
Pregnane X Receptor Signaling Pathway and Vitamin K: Molecular Mechanisms and Clinical Relevance in Human Health


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
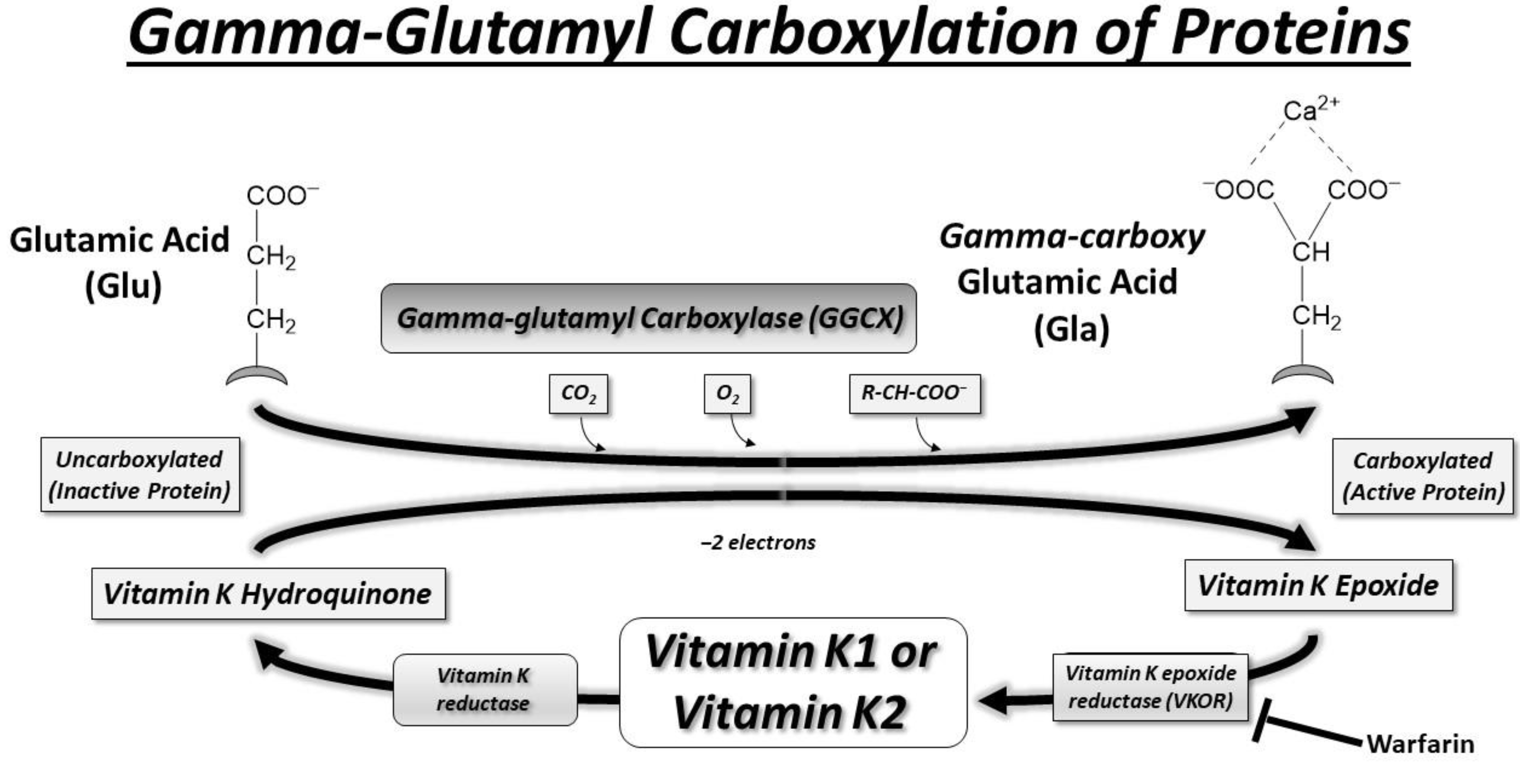
2. Vitamin K and γ-Glutamyl Carboxylase
3. Gla Proteins and Human Health
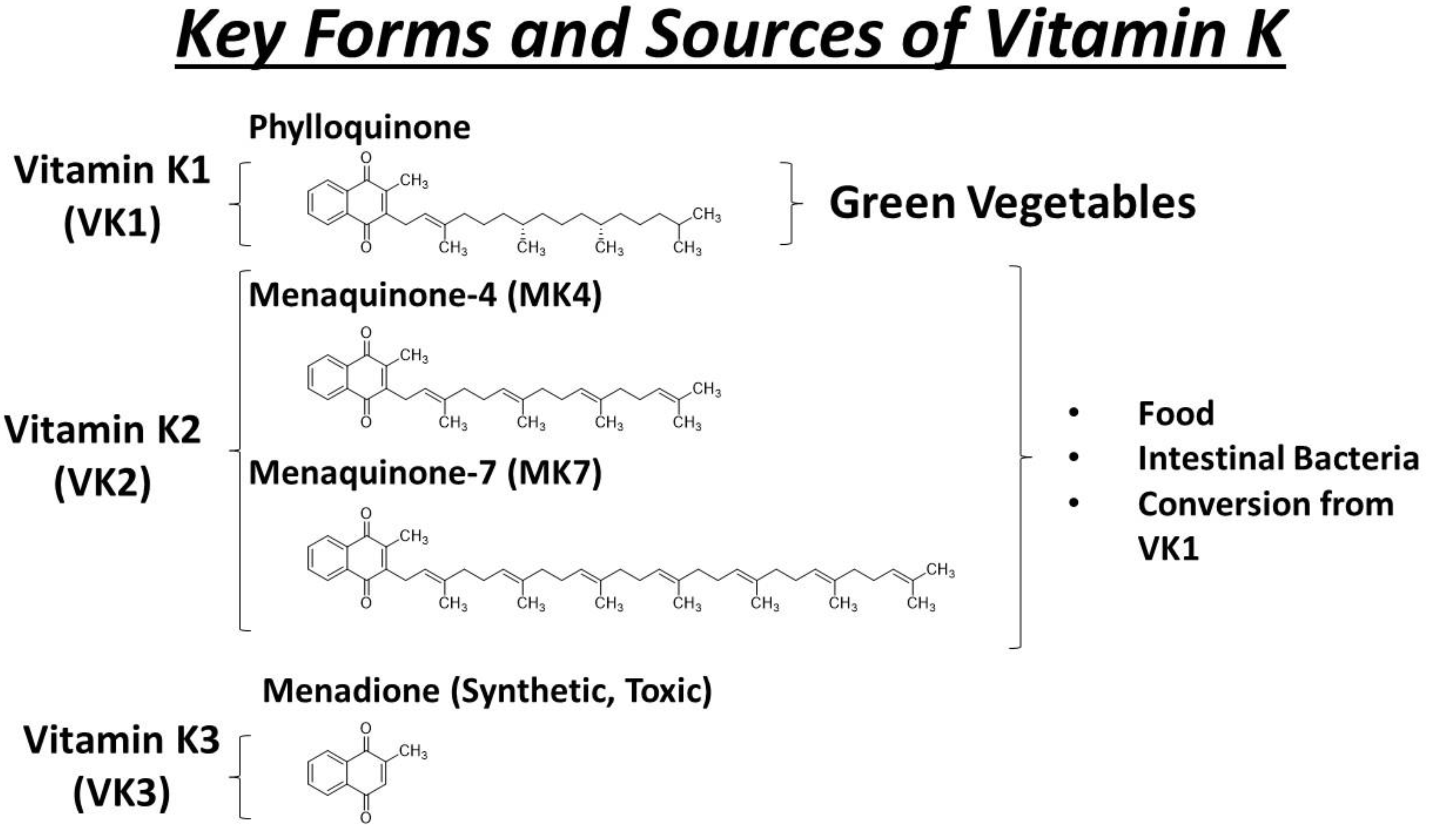
4. Key Forms and Sources of Vitamin K
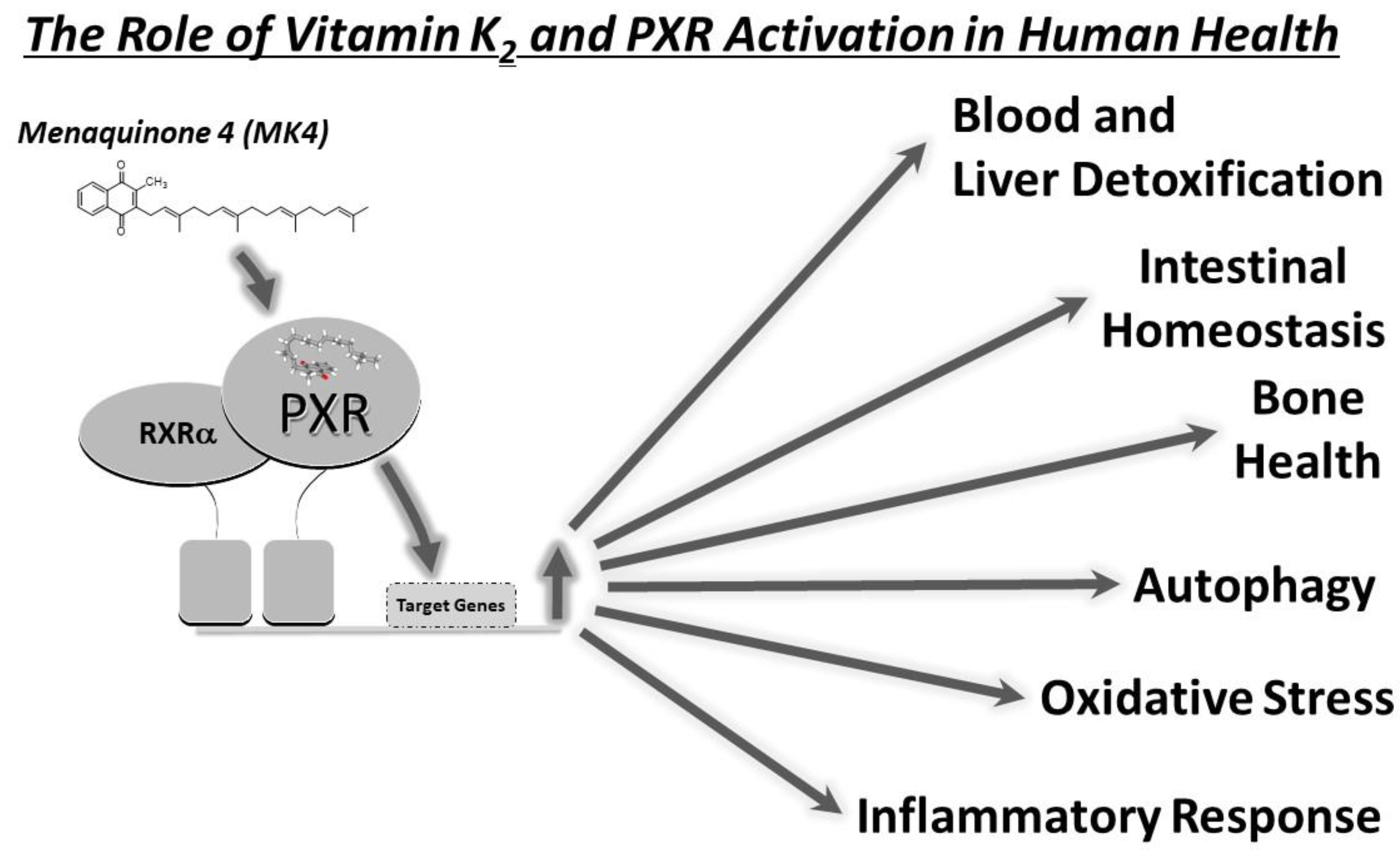
5. MK-4 and PXR
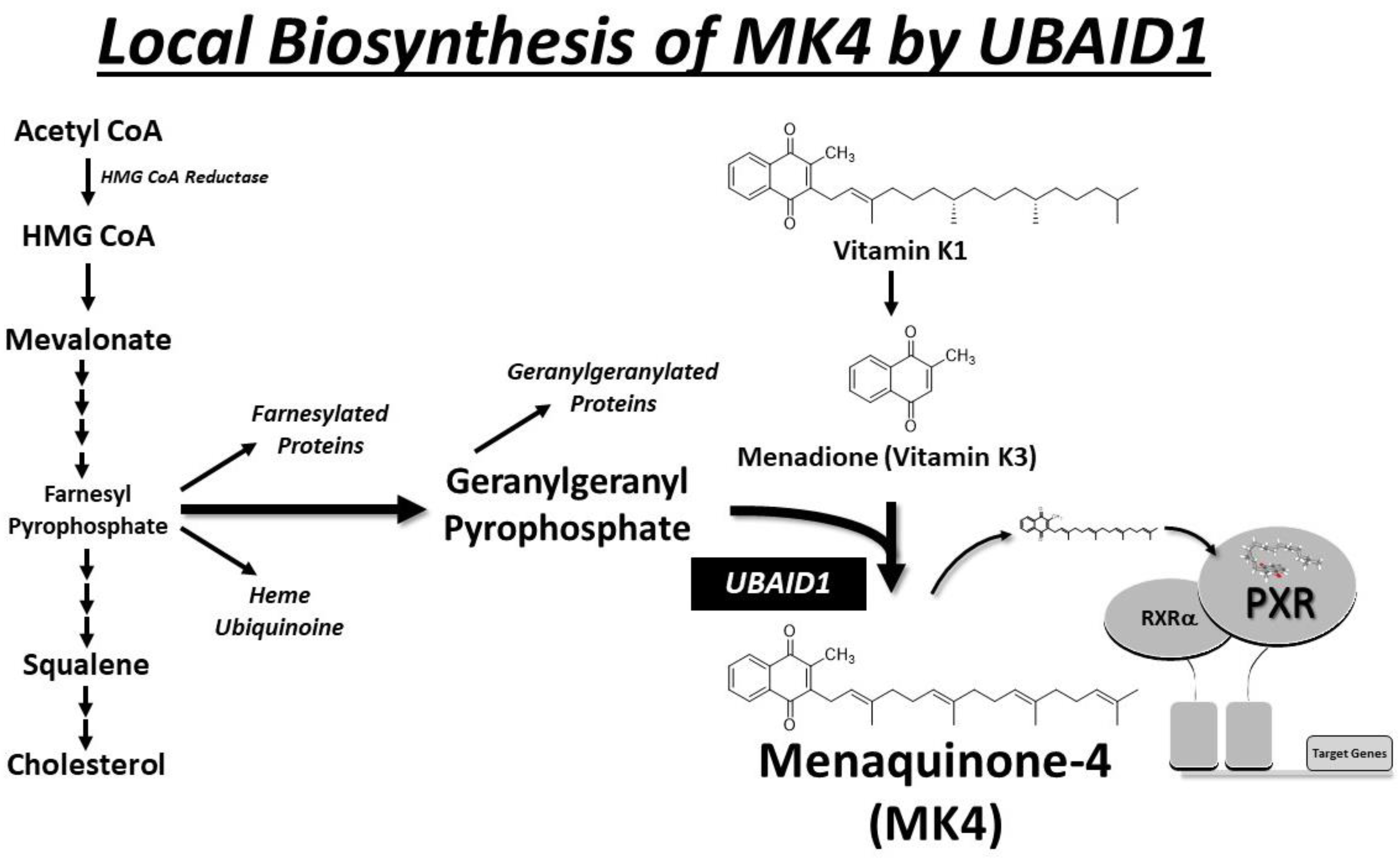
6. Local Biosynthesis of MK-4
7. Vitamin K2, PXR Activation, and Inflammatory Bowel Disease
8. Vitamin K2, PXR Activation, and Cancer
9. PXR Activation, Autophagy, and Bone Health
10. Therapeutic Implications of PXR Activation by Vitamin K
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bertilsson, G.; Heidrich, J.; Svensson, K.; Asman, M.; Jendeberg, L.; Sydow-Bäckman, M.; Ohlsson, R.; Postlind, H.; Blomquist, P.; Berkenstam, A. Identification of a human nuclear receptor defines a new signaling pathway for CYP3A induction. Proc. Natl. Acad. Sci. USA 1998, 95, 12208–12213. [Google Scholar] [CrossRef] [PubMed]
- Blumberg, B.; Sabbagh, W., Jr.; Juguilon, H.; Bolado, J., Jr.; van Meter, C.M.; Ong, E.S.; Evans, R.M. SXR, a novel steroid and xenobiotic-sensing nuclear receptor. Genes Dev. 1998, 12, 3195–3205. