
Shining the Light on Astrocytic Ensembles



Abstract
:1. Introduction
2. Available Methods for Tracking Astrocytes

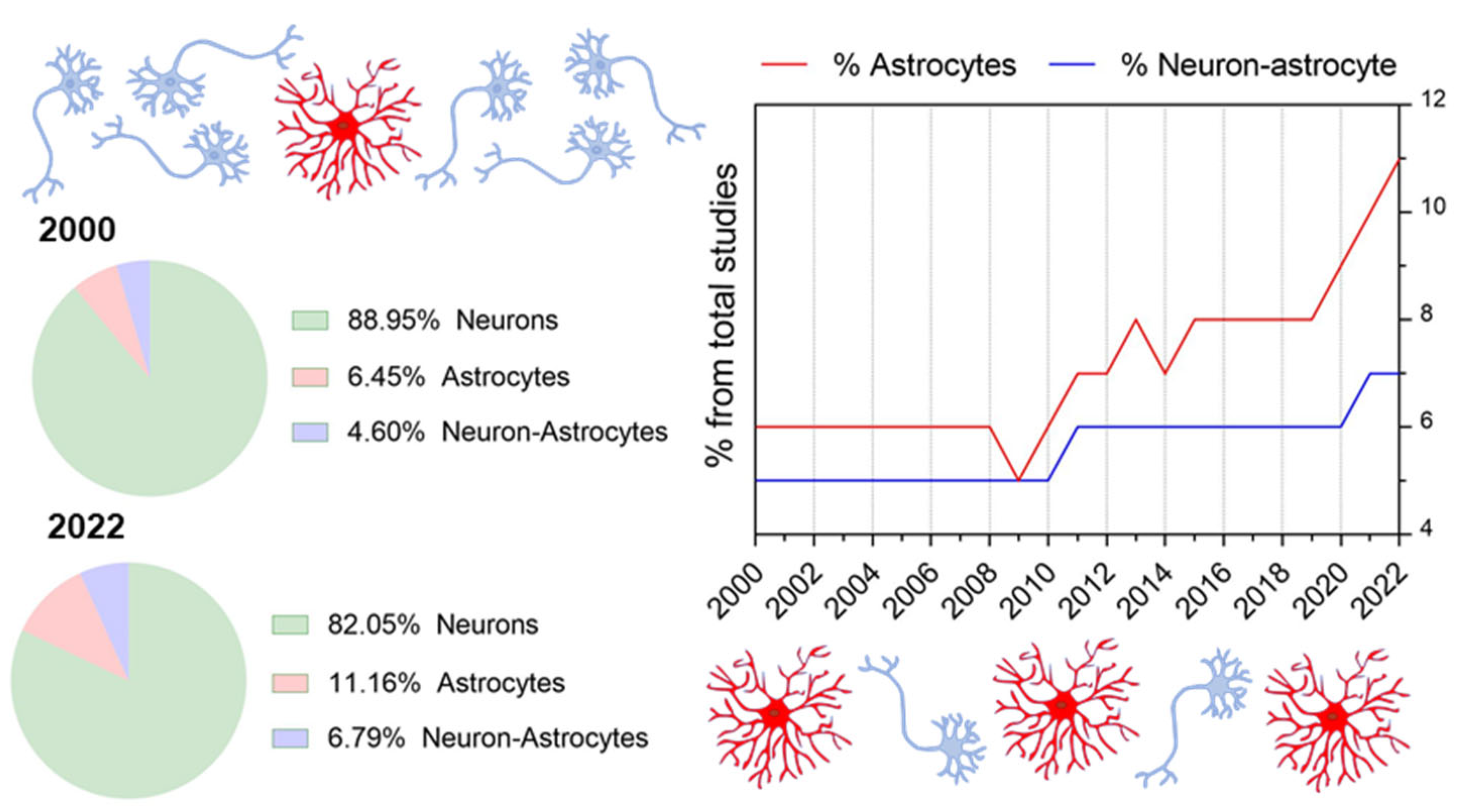{kind=link}
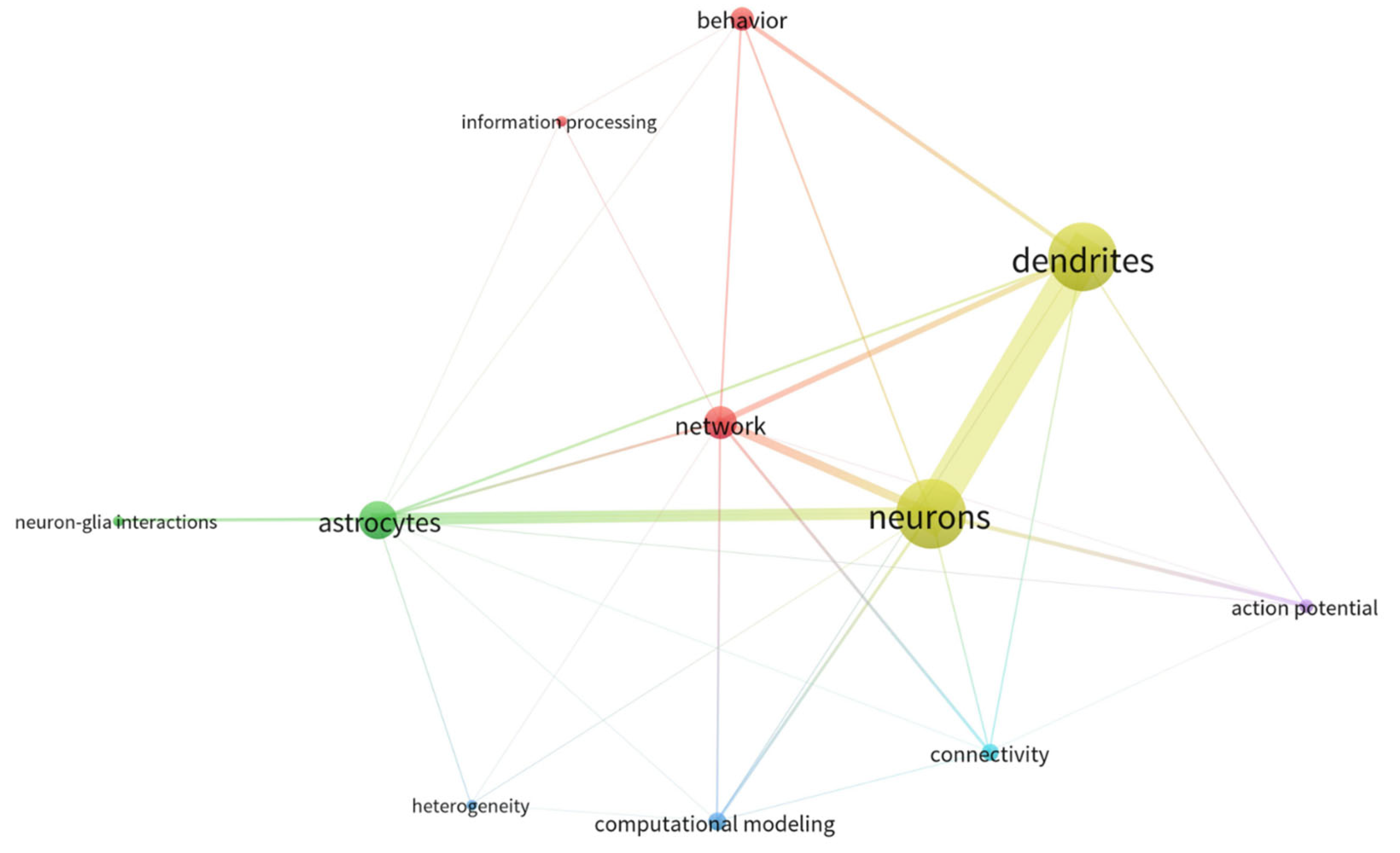{kind=link}
{kind=link}
| Experimental Techniques | ||
|---|---|---|
| Structure and/or Morphology of Astrocytes | References | |
| Labeling |
| [63] |
| [64] | |
| [65] | |
| [37] | |
| [66] | |
| Imaging |
| [67,68,69,70,71,72] |
| [118,119] | |
| [120] | |
| [121] | |
| [122,123] | |
| Labeling and Imaging |
| [73] |
| Labeling and Imaging and FRET Phenomenon |
| [30,74] |
| Genetic Programs and Molecular Profiles | ||
| RNA-Seq and Single Cell RNA-Seq |
| [80] |
| [81,82] | |
| [83,84] | |
| [85,86] | |
| [87,88] | |
| Spatial Transcriptomics |
| [92] |
| [93] | |
| [94] | |
| Activity of Astrocytes | ||
| Ca2+ Specific Dyes for Intracellular Measurements |
| [59,60] |
| Genetically Encoded Ca2+Indicators(GECIs) |
| [3,97] |
| [98,99,100] | |
| [58,101,102,106,107] | |
| Activation of Astrocytes |
| [109,110] |
| [111] | |
| [130] | |
| [30,113] | |
| [114] | |
| Inhibition of astrocytes |
| [115,116] |
| [117] | |
| Analytical Tools | References | Properties |
|---|---|---|
| Astrocyte Quantitative Analysis (AQuA) | [125] |
|
| GECIquant | [126] |
|
| CaSCaDe | [127] |
|
| Partition in Regular Quadrants (PRQ) | [3] |
|
| Calsee | [128] |
|
| Astral | [129] |
|
3. Astrocytes Add Complexity and Increase Integration Versatility
3.1. Definition of Functional Astrocytic Ensembles
3.2. Could Astrocytes Undergo Specialization?
3.3. The In Vivo Study of Astrocytic Processing in Behavior
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yu, X. Hippocampal Astrocytes Represent Navigation Space. PLoS Biol. 2022, 20, e3001568. [Google Scholar] [CrossRef] [PubMed]
- Doron, A.; Rubin, A.; Benmelech-Chovav, A.; Benaim, N.; Carmi, T.; Refaeli, R.; Novick, N.; Kreisel, T.; Ziv, Y.; Goshen, I. Hippocampal Astrocytes Encode Reward Location. Nature 2022, 609, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Serra, I.; Esparza, J.; Delgado, L.; Martín-Monteagudo, C.; Puigròs, M.; Podlesniy, P.; Trullás, R.; Navarrete, M. Ca2+-Modulated Photoactivatable Imaging Reveals Neuron-Astrocyte Glutamatergic Circuitries within the Nucleus Accumbens. Nat. Commun. 2022, 13, 5272. [Google Scholar] [CrossRef] [PubMed]
- Volterra, A.; Liaudet, N.; Savtchouk, I. Astrocyte Ca2+ Signalling: An Unexpected Complexity. Nat. Rev. Neurosci. 2014, 15, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J.; Bootman, M.D.; Roderick, H.L. Calcium Signalling: Dynamics, Homeostasis and Remodelling. Nat. Rev. Mol. Cell Biol. 2003, 4, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; McCarthy, K.D. Astrocyte Calcium Signaling: From Observations to Functions and the Challenges Therein. Cold Spring Harb. Perspect. Biol. 2015, 7, a020404. [Google Scholar] [CrossRef]
- Nett, W.J.; Oloff, S.H.; Mccarthy, K.D. Hippocampal Astrocytes In Situ Exhibit Calcium Oscillations That Occur Independent of Neuronal Activity. J. Neurophysiol. 2002, 87, 528–537. [Google Scholar] [CrossRef]
- Ding, F.; O’Donnell, J.; Thrane, A.S.; Zeppenfeld, D.; Kang, H.; Xie, L.; Wang, F.; Nedergaard, M. A1-Adrenergic Receptors Mediate Coordinated Ca2+ Signaling of Cortical Astrocytes in Awake, Behaving Mice. Cell Calcium 2013, 54, 387–394. [Google Scholar] [CrossRef]
- Cornell-Bell, A.H.; Finkbeiner, S.M.; Cooper, M.S.; Smith, S.J. Glutamate Induces Calcium Waves in Cultured Astrocytes: Long-Range Glial Signaling. Science 1990, 247, 470–473. [Google Scholar] [CrossRef]
- Palygin, O.; Lalo, U.; Verkhratsky, A.; Pankratov, Y. Ionotropic NMDA and P2X1/5 Receptors Mediate Synaptically Induced Ca2+ Signalling in Cortical Astrocytes. Cell Calcium 2010, 48, 225–231. [Google Scholar] [CrossRef]
- Araque, A.; Carmignoto, G.; Haydon, P.G. Dynamic Signaling between Astrocytes and Neurons. Annu. Rev. Physiol. 2001, 63, 795–813. [Google Scholar] [CrossRef] [PubMed]
- Araque, A.; Carmignoto, G.; Haydon, P.G.; Oliet, S.H.R.; Robitaille, R.; Volterra, A. Gliotransmitters Travel in Time and Space. Neuron 2014, 81, 728–739. [Google Scholar] [CrossRef] [PubMed]
- Halassa, M.M.; Haydon, P.G. Integrated Brain Circuits: Astrocytic Networks Modulate Neuronal Activity and Behavior. Annu. Rev. Physiol. 2010, 72, 335. [Google Scholar] [CrossRef] [PubMed]
- Perea, G.; Navarrete, M.; Araque, A. Tripartite Synapses: Astrocytes Process and Control Synaptic Information. Trends Neurosci. 2009, 32, 421–431. [Google Scholar] [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from Brain Glue to Communication Elements: The Revolution Continues. Nat. Rev. Neurosci. 2005, 6, 626–640. [Google Scholar] [CrossRef]
- Aguado, F.; Espinosa-Parrilla, J.F.; Carmona, M.A.; Soriano, E. Neuronal Activity Regulates Correlated Network Properties of Spontaneous Calcium Transients in Astrocytes In Situ. J. Neurosci. 2002, 22, 9430–9444. [Google Scholar] [CrossRef]
- Chung, W.S.; Allen, N.J.; Eroglu, C. Astrocytes Control Synapse Formation, Function, and Elimination. Cold Spring Harb. Perspect. Biol. 2015, 7, a020370. [Google Scholar] [CrossRef]
- Baldwin, K.T.; Eroglu, C. Molecular Mechanisms of Astrocyte-Induced Synaptogenesis. Curr. Opin. Neurobiol. 2017, 45, 113–120. [Google Scholar] [CrossRef]
- Allen, N.J.; Eroglu, C. Cell Biology of Astrocyte-Synapse Interactions. Neuron 2017, 96, 697–708. [Google Scholar] [CrossRef]
- Stork, T.; Sheehan, A.; Tasdemir-Yilmaz, O.E.; Freeman, M.R. Neuron-Glia Interactions through the Heartless FGF Receptor Signaling Pathway Mediate Morphogenesis of Drosophila Astrocytes. Neuron 2014, 83, 388–403. [Google Scholar] [CrossRef]
