
Poly-L-Lactic Acid Fillers Improved Dermal Collagen Synthesis by Modulating M2 Macrophage Polarization in Aged Animal Skin

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. PLLA Preparation
2.1.1. PLLA Synthesis
2.1.2. Scanning Electron Microscopy (SEM) Imaging for Surface Observations of PLLA
2.2. Experimental Models
2.2.1. PLLA Treatment in H2O2-Induced Senescent Macrophages
2.2.2. PLLA Treatment of H2O2-Induced Senescent Fibroblasts
2.2.3. Treatment of Senescent Fibroblasts with CM from PLLA-Treated Senescent Macrophages
2.2.4. PLLA Treatment in Young and Aged Mice
2.3. Senescence-Associated β-Galactosidase (SA-β-gal) Activity
2.4. Western Blotting
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Immunocytochemistry
2.7. Staining with 3,3-Diaminobenzidine (DAB)
2.8. Proliferation Assay
2.9. Immunofluorescence
2.10. Histological Analysis
2.10.1. Masson’s Trichrome Staining
2.10.2. Herovici’s Staining
2.10.3. Verhoeff’s Staining
2.11. Statistical Analysis
3. Results
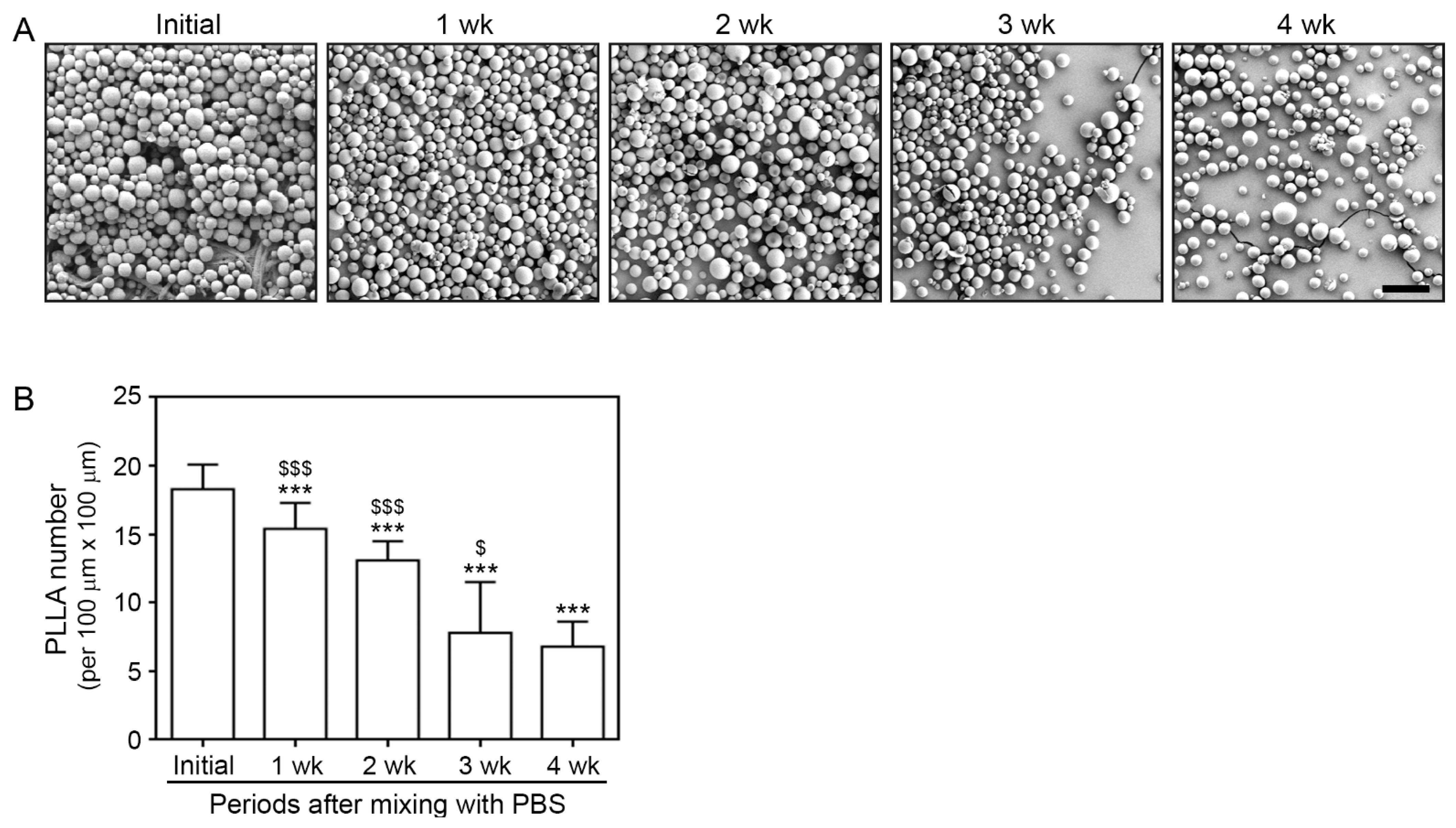
3.1. Morphology and Degradation Pattern of PLLA
3.2. PLLA Increased IL-4 and IL-13 Expression and M2 Polrization in H2O2-Treated Macrophages and Aged Skin
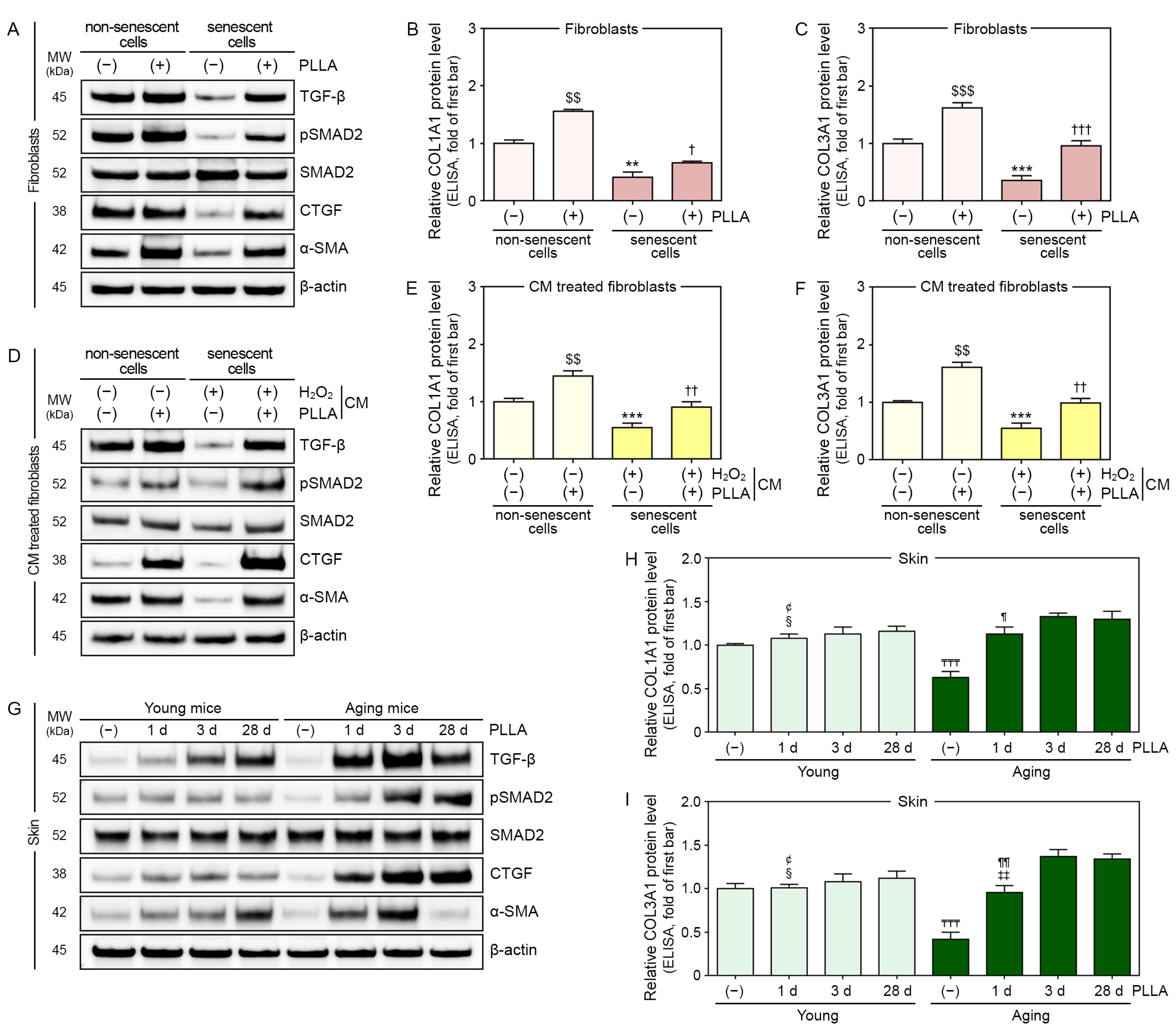
3.3. PLLA Upregulated Expression of TGF-β, pSMAD2/SMAD2, and Connective Tissue Growth Factor (CTGF) in H2O2-Treated Fibroblasts
3.4. PLLA Upregulated the Expression of TGF-β, pSMAD2/SMAD2, and CTGF in Fibroblasts Treated with CM from H2O2-Treated Macrophages and Aged Skin
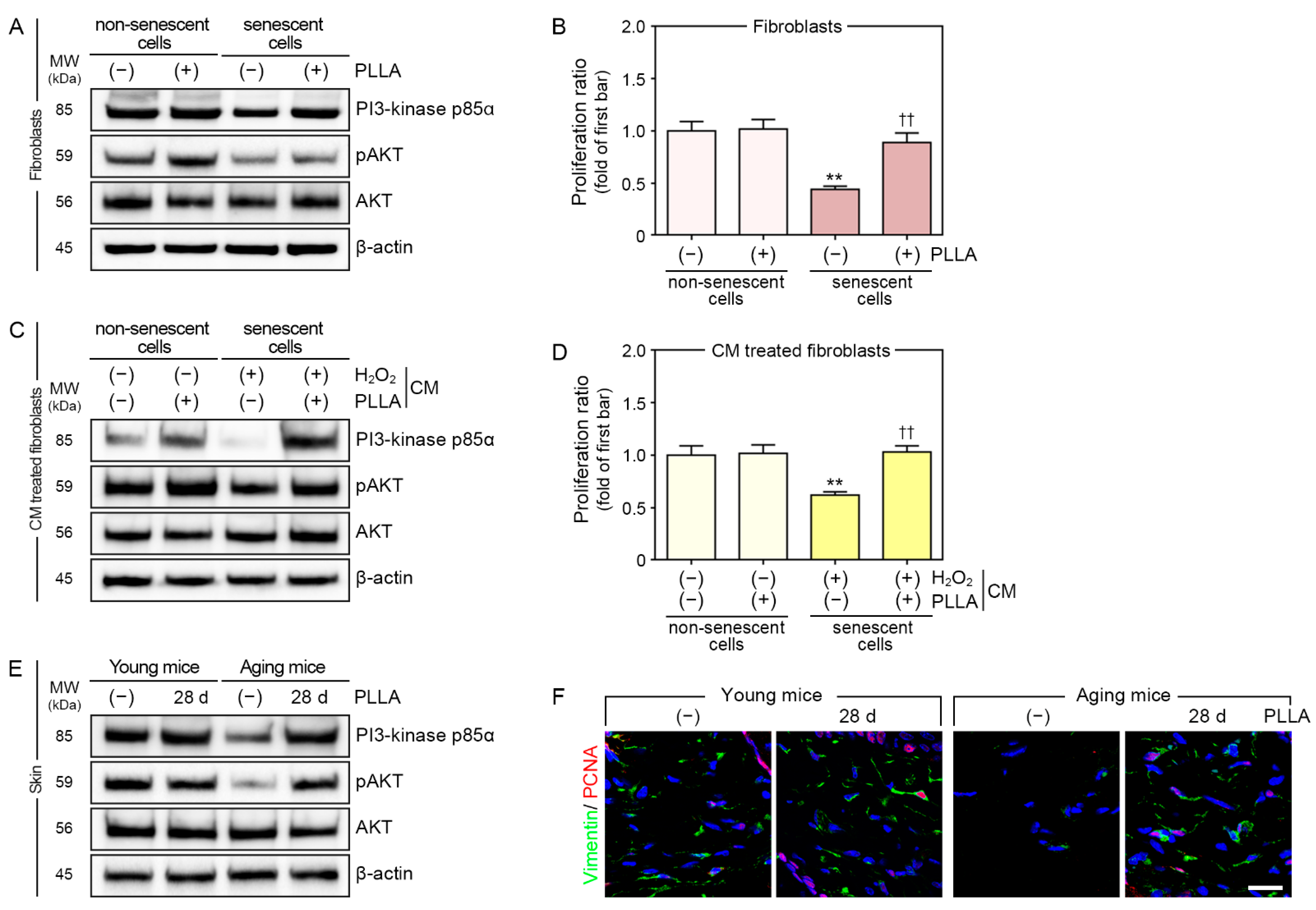
3.5. PLLA Upregulated Phosphatidylinositol 3-Kinase p85α (PI3-Kinase p85α)/Protein Kinase B (AKT) Signaling and Fibroblast Proliferation
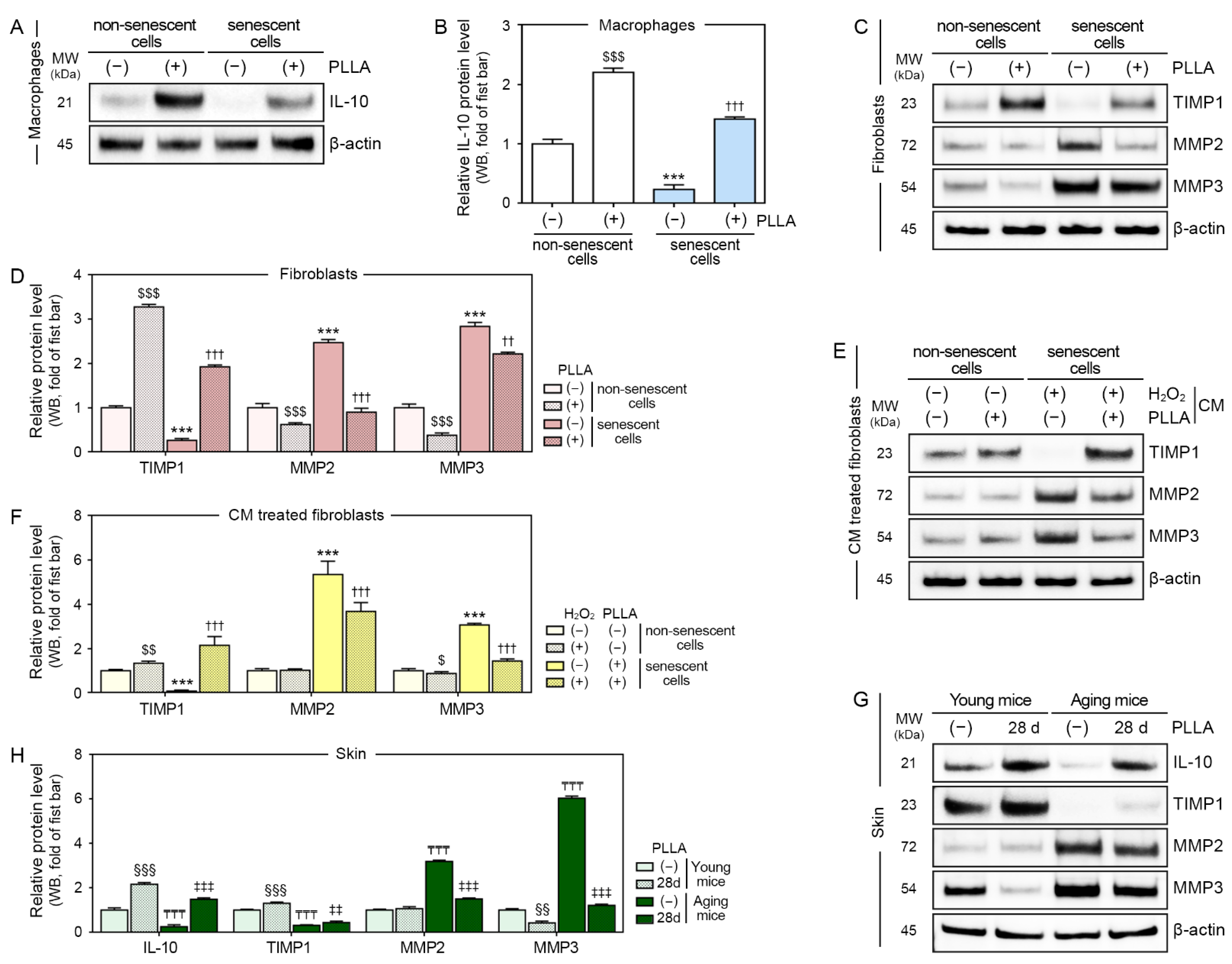
3.6. PLLA Increased the Expression of IL-10 and TIMP1 and Decreased the Expression of MMP2 and MMP3
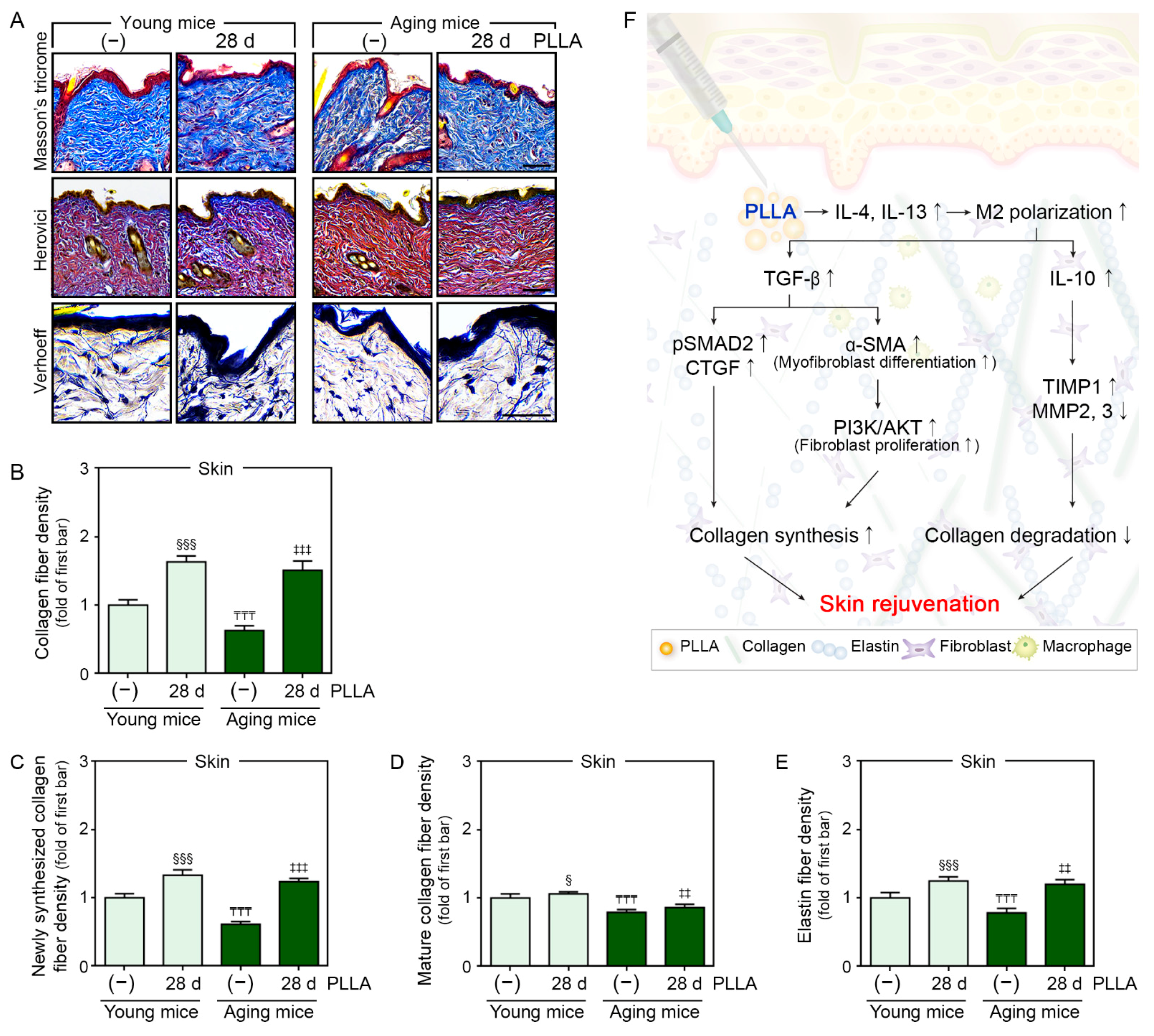
3.7. PLLA Increased Collagen and Elastin Fiber Content in Aged Skin
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sadick, N.S.; Manhas-Bhutani, S.; Krueger, N. A novel approach to structural facial volume replacement. Aesthetic Plast. Surg. 2013, 37, 266–276. [Google Scholar] [CrossRef]
