. Rep. 2018, 18, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Kinney, S.R.M.; Carlson, L.; Ser-Dolansky, J.; Thompson, C.; Shah, S.; Gambrah, A.; Xing, W.; Schneider, S.S.; Mathias, C.B. Curcumin Ingestion Inhibits Mastocytosis and Suppresses Intestinal Anaphylaxis in a Murine Model of Food Allergy. PLoS ONE 2015, 10, e0132467. [Google Scholar] [CrossRef]
- Nishi, K.; Kanayama, Y.; Kim, I.-H.; Nakata, A.; Nishiwaki, H.; Sugahara, T. Docosahexaenoyl ethanolamide mitigates IgE-mediated allergic reactions by inhibiting mast cell degranulation and regulating allergy-related immune cells. Sci. Rep. 2019, 9, 16213. [Google Scholar] [CrossRef]
- Cruz, S.L.; Sánchez-Miranda, E.; Castillo-Arellano, J.I.; Cervantes-Villagrana, R.D.; Ibarra-Sánchez, A.; González-Espinosa, C. Anandamide inhibits FcεRI-dependent degranulation and cytokine synthesis in mast cells through CB2 and GPR55 receptor activation. Possible involvement of CB2-GPR55 heteromers. Int. Immunopharmacol. 2018, 64, 298–307. [Google Scholar] [CrossRef]
- Facci, L.; Dal Toso, R.; Romanello, S.; Buriani, A.; Skaper, S.D.; Leon, A. Mast cells express a peripheral cannabinoid receptor with differential sensitivity to anandamide and palmitoylethanolamide. Proc. Natl. Acad. Sci. USA 1995, 92, 3376–3380. [Google Scholar] [CrossRef]
- Petrosino, S.; Schiano Moriello, A.; Verde, R.; Allarà, M.; Imperatore, R.; Ligresti, A.; Mahmoud, A.M.; Peritore, A.F.; Iannotti, F.A.; Di Marzo, V. Palmitoylethanolamide counteracts substance P-induced mast cell activation in vitro by stimulating diacylglycerol lipase activity. J. Neuroinflammation 2019, 16, 274. [Google Scholar] [CrossRef]
- Cantarella, G.; Scollo, M.; Lempereur, L.; Saccani-Jotti, G.; Basile, F.; Bernardini, R. Endocannabinoids inhibit release of nerve growth factor by inflammation-activated mast cells. Biochem. Pharmacol. 2011, 82, 380–388. [Google Scholar] [CrossRef]
- Kolkhir, P.; Elieh-Ali-Komi, D.; Metz, M.; Siebenhaar, F.; Maurer, M. Understanding human mast cells: Lesson from therapies for allergic and non-allergic diseases. Nat. Rev. Immunol. 2022, 22, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Thong, B.Y.H.; Cheng, Y.K.; Leong, K.P.; Tang, C.Y.; Chng, H.H. Anaphylaxis in adults referred to a clinical immunology/allergy centre in Singapore. Singap. Med. J. 2005, 46, 529–534. [Google Scholar]
- Webb, L.M.; Lieberman, P. Anaphylaxis: A review of 601 cases. Ann. Allergy Asthma Immunol. 2006, 97, 39–43. [Google Scholar] [CrossRef]
- Gasser, P.; Eggel, A. Targeting IgE in allergic disease. Curr. Opin. Immunol. 2018, 54, 86–92. [Google Scholar] [CrossRef]
- Rigoni, A.; Colombo, M.P.; Pucillo, C. Mast cells, basophils and eosinophils: From allergy to cancer. Semin. Immunol. 2018, 35, 29–34. [Google Scholar] [CrossRef]
- Brown, J.C.; Simons, E.; Rudders, S.A. Epinephrine in the Management of Anaphylaxis. J. Allergy Clin. Immunol. Pract. 2020, 8, 1186–1195. [Google Scholar] [CrossRef]