- Oberheim, N.A.; Wang, X.; Goldman, S.; Nedergaard, M. Astrocytic Complexity Distinguishes the Human Brain. Trends Neurosci. 2006, 29, 547–553. [Google Scholar] [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite Synapses: Glia, the Unacknowledged Partner. Trends Neurosci. 1999, 22, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Kleim, J.A.; Markham, J.A.; Vij, K.; Freese, J.L.; Ballard, D.H.; Greenough, W.T. Motor Learning Induces Astrocytic Hypertrophy in the Cerebellar Cortex. Behav. Brain Res. 2007, 178, 244–249. [Google Scholar] [CrossRef]
- Allen, N.J. Astrocyte Regulation of Synaptic Behavior. Annu. Rev. Cell Dev. Biol. 2014, 30, 439–463. [Google Scholar] [CrossRef] [PubMed]
- Bernardinelli, Y.; Muller, D.; Nikonenko, I. Astrocyte-Synapse Structural Plasticity. Neural Plast. 2014, 2014, 1. [Google Scholar] [CrossRef]
- Arizono, M.; Inavalli, V.V.G.K.; Bancelin, S.; Fernández-Monreal, M.; Nägerl, U.V. Super-Resolution Shadow Imaging Reveals Local Remodeling of Astrocytic Microstructures and Brain Extracellular Space after Osmotic Challenge. Glia 2021, 69, 1605–1613. [Google Scholar] [CrossRef]
- Theodosis, D.T.; Poulain, D.A.; Oliet, S.H.R. Activity-Dependent Structural and Functional Plasticity of Astrocyte-Neuron Interactions. Physiol. Rev. 2008, 88, 983–1008. [Google Scholar] [CrossRef] [PubMed]
- Hirrlinger, J.; Hülsmann, S.; Kirchhoff, F. Astroglial Processes Show Spontaneous Motility at Active Synaptic Terminals In Situ. Eur. J. Neurosci. 2004, 20, 2235–2239. [Google Scholar] [CrossRef]
- Xu-Friedman, M.A.; Harris, K.M.; Regehr, W.G. Three-Dimensional Comparison of Ultrastructural Characteristics at Depressing and Facilitating Synapses onto Cerebellar Purkinje Cells. J. Neurosci. 2001, 21, 6666–6672. [Google Scholar] [CrossRef]
- Bernardinelli, Y.; Randall, J.; Janett, E.; Nikonenko, I.; König, S.; Jones, E.V.; Flores, C.E.; Murai, K.K.; Bochet, C.G.; Holtmaat, A.; et al. Activity-Dependent Structural Plasticity of Perisynaptic Astrocytic Domains Promotes Excitatory Synapse Stability. Curr. Biol. 2014, 24, 1679–1688. [Google Scholar] [CrossRef]
- Han, J.; Kesner, P.; Metna-Laurent, M.; Duan, T.; Xu, L.; Georges, F.; Koehl, M.; Abrous, D.N.; Mendizabal-Zubiaga, J.; Grandes, P.; et al. Acute Cannabinoids Impair Working Memory through Astroglial CB1 Receptor Modulation of Hippocampal LTD. Cell 2012, 148, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Duarte, A.; Roberts, A.J.; Ouyang, K.; Sejnowski, T.J. Impairments in Remote Memory Caused by the Lack of Type 2 IP3 Receptors. Glia 2019, 67, 1976–1989. [Google Scholar] [CrossRef]
- Sherwood, M.W.; Arizono, M.; Panatier, A.; Mikoshiba, K.; Oliet, S.H.R. Astrocytic IP3Rs: Beyond IP3R2. Front. Cell Neurosci. 2021, 15, 695817. [Google Scholar] [CrossRef] [PubMed]
- Mederos, S.; Sánchez-Puelles, C.; Esparza, J.; Valero, M.; Ponomarenko, A.; Perea, G. GABAergic Signaling to Astrocytes in the Prefrontal Cortex Sustains Goal-Directed Behaviors. Nat. Neurosci. 2021, 24, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, M.; Cuartero, M.I.; Palenzuela, R.; Draffin, J.E.; Konomi, A.; Serra, I.; Colié, S.; Castaño-Castaño, S.; Hasan, M.T.; Nebreda, Á.R.; et al. Astrocytic P38α MAPK Drives NMDA Receptor-Dependent Long-Term Depression and Modulates Long-Term Memory. Nat. Commun. 2019, 10, 2968. [Google Scholar] [CrossRef]
- Lutz, S.E.; Zhao, Y.; Gulinello, M.; Lee, S.C.; Raine, C.S.; Brosnan, C.F. Deletion of Astrocyte Connexins 43 and 30 Leads to a Dysmyelinating Phenotype and Hippocampal CA1 Vacuolation. J. Neurosci. 2009, 29, 7743–7752. [Google Scholar] [CrossRef] [PubMed]
- Stogsdill, J.A.; Ramirez, J.; Liu, D.; Kim, Y.-H.; Baldwin, K.T.; Enustun, E.; Ejikeme, T.; Ji, R.-R.; Eroglu, C. Astrocytic Neuroligins Control Astrocyte Morphogenesis and Synaptogenesis. Nature 2017, 551, 192–197. [Google Scholar] [CrossRef]
- Kol, A.; Adamsky, A.; Groysman, M.; Kreisel, T.; London, M.; Goshen, I. Astrocytes Contribute to Remote Memory Formation by Modulating Hippocampal-Cortical Communication during Learning. Nat. Neurosci. 2020, 23, 1229–1239. [Google Scholar] [CrossRef]
- Vaccari Cardoso, B.; Shevelkin, A.V.; Terrillion, C.; Mychko, O.; Mosienko, V.; Kasparov, S.; Pletnikov, M.V.; Teschemacher, A.G. Reducing L-Lactate Release from Hippocampal Astrocytes by Intracellular Oxidation Increases Novelty Induced Activity in Mice. Glia 2021, 69, 1241–1250. [Google Scholar] [CrossRef]
- Beltrán-Castillo, S.; Olivares, M.J.; Contreras, R.A.; Zúñiga, G.; Llona, I.; Von Bernhardi, R.; Eugenín, J.L. D-Serine Released by Astrocytes in Brainstem Regulates Breathing Response to CO2 Levels. Nat. Commun. 2017, 8, 838. [Google Scholar] [CrossRef]
- Bhandare, A.; van de Wiel, J.; Roberts, R.; Braren, I.; Huckstepp, R.; Dale, N. Analyzing the Brainstem Circuits for Respiratory Chemosensitivity in Freely Moving Mice. ELife 2022, 11, e70671. [Google Scholar] [CrossRef] [PubMed]
- Czeisler, C.M.; Silva, T.M.; Fair, S.R.; Liu, J.; Tupal, S.; Kaya, B.; Cowgill, A.; Mahajan, S.; Silva, P.E.; Wang, Y.; et al. The Role of PHOX2B-derived Astrocytes in Chemosensory Control of Breathing and Sleep Homeostasis. J. Physiol. 2019, 597, 2225. [Google Scholar] [CrossRef]
- Okada, Y.; Sasaki, T.; Oku, Y.; Takahashi, N.; Seki, M.; Ujita, S.; Tanaka, K.F.; Matsuki, N.; Ikegaya, Y. Preinspiratory Calcium Rise in Putative Pre-Bötzinger Complex Astrocytes. J. Physiol. 2012, 590, 4933–4944. [Google Scholar] [CrossRef] [PubMed]
- Oku, Y.; Fresemann, J.; Miwakeichi, F.; Hülsmann, S. Respiratory Calcium Fluctuations in Low-Frequency Oscillating Astrocytes in the Pre-Bötzinger Complex. Respir. Physiol. Neurobiol. 2016, 226, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Bardoni, R.; Ghirri, A.; Zonta, M.; Betelli, C.; Vitale, G.; Ruggieri, V.; Sandrini, M.; Carmignoto, G. Glutamate-Mediated Astrocyte-to-Neuron Signalling in the Rat Dorsal Horn. J. Physiol. 2010, 588, 831–846. [Google Scholar] [CrossRef]