- Johl, S.S.; Burgett, R.A. Dermal filler agents: A practical review. Curr. Opin. Ophthalmol. 2006, 7, 471–479. [Google Scholar] [CrossRef]
- Sánchez-Carpintero, I.; Candelas, D.; Ruiz-Rodríguez, R. Dermal fillers: Types, indications, and complications. Actas Dermosifiliogr. 2010, 101, 381–393. [Google Scholar] [CrossRef]
- Kruglikov, I.L.; Wollina, U. Soft tissue fillers as non-specific modulators of adipogenesis: Change of the paradigm? Exp. Dermatol. 2015, 24, 912–915. [Google Scholar] [CrossRef]
- Wise, J.B.; Greco, T. Injectable treatments for the aging face. Facial Plast. Surg. 2006, 22, 140–146. [Google Scholar] [CrossRef]
- Wu, D.C.; Karnik, J.; Margarella, T.; Nguyen, V.L.; Calame, A.; Goldman, M.P. Evaluation of the in vivo effects of various laser, light, or ultrasound modalities on human skin treated with a collagen and polymethylmethacrylate microsphere dermal filler product. Lasers Surg. Med. 2016, 48, 811–819. [Google Scholar] [CrossRef]
- Lee, J.C.; Lorenc, Z.P. Synthetic fillers for facial rejuvenation. Clin. Plast. Surg. 2016, 43, 497–503. [Google Scholar] [CrossRef]
- Yu, H.X.; Huang, N.X.; Wang, C.S.; Tang, Z.L. Modeling of poly(L-lactide) thermal degradation: Theoretical prediction of molecular weight and polydispersity index. J. Appl. Polym. Sci. 2003, 88, 2557–2562. [Google Scholar] [CrossRef]
- Hu, K.; Lv, Q.; Cui, F.Z.; Xu, L.; Jiao, Y.P.; Wang, Y.; Feng, Q.L. A novel poly(L-lactide) (PLLA)/Fibroin hybrid scaffold to promote hepatocyte viability and decrease macrophage responses. J. Bioact. Compat. Polym. 2007, 22, 395–410. [Google Scholar] [CrossRef]
- Moyle, G.J.; Lysakova, L.; Brown, S.; Sibtain, N.; Healy, J.; Priest, C.; Mandalia, S.; Barton, S.E. A randomized open-label study of immediate versus delayed polylactic acid injections for the cosmetic management of facial lipoatrophy in persons with HIV infection. HIV Med. 2004, 5, 82–87. [Google Scholar] [CrossRef]
- Brady, J.M.; Cutright, D.E.; Miller, R.A.; Barristone, G.C. Resorption rate, route, route of elimination, and ultrastructure of the implant site of polylactic acid in the abdominal wall of the rat. J. Biomed. Mater. Res. 1973, 7, 155–166. [Google Scholar] [CrossRef]