- Kemp, S.F.; Lockey, R.F.; Simons, F.E.R. Epinephrine: The Drug of Choice for Anaphylaxis—A Statement of the World Allergy Organization. World Allergy Organ. J. 2008, 1, S18–S26. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Muñoz-Furlong, A.; Campbell, R.L.; Adkinson, N.F.; Allan Bock, S.; Branum, A.; Brown, S.G.A.; Camargo, C.A.; Cydulka, R.; Galli, S.J.; et al. Second Symposium on the Definition and Management of Anaphylaxis: Summary Report—Second National Institute of Allergy and Infectious Disease/Food Allergy and Anaphylaxis Network Symposium. Ann. Emerg. Med. 2006, 47, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Z.; Cui, L.; Xu, Y.; Guan, K.; Zhao, B. Anaphylactic risk related to omalizumab, benralizumab, reslizumab, mepolizumab, and dupilumab. Clin. Transl. Allergy 2021, 11, e12038. [Google Scholar] [CrossRef]
- Otani, T.; Iwamoto, H.; Horimasu, Y.; Yamaguchi, K.; Sakamoto, S.; Masuda, T.; Miyamoto, S.; Nakashima, T.; Fujitaka, K.; Hamada, H.; et al. Effect of dupilumab in a patient with severe asthma complicated with recurrent anaphylaxis: A case report. J. Investig. Allergol. Clin. Immunol. 2022, 33, 3. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.-Y.; Ha, D.H.; Rah, S.-Y.; Lee, D.-H.; Lee, S.-M.; Park, B.-H. Sirtuin 6 is a negative regulator of FcεRI signaling and anaphylactic responses. J. Allergy Clin. Immunol. 2022, 149, 156–167.e7. [Google Scholar] [CrossRef]
- Dispenza, M.C.; Krier-Burris, R.A.; Chhiba, K.D.; Undem, B.J.; Robida, P.A.; Bochner, B.S. Bruton’s tyrosine kinase inhibition effectively protects against human IgE-mediated anaphylaxis. J. Clin. Investig. 2020, 130, 4759–4770. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.C.; Perkins, C.; Potter, C.; Parsons, D.; Schuman, R.; Khodoun, M.V.; Samavedam, U.; Strait, R.; Finkelman, F.D. Optimizing drug inhibition of IgE-mediated anaphylaxis in mice. J. Allergy Clin. Immunol. 2022, 149, 671–684.e9. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Fan, T.; Huang, J. Anemoside B4 protects against IgE-dependent allergic responses by suppressing the PLC/IP3 and JAK/STAT3 pathways. Chem. Biol. Interact. 2022, 366, 110153. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Sun, J.; Wang, L.; Zhu, J.; Cui, W.; Hou, H.; Zhang, J.; Zhou, C.; Yan, X. Isolation and Purification of Fucoxanthin from Brown Seaweed Sargassum horneri Using Open ODS Column Chromatography and Ethanol Precipitation. Molecules 2021, 26, 3777. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Ma, Y. Restorative Effect of Fucoxanthin in an Ovalbumin-Induced Allergic Rhinitis Animal Model through NF-κB p65 and STAT3 Signaling. J. Environ. Pathol. Toxicol. Oncol. 2019, 38, 365–375. [Google Scholar] [CrossRef]
- Zhong, W.-C.; Li, E.-C.; Hao, R.-R.; Zhang, J.-F.; Jin, H.-T.; Lin, S. Anti-anaphylactic potential of benzoylpaeoniflorin through inhibiting HDC and MAPKs from Paeonia lactiflora. Chin. J. Nat. Med. 2021, 19, 825–835. [Google Scholar] [CrossRef]
- Xu, C.; Li, L.; Wang, C.; Jiang, J.; Li, L.; Zhu, L.; Jin, S.; Jin, Z.; Lee, J.J.; Li, G.; et al. Effects of G-Rh2 on mast cell-mediated anaphylaxis via AKT-Nrf2/NF-κB and MAPK-Nrf2/NF-κB pathways. J. Ginseng Res. 2022, 46, 550–560. [Google Scholar] [CrossRef] [PubMed]