- Broadhead, M.J.; Miles, G.B. Bi-Directional Communication Between Neurons and Astrocytes Modulates Spinal Motor Circuits. Front. Cell. Neurosci. 2020, 14, 30. [Google Scholar] [CrossRef]
- Dombeck, D.A.; Khabbaz, A.N.; Collman, F.; Adelman, T.L.; Tank, D.W. Imaging Large-Scale Neural Activity with Cellular Resolution in Awake, Mobile Mice. Neuron 2007, 56, 43–57. [Google Scholar] [CrossRef]
- Hegyi, Z.; Oláh, T.; Koszeghy, Á.; Pisticelli, F.; Holló, K.; Pál, B.; Csernoch, L.; di Marzo, V.; Antal, M. CB 1 Receptor Activation Induces Intracellular Ca2+ Mobilization and 2-Arachidonoylglycerol Release in Rodent Spinal Cord Astrocytes. Sci. Rep. 2018, 8, 10562. [Google Scholar] [CrossRef]
- Sekiguchi, K.J.; Shekhtmeyster, P.; Merten, K.; Arena, A.; Cook, D.; Hoffman, E.; Ngo, A.; Nimmerjahn, A. Imaging Large-Scale Cellular Activity in Spinal Cord of Freely Behaving Mice. Nat. Commun. 2016, 7, 11450. [Google Scholar] [CrossRef]
- Brocard, F.; Verdier, D.; Arsenault, I.; Lund, J.P.; Kolta, A. Emergence of Intrinsic Bursting in Trigeminal Sensory Neurons Parallels the Acquisition of Mastication in Weanling Rats. J. Neurophysiol. 2006, 96, 2410–2424. [Google Scholar] [CrossRef]
- Condamine, S.; Lavoie, R.; Verdier, D.; Kolta, A. Functional Rhythmogenic Domains Defined by Astrocytic Networks in the Trigeminal Main Sensory Nucleus. Glia 2018, 66, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Morquette, P.; Verdier, D.; Kadala, A.; Féthière, J.; Philippe, A.G.; Robitaille, R.; Kolta, A. An Astrocyte-Dependent Mechanism for Neuronal Rhythmogenesis. Nat. Neurosci. 2015, 18, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Damulewicz, M.; Doktór, B.; Baster, Z.; Pyza, E. The Role of Glia Clocks in the Regulation of Sleep in Drosophila Melanogaster. J. Neurosci. 2022, 42, 6848–6860. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; Edwards, M.D.; Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Cell-Autonomous Clock of Astrocytes Drives Circadian Behavior in Mammals. Science 2019, 363, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Brancaccio, M.; Patton, A.P.; Chesham, J.E.; Maywood, E.S.; Hastings, M.H. Astrocytes Control Circadian Timekeeping in the Suprachiasmatic Nucleus via Glutamatergic Signaling. Neuron 2017, 93, 1420–1435.e5. [Google Scholar] [CrossRef] [PubMed]
- Patton, A.P.; Smyllie, N.J.; Chesham, J.E.; Hastings, M.H. Astrocytes Sustain Circadian Oscillation and Bidirectionally Determine Circadian Period, But Do Not Regulate Circadian Phase in the Suprachiasmatic Nucleus. J. Neurosci. 2022, 42, 5522–5537. [Google Scholar] [CrossRef]
- Lee, H.S.; Ghetti, A.; Pinto-Duarte, A.; Wang, X.; Dziewczapolski, G.; Galimi, F.; Huitron-Resendiz, S.; Piña-Crespo, J.C.; Roberts, A.J.; Verma, I.M.; et al. Astrocytes Contribute to Gamma Oscillations and Recognition Memory. Proc. Natl. Acad. Sci. USA 2014, 111, E3343. [Google Scholar] [CrossRef]
- Hirrlinger, J.; Nimmerjahn, A. A Perspective on Astrocyte Regulation of Neural Circuit Function and Animal Behavior. Glia 2022, 70, 1554–1580. [Google Scholar] [CrossRef]
- Yu, X.; Nagai, J.; Khakh, B.S. Improved Tools to Study Astrocytes. Nat. Rev. Neurosci. 2020, 21, 121–138. [Google Scholar] [CrossRef]
- Gorzo, K.A.; Gordon, G.R. Photonics Tools Begin to Clarify Astrocyte Calcium Transients. Neurophotonics 2022, 9, 021907. [Google Scholar] [CrossRef]
- Gorina, Y.V.; Salmina, A.B.; Erofeev, A.I.; Gerasimov, E.I.; Bolshakova, A.V.; Balaban, P.M.; Bezprozvanny, I.B.; Vlasova, O.L. Astrocyte Activation Markers. Biochemistry 2022, 87, 851–870. [Google Scholar] [CrossRef] [PubMed]
- Bindocci, E.; Savtchouk, I.; Liaudet, N.; Becker, D.; Carriero, G.; Volterra, A. Three-Dimensional Ca2+ Imaging Advances Understanding of Astrocyte Biology. Science 2017, 356, eaai8185. [Google Scholar] [CrossRef] [PubMed]
- Bushong, E.A.; Martone, M.E.; Jones, Y.Z.; Ellisman, M.H. Protoplasmic Astrocytes in CA1 Stratum Radiatum Occupy Separate Anatomical Domains. J. Neurosci. 2002, 22, 183–192. [Google Scholar] [CrossRef]
- Holt, L.M.; Hernandez, R.D.; Pacheco, N.L.; Torres Ceja, B.; Hossain, M.; Olsen, M.L. Astrocyte Morphogenesis Is Dependent on BDNF Signaling via Astrocytic TrkB.T1. Elife 2019, 8, e44667. [Google Scholar] [CrossRef]
- Lanjakornsiripan, D.; Pior, B.J.; Kawaguchi, D.; Furutachi, S.; Tahara, T.; Katsuyama, Y.; Suzuki, Y.; Fukazawa, Y.; Gotoh, Y. Layer-Specific Morphological and Molecular Differences in Neocortical Astrocytes and Their Dependence on Neuronal Layers. Nat. Commun. 2018, 9, 1623. [Google Scholar] [CrossRef]
- Lu, X.H.; Yang, X.W. Genetically-Directed Sparse Neuronal Labeling in BAC Transgenic Mice through Mononucleotide Repeat Frameshift. Sci. Rep. 2017, 7, 43915. [Google Scholar] [CrossRef] [PubMed]
- Mathiisen, T.M.; Lehre, K.P.; Danbolt, N.C.; Ottersen, O.P. The Perivascular Astroglial Sheath Provides a Complete Covering of the Brain Microvessels: An Electron Microscopic 3D Reconstruction. Glia 2010, 58, 1094–1103. [Google Scholar] [CrossRef]
- Witcher, M.R.; Park, Y.D.; Lee, M.R.; Sharma, S.; Harris, K.M.; Kirov, S.A. Three-Dimensional Relationships Between Perisynaptic Astroglia and Human Hippocampal Synapses. Glia 2010, 58, 572. [Google Scholar] [CrossRef]
- Calì, C.; Agus, M.; Kare, K.; Boges, D.J.; Lehväslaiho, H.; Hadwiger, M.; Magistretti, P.J. 3D Cellular Reconstruction of Cortical Glia and Parenchymal Morphometric Analysis from Serial Block-Face Electron Microscopy of Juvenile Rat. Prog. Neurobiol. 2019, 183, 101696. [Google Scholar] [CrossRef]
- Watanabe, K.; Takeishi, H.; Hayakawa, T.; Sasaki, H. Three-Dimensional Organization of the Perivascular Glial Limiting Membrane and Its Relationship with the Vasculature: A Scanning Electron Microscope Study. Okajimas Folia Anat. Jpn. 2010, 87, 109–121. [Google Scholar] [CrossRef]
- Ventura, R.; Harris, K.M. Three-Dimensional Relationships between Hippocampal Synapses and Astrocytes. J. Neurosci. 1999, 19, 6897–6906. [Google Scholar] [CrossRef] [PubMed]