- Fitzgerald, R.; Graivier, M.H.; Kane, M.; Lorenc, Z.P.; Vleggaar, D.; Werschler, W.P.; Jeffrey, M.K. Update on facial aging. Aesthet. Surg. J. 2010, 30, 11S–24S. [Google Scholar] [CrossRef]
- unge, K.; Binnebösel, M.; Von Trotha, K.T.; Rosch, R.; Klinge, U.; Neumann, U.P.; Jansen, P.L. Mesh biocompatibility: Effects of cellular inflammation and tissue remodelling. Langenbecks Arch. Surg. 2012, 397, 255–270. [Google Scholar]
- Wilson, C.J.; Clegg, R.E.; Leavesley, D.I.; Pearcy, M.J. Mediation of biomaterial-cell interactions by adsorbed proteins: A review. Tissue Eng. 2005, 11, 1–18. [Google Scholar] [CrossRef]
- Helming, L.; Gordon, S. Molecular mediators of macrophage fusion. Trends Cell. Biol. 2009, 19, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Stein, P.; Vitavska, O.; Kind, P.; Hoppe, W.; Wieczorek, H.; Schürer, N.Y. The biological basis for poly-L-lactic acid-induced augmentation. J. Dermatol. Sci. 2015, 78, 26–33. [Google Scholar] [CrossRef]
- Duggan, S.; Essig, F.; Hünniger, K.; Mokhtari, Z.; Bauer, L.; Lehnert, T.; Brandes, S.; Häder, A.; Jacobsen, I.D.; Martin, R.; et al. Neutrophil activation by Candida glabrata but not Candida albicans promotes fungal uptake by monocytes. Cell. Microbiol. 2015, 17, 1259–1276. [Google Scholar] [CrossRef] [PubMed]
- Reedy, J.L.; Negoro, P.E.; Feliu, M.; Lord, A.K.; Khan, N.S.; Lukason, D.P.; Wiederhold, N.P.; Tam, J.M.; Mansour, M.K.; Patterson, T.F.; et al. The carbohydrate lectin receptor dectin-1 mediates the immune response to exserohilum rostratum. Infect. Immun. 2017, 85, e00903–e00916. [Google Scholar] [CrossRef]
- Heung, L.J. Monocytes and the host response to fungal pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Suzuki. K.; Meguro, K.; Nakagomi, D.; Nakajima, H. Roles of alternatively activated M2 macrophages in allergic contact dermatitis. Allergol. Int. 2017, 66, 392–397. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.-F.; Gao, C.-C.; Yi, J.; Zhao, J.-L.; Liang, S.-Q.; Zhao, Y.; Ye, Y.-C.; Bai, J.; Zheng, Q.-J.; Dou, K.-F.; et al. Cytotherapy with M1-polarized macrophages ameliorates liver fibrosis by modulating immune microenvironment in mice. J. Hepatol. 2017, 67, 770–779. [Google Scholar] [CrossRef]