- Kee, J.-Y.; Hong, S.-H. Ginsenoside Rg3 suppresses mast cell–mediated allergic inflammation via mitogen-activated protein kinase signaling pathway. J. Ginseng Res. 2019, 43, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-K.; Choo, M.-K.; Kim, E.-J.; Han, M.J.; Kim, D.-H. Antiallergic Activity of Ginsenoside Rh2. Biol. Pharm. Bull. 2003, 26, 1581–1584. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-K.; Choo, M.-K.; Han, M.J.; Kim, D.-H. Ginsenoside Rh1 Possesses Antiallergic and Anti-Inflammatory Activities. Int. Arch. Allergy Immunol. 2004, 133, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Wang, Y.; Li, C.; Zhang, Y.; Hu, S.; Gao, J.; Liu, R.; An, H. α-Linolenic acid attenuates pseudo-allergic reactions by inhibiting Lyn kinase activity. Phytomedicine 2021, 80, 153391. [Google Scholar] [CrossRef]
- Rivera, J.; Olivera, A. Src family kinases and lipid mediators in control of allergic inflammation. Immunol. Rev. 2007, 217, 255–268. [Google Scholar] [CrossRef]
- Wang, Y.; Ding, Y.; Li, C.; Gao, J.; Wang, X.; An, H. Alpha-linolenic acid inhibits IgE-mediated anaphylaxis by inhibiting Lyn kinase and suppressing mast cell activation. Int. Immunopharmacol. 2022, 103, 108449. [Google Scholar] [CrossRef]
- Ashikari, T.; Hachisu, M.; Nagata, K.; Ando, D.; Iizuka, Y.; Ito, N.; Ito, K.; Ikeda, Y.; Matsubara, H.; Yashiro, T.; et al. Salicylaldehyde Suppresses IgE-Mediated Activation of Mast Cells and Ameliorates Anaphylaxis in Mice. Int. J. Mol. Sci. 2022, 23, 8826. [Google Scholar] [CrossRef]
- Simons, K.J.; Simons, F.E.R. Epinephrine and its use in anaphylaxis: Current issues. Curr. Opin. Allergy Clin. Immunol. 2010, 10, 354–361. [Google Scholar] [CrossRef]
- Gilfillan, A.M.; Beaven, M.A. Regulation of Mast Cell Responses in Health and Disease. Crit. Rev. Immunol. 2011, 31, 475–530. [Google Scholar] [CrossRef]
- Weller, C.L.; Collington, S.J.; Williams, T.; Lamb, J.R. Mast cells in health and disease. Clin. Sci. 2011, 120, 473–484. [Google Scholar] [CrossRef]
- Dawicki, W.; Marshall, J.S. New and emerging roles for mast cells in host defence. Curr. Opin. Immunol. 2007, 19, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Oldford, S.A.; Marshall, J.S. Mast cells as targets for immunotherapy of solid tumors. Mol. Immunol. 2015, 63, 113–124. [Google Scholar] [CrossRef]
- Theoharides, T.C.; Alysandratos, K.-D.; Angelidou, A.; Delivanis, D.-A.; Sismanopoulos, N.; Zhang, B.; Asadi, S.; Vasiadi, M.; Weng, Z.; Miniati, A.; et al. Mast cells and inflammation. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2012, 1822, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Theoharides, T.C.; Conti, P. Mast cells: The JEKYLL and HYDE of tumor growth. Trends Immunol. 2004, 25, 235–241. [Google Scholar] [CrossRef]
- Aponte-López, A.; Fuentes-Pananá, E.M.; Cortes-Muñoz, D.; Muñoz-Cruz, S. Mast Cell, the Neglected Member of the Tumor Microenvironment: Role in Breast Cancer. J. Immunol. Res. 2018, 2018, 2584243. [Google Scholar] [CrossRef]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Donato, G.; Montemurro, S.; Ruggieri, E.; Patruno, R.; Marech, I.; Cariello, M.; Vacca, A.; et al. Correlation between Serum Tryptase, Mast Cells Positive to Tryptase and Microvascular Density in Colo-Rectal Cancer Patients: Possible Biological-Clinical Significance. PLoS ONE 2014, 9, e99512. [Google Scholar] [CrossRef]