- Patrushev, I.; Gavrilov, N.; Turlapov, V.; Semyanov, A. Subcellular Location of Astrocytic Calcium Stores Favors Extrasynaptic Neuron-Astrocyte Communication. Cell Calcium 2013, 54, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Refaeli, R.; Goshen, I. Investigation of Spatial Interaction between Astrocytes and Neurons in Cleared Brains. J. Vis. Exp. 2022, 2022, e63679. [Google Scholar] [CrossRef]
- Octeau, J.C.; Chai, H.; Jiang, R.; Bonanno, S.L.; Martin, K.C.; Khakh, B.S. An Optical Neuron-Astrocyte Proximity Assay at Synaptic Distance Scales. Neuron 2018, 98, 49–66.e9. [Google Scholar] [CrossRef]
- Endo, F.; Kasai, A.; Soto, J.S.; Yu, X.; Qu, Z.; Hashimoto, H.; Gradinaru, V.; Kawaguchi, R.; Khakh, B.S. Molecular Basis of Astrocyte Diversity and Morphology across the CNS in Health and Disease. Science 2022, 378, 1631–1635. [Google Scholar] [CrossRef]
- Lavialle, M.; Aumann, G.; Anlauf, E.; Pröls, F.; Arpin, M.; Derouiche, A. Structural Plasticity of Perisynaptic Astrocyte Processes Involves Ezrin and Metabotropic Glutamate Receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 12915–12919. [Google Scholar] [CrossRef]
- Perez-Alvarez, A.; Navarrete, M.; Covelo, A.; Martin, E.D.; Araque, A. Structural and Functional Plasticity of Astrocyte Processes and Dendritic Spine Interactions. J. Neurosci. 2014, 34, 12738–12744. [Google Scholar] [CrossRef]
- Chai, H.; Diaz-Castro, B.; Shigetomi, E.; Monte, E.; Octeau, J.C.; Yu, X.; Cohn, W.; Rajendran, P.S.; Vondriska, T.M.; Whitelegge, J.P.; et al. Neural Circuit-Specialized Astrocytes: Transcriptomic, Proteomic, Morphological, and Functional Evidence. Neuron 2017, 95, 531–549.e9. [Google Scholar] [CrossRef]
- Adil, A.; Kumar, V.; Jan, A.T.; Asger, M. Single-Cell Transcriptomics: Current Methods and Challenges in Data Acquisition and Analysis. Front. Neurosci. 2021, 15, 591122. [Google Scholar] [CrossRef]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef]
- Barres, B.A.; Silverstein, B.E.; Corey, D.R.; Chun, L.L.Y. Immunological, Morphological, and Electrophysiological Variation among Retinal Ganglion Cells Purified by Panning. Neuron 1988, 1, 791–803. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sloan, S.A.; Clarke, L.E.; Caneda, C.; Plaza, C.A.; Blumenthal, P.D.; Vogel, H.; Steinberg, G.K.; Edwards, M.S.B.; Li, G.; et al. Purification and Characterization of Progenitor and Mature Human Astrocytes Reveals Transcriptional and Functional Differences with Mouse. Neuron 2016, 89, 37–53. [Google Scholar] [CrossRef] [PubMed]
- Holt, L.M.; Stoyanof, S.T.; Olsen, M.L. Magnetic Cell Sorting for In Vivo and In Vitro Astrocyte, Neuron, and Microglia Analysis. Curr. Protoc. Neurosci. 2019, 88, e71. [Google Scholar] [CrossRef]
- Holt, L.M.; Olsen, M.L. Novel Applications of Magnetic Cell Sorting to Analyze Cell-Type Specific Gene and Protein Expression in the Central Nervous System. PLoS ONE 2016, 11, e0150290. [Google Scholar] [CrossRef] [PubMed]
- Heiman, M.; Schaefer, A.; Gong, S.; Peterson, J.D.; Day, M.; Ramsey, K.E.; Suárez-Fariñas, M.; Schwarz, C.; Stephan, D.A.; Surmeier, D.J.; et al. A Translational Profiling Approach for the Molecular Characterization of CNS Cell Types. Cell 2008, 135, 738–748. [Google Scholar] [CrossRef] [PubMed]
- Doyle, J.P.; Dougherty, J.D.; Heiman, M.; Schmidt, E.F.; Stevens, T.R.; Ma, G.; Bupp, S.; Shrestha, P.; Shah, R.D.; Doughty, M.L.; et al. Application of a Translational Profiling Approach for the Comparative Analysis of CNS Cell Types. Cell 2008, 135, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Sanz, E.; Bean, J.C.; Carey, D.P.; Quintana, A.; McKnight, G.S. RiboTag: Ribosomal Tagging Strategy to Analyze Cell-Type-Specific MRNA Expression In Vivo. Curr. Protoc. Neurosci. 2019, 88, e77. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Ferrer, I.; Khakh, B.S.; Díaz-Castro, B. Cell-Specific RNA Purification to Study Translatomes of Mouse Central Nervous System. STAR Protoc. 2022, 3, 101397. [Google Scholar] [CrossRef]
- Rao, A.; Barkley, D.; França, G.S.; Yanai, I. Exploring Tissue Architecture Using Spatial Transcriptomics. Nature 2021, 596, 211. [Google Scholar] [CrossRef]
- Dries, R.; Chen, J.; Del Rossi, N.; Khan, M.M.; Sistig, A.; Yuan, G.C. Advances in Spatial Transcriptomic Data Analysis. Genome Res. 2021, 31, 1706. [Google Scholar] [CrossRef]
- Williams, C.G.; Lee, H.J.; Asatsuma, T.; Vento-Tormo, R.; Haque, A. An Introduction to Spatial Transcriptomics for Biomedical Research. Genome Med 2022, 14, 68. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, P.L.; Salmén, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and Analysis of Gene Expression in Tissue Sections by Spatial Transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex Digital Spatial Profiling of Proteins and RNA in Fixed Tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Bhatt, R.; Brown, C.; Brown, E.A.; Buhr, D.L.; Chantranuvatana, K.; Danaher, P.; Dunaway, D.; Garrison, R.G.; Geiss, G.; et al. High-Plex Multiomic Analysis in FFPE at Subcellular Level by Spatial Molecular Imaging. Nat. Biotechnol. 2022, 40, 1794–1806. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A New Generation of Ca2+ Indicators with Greatly Improved Fluorescence Properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef] [PubMed]
- Minta, A.; Kao, J.P.Y.; Tsien, R.Y. Fluorescent Indicators for Cytosolic Calcium Based on Rhodamine and Fluorescein Chromophores. J. Biol. Chem. 1989, 264, 8171–8178. [Google Scholar] [CrossRef] [PubMed]
- Nakai, J.; Ohkura, M.; Imoto, K. A High Signal-to-Noise Ca(2+) Probe Composed of a Single Green Fluorescent Protein. Nat. Biotechnol. 2001, 19, 137–141. [Google Scholar] [CrossRef]
