
Epigenetic Changes Associated with Different Types of Stressors and Suicide

Abstract
:1. Introduction
2. Acute Stress
3. Chronic Stress
4. Early Childhood Stress
5. Traumatic Stress
6. Suicide
7. Transgenerational Effects
8. Future Directions/Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Berger, S.L.; Kouzarides, T.; Shiekhattar, R.; Shilatifard, A. An operational definition of epigenetics. Genes Dev. 2009, 23, 781–783. [Google Scholar] [CrossRef]
- Cheeseman, K.; Weitzman, J.B. Host-parasite interactions: An intimate epigenetic relationship. Cell Microbiol. 2015, 17, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, P.; Cartron, P.F.; Serandour, A.A.; Hervouet, E. From 1957 to Nowadays: A Brief History of Epigenetics. Int. J. Mol. Sci. 2020, 21, 7571. [Google Scholar] [CrossRef]
- Smith, J.A.; Zhao, W.; Wang, X.; Ratliff, S.M.; Mukherjee, B.; Kardia, S.L.R.; Liu, Y.; Roux, A.V.D.; Needham, B.L. Neighborhood characteristics influence DNA methylation of genes involved in stress response and inflammation: The Multi-Ethnic Study of Atherosclerosis. Epigenetics 2017, 12, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Makunin, I.V. Non-coding RNA. Hum. Mol. Genet. 2006, 15 Spec No 1, R17–R29. [Google Scholar] [CrossRef]
- Alegría-Torres, J.A.; Baccarelli, A.; Bollati, V. Epigenetics and lifestyle. Epigenomics 2011, 3, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Gatta, E.; Saudagar, V.; Auta, J.; Grayson, D.R.; Guidotti, A. Epigenetic landscape of stress surfeit disorders: Key role for DNA methylation dynamics. Int. Rev. Neurobiol. 2021, 156, 127–183. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Panahi, Y.; Sahraei, H.; Johnston, T.P.; Sahebkar, A. The impact of stress on body function: A review. Excli. J. 2017, 16, 1057–1072. [Google Scholar] [CrossRef]
- Biesmans, S.; Acton, P.D.; Cotto, C.; Langlois, X.; Ver Donck, L.; Bouwknecht, J.A.; Aelvoet, S.A.; Hellings, N.; Meert, T.F.; Nuydens, R. Effect of stress and peripheral immune activation on astrocyte activation in transgenic bioluminescent Gfap-luc mice. Glia 2015, 63, 1126–1137. [Google Scholar] [CrossRef]
- Dragoş, D.; Tănăsescu, M.D. The effect of stress on the defense systems. J. Med. Life 2010, 3, 10–18. [Google Scholar]
- Dong, T.; Zhi, L.; Bhayana, B.; Wu, M.X. Cortisol-induced immune suppression by a blockade of lymphocyte egress in traumatic brain injury. J. Neuroinflamm. 2016, 13, 197. [Google Scholar] [CrossRef]
- Howie, H.; Rijal, C.M.; Ressler, K.J. A review of epigenetic contributions to post-traumatic stress disorder. Dialogues Clin. Neurosci. 2019, 21, 417–428. [Google Scholar] [CrossRef]
- Pai, A.; Suris, A.M.; North, C.S. Posttraumatic Stress Disorder in the DSM-5: Controversy, Change, and Conceptual Considerations. Behav. Sci. 2017, 7, 7. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders: DSM-5TM, 5th ed.; American Psychiatric Publishing, a Division of American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Guo, B.; Feng, D.; Xu, Z.; Qi, P.; Yan, X. Acute benzo[a]pyrene exposure induced oxidative stress, neurotoxicity and epigenetic change in blood clam Tegillarca granosa. Sci. Rep. 2021, 11, 18744. [Google Scholar] [CrossRef]
- Apsley, A.T.; Etzel, L.; Hastings, W.J.; Heim, C.C.; Noll, J.G.; O’donnell, K.J.; Schreier, H.M.C.; Shenk, C.E.; Ye, Q.; Shalev, I. Investigating the effects of maltreatment and acute stress on the concordance of blood and DNA methylation methods of estimating immune cell proportions. Clin. Epigenetics 2023, 15, 1–13. [Google Scholar] [CrossRef]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477–501. [Google Scholar] [CrossRef]
- Li, S.; Papale, L.A.; Kintner, D.B.; Sabat, G.; Barrett-Wilt, G.A.; Cengiz, P.; Alisch, R.S. Hippocampal increase of 5-hmC in the glucocorticoid receptor gene following acute stress. Behav. Brain Res. 2015, 286, 236–240. [Google Scholar] [CrossRef]
- Li, S.; Papale, L.A.; Zhang, Q.; Madrid, A.; Chen, L.; Chopra, P.; Keleş, S.; Jin, P.; Alisch, R.S. Genome-wide alterations in hippocampal 5-hydroxymethylcytosine links plasticity genes to acute stress. Neurobiol. Dis. 2016, 86, 99–108. [Google Scholar] [CrossRef]
- de Rooij, S.R.; Costello, P.M.; Veenendaal, M.V.E.; Lillycrop, K.A.; Gluckman, P.D.; Hanson, M.A.; Painter, R.C.; Roseboom, T.J. Associations between DNA methylation of a glucocorticoid receptor promoter and acute stress responses in a large healthy adult population are largely explained by lifestyle and educational differences. Psychoneuroendocrinology 2012, 37, 782–788. [Google Scholar] [CrossRef]
- Mifsud, K.R.; Saunderson, E.A.; Spiers, H.; Carter, S.D.; Trollope, A.F.; Mill, J.; Reul, J.M. Rapid Down-Regulation of Glucocorticoid Receptor Gene Expression in the Dentate Gyrus after Acute Stress in vivo: Role of DNA Methylation and MicroRNA Activity. Neuroendocrinology 2017, 104, 157–169. [Google Scholar] [CrossRef]
- Hunter, R.G.; Murakami, G.; Dewell, S.; Seligsohn, M.; Baker, M.E.; Datson, N.A.; McEwen, B.S.; Pfaff, D.W. Acute stress and hippocampal histone H3 lysine 9 trimethylation, a retrotransposon silencing response. Proc. Natl. Acad. Sci. USA 2012, 109, 17657–17662. [Google Scholar] [CrossRef]
- Zheng, H.T.; Zhuang, Z.X.; Chen, C.J.; Liao, H.Y.; Chen, H.L.; Hsueh, H.C.; Chen, C.F.; Chen, S.E.; Huang, S.Y. Effects of acute heat stress on protein expression and histone modification in the adrenal gland of male layer-type country chickens. Sci. Rep. 2021, 11, 6499. [Google Scholar] [CrossRef]
- Hunter, R.G.; McCarthy, K.J.; Milne, T.A.; Pfaff, D.W.; McEwen, B.S. Regulation of hippocampal H3 histone methylation by acute and chronic stress. Proc. Natl. Acad. Sci. USA 2009, 106, 20912–20917. [Google Scholar] [CrossRef]
- Ieraci, A.; Mallei, A.; Musazzi, L.; Popoli, M. Physical exercise and acute restraint stress differentially modulate hippocampal brain-derived neurotrophic factor transcripts and epigenetic mechanisms in mice. Hippocampus 2015, 25, 1380–1392. [Google Scholar] [CrossRef]
- Mannironi, C.; Camon, J.; De Vito, F.; Biundo, A.; De Stefano, M.E.; Persiconi, I.; Bozzoni, I.; Fragapane, P.; Mele, A.; Presutti, C. Acute stress alters amygdala microRNA miR-135a and miR-124 expression: Inferences for corticosteroid dependent stress response. PLoS ONE 2013, 8, e73385. [Google Scholar] [CrossRef]
- Holzscheck, N.; Söhle, J.; Schläger, T.; Falckenhayn, C.; Grönniger, E.; Kolbe, L.; Wenck, H.; Terstegen, L.; Kaderali, L.; Winnefeld, M.; et al. Concomitant DNA methylation and transcriptome signatures define epidermal responses to acute solar UV radiation. Sci. Rep. 2020, 10, 12918. [Google Scholar] [CrossRef]
- Unternaehrer, E.; Luers, P.; Mill, J.; Dempster, E.; Meyer, A.H.; Staehli, S.; Lieb, R.; Hellhammer, D.H.; Meinlschmidt, G. Dynamic changes in DNA methylation of stress-associated genes (OXTR, BDNF ) after acute psychosocial stress. Transl. Psychiatry 2012, 2, e150. [Google Scholar] [CrossRef]
- Falkenberg, V.R.; Whistler, T.; Murray, J.R.; Unger, E.R.; Rajeevan, M.S. Acute psychosocial stress-mediated changes in the expression and methylation of perforin in chronic fatigue syndrome. Genet. Epigenet. 2013, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- McEwen, B.S. Neurobiological and Systemic Effects of Chronic Stress. Chronic Stress 2017, 1, 2470547017692328. [Google Scholar] [CrossRef]
- Sanacora, G.; Yan, Z.; Popoli, M. The stressed synapse 2.0: Pathophysiological mechanisms in stress-related neuropsychiatric disorders. Nat. Rev. Neurosci. 2022, 23, 86–103. [Google Scholar] [CrossRef]
- Dickens, M.; Romero, L.M.; Cyr, N.E.; Dunn, I.C.; Meddle, S.L. Chronic stress alters glucocorticoid receptor and mineralocorticoid receptor mRNA expression in the European starling (Sturnus vulgaris) brain. J. Neuroendocrinol. 2009, 21, 832–840. [Google Scholar] [CrossRef]
- Kitraki, E.; Karandrea, D.; Kittas, C. Long-lasting effects of stress on glucocorticoid receptor gene expression in the rat brain. Neuroendocrinology 1999, 69, 331–338. [Google Scholar] [CrossRef]
- Louwies, T.; Greenwood-Van Meerveld, B. Chronic stress increases DNA methylation of the GR promoter in the central nucleus of the amygdala of female rats. Neurogastroenterol. Motil. 2022, 34, e14377. [Google Scholar] [CrossRef]
- Louwies, T.; Orock, A.; Greenwood-Van Meerveld, B. Stress-induced visceral pain in female rats is associated with epigenetic remodeling in the central nucleus of the amygdala. Neurobiol. Stress 2021, 15, 100386. [Google Scholar] [CrossRef]