- Elezoglu, B.; Tolunay, S. The relationship between the stromal mast cell number, microvessel density, c-erbb-2 staining and survival and prognostic factors in colorectal carcinoma. Turk. J. Pathol. 2012, 28, 110. [Google Scholar] [CrossRef]
- Gulubova, M.; Vlaykova, T. Prognostic significance of mast cell number and microvascular density for the survival of patients with primary colorectal cancer. J. Gastroenterol. Hepatol. 2009, 24, 1265–1275. [Google Scholar] [CrossRef]
- Wu, X.; Zou, Y.; He, X.; Yuan, R.; Chen, Y.; Lan, N.; Lian, L.; Wang, F.; Fan, X.; Zeng, Y.; et al. Tumor-Infiltrating Mast Cells in Colorectal Cancer as a Poor Prognostic Factor. Int. J. Surg. Pathol. 2013, 21, 111–120. [Google Scholar] [CrossRef]
- Ammendola, M.; Marech, I.; Sammarco, G.; Zuccalà, V.; Luposella, M.; Zizzo, N.; Patruno, R.; Crovace, A.; Ruggieri, E.; Zito, A.; et al. Infiltrating Mast Cells Correlate with Angiogenesis in Bone Metastases from Gastric Cancer Patients. Int. J. Mol. Sci. 2015, 16, 3237–3250. [Google Scholar] [CrossRef] [PubMed]
- Ammendola, M.; Sacco, R.; Zuccalà, V.; Luposella, M.; Patruno, R.; Gadaleta, P.; Zizzo, N.; Gadaleta, C.; De Sarro, G.; Sammarco, G.; et al. Mast Cells Density Positive to Tryptase Correlate with Microvascular Density in both Primary Gastric Cancer Tissue and Loco-Regional Lymph Node Metastases from Patients That Have Undergone Radical Surgery. Int. J. Mol. Sci. 2016, 17, 1905. [Google Scholar] [CrossRef]
- Micu, G.V.; Stăniceanu, F.; Sticlaru, L.C.; Popp, C.G.; Bastian, A.E.; Gramada, E.; Pop, G.; Mateescu, R.B.; Rimbaş, M.; Archip, B.; et al. Correlations Between the Density of Tryptase Positive Mast Cells (DMCT) and that of New Blood Vessels (CD105+) in Patients with Gastric Cancer. Rom. J. Intern. Med. 2016, 54, 113–120. [Google Scholar] [CrossRef]
- Ribatti, D.; Guidolin, D.; Marzullo, A.; Nico, B.; Annese, T.; Benagiano, V.; Crivellato, E. Mast cells and angiogenesis in gastric carcinoma: Mast cells and angiogenesis in gastric carcinoma. Int. J. Exp. Pathol. 2010, 91, 350–356. [Google Scholar] [CrossRef]
- Cai, S.-W.; Yang, S.-Z.; Gao, J.; Pan, K.; Chen, J.-Y.; Wang, Y.-L.; Wei, L.-X.; Dong, J.-H. Prognostic significance of mast cell count following curative resection for pancreatic ductal adenocarcinoma. Surgery 2011, 149, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.Z.; Ma, Y.; Ji, B.; Wang, H.; Deng, D.; Liu, Y.; Abbruzzese, J.L.; Liu, Y.; Logsdon, C.D.; Hwu, P. Mast Cells in Tumor Microenvironment Promotes the In Vivo Growth of Pancreatic Ductal Adenocarcinoma. Clin. Cancer Res. 2011, 17, 7015–7023. [Google Scholar] [CrossRef] [PubMed]
- Porcelli, L.; Iacobazzi, R.; Di Fonte, R.; Serratì, S.; Intini, A.; Solimando, A.; Brunetti, O.; Calabrese, A.; Leonetti, F.; Azzariti, A.; et al. CAFs and TGF-β Signaling Activation by Mast Cells Contribute to Resistance to Gemcitabine/Nabpaclitaxel in Pancreatic Cancer. Cancers 2019, 11, 330. [Google Scholar] [CrossRef]
- Strouch, M.J.; Cheon, E.C.; Salabat, M.R.; Krantz, S.B.; Gounaris, E.; Melstrom, L.G.; Dangi-Garimella, S.; Wang, E.; Munshi, H.G.; Khazaie, K.; et al. Crosstalk between Mast Cells and Pancreatic Cancer Cells Contributes to Pancreatic Tumor Progression. Clin. Cancer Res. 2010, 16, 2257–2265. [Google Scholar] [CrossRef]