- Oh, S.; Rho, N.-K.; Byun, K.-A.; Yang, J.Y.; Sun, H.J.; Jang, M.; Kang, D.; Son, K.H.; Byun, K. Combined treatment of monopolar and bipolar radiofrequency increases skin elasticity by decreasing the accumulation of advanced glycated end products in aged animal skin. Int. J. Mol. Sci. 2022, 23, 2993. [Google Scholar] [CrossRef]
- Park, J.E.; Pyun, H.B.; Woo, S.W.; Jeong, J.H.; Hwang, J.K. The protective effect of Kaempferia parviflora extract on UVB-induced skin photoaging in hairless mice. Photodermatol. Photoimmunol. Photomed. 2014, 30, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Imokawa, G.; Ishida, K. Biological mechanisms underlying the ultraviolet radiation-induced formation of skin wrinkling and sagging I: Reduced skin elasticity, highly associated with enhanced dermal elastase activity, triggers wrinkling and sagging. Int. J. Mol. Sci. 2015, 16, 7753–7775. [Google Scholar] [CrossRef]
- Kim, H.S.; Park, S.Y.; Moon, S.H.; Lee, J.D.; Kim, S. Autophagy in human skin fibroblasts: Impact of age. Int. J. Mol. Sci. 2018, 19, 2254. [Google Scholar] [CrossRef]
- Mokos, Z.B.; Ćurković, D.; Kostović, K.; Čeović, R. Facial changes in the mature patient. Clin. Dermatol. 2018, 36, 152–158. [Google Scholar] [CrossRef]
- Varani, J.; Dame, M.K.; Rittie, L.; Fligiel, S.E.; Kang, S.; Fisher, G.J.; Voorhees, J.J. Decreased collagen production in chronologically aged skin: Roles of age-dependent alteration in fibroblast function and defective mechanical stimulation. Am. J. Pathol. 2006, 168, 1861–1868. [Google Scholar] [CrossRef]
- Cabral, L.R.B.; Teixeira, L.N.; Gimenez, R.P.; Demasi, A.P.D.; de Brito, R.B.; de Araújo, V.C.; Martinez, E.F. Effect of hyaluronic acid and poly-L-lactic acid dermal fillers on collagen synthesis: An in vitro and in vivo study. Clin. Cosmet. Investig. Dermatol. 2020, 13, 701–710. [Google Scholar] [CrossRef]
- Lin, C.Y.; Lin, J.Y.; Yang, D.Y.; Lee, S.H.; Kim, J.Y.; Kang, M. Efficacy and safety of poly-D,L-lactic acid microspheres as subdermal fillers in animals. Plast. Aesthet. Res. 2019, 6, 16. [Google Scholar] [CrossRef]