- Witzmann, S.R.; Turner, J.D.; Mériaux, S.B.; Meijer, O.C.; Muller, C.P. Epigenetic regulation of the glucocorticoid receptor promoter 1(7) in adult rats. Epigenetics 2012, 7, 1290–1301. [Google Scholar] [CrossRef]
- Desarnaud, F.; Jakovcevski, M.; Morellini, F.; Schachner, M. Stress downregulates hippocampal expression of the adhesion molecules NCAM and CHL1 in mice by mechanisms independent of DNA methylation of their promoters. Cell Adh. Migr. 2008, 2, 38–44. [Google Scholar] [CrossRef]
- Leung, C.S.; Kosyk, O.; Welter, E.M.; Dietrich, N.; Archer, T.K.; Zannas, A.S. Chronic stress-driven glucocorticoid receptor activation programs key cell phenotypes and functional epigenomic patterns in human fibroblasts. iScience 2022, 25, 104960. [Google Scholar] [CrossRef]
- Bartolomucci, A.; Palanza, P.; Sacerdote, P.; Panerai, A.E.; Sgoifo, A.; Dantzer, R.; Parmigiani, S. Social factors and individual vulnerability to chronic stress exposure. Neurosci. Biobehav. Rev. 2005, 29, 67–81. [Google Scholar] [CrossRef]
- Le François, B.; Soo, J.; Millar, A.M.; Daigle, M.; Le Guisquet, A.M.; Leman, S.; Minier, F.; Belzung, C.; Albert, P.R. Chronic mild stress and antidepressant treatment alter 5-HT1A receptor expression by modifying DNA methylation of a conserved Sp4 site. Neurobiol. Dis. 2015, 82, 332–341. [Google Scholar] [CrossRef]
- Wiley, J.W.; Zong, Y.; Zheng, G.; Zhu, S.; Hong, S. Histone H3K9 methylation regulates chronic stress and IL-6-induced colon epithelial permeability and visceral pain. Neurogastroenterol. Motil. 2020, 32, e13941. [Google Scholar] [CrossRef]
- Simons, R.L.; Lei, M.K.; Beach, S.R.H.; Cutrona, C.E.; Philibert, R.A. Methylation of the oxytocin receptor gene mediates the effect of adversity on negative schemas and depression. Dev. Psychopathol. 2017, 29, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Babenko, O.; Golubov, A.; Ilnytskyy, Y.; Kovalchuk, I.; Metz, G.A. Genomic and epigenomic responses to chronic stress involve miRNA-mediated programming. PLoS ONE 2012, 7, e29441. [Google Scholar] [CrossRef]
- Hing, B.; Braun, P.; Cordner, Z.A.; Ewald, E.R.; Moody, L.; McKane, M.; Willour, V.L.; Tamashiro, K.L.; Potash, J.B. Chronic social stress induces DNA methylation changes at an evolutionary conserved intergenic region in chromosome X. Epigenetics 2018, 13, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, T.; Liu, Y.Q.; Zhang, H.; Xue, S.M.; Li, G.; Cheng, H.M.; Cao, J.M. Contribution of DNA methylation in chronic stress-induced cardiac remodeling and arrhythmias in mice. FASEB J. 2019, 33, 12240–12252. [Google Scholar] [CrossRef] [PubMed]
- Niknazar, S.; Nahavandi, A.; Peyvandi, A.A.; Peyvandi, H.; Akhtari, A.S.; Karimi, M. Comparison of the Adulthood Chronic Stress Effect on Hippocampal BDNF Signaling in Male and Female Rats. Mol. Neurobiol. 2016, 53, 4026–4033. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Miyaki, K.; Suzuki, T.; Sasaki, Y.; Tsutsumi, A.; Kawakami, N.; Shimazu, A.; Takahashi, M.; Inoue, A.; Kan, C.; et al. Altered DNA methylation status of human brain derived neurotrophis factor gene could be useful as biomarker of depression. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2014, 165b, 357–364. [Google Scholar] [CrossRef]
- Wang, S.-D.; Wang, X.; Zhao, Y.; Xue, B.-H.; Wang, X.-T.; Chen, Y.-X.; Zhang, Z.-Q.; Tian, Y.-R.; Xie, F.; Qian, L.-J. Homocysteine-Induced Disturbances in DNA Methylation Contribute to Development of Stress-Associated Cognitive Decline in Rats. Neurosci. Bull. 2022, 38, 887–900. [Google Scholar] [CrossRef]
- Alasaari, J.S.; Lagus, M.; Ollila, H.M.; Toivola, A.; Kivimäki, M.; Vahtera, J.; Kronholm, E.; Härmä, M.; Puttonen, S.; Paunio, T. Environmental stress affects DNA methylation of a CpG rich promoter region of serotonin transporter gene in a nurse cohort. PLoS ONE 2012, 7, e45813. [Google Scholar] [CrossRef]
- Austin, A. Association of Adverse Childhood Experiences with Life Course Health and Development. N. C. Med. J. 2018, 79, 99–103. [Google Scholar] [CrossRef]
- Thumfart, K.M.; Jawaid, A.; Bright, K.; Flachsmann, M.; Mansuy, I.M. Epigenetics of childhood trauma: Long term sequelae and potential for treatment. Neurosci. Biobehav. Rev. 2022, 132, 1049–1066. [Google Scholar] [CrossRef]
- Miguel, P.M.; Pereira, L.O.; Silveira, P.P.; Meaney, M.J. Early environmental influences on the development of children’s brain structure and function. Dev. Med. Child Neurol. 2019, 61, 1127–1133. [Google Scholar] [CrossRef] [PubMed]
- Shields, A.E.; Wise, L.A.; Ruiz-Narvaez, E.A.; Seddighzadeh, B.; Byun, H.M.; Cozier, Y.C.; Rosenberg, L.; Palmer, J.R.; Baccarelli, A.A. Childhood abuse, promoter methylation of leukocyte NR3C1 and the potential modifying effect of emotional support. Epigenomics 2016, 8, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Tyrka, A.R.; Price, L.H.; Marsit, C.; Walters, O.C.; Carpenter, L.L. Childhood adversity and epigenetic modulation of the leukocyte glucocorticoid receptor: Preliminary findings in healthy adults. PLoS ONE 2012, 7, e30148. [Google Scholar] [CrossRef]
- Misiak, B.; Karpiński, P.; Szmida, E.; Grąźlewski, T.; Jabłoński, M.; Cyranka, K.; Rymaszewska, J.; Piotrowski, P.; Kotowicz, K.; Frydecka, D. Adverse Childhood Experiences and Methylation of the FKBP5 Gene in Patients with Psychotic Disorders. J. Clin. Med. 2020, 9, 3792. [Google Scholar] [CrossRef]
- Vangeel, E.B.; Kempke, S.; Bakusic, J.; Godderis, L.; Luyten, P.; Van Heddegem, L.; Compernolle, V.; Persoons, P.; Lambrechts, D.; Izzi, B.; et al. Glucocorticoid receptor DNA methylation and childhood trauma in chronic fatigue syndrome patients. J. Psychosom. Res. 2018, 104, 55–60. [Google Scholar] [CrossRef]
- Marzi, S.J.; Sugden, K.; Arseneault, L.; Belsky, D.W.; Burrage, J.; Corcoran, D.L.; Danese, A.; Fisher, H.L.; Hannon, E.; Moffitt, T.E.; et al. Analysis of DNA Methylation in Young People: Limited Evidence for an Association between Victimization Stress and Epigenetic Variation in Blood. Am. J. Psychiatry 2018, 175, 517–529. [Google Scholar] [CrossRef]
- Chubar, V.; Vaessen, T.; Noortgate, W.V.D.; Lutin, E.; Bosmans, G.; Bekaert, B.; Van Leeuwen, K.; Calders, F.; Weyn, S.; Bijttebier, P.; et al. Mild daily stress, in interaction with NR3C1 DNA methylation levels, is linked to alterations in the HPA axis and ANS response to acute stress in early adolescents. Psychoneuroendocrinology 2023, 150, 106045. [Google Scholar] [CrossRef]
- Mehta, D.; Klengel, T.; Conneely, K.N.; Smith, A.K.; Altmann, A.; Pace, T.W.; Rex-Haffner, M.; Loeschner, A.; Gonik, M.; Mercer, K.B.; et al. Childhood maltreatment is associated with distinct genomic and epigenetic profiles in posttraumatic stress disorder. Proc. Natl. Acad. Sci. USA 2013, 110, 8302–8307. [Google Scholar] [CrossRef]
- Ramo-Fernández, L.; Boeck, C.; Koenig, A.M.; Schury, K.; Binder, E.B.; Gündel, H.; Fegert, J.M.; Karabatsiakis, A.; Kolassa, I.T. The effects of childhood maltreatment on epigenetic regulation of stress-response associated genes: An intergenerational approach. Sci. Rep. 2019, 9, 983. [Google Scholar] [CrossRef]
- Alexander, N.; Kirschbaum, C.; Stalder, T.; Muehlhan, M.; Vogel, S. No association between FKBP5 gene methylation and acute and long-term cortisol output. Transl. Psychiatry 2020, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Roth, T.L.; Lubin, F.D.; Funk, A.J.; Sweatt, J.D. Lasting epigenetic influence of early-life adversity on the BDNF gene. Biol. Psychiatry 2009, 65, 760–769. [Google Scholar] [CrossRef]
- Wang, Q.; Shelton, R.C.; Dwivedi, Y. Interaction between early-life stress and FKBP5 gene variants in major depressive disorder and post-traumatic stress disorder: A systematic review and meta-analysis. J. Affect. Disord. 2018, 225, 422–428. [Google Scholar] [CrossRef]
- Hossack, M.R.; Reid, M.W.; Aden, J.K.; Gibbons, T.; Noe, J.C.; Willis, A.M. Adverse Childhood Experience, Genes, and PTSD Risk in Soldiers: A Methylation Study. Mil. Med. 2020, 185, 377–384. [Google Scholar] [CrossRef]
- Smearman, E.L.; Almli, L.M.; Conneely, K.N.; Brody, G.H.; Sales, J.M.; Bradley, B.; Ressler, K.J.; Smith, A.K. Oxytocin Receptor Genetic and Epigenetic Variations: Association with Child Abuse and Adult Psychiatric Symptoms. Child Dev. 2016, 87, 122–134. [Google Scholar] [CrossRef]
- Gouin, J.P.; Zhou, Q.Q.; Booij, L.; Boivin, M.; Côté, S.M.; Hébert, M.; Ouellet-Morin, I.; Szyf, M.; Tremblay, R.E.; Turecki, G.; et al. Associations among oxytocin receptor gene (OXTR) DNA methylation in adulthood, exposure to early life adversity, and childhood trajectories of anxiousness. Sci. Rep. 2017, 7, 7446. [Google Scholar] [CrossRef]