- Carpenco, E.; Ceauşu, R.A.; Cimpean, A.M.; Gaje, P.N.; Șaptefraţi, L.; Fulga, V.; David, V.; Raica, M. Mast Cells as an Indicator and Prognostic Marker in Molecular Subtypes of Breast Cancer. In Vivo 2019, 33, 743–748. [Google Scholar] [CrossRef]
- Fakhrjou, A.; Naghavi-Behzad, M.; Montazeri, V.; Karkon-Shayan, F.; Norouzi-Panahi, L.; Piri, R. The relationship between histologic grades of invasive carcinoma of breast ducts and mast cell infiltration. South Asian J. Cancer 2016, 05, 005–007. [Google Scholar] [CrossRef]
- Keser, S.H.; Kandemir, N.O.; Ece, D.; Gecmen, G.G.; Gul, A.E.; Barisik, N.O.; Sensu, S.; Buyukuysal, C.; Barut, F. Relationship of mast cell density with lymphangiogenesis and prognostic parameters in breast carcinoma. Kaohsiung J. Med. Sci. 2017, 33, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Marech, I.; Ammendola, M.; Sacco, R.; Capriuolo, G.S.; Patruno, R.; Rubini, R.; Luposella, M.; Zuccalà, V.; Savino, E.; Gadaleta, C.D.; et al. Serum tryptase, mast cells positive to tryptase and microvascular density evaluation in early breast cancer patients: Possible translational significance. BMC Cancer 2014, 14, 534. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.M.; Reuben, A.; Barua, S.; Jiang, H.; Zhang, S.; Wang, L.; Gopalakrishnan, V.; Hudgens, C.W.; Tetzlaff, M.T.; Reuben, J.M.; et al. Poor Response to Neoadjuvant Chemotherapy Correlates with Mast Cell Infiltration in Inflammatory Breast Cancer. Cancer Immunol. Res. 2019, 7, 1025–1035. [Google Scholar] [CrossRef]
- Carlini, M.J.; Dalurzo, M.C.L.; Lastiri, J.M.; Smith, D.E.; Vasallo, B.C.; Puricelli, L.I.; de Cidre, L.S.L. Mast cell phenotypes and microvessels in non–small cell lung cancer and its prognostic significance. Hum. Pathol. 2010, 41, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Imada, A.; Shijubo, N.; Kojima, H.; Abe, S. Mast cells correlate with angiogenesis and poor outcome in stage I lung adenocarcinoma. Eur. Respir. J. 2000, 15, 1087–1093. [Google Scholar] [CrossRef]
- Takanami, I.; Takeuchi, K.; Naruke, M. Mast cell density is associated with angiogenesis and poor prognosis in pulmonary adenocarcinoma. Cancer 2000, 88, 2686–2692. [Google Scholar] [CrossRef]
- Nonomura, N.; Takayama, H.; Nishimura, K.; Oka, D.; Nakai, Y.; Shiba, M.; Tsujimura, A.; Nakayama, M.; Aozasa, K.; Okuyama, A. Decreased number of mast cells infiltrating into needle biopsy specimens leads to a better prognosis of prostate cancer. Br. J. Cancer 2007, 97, 952–956. [Google Scholar] [CrossRef]
- Pittoni, P.; Tripodo, C.; Piconese, S.; Mauri, G.; Parenza, M.; Rigoni, A.; Sangaletti, S.; Colombo, M.P. Mast Cell Targeting Hampers Prostate Adenocarcinoma Development but Promotes the Occurrence of Highly Malignant Neuroendocrine Cancers. Cancer Res. 2011, 71, 5987–5997. [Google Scholar] [CrossRef]
- Bahri, R.; Kiss, O.; Prise, I.; Garcia-Rodriguez, K.M.; Atmoko, H.; Martínez-Gómez, J.M.; Levesque, M.P.; Dummer, R.; Smith, M.P.; Wellbrock, C.; et al. Human Melanoma-Associated Mast Cells Display a Distinct Transcriptional Signature Characterized by an Upregulation of the Complement Component 3 That Correlates With Poor Prognosis. Front. Immunol. 2022, 13, 861545. [Google Scholar] [CrossRef]