- Redolfi, N.; Greotti, E.; Zanetti, G.; Hochepied, T.; Fasolato, C.; Pendin, D.; Pozzan, T. A New Transgenic Mouse Line for Imaging Mitochondrial Calcium Signals. Function 2021, 2, zqab012. [Google Scholar] [CrossRef]
- Srinivasan, R.; Lu, T.Y.; Chai, H.; Xu, J.; Huang, B.S.; Golshani, P.; Coppola, G.; Khakh, B.S. New Transgenic Mouse Lines for Selectively Targeting Astrocytes and Studying Calcium Signals in Astrocyte Processes In Situ and In Vivo. Neuron 2016, 92, 1181–1195. [Google Scholar] [CrossRef]
- Zhuo, L.; Theis, M.; Alvarez-Maya, I.; Brenner, M.; Willecke, K.; Messing, A. HGFAP-Cre Transgenic Mice for Manipulation of Glial and Neuronal Function In Vivo. Genesis 2001, 31, 85–94. [Google Scholar] [CrossRef]
- Vives, V.; Alonso, G.; Solal, A.C.; Joubert, D.; Legraverend, C. Visualization of S100B-Positive Neurons and Glia in the Central Nervous System of EGFP Transgenic Mice. J. Comp. Neurol. 2003, 457, 404–419. [Google Scholar] [CrossRef]
- Nolte, C.; Matyash, M.; Pivneva, T.; Schipke, C.G.; Ohlemeyer, C.; Hanisch, U.K.; Kirchhoff, F.; Kettenmann, H. GFAP Promoter-Controlled EGFP-Expressing Transgenic Mice: A Tool to Visualize Astrocytes and Astrogliosis in Living Brain Tissue. Glia 2001, 33, 72–86. [Google Scholar] [CrossRef]
- Shigetomi, E.; Bushong, E.A.; Haustein, M.D.; Tong, X.; Jackson-Weaver, O.; Kracun, S.; Xu, J.; Sofroniew, M.V.; Ellisman, M.H.; Khakh, B.S. Imaging Calcium Microdomains within Entire Astrocyte Territories and Endfeet with GCaMPs Expressed Using Adeno-Associated Viruses. J. Gen. Physiol. 2013, 141, 633–647. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, P.A.; Bland, R.J.; Mouravlev, A.; Young, D.; During, M.J. Efficient Gene Delivery and Selective Transduction of Glial Cells in the Mammalian Brain by AAV Serotypes Isolated from Nonhuman Primates. Mol. Ther. 2009, 17, 1692–1702. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; He, W.; Yang, C.; Li, J.; Jian, T.; Liang, S.; Chen, T.; Feng, H.; Chen, X.; Liao, X.; et al. Monitoring Astrocytic Ca2+ Activity in Freely Behaving Mice. Front. Cell. Neurosci. 2020, 14, 410. [Google Scholar] [CrossRef]
- Georgiou, L.; Echeverría, A.; Georgiou, A.; Kuhn, B. Ca+ Activity Maps of Astrocytes Tagged by Axoastrocytic AAV Transfer. Sci. Adv. 2022, 8, 5371. [Google Scholar] [CrossRef]
- Li, M.; Liu, Z.; Wu, Y.; Zheng, N.; Liu, X.; Cai, A.; Zheng, D.; Zhu, J.; Wu, J.; Xu, L.; et al. In Vivo Imaging of Astrocytes in the Whole Brain with Engineered AAVs and Diffusion-Weighted Magnetic Resonance Imaging. Mol. Psychiatry 2022, 1–8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Nagai, J.; Yu, X.; Papouin, T.; Cheong, E.; Freeman, M.R.; Monk, K.R.; Hastings, M.H.; Haydon, P.G.; Rowitch, D.; Shaham, S.; et al. Behaviorally Consequential Astrocytic Regulation of Neural Circuits. Neuron 2021, 109, 576–596. [Google Scholar] [CrossRef]
- Scofield, M.D.; Boger, H.A.; Smith, R.J.; Li, H.; Haydon, P.G.; Kalivas, P.W. Gq-DREADD Selectively Initiates Glial Glutamate Release and Inhibits Cue-Induced Cocaine Seeking. Biol. Psychiatry 2015, 78, 441. [Google Scholar] [CrossRef]
- Agulhon, C.; Boyt, K.M.; Xie, A.X.; Friocourt, F.; Roth, B.L.; Mccarthy, K.D. Modulation of the Autonomic Nervous System and Behaviour by Acute Glial Cell Gq Protein-Coupled Receptor Activation in Vivo. J. Physiol. 2013, 591, 5599. [Google Scholar] [CrossRef]
- Perea, G.; Yang, A.; Boyden, E.S.; Sur, M. Optogenetic Astrocyte Activation Modulates Response Selectivity of Visual Cortex Neurons In Vivo. Nat. Commun. 2014, 5, 3262. [Google Scholar] [CrossRef] [PubMed]
- Mederos, S.; Hernández-Vivanco, A.; Ramírez-Franco, J.; Martín-Fernández, M.; Navarrete, M.; Yang, A.; Boyden, E.S.; Perea, G. Melanopsin for Precise Optogenetic Activation of Astrocyte-Neuron Networks. Glia 2019, 67, 915–934. [Google Scholar] [CrossRef] [PubMed]
- Volgraf, M.; Gorostiza, P.; Numano, R.; Kramer, R.H.; Isacoff, E.Y.; Trauner, D. Allosteric Control of an Ionotropic Glutamate Receptor with an Optical Switch. Nat. Chem. Biol. 2006, 2, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Airan, R.D.; Thompson, K.R.; Fenno, L.E.; Bernstein, H.; Deisseroth, K. Temporally Precise In Vivo Control of Intracellular Signalling. Nature 2009, 458, 1025–1029. [Google Scholar] [CrossRef] [PubMed]
- Nagai, J.; Bellafard, A.; Qu, Z.; Yu, X.; Ollivier, M.; Gangwani, M.R.; Diaz-Castro, B.; Coppola, G.; Schumacher, S.M.; Golshani, P.; et al. Specific and Behaviorally Consequential Astrocyte G q GPCR Signaling Attenuation in Vivo with IβARK. Neuron 2021, 109, 2256–2274.e9. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zima, A.V.; Sheikh, F.; Blatter, L.A.; Chen, J. Endothelin-1–Induced Arrhythmogenic Ca2+ Signaling Is Abolished in Atrial Myocytes of Inositol-1,4,5-Trisphosphate(IP3)–Receptor Type 2–Deficient Mice. Circ. Res. 2005, 96, 1274–1281. [Google Scholar] [CrossRef]
- Yu, X.; Taylor, A.M.W.; Nagai, J.; Golshani, P.; Evans, C.J.; Coppola, G.; Khakh, B.S. Reducing Astrocyte Calcium Signaling In Vivo Alters Striatal Microcircuits and Causes Repetitive Behavior. Neuron 2018, 99, 1170–1187.e9. [Google Scholar] [CrossRef]
- Helmchen, F.; Denk, W. Deep Tissue Two-Photon Microscopy. Nat. Methods 2005, 2, 932–940. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.T.; Theer, P.; Denk, W. Two-Photon Imaging to a Depth of 1000 Μm in Living Brains by Use of a Ti:Al2O3 Regenerative Amplifier. Opt. Lett. 2003, 28, 1022–1024. [Google Scholar] [CrossRef]
- Centonze, V.E.; White, J.G. Multiphoton Excitation Provides Optical Sections from Deeper within Scattering Specimens than Confocal Imaging. Biophys. J 1998, 75, 2015. [Google Scholar] [CrossRef] [PubMed]
- Tonnesen, J.; Nägerl, U.V. Two-Color STED Imaging of Synapses in Living Brain Slices. Methods Mol. Biol. 2013, 950, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Ritter, J.G.; Veith, R.; Veenendaal, A.; Siebrasse, J.P.; Kubitscheck, U. Light Sheet Microscopy for Single Molecule Tracking in Living Tissue. PLoS ONE 2010, 5, e11639. [Google Scholar] [CrossRef] [PubMed]