- Womersley, J.S.; Hemmings, S.M.J.; Ziegler, C.; Gutridge, A.; Ahmed-Leitao, F.; Rosenstein, D.; Domschke, K.; Seedat, S. Childhood emotional neglect and oxytocin receptor variants: Association with limbic brain volumes. World J. Biol. Psychiatry 2020, 21, 513–528. [Google Scholar] [CrossRef]
- Parianen Lesemann, F.H.; Spencer, H.; Montoya, E.R.; Kraaijenvanger, E.J.; He, Y.; Branje, S.; Boks, M.P.; Bos, P.A. Methylation of oxytocin related genes and early life trauma together shape the N170 response to human faces. Eur. Neuropsychopharmacol. 2020, 39, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Beach, S.R.H.; Brody, G.H.; Todorov, A.A.; Gunter, T.D.; Philibert, R.A. Methylation at SLC6A4 is linked to family history of child abuse: An examination of the Iowa Adoptee sample. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2010, 153b, 710–713. [Google Scholar] [CrossRef]
- Vijayendran, M.; Beach, S.R.; Plume, J.M.; Brody, G.H.; Philibert, R.A. Effects of genotype and child abuse on DNA methylation and gene expression at the serotonin transporter. Front. Psychiatry 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.J.; Kim, J.M.; Stewart, R.; Kim, S.Y.; Bae, K.Y.; Kim, S.W.; Shin, I.S.; Shin, M.G.; Yoon, J.S. Association of SLC6A4 methylation with early adversity, characteristics and outcomes in depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2013, 44, 23–28. [Google Scholar] [CrossRef]
- Booij, L.; Szyf, M.; Carballedo, A.; Frey, E.M.; Morris, D.; Dymov, S.; Vaisheva, F.; Ly, V.; Fahey, C.; Meaney, J.; et al. DNA methylation of the serotonin transporter gene in peripheral cells and stress-related changes in hippocampal volume: A study in depressed patients and healthy controls. PLoS ONE 2015, 10, e0119061. [Google Scholar] [CrossRef]
- Labonté, B.; Suderman, M.; Maussion, G.; Navaro, L.; Yerko, V.; Mahar, I.; Bureau, A.; Mechawar, N.; Szyf, M.; Meaney, M.J.; et al. Genome-wide epigenetic regulation by early-life trauma. Arch. Gen. Psychiatry 2012, 69, 722–731. [Google Scholar] [CrossRef] [PubMed]
- Lutz, P.E.; Tanti, A.; Gasecka, A.; Barnett-Burns, S.; Kim, J.J.; Zhou, Y.; Chen, G.G.; Wakid, M.; Shaw, M.; Almeida, D.; et al. Association of a History of Child Abuse with Impaired Myelination in the Anterior Cingulate Cortex: Convergent Epigenetic, Transcriptional, and Morphological Evidence. Am. J. Psychiatry 2017, 174, 1185–1194. [Google Scholar] [CrossRef]
- Houtepen, L.C.; Vinkers, C.H.; Carrillo-Roa, T.; Hiemstra, M.; van Lier, P.A.; Meeus, W.; Branje, S.; Heim, C.M.; Nemeroff, C.B.; Mill, J.; et al. Genome-wide DNA methylation levels and altered cortisol stress reactivity following childhood trauma in humans. Nat. Commun. 2016, 7, 10967. [Google Scholar] [CrossRef]
- Nätt, D.; Johansson, I.; Faresjö, T.; Ludvigsson, J.; Thorsell, A. High cortisol in 5-year-old children causes loss of DNA methylation in SINE retrotransposons: A possible role for ZNF263 in stress-related diseases. Clin. Epigenetics 2015, 7, 91. [Google Scholar] [CrossRef]
- Volk, N.; Pape, J.C.; Engel, M.; Zannas, A.S.; Cattane, N.; Cattaneo, A.; Binder, E.B.; Chen, A. Amygdalar MicroRNA-15a is Essential for Coping with Chronic Stress. Cell Rep. 2016, 17, 1882–1891. [Google Scholar] [CrossRef]
- Boersma, G.J.; Lee, R.S.; Cordner, Z.A.; Ewald, E.R.; Purcell, R.H.; Moghadam, A.A.; Tamashiro, K.L. Prenatal stress decreases Bdnf expression and increases methylation of Bdnf exon IV in rats. Epigenetics 2014, 9, 437–447. [Google Scholar] [CrossRef]
- Bockmühl, Y.; Patchev, A.V.; Madejska, A.; Hoffmann, A.; Sousa, J.C.; Sousa, N.; Holsboer, F.; Almeida, O.F.; Spengler, D. Methylation at the CpG island shore region upregulates Nr3c1 promoter activity after early-life stress. Epigenetics 2015, 10, 247–257. [Google Scholar] [CrossRef]
- Kember, R.L.; Dempster, E.L.; Lee, T.H.; Schalkwyk, L.C.; Mill, J.; Fernandes, C. Maternal separation is associated with strain-specific responses to stress and epigenetic alterations to Nr3c1, Avp, and Nr4a1 in mouse. Brain Behav. 2012, 2, 455–467. [Google Scholar] [CrossRef]
- Franklin, T.B.; Russig, H.; Weiss, I.C.; Gräff, J.; Linder, N.; Michalon, A.; Vizi, S.; Mansuy, I.M. Epigenetic transmission of the impact of early stress across generations. Biol. Psychiatry 2010, 68, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, M.C.; Adrover, E.; Pallarés, M.E.; Antonelli, M.C.; Frasch, A.C.; Brocco, M.A. Prenatal stress changes the glycoprotein GPM6A gene expression and induces epigenetic changes in rat offspring brain. Epigenetics 2014, 9, 152–160. [Google Scholar] [CrossRef]
- Fransquet, P.D.; Hjort, L.; Rushiti, F.; Wang, S.J.; Krasniqi, S.P.; Çarkaxhiu, S.I.; Arifaj, D.; Xhemaili, V.D.; Salihu, M.; Leku, N.A.; et al. DNA methylation in blood cells is associated with cortisol levels in offspring of mothers who had prenatal post-traumatic stress disorder. Stress Health 2022, 38, 755–766. [Google Scholar] [CrossRef]
- D’Addario, S.L.; Municchi, D.; Mancini, C.; Ielpo, D.; Babicola, L.; Di Segni, M.; Iacono, L.L.; Ferlazzo, F.; Cifani, C.; Andolina, D.; et al. The long-lasting effects of early life adversities are sex dependent: The signature of miR-34a. J. Affect. Disord. 2022. [Google Scholar] [CrossRef]
- Grasso, D.J.; Drury, S.; Briggs-Gowan, M.; Johnson, A.; Ford, J.; Lapidus, G.; Scranton, V.; Abreu, C.; Covault, J. Adverse childhood experiences, posttraumatic stress, and FKBP5 methylation patterns in postpartum women and their newborn infants. Psychoneuroendocrinology 2020, 114, 104604. [Google Scholar] [CrossRef] [PubMed]
- Mulder, R.H.; Walton, E.; Neumann, A.; Houtepen, L.C.; Felix, J.F.; Bakermans-Kranenburg, M.J.; Suderman, M.; Tiemeier, H.; van IJzendoorn, M.H.; Relton, C.L.; et al. Epigenomics of being bullied: Changes in DNA methylation following bullying exposure. Epigenetics 2020, 15, 750–764. [Google Scholar] [CrossRef] [PubMed]
- van der Knaap, L.J.; Riese, H.; Hudziak, J.J.; Verbiest, M.M.; Verhulst, F.C.; Oldehinkel, A.J.; van Oort, F.V. Glucocorticoid receptor gene (NR3C1) methylation following stressful events between birth and adolescence. The TRAILS study. Transl. Psychiatry 2014, 4, e381. [Google Scholar] [CrossRef]
- Papale, L.A.; Madrid, A.; Li, S.; Alisch, R.S. Early-life stress links 5-hydroxymethylcytosine to anxiety-related behaviors. Epigenetics 2017, 12, 264–276. [Google Scholar] [CrossRef]
- Silva, A.S.; Toffoli, L.V.; Estrada, V.B.; Veríssimo, L.F.; Francis-Oliveira, J.; Moreira, E.G.; Gomes, M.V.; Pelosi, G.G. Maternal exposure to fluoxetine during gestation and lactation induces long lasting changes in the DNA methylation profile of offspring’s brain and affects the social interaction of rat. Brain Res. Bull. 2018, 142, 409–413. [Google Scholar] [CrossRef]
- Kertes, D.A.; Bhatt, S.S.; Kamin, H.S.; Hughes, D.A.; Rodney, N.C.; Mulligan, C.J. BNDF methylation in mothers and newborns is associated with maternal exposure to war trauma. Clin. Epigenetics 2017, 9, 68. [Google Scholar] [CrossRef]
- Martin, C.A.; Vorn, R.; Schrieber, M.; Lai, C.; Yun, S.; Kim, H.S.; Gill, J. Identification of DNA Methylation Changes That Predict Onset of Post-traumatic Stress Disorder and Depression Following Physical Trauma. Front. Neurosci. 2021, 15, 738347. [Google Scholar] [CrossRef] [PubMed]
- Misiak, B.; Samochowiec, J.; Konopka, A.; Gawrońska-Szklarz, B.; Beszłej, J.A.; Szmida, E.; Karpiński, P. Clinical Correlates of the NR3C1 Gene Methylation at Various Stages of Psychosis. Int. J. Neuropsychopharmacol. 2021, 24, 322–332. [Google Scholar] [CrossRef]
- Misiak, B.; Szmida, E.; Karpiński, P.; Loska, O.; Sąsiadek, M.M.; Frydecka, D. Lower LINE-1 methylation in first-episode schizophrenia patients with the history of childhood trauma. Epigenomics 2015, 7, 1275–1285. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Ancelin, M.-L.; Ritchie, K.; Ryan, J. Childhood adverse events and BDNF promoter methylation in later-life. Front. Psychiatry 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, D.; Amstadter, A.B.; Pritikin, J.N.; Brick, T.R.; Neale, M.C. Associations between Traumatic Stress, Brain Volumes and Post-traumatic Stress Disorder Symptoms in Children: Data from the ABCD Study. Behav. Genet. 2022, 52, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Klinger-König, J.; Hertel, J.; Van der Auwera, S.; Frenzel, S.; Pfeiffer, L.; Waldenberger, M.; Golchert, J.; Teumer, A.; Nauck, M.; Homuth, G.; et al. Methylation of the FKBP5 gene in association with FKBP5 genotypes, childhood maltreatment and depression. Neuropsychopharmacology 2019, 44, 930–938. [Google Scholar] [CrossRef]