- Ammendola, M.; Sacco, R.; Sammarco, G.; Luposella, M.; Patruno, R.; Gadaleta, C.D.; De Sarro, G.; Ranieri, G. Mast Cell-Targeted Strategies in Cancer Therapy. Transfus. Med. Hemotherapy 2016, 43, 109–113. [Google Scholar] [CrossRef]
- Faustino-Rocha, A.I.; Gama, A.; Oliveira, P.A.; Vanderperren, K.; Saunders, J.H.; Pires, M.J.; Ferreira, R.; Ginja, M. Modulation of mammary tumor vascularization by mast cells: Ultrasonographic and histopathological approaches. Life Sci. 2017, 176, 35–41. [Google Scholar] [CrossRef]
- Iqbal, N.; Iqbal, N. Imatinib: A Breakthrough of Targeted Therapy in Cancer. Chemother. Res. Pract. 2014, 2014, 357027. [Google Scholar] [CrossRef] [PubMed]
- Cimpean, A.M.; Raica, M. The Hidden Side of Disodium Cromolyn: From Mast Cell Stabilizer to an Angiogenic Factor and Antitumor Agent. Arch. Immunol. Ther. Exp. 2016, 64, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Marech, I.; Ammendola, M.; Gadaleta, C.; Zizzo, N.; Oakley, C.; Gadaleta, C.D.; Ranieri, G. Possible biological and translational significance of mast cells density in colorectal cancer. World J. Gastroenterol. 2014, 20, 8910–8920. Available online: https://www.wjgnet.com/1007-9327/full/v20/i27/8910.htm (accessed on 9 March 2023). [PubMed]
- Alzforum. Mastinib. Available online: www.alzforum.org/therapeutics/mastinib (accessed on 20 January 2023).
- Dubreuil, P.; Letard, S.; Ciufolini, M.; Gros, L.; Humbert, M.; Castéran, N.; Borge, L.; Hajem, B.; Lermet, A.; Sippl, W.; et al. Masitinib (AB1010), a Potent and Selective Tyrosine Kinase Inhibitor Targeting KIT. PLoS ONE 2009, 4, e7258. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Mast cells as therapeutic target in cancer. Eur. J. Pharmacol. 2016, 778, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Demetri, G.D.; van Oosterom, A.T.; Garrett, C.R.; Blackstein, M.E.; Shah, M.H.; Verweij, J.; McArthur, G.; Judson, I.R.; Heinrich, M.C.; Morgan, J.A.; et al. Efficacy and safety of sunitinib in patients with advanced gastrointestinal stromal tumour after failure of imatinib: A randomised controlled trial. Lancet 2006, 368, 1329–1338. [Google Scholar] [CrossRef]
- Ohno, H.; Kosaki, G.; Kambayashi, J.; Imaoka, S.; Hirata, F. FOY: [Ethylp-(6-guanidinohexanoyloxy) benzoate] methanesulfonate as a serine proteinase inhibitor. I. Inhibition of thrombin and factor Xa in vitro. Thromb. Res. 1980, 19, 579–588. [Google Scholar] [CrossRef]
- Tamura, Y.; Hirado, M.; Okamura, K.; Minato, Y.; Fujii, S. Synthetic inhibitors of trypsin, plasmin, kallikrein, thrombin, and C1 esterase. Biochim. Biophys. Acta BBA Enzymol. 1977, 484, 417–422. [Google Scholar] [CrossRef]
- Yoon, W.-H.; Jung, Y.-J.; Kim, T.-D.; Li, G.; Park, B.-J.; Kim, J.-Y.; Lee, Y.-C.; Kim, J.-M.; Park, J.-I.; Park, H.-D.; et al. Gabexate Mesilate Inhibits Colon Cancer Growth, Invasion, and Metastasis by Reducing Matrix Metalloproteinases and Angiogenesis. Clin. Cancer Res. 2004, 10, 4517–4526. [Google Scholar] [CrossRef]
- PubChem. Nafamostat Mesylate. Available online: Pubchem.ncbi.nlm.nih.gov/compound/Nafamostat-mesylate (accessed on 20 January 2023).