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Moore, J.T.; Wade, L.; Staudinger, J.L.; Watson, M.A.; Jones, S.A.; McKee, D.D.; Oliver, B.B.; Willson, T.M.; Zetterström, R.H.; et al. An orphan nuclear receptor activated by pregnanes defines a novel steroid signaling pathway. Cell 1998, 92, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J.M.; McKee, D.D.; Watson, M.A.; Willson, T.M.; Moore, J.T.; Kliewer, S.A. The human orphan nuclear receptor PXR is activated by compounds that regulate CYP3A4 gene expression and cause drug interactions. J. Clin. Investig. 1998, 102, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, B.; Hodgson, E.; Liddle, C. The orphan human pregnane X receptor mediates the transcriptional activation of CYP3A4 by rifampicin through a distal enhancer module. Mol. Pharmacol. 1999, 56, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Kliewer, S.A.; Lehmann, J.M.; Milburn, M.V.; Willson, T.M. The PPARs and PXRs: Nuclear xenobiotic receptors that define novel hormone signaling pathways. Recent Prog. Horm. Res. 1999, 54, 345–367; discussion 367–348. [Google Scholar]
- Moore, D.D. Regulation of drug transport by new xenobiotic receptors. Pharmacogenomics J. 2001, 1, 224–225. [Google Scholar] [CrossRef] [PubMed]