- Gerhart, J.I.; Canetti, D.; Hobfoll, S.E. Traumatic stress in overview: Definition, context, scope, and long-term outcomes. In Traumatic Stress and Long-Term Recovery; Springer: Berlin/Heidelberg, Germany, 2015; pp. 3–24. [Google Scholar]
- Logue, M.W.; Miller, M.W.; Wolf, E.J.; Huber, B.R.; Morrison, F.G.; Zhou, Z.; Zheng, Y.; Smith, A.K.; Daskalakis, N.P.; Ratanatharathorn, A.; et al. An epigenome-wide association study of posttraumatic stress disorder in US veterans implicates several new DNA methylation loci. Clin. Epigenetics 2020, 12, 46. [Google Scholar] [CrossRef]
- Katrinli, S.; Maihofer, A.X.; Wani, A.H.; Pfeiffer, J.R.; Ketema, E.; Ratanatharathorn, A.; Baker, D.G.; Boks, M.P.; Geuze, E.; Kessler, R.C.; et al. Epigenome-wide meta-analysis of PTSD symptom severity in three military cohorts implicates DNA methylation changes in genes involved in immune system and oxidative stress. Mol. Psychiatry 2022, 27, 1720–1728. [Google Scholar] [CrossRef]
- Pape, J.C.; Carrillo-Roa, T.; Rothbaum, B.O.; Nemeroff, C.B.; Czamara, D.; Zannas, A.S.; Iosifescu, D.; Mathew, S.J.; Neylan, T.C.; Mayberg, H.S.; et al. DNA methylation levels are associated with CRF(1) receptor antagonist treatment outcome in women with post-traumatic stress disorder. Clin. Epigenetics 2018, 10, 136. [Google Scholar] [CrossRef]
- Uddin, M.; Galea, S.; Chang, S.C.; Aiello, A.E.; Wildman, D.E.; de los Santos, R.; Koenen, K.C. Gene expression and methylation signatures of MAN2C1 are associated with PTSD. Dis. Markers 2011, 30, 111–121. [Google Scholar] [CrossRef]
- Hammamieh, R.; Chakraborty, N.; Gautam, A.; Muhie, S.; Yang, R.; Donohue, D.; Kumar, R.; Daigle, B.J., Jr.; Zhang, Y.; Amara, D.A.; et al. Whole-genome DNA methylation status associated with clinical PTSD measures of OIF/OEF veterans. Transl. Psychiatry 2017, 7, e1169. [Google Scholar] [CrossRef] [PubMed]
- Rutten, B.P.F.; Vermetten, E.; Vinkers, C.H.; Ursini, G.; Daskalakis, N.P.; Pishva, E.; de Nijs, L.; Houtepen, L.C.; Eijssen, L.; Jaffe, A.E.; et al. Longitudinal analyses of the DNA methylome in deployed military servicemen identify susceptibility loci for post-traumatic stress disorder. Mol. Psychiatry 2018, 23, 1145–1156. [Google Scholar] [CrossRef]
- Rusiecki, J.A.; Byrne, C.; Galdzicki, Z.; Srikantan, V.; Chen, L.; Poulin, M.; Yan, L.; Baccarelli, A. PTSD and DNA Methylation in Select Immune Function Gene Promoter Regions: A Repeated Measures Case-Control Study of U.S. Military Service Members. Front. Psychiatry 2013, 4, 56. [Google Scholar] [CrossRef]
- Wang, B.; Suen, C.W.; Ma, H.; Wang, Y.; Kong, L.; Qin, D.; Lee, Y.W.W.; Li, G. The Roles of H19 in Regulating Inflammation and Aging. Front. Immunol. 2020, 11, 579687. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, K.; Doggen, K.; De Keulenaer, G.W. Role of neuregulin-1/ErbB signaling in cardiovascular physiology and disease: Implications for therapy of heart failure. Circulation 2007, 116, 954–960. [Google Scholar] [CrossRef]
- Branham, E.M.; McLean, S.A.; Deliwala, I.; Mauck, M.C.; Zhao, Y.; McKibben, L.A.; Lee, A.; Spencer, A.B.; Zannas, A.S.; Lechner, M.; et al. CpG Methylation Levels in HPA Axis Genes Predict Chronic Pain Outcomes Following Trauma Exposure. J. Pain 2023. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, G.; Han, Y.; Zhang, J.; Cao, C.; Wang, L.; Zhang, K. The mechanisms of histone modification in post-traumatic stress disorder. Adv. Psychol. Sci. 2022, 30, 98. [Google Scholar] [CrossRef]
- Wolf, E.J.; Maniates, H.; Nugent, N.; Maihofer, A.X.; Armstrong, D.; Ratanatharathorn, A.; Ashley-Koch, A.E.; Garrett, M.; Kimbrel, N.A.; Lori, A.; et al. Traumatic stress and accelerated DNA methylation age: A meta-analysis. Psychoneuroendocrinology 2018, 92, 123–134. [Google Scholar] [CrossRef]
- Snijders, C.; Maihofer, A.X.; Ratanatharathorn, A.; Baker, D.G.; Boks, M.P.; Geuze, E.; Jain, S.; Kessler, R.C.; Pishva, E.; Risbrough, V.B.; et al. Longitudinal epigenome-wide association studies of three male military cohorts reveal multiple CpG sites associated with post-traumatic stress disorder. Clin. Epigenetics 2020, 12, 11. [Google Scholar] [CrossRef]
- Yehuda, R.; Flory, J.D.; Bierer, L.M.; Henn-Haase, C.; Lehrner, A.; Desarnaud, F.; Makotkine, I.; Daskalakis, N.P.; Marmar, C.R.; Meaney, M.J. Lower methylation of glucocorticoid receptor gene promoter 1F in peripheral blood of veterans with posttraumatic stress disorder. Biol. Psychiatry 2015, 77, 356–364. [Google Scholar] [CrossRef]
- Bam, M.; Yang, X.; Busbee, B.P.; Aiello, A.E.; Uddin, M.; Ginsberg, J.P.; Galea, S.; Nagarkatti, P.S.; Nagarkatti, M. Increased H3K4me3 methylation and decreased miR-7113-5p expression lead to enhanced Wnt/β-catenin signaling in immune cells from PTSD patients leading to inflammatory phenotype. Mol. Med. 2020, 26, 110. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, S.J.; Chung, H.G.; Choi, J.H.; Kim, S.H.; Kang, J.I. Epigenetic alterations of the BDNF gene in combat-related post-traumatic stress disorder. Acta Psychiatr. Scand. 2017, 135, 170–179. [Google Scholar] [CrossRef]
- Ressler, K.J.; Mercer, K.B.; Bradley, B.; Jovanovic, T.; Mahan, A.; Kerley, K.; Norrholm, S.D.; Kilaru, V.; Smith, A.K.; Myers, A.J.; et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011, 470, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Nawijn, L.; Krzyzewska, I.M.; van Zuiden, M.; Henneman, P.; Koch, S.B.J.; Mul, A.N.; Frijling, J.L.; Veltman, D.J.; Mannens, M.; Olff, M. Oxytocin receptor gene methylation in male and female PTSD patients and trauma-exposed controls. Eur. Neuropsychopharmacol. 2019, 29, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Koenen, K.C.; Uddin, M.; Chang, S.C.; Aiello, A.E.; Wildman, D.E.; Goldmann, E.; Galea, S. SLC6A4 methylation modifies the effect of the number of traumatic events on risk for posttraumatic stress disorder. Depress. Anxiety 2011, 28, 639–647. [Google Scholar] [CrossRef]
- Haghighi, F.; Xin, Y.; Chanrion, B.; O’Donnell, A.H.; Ge, Y.; Dwork, A.J.; Arango, V.; Mann, J.J. Increased DNA methylation in the suicide brain. Dialogues Clin. Neurosci. 2014, 16, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; El Hajj, N.; Müller, F.; Navarro, B.; Haaf, T. Epigenetic Dysregulation in the Prefrontal Cortex of Suicide Completers. Cytogenet. Genome Res. 2015, 146, 19–27. [Google Scholar] [CrossRef]
- Nagy, C.; Suderman, M.; Yang, J.; Szyf, M.; Mechawar, N.; Ernst, C.; Turecki, G. Astrocytic abnormalities and global DNA methylation patterns in depression and suicide. Mol. Psychiatry 2015, 20, 320–328. [Google Scholar] [CrossRef]
- Ropret, S.; Kouter, K.; Zupanc, T.; Videtic Paska, A. BDNF methylation and mRNA expression in brain and blood of completed suicides in Slovenia. World J. Psychiatry 2021, 11, 1301–1313. [Google Scholar] [CrossRef]
- Maussion, G.; Yang, J.; Suderman, M.; Diallo, A.; Nagy, C.; Arnovitz, M.; Mechawar, N.; Turecki, G. Functional DNA methylation in a transcript specific 3′UTR region of TrkB associates with suicide. Epigenetics 2014, 9, 1061–1070. [Google Scholar] [CrossRef]
- Keller, S.; Sarchiapone, M.; Zarrilli, F.; Tomaiuolo, R.; Carli, V.; Angrisano, T.; Videtic, A.; Amato, F.; Pero, R.; di Giannantonio, M.; et al. TrkB gene expression and DNA methylation state in Wernicke area does not associate with suicidal behavior. J. Affect. Disord. 2011, 135, 400–404. [Google Scholar] [CrossRef] [PubMed]
- Keller, S.; Sarchiapone, M.; Zarrilli, F.; Videtic, A.; Ferraro, A.; Carli, V.; Sacchetti, S.; Lembo, F.; Angiolillo, A.; Jovanovic, N.; et al. Increased BDNF promoter methylation in the Wernicke area of suicide subjects. Arch. Gen. Psychiatry 2010, 67, 258–267. [Google Scholar] [CrossRef]
- Jamshidi, E.; Boström, A.E.D.; Wilczek, A.; Nilsonne, Å.; Åsberg, M.; Jokinen, J. Increased Methylation of Brain-Derived Neurotrophic Factor (BDNF) Is Related to Emotionally Unstable Personality Disorder and Severity of Suicide Attempt in Women. Cells 2023, 12, 350. [Google Scholar] [CrossRef]