{kind=link}
{kind=link}
{kind=link}
| Name of the Drug | Origin or Family of the Drug Molecule | Target | Disease, Animal or Cell Model | References |
|---|---|---|---|---|
| Olopatadine | Antihistamines | stabilization of cell membrane | various mast cells | [15] |
| Budesonide | Glucocorticosteroids | stabilization of cell membrane | airway mucosal biopsy specimen from human | [19] |
| Omalizumab | Monoclonal antibodies | IgE | FDA-approved drug | [20] |
| Ligelizumab | IgE | clinical trials | ||
| Dupilumab | interleukin signaling | FDA-approved drug | ||
| MTPS9579A | tryptase | clinical trials | ||
| CDX-0159 | c-kit/CD117 receptor | clinical trials | ||
| Vit. C | Vitamins | inhibiting histidine decarboxylase activity (the enzyme responsible for the production of histamine from histidine) | blood, urine, and tissue (lung, gastric mucosa, and spleen) of guinea pigs; blood serum of human patients with allergic and non-allergic diseases | [21] |
| Vit. D | binding to the gene transcription apparatus (regulating gene expression of selected mediators like TNF-α) | Animal models: Male BALB/c mice, Sprague Dawley rats; Cell lines: human mast cell line HMC1, rat mast cell line RBL-2H3, mouse mast cell lines p815, and mouse BMMCs | [23] | |
| Fisetin, kaempferol, myricetin, quercetin, and rutin | Flavonoids | precise target unknown (inhibited histamine release and intracellular calcium ions) | rat mast cells RBL-2H3 | [27] |
| Salvinorin A | Diterpenoid | precise target unknown (reduced release of β-hex, histamine, IL-4 and TNF-α) | rat mast cells RBL-2H3 | [29] |
| Coptisine | Alkaloids | precise target unknown (decreased levels of β-hex, histamine, IL-4 and TNF-α) | rat mast cells RBL-2H3 | [30] |
| Curcumin, monomethylcurcumin and bisdemethoxycurcumin | Curcuminoids | precise target unknown (reduced release of β-hex and inhibition of IL-4 and TNF-α production) | rat mast cells RBL-2H3; BALB/c mice and mouse BMMCs | [26,32] |
| Docosahexaenoyl ethanolamide | Derivatives of fatty acids | IgE-mediated degranulation (decreased release of β-hex and calcium ions influx) | rat mast cells RBL-2H3 | [33] |
| Anandamide | FcεRI-dependent degranulation (reduced intracellular calcium ions) | mouse BMMCs | [34] | |
| Palmitoylethanolamide | precise target unknown (reducing serotonin, histamine, β-hex and NGF) | rat mast cells RBL-2H3 | [35,37,38] |
| Name of the Drug | Origin or Family of the Drug Molecule | Target | Disease, Animal or Cell Model | References |
|---|---|---|---|---|
| Epinephrine | Adrenergic drug | Adrenergic receptors: α1 receptor α2 receptor β1 receptor β2 receptor | Emergency treatment for acute allergic reactions (anaphylaxis) caused by peanuts or other foods, medications, insect bites and stings, and other allergens, as well as exercise-induced or idiopathic anaphylaxis. | [47,48,67] |
| Albuterol | β2 adrenergic receptor | Prevention and treatment of difficulty breathing, wheezing, shortness of breath, coughing, and chest tightness caused by asthma and chronic obstructive pulmonary disease. | [49] | |
| Dupilumab | Monoclonal antibody | IL-4Rα | Atopic dermatitis. | [50,51] |
| Sirtuin 6 | NAD+-dependent deacetylase | Suppressor of PTPRC (Protein tyrosine phosphatase, receptor-type C) transcription | Murine bone marrow-derived mast cells (BMMCs), Human cord blood-derived mast cells, Myeloid Sirt6 KO mice (Sirt6flox/flox; LysM-Cre) Mast cell-deficient KitW-sh/W-sh mice | [52] |
| Ibrutinib | Bruton’s tyrosine kinase inhibitors | Bruton’s tyrosine kinase | Mantle cell lymphoma, chronic lymphotic leukemia and Waldenstroms macroglobuline-mia. Clinically tested for food-induced anaphylaxis. | NCT03149315 [53] |