- Keller, P.J.; Dodt, H.U. Light Sheet Microscopy of Living or Cleared Specimens. Curr. Opin. Neurobiol. 2012, 22, 138–143. [Google Scholar] [CrossRef] [PubMed]
- di Castro, M.A.; Chuquet, J.; Liaudet, N.; Bhaukaurally, K.; Santello, M.; Bouvier, D.; Tiret, P.; Volterra, A. Local Ca2+ Detection and Modulation of Synaptic Release by Astrocytes. Nat. Neurosci. 2011, 14, 1276–1284. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; DelRosso, N.V.; Vaidyanathan, T.V.; Cahill, M.K.; Reitman, M.E.; Pittolo, S.; Mi, X.; Yu, G.; Poskanzer, K.E. Accurate Quantification of Astrocyte and Neurotransmitter Fluorescence Dynamics for Single-Cell and Population-Level Physiology. Nat. Neurosci. 2019, 22, 1936–1944. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, R.; Huang, B.S.; Venugopal, S.; Johnston, A.D.; Chai, H.; Zeng, H.; Golshani, P.; Khakh, B.S. Ca2+ Signaling in Astrocytes from Ip3r2−/− Mice in Brain Slices and during Startle Responses in Vivo. Nat. Neurosci. 2015, 18, 708–717. [Google Scholar] [CrossRef]
- Agarwal, A.; Wu, P.H.; Hughes, E.G.; Fukaya, M.; Tischfield, M.A.; Langseth, A.J.; Wirtz, D.; Bergles, D.E. Transient Opening of the Mitochondrial Permeability Transition Pore Induces Microdomain Calcium Transients in Astrocyte Processes. Neuron 2017, 93, 587–605.e7. [Google Scholar] [CrossRef]
- Lines, J.; Martin, E.D.; Kofuji, P.; Aguilar, J.; Araque, A. Astrocytes Modulate Sensory-Evoked Neuronal Network Activity. Nat. Commun. 2020, 11, 3689. [Google Scholar] [CrossRef]
- Dzyubenko, E.; Prazuch, W.; Pillath-Eilers, M.; Polanska, J.; Hermann, D.M. Analysing Intercellular Communication in Astrocytic Networks Using “Astral”. Front. Cell. Neurosci. 2021, 15, 219. [Google Scholar] [CrossRef]
- Mederos, S.; González-Arias, C.; Perea, G. Melanopsin for Time-Controlling Activation of Astrocyte -Neuron Networks. Methods Mol. Biol. 2020, 2173, 53–69. [Google Scholar] [CrossRef]
- Kastanenka, K.V.; Moreno-Bote, R.; de Pittà, M.; Perea, G.; Eraso-Pichot, A.; Masgrau, R.; Poskanzer, K.E.; Galea, E. A Roadmap to Integrate Astrocytes into Systems Neuroscience. Glia 2020, 68, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Khakh, B.S. SnapShot: Astrocyte Interactions. Cell 2022, 185, 220–220.e1. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Nold, A.; Tchumatchenko, T. Modulation of Working Memory Duration by Synaptic and Astrocytic Mechanisms. PLoS Comput. Biol. 2022, 18, e1010543. [Google Scholar] [CrossRef]
- Nande, A.; Dubinkina, V.; Ravasio, R.; Zhang, G.H.; Berman, G.J. Bottlenecks, Modularity, and the Neural Control of Behavior. Front. Behav. Neurosci. 2022, 16, 835753. [Google Scholar] [CrossRef]
- Shein-Idelson, M.; Cohen, G.; Ben-Jacob, E.; Hanein, Y. Modularity Induced Gating and Delays in Neuronal Networks. PLoS Comput. Biol. 2016, 12, e1004883. [Google Scholar] [CrossRef]
- Cangelosi, A.; Invitto, S. Human-Robot Interaction and Neuroprosthetics: A Review of New Technologies. IEEE Consum. Electron. Mag. 2017, 6, 24–33. [Google Scholar] [CrossRef]
- Chang, L.; Tsao, D.Y. The Code for Facial Identity in the Primate Brain. Cell 2017, 169, 1013–1028.e14. [Google Scholar] [CrossRef]
- Kiani, R.; Cueva, C.J.; Reppas, J.B.; Newsome, W.T. Dynamics of Neural Population Responses in Prefrontal Cortex Indicate Changes of Mind on Single Trials. Curr. Biol. 2014, 24, 1542–1547. [Google Scholar] [CrossRef]
- Nogueira, R.; Abolafia, J.M.; Drugowitsch, J.; Balaguer-Ballester, E.; Sanchez-Vives, M.V.; Moreno-Bote, R. Lateral Orbitofrontal Cortex Anticipates Choices and Integrates Prior with Current Information. Nat. Commun. 2017, 8, 14823. [Google Scholar] [CrossRef]
- Kofuji, P.; Araque, A. Annual Review of Neuroscience Astrocytes and Behavior. Annu. Rev. Neurosci. 2021, 44, 49–67. [Google Scholar] [CrossRef]
- Nimmerjahn, A.; Hirrlinger, J. Astrocyte Regulation of Neural Circuit Function and Animal Behavior. Glia 2022, 70, 1453–1454. [Google Scholar] [CrossRef] [PubMed]
- Tso, C.F.; Simon, T.; Greenlaw, A.C.; Puri, T.; Mieda, M.; Herzog, E.D. Astrocytes Regulate Daily Rhythms in the Suprachiasmatic Nucleus and Behavior. Curr. Biol. 2017, 27, 1055–1061. [Google Scholar] [CrossRef] [PubMed]
- van Horn, M.R.; Benfey, N.J.; Shikany, C.; Severs, L.J.; Deemyad, T. Neuron-Astrocyte Networking: Astrocytes Orchestrate and Respond to Changes in Neuronal Network Activity across Brain States and Behaviors. J. Neurophysiol. 2021, 126, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Gordleeva, S.Y.; Tsybina, Y.A.; Krivonosov, M.I.; Ivanchenko, M.V.; Zaikin, A.A.; Kazantsev, V.B.; Gorban, A.N. Modeling Working Memory in a Spiking Neuron Network Accompanied by Astrocytes. Front. Cell. Neurosci. 2021, 15, 86. [Google Scholar] [CrossRef] [PubMed]
- de Pittà, M.; Brunel, N. Modulation of Synaptic Plasticity by Glutamatergic Gliotransmission: A Modeling Study. Neural Plast. 2016, 2016, 7607924. [Google Scholar] [CrossRef]
- Sajedinia, Z.; Hélie, S. A New Computational Model for Astrocytes and Their Role in Biologically Realistic Neural Networks. Comput. Intell. Neurosci. 2018, 2018, 3689487. [Google Scholar] [CrossRef] [PubMed]
- Mesejo, P.; Ibáñez, O.; Fernández-Blanco, E.; Cedrón, F.; Pazos, A.; Porto-Pazos, A.B. 2nd Reading Artificial Neuron-Glia Networks Learning Approach Based on Cooperative Coevolution. Int. J. Neural. Syst. 2015, 25, 1550012. [Google Scholar] [CrossRef]
- Porto-Pazos, A.B.; Veiguela, N.; Mesejo, P.; Navarrete, M.; Alvarellos, A.; Ibáñez, O.; Pazos, A.; Araque, A. Artificial Astrocytes Improve Neural Network Performance. PLoS ONE 2011, 6, e19109. [Google Scholar] [CrossRef]
- Houades, V.; Koulakoff, A.; Ezan, P.; Seif, I.; Giaume, C. Gap Junction-Mediated Astrocytic Networks in the Mouse Barrel Cortex. J. Neurosci. 2008, 28, 5207–5217. [Google Scholar] [CrossRef]