- Kwon, T.; Han, S.W.; Yeo, I.K.; Kim, J.H.; Kim, J.M.; Hong, J.; Lee, B.; Lee, S.; Moon, H.S.; Kwon, H.J.; et al. Biostimulatory effects of polydioxanone, poly-D,L lactic acid, and polycaprolactone fillers in mouse model. J. Cosmet. Dermatol. 2019, 18, 1002–1008. [Google Scholar] [CrossRef]
- Savioli Lopes, M.; Jardini, A.; Maciel Filho, R. Synthesis and characterizations of poly (lactic acid) by ring-opening polymerization for biomedical applications. Chem. Eng. Trans. 2014, 38, 331–336. [Google Scholar]
- Pholharn, D.; Srithep, Y.; Morris, J. Ring opening polymerization of poly(Llactide) by macroinitiator. AIP Conf. Proc. 2019, 2065, 030016. [Google Scholar]
- Elmowafy, E.M.; Tiboni, M.; Soliman, M.E. Biocompatibility, biodegradation and biomedical applications of poly(lactic acid)/poly(lactic-co-glycolic acid) micro and nanoparticles. J. Pharm. Investig. 2019, 49, 347–380. [Google Scholar] [CrossRef]
- Pholharn, D.; Srithep, Y.; Morris, J. Effect of initiators on synthesis of poly(L-lactide) by ring opening polymerization. IOP Conf. Ser: Mater. Sci. Eng. 2017, 213, 012022. [Google Scholar] [CrossRef]
- Wu, Y.H.; Cheng, M.L.; Ho, H.Y.; Chiu, D.T.; Wang, T.C. Telomerase prevents accelerated senescence in glucose-6-phosphate dehydrogenase (G6PD)-deficient human fibroblasts. J. Biomed. Sci. 2009, 16, 18. [Google Scholar] [CrossRef]
- Du, L.; Chen, E.; Wu, T.; Ruan, Y.; Wu, S. Resveratrol attenuates hydrogen peroxide-induced aging through upregulation of autophagy in human umbilical vein endothelial cells. Drug. Des. Devel Ther. 2019, 13, 747–755. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, B.; Byun, H.J.; Yu, L.; Nguyen, T.M.; Nguyen, T.H.; Do, P.A.; Kim, E.J.; Cheong, K.A.; Kim, K.S.; et al. Resolvin D1 Suppresses H2O2-Induced Senescence in Fibroblasts by Inducing Autophagy through the miR-1299/ARG2/ARL1 Axis. Antioxidants 2021, 10, 1924. [Google Scholar] [CrossRef]
- Wu, R.X.; Ma, C.; Liang, Y.; Chen, F.M.; Liu, X. ECM-mimicking nanofibrous matrix coaxes macrophages toward an anti-inflammatory phenotype: Cellular behaviors and transcriptome analysis. Appl. Mater. Today 2020, 18, 100508. [Google Scholar] [CrossRef]
- Shi, X.; Young, C.D.; Zhou, H.; Wang, X. Transforming growth factor-β signaling in fibrotic diseases and cancer-associated fibroblasts. Biomolecules 2020, 10, 1666. [Google Scholar] [CrossRef]