- Haghighi, F.; Galfalvy, H.; Echen, S.; Ehuang, Y.-Y.; Cooper, T.B.; Burke, A.K.; Oquendo, M.A.; Emann, J.J.; Esublette, M.E. DNA Methylation Perturbations in Genes Involved in Polyunsaturated Fatty Acid Biosynthesis Associated with Depression and Suicide Risk. Front. Neurol. 2015, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Gaine, M.E.; Seifuddin, F.; Sabunciyan, S.; Lee, R.S.; Benke, K.S.; Monson, E.T.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Differentially methylated regions in bipolar disorder and suicide. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2019, 180, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.M.; Crawford, B.; Dempster, E.L.; Hannon, E.; Burrage, J.; Turecki, G.; Kaminsky, Z.; Mill, J. Methylomic profiling of cortex samples from completed suicide cases implicates a role for PSORS1C3 in major depression and suicide. Transl. Psychiatry 2017, 7, e989. [Google Scholar] [CrossRef]
- Warrener, C.D.; Valentin, E.M.; Gallin, C.; Richey, L.; Ross, D.B.; Hood, C.J.; Lori, A.; Cubells, J.; Rauch, S.A.M.; Rilling, J.K. The role of oxytocin signaling in depression and suicidality in returning war veterans. Psychoneuroendocrinology 2021, 126, 105085. [Google Scholar] [CrossRef]
- Boström, A.E.D.; Jamshidi, E.; Manu, D.-M.; Kular, L.; Schiöth, H.B.; Åsberg, M.; Jokinen, J. Epigenetic changes in the CYP2D6 gene are related to severity of suicide attempt: A cross-sectional study of suicide attempters. J. Psychiatr. Res. 2023, 160, 217–224. [Google Scholar] [CrossRef]
- Kouter, K.; Zupanc, T.; Videtič Paska, A. Genome-wide DNA methylation in suicide victims revealing impact on gene expression. J. Affect. Disord. 2019, 253, 419–425. [Google Scholar] [CrossRef]
- Kouter, K.; Zupanc, T.; Videtič Paska, A. Targeted sequencing approach: Comprehensive analysis of DNA methylation and gene expression across blood and brain regions in suicide victims. World J. Biol. Psychiatry 2022, 24, 12–23. [Google Scholar] [CrossRef]
- Rizavi, H.S.; Khan, O.S.; Zhang, H.; Bhaumik, R.; Grayson, D.R.; Pandey, G.N. Methylation and expression of glucocorticoid receptor exon-1 variants and FKBP5 in teenage suicide-completers. Transl. Psychiatry 2023, 13, 1–9. [Google Scholar] [CrossRef]
- Zhu, J.-H.; Bo, H.-H.; Liu, B.-P.; Jia, C.-X. The associations between DNA methylation and depression: A systematic review and meta-analysis. J. Affect. Disord. 2023, 327, 439–450. [Google Scholar] [CrossRef]
- Policicchio, S.; Washer, S.; Viana, J.; Iatrou, A.; Burrage, J.; Hannon, E.; Turecki, G.; Kaminsky, Z.; Mill, J.; Dempster, E.L.; et al. Genome-wide DNA methylation meta-analysis in the brains of suicide completers. Transl. Psychiatry 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Matrisciano, F.; Pinna, G. PPAR-α Hypermethylation in the Hippocampus of Mice Exposed to Social Isolation Stress is Associated with Enhanced Neuroinflammation and Aggressive Behavior. Int. J. Mol. Sci. 2021, 22, 10678. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Transgenerational Epigenetic Contributions to Stress Responses: Fact or Fiction? PLoS Biol. 2016, 14, e1002426. [Google Scholar] [CrossRef]
- Skinner, M.K. Environmental stress and epigenetic transgenerational inheritance. BMC Med. 2014, 12, 153. [Google Scholar] [CrossRef] [PubMed]
- Švorcová, J. Transgenerational Epigenetic Inheritance of Traumatic Experience in Mammals. Genes 2023, 14, 120. [Google Scholar] [CrossRef]
- Mychasiuk, R.; Harker, A.; Ilnytskyy, S.; Gibb, R. Paternal stress prior to conception alters DNA methylation and behaviour of developing rat offspring. Neuroscience 2013, 241, 100–105. [Google Scholar] [CrossRef]
- Guidi, J.; Lucente, M.; Sonino, N.; Fava, G.A. Allostatic Load and Its Impact on Health: A Systematic Review. Psychother. Psychosom. 2021, 90, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Caradonna, S.G.; Paul, M.R.; Marrocco, J. An allostatic epigenetic memory on chromatin footprints after double-hit acute stress. Neurobiol. Stress 2022, 20, 100475. [Google Scholar] [CrossRef]
- Chitrala, K.N.; Hernandez, D.G.; Nalls, M.A.; Mode, N.A.; Zonderman, A.B.; Ezike, N.; Evans, M.K. Race-specific alterations in DNA methylation among middle-aged African Americans and Whites with metabolic syndrome. Epigenetics 2020, 15, 462–482. [Google Scholar] [CrossRef] [PubMed]
- Ferioli, M.; Zauli, G.; Maiorano, P.; Milani, D.; Mirandola, P.; Neri, L.M. Role of physical exercise in the regulation of epigenetic mechanisms in inflammation, cancer, neurodegenerative diseases, and aging process. J. Cell Physiol. 2019, 234, 14852–14864. [Google Scholar] [CrossRef]
- Harkess, K.N.; Ryan, J.; Delfabbro, P.H.; Cohen-Woods, S. Preliminary indications of the effect of a brief yoga intervention on markers of inflammation and DNA methylation in chronically stressed women. Transl. Psychiatry 2016, 6, e965. [Google Scholar] [CrossRef]
- Brown, K.M.; Hui, Q.; Huang, Y.; Taylor, J.Y.; Prescott, L.; de Mendoza, V.B.; Crusto, C.; Sun, Y.V. Association between Stress and Coping with DNA Methylation of Blood Pressure-Related Genes Among African American Women. Chronic Stress 2019, 3, 2470547019879088. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Daskalakis, N.P.; Desarnaud, F.; Makotkine, I.; Lehrner, A.L.; Koch, E.; Flory, J.D.; Buxbaum, J.D.; Meaney, M.J.; Bierer, L.M. Epigenetic Biomarkers as Predictors and Correlates of Symptom Improvement following Psychotherapy in Combat Veterans with PTSD. Front. Psychiatry 2013, 4, 118. [Google Scholar] [CrossRef] [PubMed]
- Wilker, S.; Vukojevic, V.; Schneider, A.; Pfeiffer, A.; Inerle, S.; Pauly, M.; Elbert, T.; Papassotiropoulos, A.; de Quervain, D.; Kolassa, I.-T. Epigenetics of traumatic stress: The association of NR3C1 methylation and posttraumatic stress disorder symptom changes in response to narrative exposure therapy. Transl. Psychiatry 2023, 13, 1–7. [Google Scholar] [CrossRef]
- Lewis, C.R.; Tafur, J.; Spencer, S.; Green, J.M.; Harrison, C.; Kelmendi, B.; Rabin, D.M.; Yehuda, R.; Yazar-Klosinski, B.; Cahn, B.R. Pilot study suggests DNA methylation of the glucocorticoid receptor gene (NR3C1) is associated with MDMA-assisted therapy treatment response for severe PTSD. Front. Psychiatry 2023, 14, 101. [Google Scholar] [CrossRef]
- Venditti, S.; Verdone, L.; Reale, A.; Vetriani, V.; Caserta, M.; Zampieri, M. Molecules of Silence: Effects of Meditation on Gene Expression and Epigenetics. Front. Psychol. 2020, 11, 1767. [Google Scholar] [CrossRef]


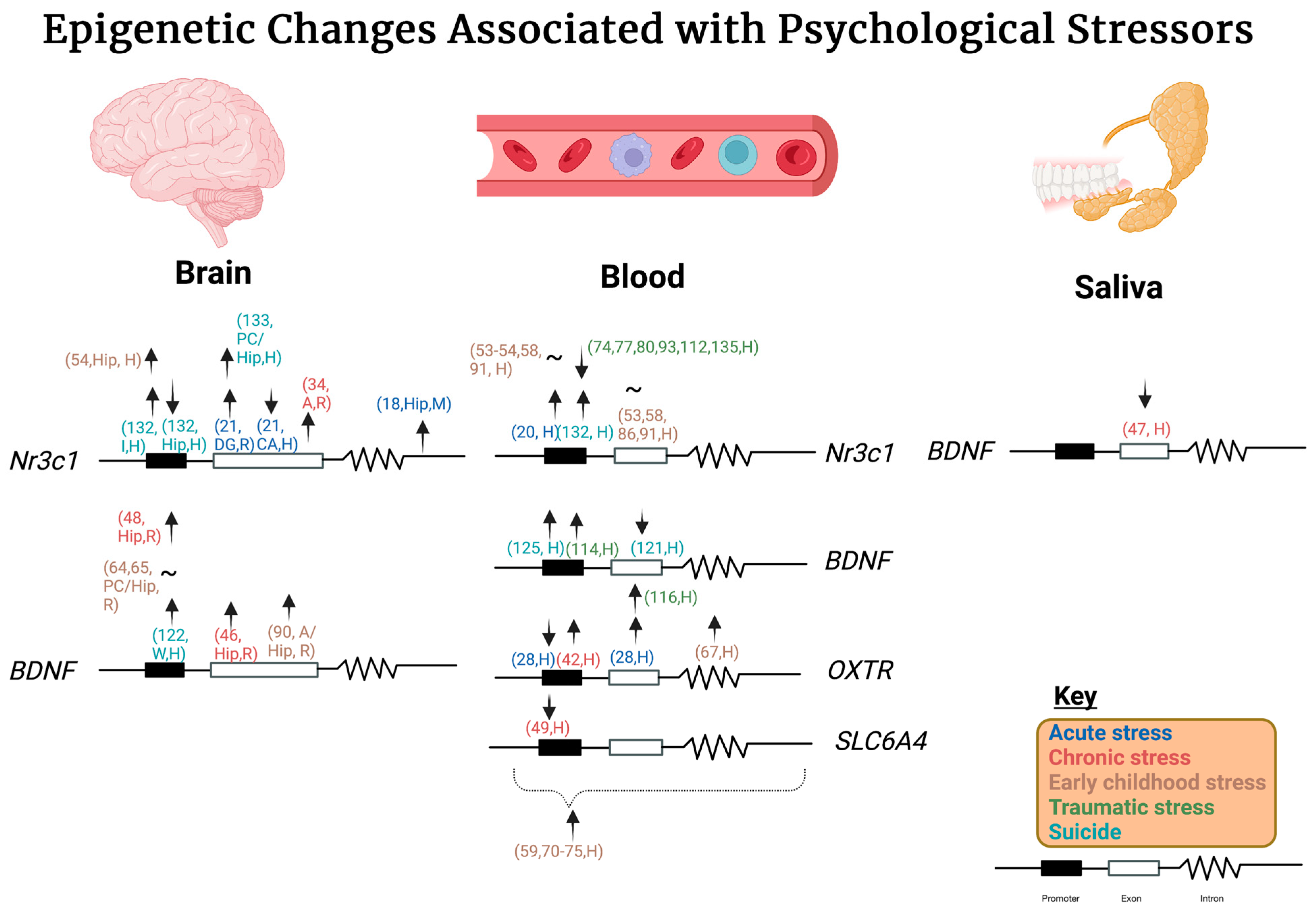{kind=link}
| Gene | Epigenetic Changes | Stress Type | Method | Model Organism | Tissue Location | |
|---|---|---|---|---|---|---|
| [16] | Whole-genome DNA methylation | Increase in methylation in concordance with increase in lymphocytes | Psychosocial stress | Illumina Infinium EPIC array | Human | Whole blood samples |
| [18] | 3′UTR Nr3c1 | Increase in 5-hmC | Acute restraint stress | Immunohistochemistry, tandem mass-spec, TAB-seq analysis | Male mice | Hippocampus |
| [20] | Promoter region 1-C of Nr3c1 | Lower levels of methylation with lower stress reactivity, but failed to survive test corrections | Psychosocial stress | Methylation-sensitive polymerase chain reaction (PCR) | Human | Fasting blood samples |
| [21] | Upstream of exon 2 in Nr3c1/Region around exon I7 Nr3c1 | No effect on CpG levels in both tissues/increase in methylation in dentate gyrus, decrease in Cornu Ammonis | Swim stress test | EZ 96-DNA methylation kit | Male Wistar rat | Dentate gyrus and Cornu Ammonis |
| [22] | Histone H3K9 at retro-transposable element loci | H3K9me3 | Acute restraint stress | ChIP sequencing against H3K9me3 | Male Sprague Dawley rats | Hippocampus |
| [23] | Histone H3K7 | H3K27me3 | Acute heat stress | Nano-UPLC-ESI-Q-TOF-MS/MS | Male Taiwan country chickens | Adrenal gland |
| [24] | Histone H3K9/H3K27 | Increase in H3K9me3 and decrease in H3K9me1 in both tissues/reduction in H3K27 me3 in both tissues | Acute restraint stress | Immunohistochemistry analysis | Adult male Sprague Dawley rats | Dentate gyrus and CA1 |
| [25] | Histone H3 in promoter region of BDNF | No methylation or acetylation change | Acute restraint stress | ChIP assay | Male mice | Hippocampus |
| [21] | MiR-124a | Increased expression | Forced swim test | MicroRNA analysis targeted against miR-124a | Male Wistar rats | Dentate gyrus |
| [26] | MiR-135a/miR-124 | Down-regulated | Acute restraint stress | Microarray analysis and reverse transcription real-time PCR | Adult male mice | Amygdala |
| [27] | CYP24A1, BRCA2, NOTCH2, FOXO3, GATA3 | Methylated | Acute exposure to UV light | Infinity methylation nEPIC array | Female Caucasian humans | Skin from lower back |