- Sul, J.Y.; Orosz, G.; Givens, R.S.; Haydon, P.G.; Haydon, P.G. Astrocytic Connectivity in the Hippocampus. Neuron Glia Biol. 2004, 1, 3. [Google Scholar] [CrossRef]
- Perea, G.; Araque, A. Properties of Synaptically Evoked Astrocyte Calcium Signal Reveal Synaptic Information Processing by Astrocytes. J. Neurosci. 2005, 25, 2192–2203. [Google Scholar] [CrossRef] [PubMed]
- Curreli, S.; Bonato, J.; Romanzi, S.; Panzeri, S.; Fellin, T. Complementary Encoding of Spatial Information in Hippocampal Astrocytes. PLoS Biol. 2022, 20, e3001530. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, K.T. Molecular Diversity of Astrocytes. Science 2022, 378, 475–476. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Sofroniew, M.V. Diversity of Astrocyte Functions and Phenotypes in Neural Circuits. Nat. Neurosci. 2015, 18, 942. [Google Scholar] [CrossRef] [PubMed]
- Khakh, B.S.; Deneen, B. The Emerging Nature of Astrocyte Diversity. Annu. Rev. Neurosci. 2019, 42, 187–207. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.H.; Raff’, M.C.; Abney, E.R.; Cohen, J.; Lindsay, R. Fibrous and Protoplasmic Astrocytes Are Biochemically and Developmentally Distinct. J. Neurosci. 1984, 4, 585–592. [Google Scholar] [CrossRef]
- Poopalasundaram, S.; Knott, C.; Shamotienko, O.G.; Foran, P.G.; Dolly, J.O.; Ghiani, C.A.; Gallo, V.; Wilkin, G.P. Glial Heterogeneity in Expression of the Inwardly Rectifying K Channel, Kir4.1, in Adult Rat CNS. Glia 2000, 30, 362–372. [Google Scholar] [CrossRef]
- Nagy, J.I.; Patel, D.; Ochalski, P.A.Y.; Stelmack, G.L. Connexin30 in Rodent, Cat and Human Brain: Selective Expression in Gray Matter Astrocytes, Co-Localization with Connexin43 at Gap Junctions and Late Developmental Appearance. Neuroscience 1999, 88, 447–468. [Google Scholar] [CrossRef]
- Goursaud, S.; Kozlova, E.N.; Maloteaux, J.M.; Hermans, E. Cultured Astrocytes Derived from Corpus Callosum or Cortical Grey Matter Show Distinct Glutamate Handling Properties. J. Neurochem. 2009, 108, 1442–1452. [Google Scholar] [CrossRef]
- Regan, M.R.; Huang, Y.H.; Yu, S.K.; Dykes-Hoberg, M.I.; Jin, L.; Watkins, A.M.; Bergles, D.E.; Rothstein, J.D. Variations in Promoter Activity Reveal a Differential Expression and Physiology of Glutamate Transporters by Glia in the Developing and Mature CNS. J. Neurosci. 2007, 27, 6607–6619. [Google Scholar] [CrossRef]
- Höft, S.; Griemsmann, S.; Seifert, G.; Steinhäuser, C. Heterogeneity in Expression of Functional Ionotropic Glutamate and GABA Receptors in Astrocytes across Brain Regions: Insights from the Thalamus. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20130602. [Google Scholar] [CrossRef] [PubMed]
- Bayraktar, O.A.; Fuentealba, L.C.; Alvarez-Buylla, A.; Rowitch, D.H. Astrocyte Development and Heterogeneity. Cold Spring Harb. Perspect. Biol. 2014, 7, a020362. [Google Scholar] [CrossRef] [PubMed]
- Chaboub, L.S.; Deneen, B. Developmental Origins of Astrocyte Heterogeneity: The Final Frontier of CNS Development. Dev. Neurosci. 2012, 34, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Kita, Y.; Kawakami, K.; Takahashi, Y.; Murakami, F. Development of Cerebellar Neurons and Glias Revealed by in Utero Electroporation: Golgi-Like Labeling of Cerebellar Neurons and Glias. PLoS ONE 2013, 8, e70091. [Google Scholar] [CrossRef]
- Oberheim, N.A.; Goldman, S.A.; Nedergaard, M. Heterogeneity of Astrocytic Form and Function. Methods Mol. Biol. 2012, 814, 23–45. [Google Scholar] [CrossRef]
- Schirmer, L.; Schafer, D.P.; Bartels, T.; Rowitch, D.H.; Calabresi, P.A. Diversity and Function of Glial Cell Types in Multiple Sclerosis. Trends Immunol. 2021, 42, 228–247. [Google Scholar] [CrossRef] [PubMed]
- Oberheim, N.A.; Takano, T.; Han, X.; He, W.; Lin, J.H.C.; Wang, F.; Xu, Q.; Wyatt, J.D.; Pilcher, W.; Ojemann, J.G.; et al. Uniquely Hominid Features of Adult Human Astrocytes. J. Neurosci. 2009, 29, 3276–3287. [Google Scholar] [CrossRef]
- Falcone, C.; Martínez-Cerdeño, V. Astrocyte Evolution and Human Specificity. Neural Regen. Res. 2023, 18, 131. [Google Scholar] [CrossRef]
- Navarrete, M.; Perea, G.; Maglio, L.; Pastor, J.; García De Sola, R.; Araque, A. Astrocyte Calcium Signal and Gliotransmission in Human Brain Tissue. Cereb. Cortex 2013, 23, 1240–1246. [Google Scholar] [CrossRef]
- Zeisel, A.; Muñoz-Manchado, A.B.; Codeluppi, S.; Lönnerberg, P.; La Manno, G.; Juréus, A.; Marques, S.; Munguba, H.; He, L.; Betsholtz, C.; et al. Cell Types in the Mouse Cortex and Hippocampus Revealed by Single-Cell RNA-Seq. Science 2015, 347, 1138–1142. [Google Scholar] [CrossRef]
- Oliveira, J.F.; Sardinha, V.M.; Guerra-Gomes, S.; Araque, A.; Sousa, N. Do Stars Govern Our Actions? Astrocyte Involvement in Rodent Behavior. Trends Neurosci. 2015, 38, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Bergles, D.E. Large-Scale Recording of Astrocyte Activity. Curr. Opin. Neurobiol. 2015, 32, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.A.; Wang, X.; Cook, D.; Carey, E.M.; Nimmerjahn, A. Imaging Spinal Cord Activity in Behaving Animals. Exp. Neurol. 2019, 320, 112974. [Google Scholar] [CrossRef] [PubMed]
- Bekar, L.K.; He, W.; Nedergaard, M. Locus Coeruleus Alpha-Adrenergic-Mediated Activation of Cortical Astrocytes In Vivo. Cereb. Cortex 2008, 18, 2789–2795. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lou, N.; Xu, Q.; Tian, G.F.; Peng, W.G.; Han, X.; Kang, J.; Takano, T.; Nedergaard, M. Astrocytic Ca2+ Signaling Evoked by Sensory Stimulation in Vivo. Nat. Neurosci. 2006, 9, 816–823. [Google Scholar] [CrossRef]
- Thrane, A.S.; Thrane, V.R.; Zeppenfeld, D.; Lou, N.; Xu, Q.; Nagelhus, E.A.; Nedergaard, M. General Anesthesia Selectively Disrupts Astrocyte Calcium Signaling in the Awake Mouse Cortex. Proc. Natl. Acad. Sci. USA 2012, 109, 18974–18979. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Mukamel, E.A.; Schnitzer, M.J. Motor Behavior Activates Bergmann Glial Networks. Neuron 2009, 62, 400–412. [Google Scholar] [CrossRef]