| Acalabrutinib | As monotherapy or in combination with obinutuzumab, it is indicated for the treatment of adult patients with previously untreated chronic lymphocytic leukemia. Clinically tested for food-induced anaphylaxis. | NCT05038904 [53] | ||
| Tirabrutinib | Treatment of recurrent or refractory primary central nervous system lymphoma and under review for the treatment of Waldenström’s macroglobulinemia and lymphoplasmacytic lymphoma. For anaphylaxis treatment tested on primary human skin-derived mast cells (SDMCs). | [53] | ||
| Anemoside B4 | PLC/IP3 and JAK/STAT3 pathways inhibitor | The exact target requires further research. | Laboratory of allergic disease 2 (LAD2) cell line and in vivo mice model. | [55] |
| Fucoxanthin | Natural compound | The exact target requires further research. | BALB/c mice | [56,57] |
| Benzoylpaeoniflorin | [58] | |||
| Ginsenosides | AKT-NrF2 and p38MAPK-Nrf pathways inhibitor | The exact target requires further research. | Cell lines: HMC-1 (human mast cell line) RBL-2H3 (rat basophilic leukemia cell line) RAW 264.7 cells (murine macrophages) RBL 2H3 cells (rat basophils) Animals: Male Hartley guinea pigs Male ICR mice SD rats | [60,61,62] |
| Alpha-linolenic acid | Lyn kinase inhibitor | Lyn kinase | Laboratory of allergic disease 2 (LAD2) cell line C57BL/6 wild-type (WT) mice | [65] |
| Salicylaldehyde | Passive anaphylaxis mouse model using C57BL/6J and BALB/c | [66] |
| Name of the Drug | Origin or Family of the Drug Molecule | Target | Disease, Animal or Cell Model | References |
|---|---|---|---|---|
| Mastinib | c-Kit tyrosine kinase inhibitor | Protein tyrosine kinase c-kit expressed on the surface of cancer cells PDGF and FGF receptors Fyn and lyn kinases | Mastocitosis Gastrointestinal stromal tumors Colon cancer Prostate cancer Pancreatic cancer | [81,101,104,105,106] NCT00998751 NCT03556956 NCT03761225 |
| Imatinib | Chronic myeloid leukemia Gastrointestinal stromal tumor | [81,101,107] | ||
| Sunitinib | Renal cell carcinoma Gastrointestinal stromal tumor | [81,101,107] | ||
| Gabexate mesylate | Serine protease inhibitor | Thrombin Plasmin Kallikrein Trypsin C1 esterase Coagulation factor Xa | Colon cancer | [81,102,108,109,110] |
| Nafomast mesylate | Prothrombin Coagulation factor X Coagulation factor XII Trypsin Kallikrein Intercellular adhesion molecules (ICAM) | Pancreatitis Antiviral and anti-cancer properties | [81,102,111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baran, J.; Sobiepanek, A.; Mazurkiewicz-Pisarek, A.; Rogalska, M.; Gryciuk, A.; Kuryk, L.; Abraham, S.N.; Staniszewska, M. Mast Cells as a Target—A Comprehensive Review of Recent Therapeutic Approaches. Cells 2023, 12, 1187. https://doi.org/10.3390/cells12081187
Baran J, Sobiepanek A, Mazurkiewicz-Pisarek A, Rogalska M, Gryciuk A, Kuryk L, Abraham SN, Staniszewska M. Mast Cells as a Target—A Comprehensive Review of Recent Therapeutic Approaches. Cells. 2023; 12(8):1187. https://doi.org/10.3390/cells12081187
Chicago/Turabian StyleBaran, Joanna, Anna Sobiepanek, Anna Mazurkiewicz-Pisarek, Marta Rogalska, Aleksander Gryciuk, Lukasz Kuryk, Soman N. Abraham, and Monika Staniszewska. 2023. "Mast Cells as a Target—A Comprehensive Review of Recent Therapeutic Approaches" Cells 12, no. 8: 1187. https://doi.org/10.3390/cells12081187
APA StyleBaran, J., Sobiepanek, A., Mazurkiewicz-Pisarek, A., Rogalska, M., Gryciuk, A., Kuryk, L., Abraham, S. N., & Staniszewska, M. (2023). Mast Cells as a Target—A Comprehensive Review of Recent Therapeutic Approaches. Cells, 12(8), 1187. https://doi.org/10.3390/cells12081187