| [27] | KRT17, CSNK2A2 | Hypomethylated | Acute exposure to UV light | Infinity methylation nEPIC array | Female Caucasian humans | Skin from lower back |
| [27] | CARD14/IRF8 | Demethylated/hypermethylated | Acute exposure to UV light | Infinity methylation nEPIC array | Female Caucasian humans | Skin from lower back |
| [28] | “OXTR1”/“OXTR2”/BDNF exon V, Vh, and Vi | Increase in methylation/decrease in methylation/no methylation differences | TSST | Sequenom EpiTYPER | Human | Blood samples |
| [29] | PRF1 CpG sites -776 and -774 | Increase in methylation | TSST | Quantitative methylation analysis by Bisulfite-pyrosequencing | Chronic fatigue syndrome humans | Blood samples |
| [19] | Enpp2/Sostdc1 | Hyper-DhMR | Acute restraint stress | Genome-wide map of 5-hmC via NEBNext ChIP-Seq and RNAseq | Mice | Hippocampus |
| [19] | Ulk4/Wnt9a | Hypo-DhMR | Acute restraint stress | Genome-wide map of 5-hmC via NEBNext ChIP-Seq and RNAseq | Mice | Hippocampus |
| [19] | Banp/Gadd45b/Cbfa2t3/Irs2/Klf15/Smtn/Spns2 | Hypo-DhMR | Acute restraint stress | Genome-wide map of 5-hmC via NEBNext ChIP-Seq and RNAseq | Mice | Hippocampus |
| Gene | Epigenetic Changes | Stress Type | Method | Model Organism | Tissue Location | |
|---|---|---|---|---|---|---|
| [34] | Nr3c1 promoter region of exon 17 | Increase in DNA methylation | Chronic water avoidance stress test (7 days) | Sodium Bisulfite sequencing, pyrosequencing | Female Fischer-344 rats | Central nucleus of the amygdala |
| [36] | Nr3c1 promoter region of exon 17 | No individual site methylation change | Chronic restraint test (1 h) | Bisulfite sequencing | Male Sprague Dawley rats | Adrenal and pituitary gland |
| [37] | Nr3c1 promoter region | No increase in DNA methylation | Social defeat test by rat exposure | Sodium Bisulfite sequencing | Mice | Hippocampus |
| [37] | NCAM/CHL1 | No DNA methylation changes | Social defeat test by rat exposure | Sodium Bisulfite sequencing | Mice | Hippocampus |
| [38] | ZBTB16 | Hypermethylation | Prolonged exposure to glucocorticoids | GenFind V3 DNA extraction and purification kit, Illumina Infinium HumanMethylationEPIC BeadChip | Humans | Fetal lung fibroblast cells |
| [40] | 5-HT1A promoter (-691 CpG) | Increase in DNA methylation | Chronic unpredictable mild stress | Bisulfite treated, PCR, followed by random selection of amplifications and DNA methylation sequencing | Male mice | Prefrontal cortex and midbrain |
| [24] | Histone H3 | H3K4me3 mild increase, H3K9me3 decrease | Chronic restraint test (7 days) | Immunohistochemistry analysis | Adult male Sprague Dawley rats | Dentate gyrus |
| [41] | Histone H3 | H3K9 methylation increase, IL-6 up-regulation | Chronic water avoidance stress (10 consecutive days) | Chip, quantitative PCR, Western blot, and immunofluorescence | Young-adult male Sprague Dawley rats | Colonic |
| [42] | OXTR promoter region | Increased DNA methylation | Adult adversity (measured via Unmet Material Needs Scale and neighborhood crime) | Illumina 450K Human Methylation BeadChip | African American Women | Blood |
| [43] | MiR-709/miR-186 | Down-regulation/up-regulation | Chronic restraint test (2 weeks) | miRNA microarray analysis using µParaflo ® Biochip | Adult male Long-Evans hooded rats | Hippocampus and prefrontal cortex |
| [44] | Drosha intron 9/chromosome X intergenic region | Decrease methylation/increased methylation | Chronic social defeat (14 days) | Methyl-Seq followed by Bisulfite-pyrosequencing | Adult male mice | Dentate gyrus |
| [45] | Desmin upstream region/Tgfb1 downstream region | Increase in DNA methylation/increase in DNA methylation | Chronic restraint test (4 weeks) | Reduced representation Bisulfite sequencing and methylation-specific PCR | Male mice | Heart |
| [45] | Ppp2r2c/Ppp2r1a/Prkca/Adra1b | Alterations in DNA methylation | Chronic restraint test (4 weeks) | Reduced representation Bisulfite sequencing and methylation-specific PCR | Male mice | Heart |
| [46] | BDNF exon IV/TrkB | Increased methylation/increased methylation | Forced swim test (21 days) | One-way ANOVA, real-time PCR, and Western blotting | Male and female Wistar rats | Hippocampus |
| [47] | BDNF CpG islands in exon 1 promoter region | Lower DNA methylation | Chronic high job stress | Illumina Infinium HumanMethylation 450 BeadChip array | Japanese workers | Leukocytes in saliva |
| [48] | BDNF promoter region | DNA hypermethylation in rats with hyperhomocysteinemia | Chronic unpredicted mild stress | Methyl-Specific PCR, Sequenom Mass Spectrometry, and enzyme-linked immunosorbent assay | Rats | Hippocampus |
| [49] | SLC6A4 promoter region | Decrease in methylation | Chronic job stress | Methylation 450K BeadChip and Bisulfite sequencing | Female nurses | Peripheral blood leukocytes |
| [4] | Non-promoter CRF/shore shelf site of SLC6A4 | DNA methylation/increased methylation | Chronic stress from living in disadvantaged neighborhoods | Illumina HumanMethylation450 BeadChip | Humans | B cells, T cells, Neutrophils, and natural killer cells found in blood |
| [4] | Non-promoter F8/non-promoter TLR1 | DNA methylation | Chronic stress from living in disadvantaged neighborhoods | Illumina HumanMethylation450 BeadChip | Humans | B cells, T cells, Neutrophils, and natural killer cells found in blood |
| Gene | Epigenetic Changes | Stress Type | Method | Model Organism | Tissue Location | |
|---|---|---|---|---|---|---|
| [84] | FRMD4A, CCDC174, FBXL2, CHD4 | Methylation status associated with cortisol levels | Torture and/or sexual assault | Epigenome-wide methylation via illuminates Infinium HumanMethylationEPIC BeadChip | Human mothers | Blood |
| [82] | MeCP2, CB1, CRFR2 | Methylation of the CpG island surrounding the transcription initiation site of MeCP2 and CB1 genes was increased/CRFR2 gene methylation in a stretch of the CpG island located 5′ of the transcription initiation site was decreased | Chronic and unpredictable maternal separation from postnatal days 1–14 | Bisulfite-converted DNA followed by pyrosequencing analysis | Mice | Germline |
| [76] | KITLG (cg27512205) | Methylation | Childhood trauma | Genome-wide analysis | Healthy humans | Blood and buccal cells |
| [77] | PRDM14 | Hypomethylated | Childhood chronic stress assessed via hair cortisol | Hair cortisol levels in relationship to whole-genome DNA-methylation sequencing | 5-year-old children | Blood and hair cortisol |
| [83] | gpm6a intron 1/miRNA-133b | No overall differences in methylation, CpG 6 methylation, and CpG 7 not methylated/overexpression of miRNA suppressed gpm6a mRNA | Restraint test on mothers in last week of gestation | Bisulfite conversion and quantitative PCR | Male offspring mice (postnatal days 28 and 60) | Hippocampus and prefrontal cortex |
| [85] | miRNA-34a | Increased | Maternal separation followed by repeated cross-fostering | Quantitative real-time PCR | Female mice | Medial prefrontal cortex and dorsal raphe nuclei |
| [80] | Nr3c1 CpG island shore region | Hypomethylation | Maternal separation | Bisulfite sequencing | Male mice (postnatal day 0) | Hypothalamic neurons |
| [86] | FKBP5 | Methylation with CC gene | Adverse childhood experiences | Bisulfite pyrosequencing and DNA methylation analysis | Postpartum women and babies (within 24 h) | Saliva |
| [58] | Nr3c1, FKBP5, BDNF, AVP, CRHR1, SLC6A4 | No robust epigenetic variation | Physical, emotional, or sexual abuse/neglect/intimate-partner violence/crime/bullying/cyber-victimization | Bisulfite sequencing with Illumina Infinium HumanMethylation450 BeadChip | Human twin cohort (assessed at ages 5, 7, 10, 12, and 18) | Peripheral blood |
| [87] | RAB14 | Decrease in methylation | Child bullying | Epigenome-wide methylation, Bisulfite conversion followed with Illumina Infinium HumanMethylation450 BeadChip | Children | Blood |
| [88] | Nr3c1 amplicon 1 | Higher methylation | Perinatal stress/stressful life events/traumatic youth experiences | Bisulfite converted DNA, PCR, reverse transcription, cleavage of RNA product followed with mass spectrometry | Children (mean age 16.1) | Blood |
| [89] | Nr3c2, Nrxn1, Nfia, and Clip1 | Variable methylation in adult female mice who had experienced early life stress | Early life stress before weaning (postnatal days 12 to 18) | Chemical labeling-based 5hmC enrichment | Female mice 3-month-old | Hypothalamus |
| [79] | BDNF exon IV | Increase methylation | Prenatal stress | Bisulfite pyrosequencing genomic DNA | Male offspring of pregnant Sprague Dawley rats | Amygdala and hippocampus |