- Sonoda, K.; Matsui, T.; Bito, H.; Ohki, K. Astrocytes in the Mouse Visual Cortex Reliably Respond to Visual Stimulation. Biochem. Biophys. Res. Commun. 2018, 505, 1216–1222. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes Are Induced by Activated Microglia. Nature 2017, 541, 481–487. [Google Scholar] [CrossRef]
- Rothhammer, V.; Borucki, D.M.; Tjon, E.C.; Takenaka, M.C.; Chao, C.C.; Ardura-Fabregat, A.; De Lima, K.A.; Gutiérrez-Vázquez, C.; Hewson, P.; Staszewski, O.; et al. Microglial Control of Astrocytes in Response to Microbial Metabolites. Nature 2018, 557, 724–728. [Google Scholar] [CrossRef]
- Vainchtein, I.D.; Chin, G.; Cho, F.S.; Kelley, K.W.; Miller, J.G.; Chien, E.C.; Liddelow, S.A.; Nguyen, P.T.; Nakao-Inoue, H.; Dorman, L.C.; et al. Astrocyte-Derived Interleukin-33 Promotes Microglial Synapse Engulfment and Neural Circuit Development. Science 2018, 359, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Tognatta, R.; Karl, M.T.; Fyffe-Maricich, S.L.; Popratiloff, A.; Garrison, E.D.; Schenck, J.K.; Abu-Rub, M.; Miller, R.H. Astrocytes Are Required for Oligodendrocyte Survival and Maintenance of Myelin Compaction and Integrity. Front. Cell. Neurosci. 2020, 14, 74. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, X.; Yang, Y.; Fancy, S.P.J.; Yi, C.; Correspondence, J.N. Astrocyte Endfoot Formation Controls the Termination of Oligodendrocyte Precursor Cell Perivascular Migration during Development Graphical Abstract Highlights d Astrocyte Endfoot Placement Terminates Developmental OPC Perivascular Migration d Astrocyte-Derived Semaphorins 3a and 6a Act to Repel OPCs from Blood Vessels d OPC Detachment from Endothelium Permits Their Subsequent Differentiation. Neuron 2023, 111, 190–201.e8. [Google Scholar] [CrossRef] [PubMed]
- Sanmarco, L.M.; Polonio, C.M.; Wheeler, M.A.; Quintana, F.J. Functional Immune Cell-Astrocyte Interactions. J. Exp. Med. 2021, 218, e20202715. [Google Scholar] [CrossRef]
- Han, R.T.; Kim, R.D.; Molofsky, A.V.; Liddelow, S.A. Astrocyte-Immune Cell Interactions in Physiology and Pathology. Immunity 2021, 54, 211–224. [Google Scholar] [CrossRef]
- Wheeler, M.A.; Clark, I.C.; Lee, H.G.; Li, Z.; Linnerbauer, M.; Rone, J.M.; Blain, M.; Akl, C.F.; Piester, G.; Giovannoni, F.; et al. Droplet-Based Forward Genetic Screening of Astrocyte–Microglia Cross-Talk. Science 2023, 379, 1023. [Google Scholar] [CrossRef]
- Wheeler, M.A.; Jaronen, M.; Covacu, R.; Zandee, S.E.J.; Scalisi, G.; Rothhammer, V.; Tjon, E.C.; Chao, C.C.; Kenison, J.E.; Blain, M.; et al. Environmental Control of Astrocyte Pathogenic Activities in CNS Inflammation. Cell 2019, 176, 581–596.e18. [Google Scholar] [CrossRef]
- Clark, I.C.; Gutiérrez-Vázquez, C.; Wheeler, M.A.; Li, Z.; Rothhammer, V.; Linnerbauer, M.; Sanmarco, L.M.; Guo, L.; Blain, M.; Zandee, S.E.J.; et al. Barcoded Viral Tracing of Single-Cell Interactions in Central Nervous System Inflammation. Science 2021, 372, eabf1230. [Google Scholar] [CrossRef]
- Wang, L.L.; Zhang, C.L. Engineering New Neurons: In Vivo Reprogramming in Mammalian Brain and Spinal Cord. Cell Tissue Res. 2017, 371, 201–212. [Google Scholar] [CrossRef]
- Gascón, S.; Masserdotti, G.; Russo, G.L.; Götz, M. Direct Neuronal Reprogramming: Achievements, Hurdles, and New Roads to Success. Cell Stem Cell 2017, 21, 18–34. [Google Scholar] [CrossRef]
- Tai, W.; Xu, X.M.; Zhang, C.L. Regeneration through in Vivo Cell Fate Reprogramming for Neural Repair. Front. Cell. Neurosci. 2020, 14, 107. [Google Scholar] [CrossRef]
- Rivetti Di Val Cervo, P.; Romanov, R.A.; Spigolon, G.; Masini, D.; Martín-Montañez, E.; Toledo, E.M.; La Manno, G.; Feyder, M.; Pifl, C.; Ng, Y.H.; et al. Induction of Functional Dopamine Neurons from Human Astrocytes in Vitro and Mouse Astrocytes in a Parkinson’s Disease Model. Nat. Biotechnol. 2017, 35, 444–452. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Tao, X.; Sui, M.; Cui, M.; Liu, D.; Wang, B.; Wang, T.; Zheng, Y.; Luo, J.; Mu, Y.; et al. Reprogramming Astrocytes to Motor Neurons by Activation of Endogenous Ngn2 and Isl1. Stem Cell Rep. 2021, 16, 1777–1791. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Xie, H.; Du, X.; Wang, L.; Jin, X.; Zhang, Q.; Han, Y.; Sun, S.; Wang, L.; Li, X.; et al. In Vivo Chemical Reprogramming of Astrocytes into Neurons. Cell Discov. 2021, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Chen, F.; Song, N.; Xie, J. In Vivo Direct Conversion of Astrocytes to Neurons Maybe a Potential Alternative Strategy for Neurodegenerative Diseases. Front. Aging Neurosci. 2021, 13, 474. [Google Scholar] [CrossRef]



| Bibliographic Search | |
|---|---|
| Source | Scopus |
| Data accessed | 26 October 2022 |
| Query | |
| Description | (neuron) OR (astrocyte) AND (mice, OR mouse, OR human) AND (dendrite) AND (process) AND (network, OR engram) |
| Documents found | 16.199 |
| Query Filters | |
| Years Range | 1980–2023 |
| Language | English |
| Map Analysis | |
| Software | 1.6.18 VOSviewer |
| Type | Co-occurrence |
| Counting Method | Full counting |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, L.; Navarrete, M. Shining the Light on Astrocytic Ensembles. Cells 2023, 12, 1253. https://doi.org/10.3390/cells12091253
Delgado L, Navarrete M. Shining the Light on Astrocytic Ensembles. Cells. 2023; 12(9):1253. https://doi.org/10.3390/cells12091253
Chicago/Turabian StyleDelgado, Laura, and Marta Navarrete. 2023. "Shining the Light on Astrocytic Ensembles" Cells 12, no. 9: 1253. https://doi.org/10.3390/cells12091253
APA StyleDelgado, L., & Navarrete, M. (2023). Shining the Light on Astrocytic Ensembles. Cells, 12(9), 1253. https://doi.org/10.3390/cells12091253