- Ricardo, S.D.; van Goor, H.; Eddy, A.A. Macrophage diversity in renal injury and repair. J. Clin. Investig. 2008, 118, 3522–3530. [Google Scholar] [CrossRef]
- Hata, A.; Chen, Y.G. TGF-beta signaling from receptors to smads. Cold Spring Harb. Perspect. Biol. 2016, 8, a022061. [Google Scholar] [CrossRef]
- Herovici, C. Picropolychrome: Histological staining technic intended for the study of normal and pathological connective tissue. Rev. Fr. Etud. Clin. Biol. 1963, 8, 88–89. [Google Scholar] [PubMed]
- Anthony, P.P. Manual of histological demonstration techniques. J. Clin. Pathol. 1975, 28, 339. [Google Scholar] [CrossRef]
- Fan, Y.; Choi, T.H.; Chung, J.H.; Jeon, Y.K.; Kim, S. Hyaluronic acid-cross-linked filler stimulates collagen type 1 and elastic fiber synthesis in skin through the TGF-β/Smad signaling pathway in a nude mouse model. J. Plast. Reconstr. Aesthet. Surg. 2019, 72, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Garcés, V.; Molina Aguilar, P.; Bea Serrano, C.; García Bustos, V.; Benavent Seguí, J.; Ferrández Izquierdo, A.; Ruiz-Saurí, A. Age-related dermal collagen changes during development, maturation and ageing- a morphometric and comparative study. J. Anat. 2014, 225, 98–108. [Google Scholar] [CrossRef]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, V. Sculptra: A stimulatory filler. Facial Plast. Surg. 2009, 25, 95–99. [Google Scholar] [CrossRef]
- Lowe, N.J. Optimizing poly-l-lactic acid use. J. Cosmet. Laser Ther. 2008, 10, 43–46. [Google Scholar] [CrossRef]
- Ray, S.; Ta, H.T. Investigating the effect of biomaterials such as poly-(l-Lactic Acid) particles on collagen synthesis in vitro: Method is matter. J. Funct. Biomater. 2020, 11, 51. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, B.; Peng, L.; Gao, S.; Guo, J.; Zhu, X. Blockade of LINC01605-enriched exosome generation in M2 macrophages impairs M2 macrophage-induced proliferation, migration, and invasion of human dermal fibroblasts. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211016724. [Google Scholar] [CrossRef]