- Schuetz, E.; Strom, S. Promiscuous regulator of xenobiotic removal. Nat. Med. 2001, 7, 536–537. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.; Liu, Y.; Madan, A.; Habeebu, S.; Klaassen, C.D. Coordinate regulation of xenobiotic and bile acid homeostasis by pregnane X receptor. Drug Metab. Dispos. Biol. Fate Chem. 2001, 29, 1467–1472. [Google Scholar] [PubMed]
- Dutta, M.; Lim, J.J.; Cui, J.Y. Pregnane X Receptor and the Gut-Liver Axis: A Recent Update. Drug Metab. Dispos. Biol. Fate Chem. 2022, 50, 478–491. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.; Xie, W. Orphan nuclear receptors, PXR and LXR: New ligands and therapeutic potential. Expert Opin. Ther. Targets 2004, 8, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Holick, M.F. Stay tuned to PXR: An orphan actor that may not be D-structive only to bone. J. Clin. Investig. 2005, 115, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Xu, C.; Staudinger, J.L. Pregnane X receptor is SUMOylated to repress the inflammatory response. J. Pharmacol. Exp. Ther. 2010, 335, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Ding, X.; Lichti, K. Pregnane X receptor and natural products: Beyond drug-drug interactions. Expert Opin. Drug Metab. Toxicol. 2006, 2, 847–857. [Google Scholar] [CrossRef]