| [90] | Global DNA methylation | Increase in DNA methylation in hippocampus | Medication used for antidepressants | Imprint methylated DNA quantification Kit | Female Wistar rats | Hippocampus, cortex, hypothalamus, and periaqueductal gray matter |
| [91] | BDNF | Increase in DNA methylation with trauma exposure | War trauma | HumanMethylation450 BeadChip | Mothers and newborns in the eastern democratic republic of Congo | Umbilical cord blood, placental tissue, maternal venous blood |
| [53] | Nr3c1 | Increase in DNA methylation | Early childhood abuse | Bi-sulfite sequencing | Women from Black Women’s Health Society | leukocytes |
| [54] | Nr3c1 | Increase in promoter methylation | Early childhood abuse | Bi-sulfite sequencing | Post-mortem suicide victims of childhood abuse | Hippocampus sections post-mortem |
| [92] | Nr3c1 | Increase in DNA methylation | Early childhood physical abuse | Bi-sulfite sequencing | Subjects with Borderline Personality Disorder | Peripheral Blood Cells |
| [55] | Nr3c1 | Increase in promoter methylation | Lack of adequate nurturing, as measured by parental loss, childhood maltreatment, and parental care | Bi-sulfite sequencing | Healthy adults | Leukocytes |
| [93] | Nr3c1 | Lower Nr3c1 methylation | Schizophrenia patients with ACES | Pyrosequencing | Human | Leukocytes |
| [59] | Nr3c1 CpG sites | Increased methylation | TSST for an acute stress | Pyrosequencing | Human | Saliva |
| [60] | FKBP5 | Demethylation in functional glucocorticoid response elements of FKBP5 gene | Childhood trauma | Bisulfite pyrosequencing | Human | Whole blood cells |
| [94] | LINE1 | Lower LINE1 methylation | Schizophrenia patients with ACES | Pyrosequencing | Human | Whole blood leukocytes |
| [61] | FKBP5 | Decreased levels of DNA methylation | mother-infant dyads shortly after parturition | Mass array spectrometry | Human | Immune cells from blood |
| [56] | FKBP5 | ACES associated with lower methylation levels at CpG site | Patients with psychotic disorders (with and without ACES) | Pyrosequencing | Human | Peripheral Blood leukocytes |
| [63] | BDNF | Hypermethylation of promoter | Maltreatment compared to positive caregiving | Methylation-specific real-time PCR (MSP) or direct bisulfite DNA sequencing PCR (BSP) on bisulfite-modified DNA (Chemicon or Qiagen), or via methylated DNA immunoprecipitation using an antibody against 5-methylcytosine | Rats | Prefrontal cortex and hippocampus |
| [95] | BDNF promoter I and IV | No long-term epigenetic changes | Early life abuse, experiencing a war or natural disaster, and poverty | Sequenom MassArray | Humans | Blood and buccal tissue |
| [66] | OXTR | Hypermethylation of CpG sites predictive for pathology | 393 African American adults with or without childhood abuse | Bisulfite sequencing | Human | Whole blood |
| [67] | OXTR | ELA exposure was associated with one significant CpG site in the first intron among females, but not among males | 46 adults (23 males/23 females) with varying degrees of childhood adverse events | Pyrosequencing | Human | Whole blood |
| [70] | SLC64A | Significant effect of Child Abuse on overall methylation levels | 192 (96 males, 96 females from Iowa Adoption Study) with varying levels of childhood abuse experiences | Bisulfite conversion | Human | lymphoblasts |
| [71] | SLC64A | Examination of these four CpG residues indicated that methylation of cg22584138 was influenced by both genotype and sex abuse, whereas methylation of cg05016953 was influenced only by sex abuse history | 158 female subjects in the Iowa Adoption Studies | Illumina HumanMethylation450 BeadChip | Human | lymphoblasts |
| [72] | SLC64A | Higher promoter methylation status was significantly associated with childhood adversities | 108 patients with major depressive disorders | Bisulfite sequencing | Human | Leukocytes |
| [73] | SLC64A | Childhood trauma, being male, and smaller hippocampal volume were independently associated with greater peripheral serotonin transporter methylation | Thirty-three adults with Major Depressive Disorder (MDD) (23 females) and 36 matched healthy controls (21 females) were included in the study | Pyrosequencing | Humans | Whole Blood |
| [74] | Whole genome | 362 differentially methylated promoters in individuals with a history of abuse compared with controls. Among these promoters, 248 showed hypermethylation and 114 demonstrated hypomethylation, highest methylation difference in ALS2 gene | 41 French-Canadian men (25 with a history of severe childhood abuse and 16 control subjects) | Methylated DNA immunoprecipitation (meDIP) method | Humans | Cingulate cortex |
| [75] | Whole genome | A history of child abuse was associated with cell type-specific changes in DNA methylation of oligodendrocyte genes and a global impairment of the myelin-related transcriptional program | Postmortem brain samples from human subjects (N = 78) and from a rodent model of the impact of early life environment (N = 24) were analyzed | Bisulfite sequencing | Humans | Amygdala/Post-mortem brains |
| [60] | Whole genome | Genome-wide methylation evidence of distinct biological modifications in PTSD in the presence or absence of exposure to childhood abuse | 396 with 169 trauma-exposed individuals | HumanMethylation 450k BeadChip | Humans | Whole Blood |
| [76] | KIT4G | Nine DMRs replicated across cohorts, respectively associated with the ACE score | Two cohorts (mothers from the Avon Longitudinal Study of Parents and Children, ALSPAC, n = 780 and women from the MRC National Survey of Health and Development, NSHD, n = 552 | HumanMethylation450 BeadChip | Humans | Peripheral Blood |
| [87] | RAB14 | One site, cg17312179, showed small changes in DNA methylation associated with bullying exposure and RAB14 methylation levels decreased for exposed but increased for nonexposed | Population-based Generation R Study and Avon Longitudinal Study of Parents and Children (combined n = 1352) | Bisulfite sequencing | Humans | Peripheral Blood |
| [78] | mIR-15a | Increased levels of childhood stress | Childhood adversity | Affymetrix miRNA 2.0 array | Humans | Blood cells |
| [81] | AVP | Decrease methylation | Maternal separation | Bisulfite sequencing | Mice | hippocampus |
| [96] | Nr3c1 | CM was associated with an increase in DNA methylation in an EGR1 transcription factor binding site | Childhood maltreatment (CM) | Pyrosequencing | 147 adult participants from the Detroit Neighborhood Health Study | Whole blood |
| [97] | FKBP5 | No change in methylation for childhood abuse | Childhood abuse | Pyrosequencing | 3965 subjects of the Study of Health in Pomerania | Whole blood |
| [57] | Nr3c1 | Associations between DNA methylation and severity of fatigue as well as with childhood emotional abuse in CFS patients, although these findings were not significant after correction for multiple testing | Childhood trauma | Bisulfite sequencing | 80 female CFS (chronic fatigue syndrome) patients and 91 female controls | Peripheral blood |
| [69] | OXTR | Methylation of both OXTR and OXT genes shaped the directionality of adversity effects | Memories of Childhood Trauma | Bisulfite sequencing | 81 women | Blood |
| [65] | BDNF, Nr3c1, and MAN2C1 | Increased methylation with adverse childhood experiences | Adverse childhood experience [ACE] score | Pyrosequencing | 70 active military members with and without PTSD | Peripheral blood |
| Gene | Epigenetic Changes | Stress Type | Method | Model Organism | Tissue Location | |
|---|---|---|---|---|---|---|
| [100] | F2R, CNPY2, BAIAP2L1, TBXAS1 | All sites showed lower DNA methylation | PTSD severity | MethylationEPIC BeadChip | PTSD military personnel | Blood |
| [92] | UBE2L3 | Differentials methylated and promoter was hypomethylated in PTSD and MDD patients | Traumatic event | Bisulfite sequencing | PTSD with MDD, MDD alone, and no PTSD or MDD | Peripheral blood |
| [101] | CRHR1 | Increase in methylation from baseline in rs110402 GG allele individuals after treatment | Traumatic event | Illumina 450K array and Bisulfite conversion | Women with PTSD | Blood |
| [110] | Epigenetic age | Traumatic stress associated with advanced epigenetic age | Traumatic event | Illumina Infinium HumanMethylation BeadChip | Humans | Peripheral blood |
| [102] | MAN2C1 | Higher methylation had increased risk of PTSD | Traumatic event | Bisulfite conversion followed by Infinium HumanMethylation 27K BeadChip | Humans from Detroit | Blood |