- Kreimendahl, F.; Marquardt, Y.; Apel, C.; Bartneck, M.; Zwadlo-Klarwasser, G.; Hepp, J.; Jockenhoevel, S.; Baron, J.M. Macrophages significantly enhance wound healing in a vascularized skin model. J. Biomed. Mater. Res. A 2019, 107, 1340–1350. [Google Scholar] [CrossRef] [PubMed]
- Horiba, S.; Kami, R.; Tsutsui, T.; Hosoi, J. IL-34 downregulation-associated M1/M2 macrophage imbalance is related to inflammaging in sun-exposed human skin. JID Innov. 2022, 2, 100112. [Google Scholar] [CrossRef] [PubMed]
- Alqahtani, M.S.; Syed, R.; Alshehri, M. Size-dependent phagocytic uptake and immunogenicity of gliadin nanoparticles. Polymers 2020, 12, 2576. [Google Scholar] [CrossRef]
- Vaine, C.A.; Patel, M.K.; Zhu, J.; Lee, E.; Finberg, R.W.; Hayward, R.C.; Kurt-Jones, E.A. Tuning innate immune activation by surface texturing of polymer microparticles: The role of shape in inflammasome activation. J. Immunol. 2013, 190, 3525–3532. [Google Scholar] [CrossRef]
- Mammadov, R.; Cinar, G.; Gunduz, N.; Goktas, M.; Kayhan, H.; Tohumeken, S.; Topal, A.E.; Orujalipoor, I.; Delibasi, T.; Dana, A.; et al. Virus-like nanostructures for tuning immune response. Sci. Rep. 2015, 5, 16728. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A.; Vannella, K.M. Macrophages in tissue repair, regeneration, and fibrosis. Immunity 2016, 44, 450–462. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y. TGF-β in fibrosis by acting as a conductor for contractile properties of myofibroblasts. Cell Biosci. 2019, 9, 98. [Google Scholar] [CrossRef]
- Thannickal, V.J.; Lee, D.Y.; White, E.S.; Cui, Z.; Larios, J.M.; Chacon, R.; Horowitz, J.C.; Day, R.M.; Thomas, P.E. Myofibroblast differentiation by transforming growth factor-beta1 is dependent on cell adhesion and integrin signaling via focal adhesion kinase. J. Biol. Chem. 2003, 278, 12384–12389. [Google Scholar] [CrossRef]
- Kulkarni, A.A.; Thatcher, T.H.; Olsen, K.C.; Maggirwar, S.B.; Phipps, R.P.; Sime, P.J. PPAR-γ ligands repress TGFβ-induced myofibroblast differentiation by targeting the PI3K/Akt pathway: Implications for therapy of fibrosis. PLoS ONE 2011, 6, e15909. [Google Scholar] [CrossRef]
- Kielty, C.M.; Sherratt, M.J.; Shuttleworth, C.A. Elastic fibres. J. Cell. Sci. 2002, 115, 2817–2828. [Google Scholar] [CrossRef]