- Staudinger, J.L.; Goodwin, B.; Jones, S.A.; Hawkins-Brown, D.; MacKenzie, K.I.; LaTour, A.; Liu, Y.; Klaassen, C.D.; Brown, K.K.; Reinhard, J.; et al. The nuclear receptor PXR is a lithocholic acid sensor that protects against liver toxicity. Proc. Natl. Acad. Sci. USA 2001, 98, 3369–3374. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Madan, A.; Carol, K.M.; Parkinson, A. Regulation of drug transporter gene expression by nuclear receptors. Drug Metab. Dispos. Biol. Fate Chem. 2003, 31, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Woody, S.; Sun, M.; Cui, W. Nuclear-receptor-mediated regulation of drug- and bile-acid-transporter proteins in gut and liver. Drug Metab. Rev. 2013, 45, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Xu, C.; Biswas, A.; Mani, S. Post-translational modification of pregnane x receptor. Pharmacol. Res. 2011, 64, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L.; Xu, C.; Cui, Y.J.; Klaassen, C.D. Nuclear receptor-mediated regulation of carboxylesterase expression and activity. Expert Opin. Drug Metab. Toxicol. 2010, 6, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Cui, W.; Woody, S.K.; Staudinger, J.L. Pregnane X receptor modulates the inflammatory response in primary cultures of hepatocytes. Drug Metab. Dispos. Biol. Fate Chem. 2015, 43, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Rogers, R.S.; Parker, A.; Vainer, P.D.; Elliott, E.; Sudbeck, D.; Parimi, K.; Peddada, V.P.; Howe, P.G.; D’Ambrosio, N.; Ruddy, G.; et al. The Interface between Cell Signaling Pathways and Pregnane X Receptor. Cells 2021, 10, 3262. [Google Scholar] [CrossRef]