| [104] | ZFP57, RNF39, HIST1H2APS2 | Decrease in DNA methylation with increased PTSD symptoms | Combat trauma leading to PTSD | Illumina HumanMethylation 450 BeadChip | Dutch military cohort and male US marine cohort | Blood |
| [74] | Nr3c1 promoter regions 1B and 1C | Lower methylation levels | PTSD from trauma | Sequence EpiTYPER, Sodium Bisulfite conversion | Humans with PTSD | T-lymphocyte isolated from blood |
| [99] | GOS2 cg19534438/AHRR cg05575921 | Methylated at locus/decreased methylation at locus | PTSD from trauma | EWAS using Illumina EPIC methylation BeadChip | Humans with PTSD | Blood samples and prefrontal cortex |
| [105] | H19 and IL18 | Those who did not develop PTSD had reduced %5-mC levels of H19 and IL18 after deployment | PTSD from combat trauma | Bisulfite treatment followed by pyrosequencing | Post-deployment military individuals with PTSD | Blood serum |
| [111] | HEXDC rs4789774/MAD1L1 | Development of combat-related PTSD is associated with distinct methylation patterns in HLA region, HEXDC, and MAD1L1 | PTSD from combat trauma | HumanMethylation450 BeadChip | Three cohorts of male military members | Blood |
| [91] | CRH, CRHBP, Nr3c1, and FKBP5 | Methylation changes in offspring associated with war exposure in mothers | War trauma | HumanMethylation450 BeadChip | Mothers and newborns in the eastern Democratic Republic of Congo | Umbilical cord blood, placental tissue, maternal venous blood |
| [4] | AHRR | Lower DNA methylation in PTSD | Traumatic event | Illumina HumanMethylation450 BeadChip | Military and civilian cohorts | Blood |
| [60] | SENP7 | Six genome-wide significant (GWS) CpG sites associated with past-month PTSD and three CpGs with lifetime PTSD | PTSD from the Vietnam War | Bisulfite converted analysis | 1135 male European–American U.S. veterans who participated in the National Health and Resilience in Veterans Study (NHRVS) | Semen and blood |
| [112] | Nr3c1 1F promoter | Lower methylation in veterans with PTSD | PTSD from combat trauma | Bisulfite mapping and colonial sequencing | Male combat veterans | Peripheral blood mononuclear cells |
| [113] | H3K4me3 histone modification, WNT 10B | Increase in H3K4me3 around WNT 10B promoter in patients with PTSD | PTSD from various traumatic stressors | RNA-seq, ChIP-seq, and microarray | Patients with PTSD | Peripheral blood mononuclear cells |
| [102] | NRG1, HGS | Two CpG sites significantly associated with current PTSD in NRG1 (cg23637605) and in HGS (cg19577098) | PTSD from combat trauma | Methylation microarray | Patients with PTSD | Whole blood cells |
| [103] | 5600 CpG islands | Majority of CpG islands were hypermethylated in PTSD cases | PTSD from combat trauma | Agilent whole genome methylation array/targeted bisulfite sequencing | Operation Enduring Freedom/Iraqi Freedom Combat veterans | Peripheral whole blood cells |
| [114] | BDNF | Subjects with PTSD showed a higher DNA methylation of four CpG sites at the BDNF promoter compared with those without PTSD | Combat veterans with PTSD | Pyrosequencing | Combat veterans | Peripheral blood cells |
| [115] | ADCYAP1 | Methylation of ADCYAP1R1 is associated with PTSD | Patients with PTSD | HumanMethylation27 BeadChip | Patients receiving services in the primary care clinics at Grady Memorial Hospital | Whole blood |
| [116] | OXTR exon 3 | CpG islands increase in methylation in female PTSD | PTSD | Bisulfite converted analysis | 67 human subjects (31 PTSD, 36 controls) | Whole blood |
| [117] | SLC6A4 promoter | No association, increase in methylation demonstrated protective effects | PTSD | HM27 BeadChip | 100 human subjects | Whole blood |
| [108] | POMC and CRHBP | CpG sites demonstrated increased methylation as predictor for chronic post-traumatic musculoskeletal pain | PTSD | Bisulfite conversion and Illumina Infinium Human MethylationEPIC BeadChip array | Humans | Blood |
| Gene | Epigenetic Changes | Stress Type | Method | Model Organism | Tissue Location | |
|---|---|---|---|---|---|---|
| [120] | GRIK2 intron 13/BEGAIN | Hypomethylated/increase in methylation | Suicide completion w/MDD | Genome-wide methylation sequencing via MBD-Seq | Humans | Cortical brain region |
| [121] | BDNF exon 1 | Decrease in methylation | Suicide completion via hanging | Bisulfite next-generation sequencing | Humans | Blood |
| [122] | 3′UTR TrkB-T1 | Hypermethylation | Suicide completion with low TrkB-T1 | Methylated DNA immunoprecipitation, labeling, and hybridization via microarray | Humans | BA 8 and 9 |
| [123] | BDNF promoter region IV | Hypermethylation | Suicide completion | MassArray methylation analysis | Humans | Wernicke area |
| [125] | BDNF promoter | Increased methylation | Suicidal patients with emotionally unstable personality disorder | Illumine EPIC BeadChip | Humans (women) | Blood |
| [126] | Elovl5 upstream/downstream to transcription start site | Methylation/lower CpG methylation | Suicide non-completers with MDD | Bisulfite pyrosequencing | Humans | Blood |
| [127] | ARHGEF38 | Hypermethylation across 4 CpG sites | Bipolar disorder suicide completion | SureSelect(XT) system, methyl-Seq, confirmation via pyrosequencing | Humans | BA 46 |
| [128] | PSORS1C3 | Hypomethylation | Suicide completion w/MDD | Bisulfite conversion followed by Infinium HumanMethylation450 BeadChip Array, then Bisulfite pyrosequencing confirmation | Humans | BA 11 and 25 |
| [129] | OXTR MT2 region (-901, -924, -934) | No methylation at sites | Suicidality | Methylation assay via Bisulfite conversion followed by pyrosequencing | Humans (male veterans) | Saliva |
| [130] | CYP2D6 CpG sites | Hypomethylation (males) and hypermethylation (females) | Severe suicide behavior | Illumina Infinium Methylation EPIC BeadChip | Humans | Peripheral blood |
| [131] | ZNF714/NRIP3 | Hypomethylated/mixed methylation | Suicide completion via hanging | Next-generation sequencing of genome-wide methylation analysis | Humans | BA 9 |
| [132] | Nr3c1 1B promoter | Higher methylation in the insula and blood, lower levels of methylation in hippocampus | Suicide completion | Bisulfite next-generation sequencing | Male Humans | Insula, blood, and hippocampus |
| [133] | Nr3c1 5′UTR | DNA methylation | Suicide completion | DNA methylation enrichment assay | Teenage humans | Hippocampus and prefrontal cortex |
| [132] | SLC6A4_2 amplicon | Hypermethylated | Suicide completion | Bisulfite next-generation sequencing | Male Humans | BA 46 |
| [132] | 5-HT1A | Hypomethylated in blood/hypermethylated in insula | Suicide completion | Bisulfite next-generation sequencing | Male Humans | Blood and insula |
| [132] | SKA2 | Mixed methylation data in all tissues | Suicide completion | Bisulfite next-generation sequencing | Male Humans | Hippocampus, insula, amygdala, BA 46, and blood |
| [132] | MAOA_2 amplicon | Hypomethylated in insula and BA 46 | Suicide completion | Bisulfite next-generation sequencing | Male Humans | Insula and BA 46 |
| [132] | GABRA1 | Decreased methylation in hippocampus and blood/increased methylation in insula | Suicide completion | Bisulfite next-generation sequencing | Male Humans | Hippocampus, blood, insula |
| [132] | NRIP3 amplicon | Decrease methylation | Suicide completion | Bisulfite next-generation sequencing | Male Humans | Hippocampus and insula |
| [135] | CERC2 intronic region | Hypermethylation across 4 CpG sites | Suicide completers | Illumina HumanMethylation450K BeadChip or Infinium MethylationEPIC BeadChip | Human | Cerebellum |
| [136] | PPAR-α | Increased methylation in CpG regions | Social isolation (4 weeks) model for suicide-like behavior/PTSD | Methyl-DNA immunoprecipitation | Male Swiss-webster mice | Hippocampus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dee, G.; Ryznar, R.; Dee, C. Epigenetic Changes Associated with Different Types of Stressors and Suicide. Cells 2023, 12, 1258. https://doi.org/10.3390/cells12091258
Dee G, Ryznar R, Dee C. Epigenetic Changes Associated with Different Types of Stressors and Suicide. Cells. 2023; 12(9):1258. https://doi.org/10.3390/cells12091258
Chicago/Turabian StyleDee, Garrett, Rebecca Ryznar, and Colton Dee. 2023. "Epigenetic Changes Associated with Different Types of Stressors and Suicide" Cells 12, no. 9: 1258. https://doi.org/10.3390/cells12091258
APA StyleDee, G., Ryznar, R., & Dee, C. (2023). Epigenetic Changes Associated with Different Types of Stressors and Suicide. Cells, 12(9), 1258. https://doi.org/10.3390/cells12091258