- Fisher, G.J.; Datta, S.C.; Talwar, H.S.; Wang, Z.Q.; Varani, J.; Kang, S.; Voorhees, J.J. Molecular basis of sun-induced premature skin ageing and retinoid antagonism. Nature 1996, 379, 335–339. [Google Scholar] [CrossRef] [PubMed]
- Brennan, M.; Bhatti, H.; Nerusu, K.C.; Bhagavathula, N.; Kang, S.; Fisher, G.J.; Varani, J.; Voorhees, J.J. Matrix metalloproteinase-1 is the major collagenolytic enzyme responsible for collagen damage in UV-irradiated human skin. Photochem. Photobiol. 2003, 78, 43–48. [Google Scholar] [CrossRef]
- Sternlicht, M.D.; Werb, Z. How matrix metalloproteinases regulate cell behavior. Annu. Rev. Cell Dev. Biol. 2001, 17, 463–516. [Google Scholar] [CrossRef] [PubMed]
- Ueno, T.; Tanaka, K.; Kaneko, K.; Taga, Y.; Sata, T.; Irie, S.; Hattori, S.; Ogawa-Goto, K. Enhancement of procollagen biosynthesis by p180 through augmented ribosome association on the endoplasmic reticulum in response to stimulated secretion. J. Biol. Chem. 2010, 285, 29941–29950. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, S.; Lee, J.H.; Kim, H.M.; Batsukh, S.; Sung, M.J.; Lim, T.H.; Lee, M.H.; Son, K.H.; Byun, K. Poly-L-Lactic Acid Fillers Improved Dermal Collagen Synthesis by Modulating M2 Macrophage Polarization in Aged Animal Skin. Cells 2023, 12, 1320. https://doi.org/10.3390/cells12091320
Oh S, Lee JH, Kim HM, Batsukh S, Sung MJ, Lim TH, Lee MH, Son KH, Byun K. Poly-L-Lactic Acid Fillers Improved Dermal Collagen Synthesis by Modulating M2 Macrophage Polarization in Aged Animal Skin. Cells. 2023; 12(9):1320. https://doi.org/10.3390/cells12091320
Chicago/Turabian StyleOh, Seyeon, Je Hyuk Lee, Hyoung Moon Kim, Sosorburam Batsukh, Mi Jeong Sung, Tae Hwan Lim, Myoung Hoon Lee, Kuk Hui Son, and Kyunghee Byun. 2023. "Poly-L-Lactic Acid Fillers Improved Dermal Collagen Synthesis by Modulating M2 Macrophage Polarization in Aged Animal Skin" Cells 12, no. 9: 1320. https://doi.org/10.3390/cells12091320
APA StyleOh, S., Lee, J. H., Kim, H. M., Batsukh, S., Sung, M. J., Lim, T. H., Lee, M. H., Son, K. H., & Byun, K. (2023). Poly-L-Lactic Acid Fillers Improved Dermal Collagen Synthesis by Modulating M2 Macrophage Polarization in Aged Animal Skin. Cells, 12(9), 1320. https://doi.org/10.3390/cells12091320