- Creamer, B.A.; Sloan, S.N.B.; Dennis, J.F.; Rogers, R.; Spencer, S.; McCuen, A.; Persaud, P.; Staudinger, J.L. Associations between Pregnane X Receptor and Breast Cancer Growth and Progression. Cells 2020, 9, 2295. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Inoue, S. Multiple Modes of Vitamin K Actions in Aging-Related Musculoskeletal Disorders. Int. J. Mol. Sci. 2019, 20, 2844. [Google Scholar] [CrossRef]
- Azuma, K.; Ouchi, Y.; Inoue, S. Vitamin K: Novel molecular mechanisms of action and its roles in osteoporosis. Geriatr. Gerontol. Int. 2014, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Shiba, S.; Hasegawa, T.; Ikeda, K.; Urano, T.; Horie-Inoue, K.; Ouchi, Y.; Amizuka, N.; Inoue, S. Osteoblast-Specific γ-Glutamyl Carboxylase-Deficient Mice Display Enhanced Bone Formation with Aberrant Mineralization. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2015, 30, 1245–1254. [Google Scholar] [CrossRef]
- Azuma, K.; Urano, T.; Ouchi, Y.; Inoue, S. Vitamin K2 suppresses proliferation and motility of hepatocellular carcinoma cells by activating steroid and xenobiotic receptor. Endocr. J. 2009, 56, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Urano, T.; Watabe, T.; Ouchi, Y.; Inoue, S. PROX1 suppresses vitamin K-induced transcriptional activity of Steroid and Xenobiotic Receptor. Genes Cells Devoted Mol. Cell. Mech. 2011, 16, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Cianciolo, G.; Brandi, M.L.; Ferrari, S.; Nickolas, T.L.; Tripepi, G.; Plebani, M.; Zaninotto, M.; Iervasi, G.; La Manna, G.; et al. Vitamin K and Osteoporosis. Nutrients 2020, 12, 3625. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Cianciolo, G.; Evenepoel, P.; Schurgers, L.; Plebani, M. Vitamin K in CKD Bone Disorders. Calcif. Tissue Int. 2021, 108, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Hirota, Y.; Suhara, Y. New Aspects of Vitamin K Research with Synthetic Ligands: Transcriptional Activity via SXR and Neural Differentiation Activity. Int. J. Mol. Sci. 2019, 20, 3006. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, N.; Shiraki, M. Vitamin K Nutrition and Bone Health. Nutrients 2020, 12, 1909. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.; Bak, M.J.; Narvaez, C.J. New insights into vitamin K biology with relevance to cancer. Trends Mol. Med. 2022, 28, 864–881. [Google Scholar] [CrossRef] [PubMed]
- Dam, H. The antihaemorrhagic vitamin of the chick. Biochem. J. 1935, 29, 1273–1285. [Google Scholar] [CrossRef] [PubMed]
- Binkley, S.B.; Cheney, L.C.; Holcomb, W.F.; McKee, R.W.; Thayer, S.A.; MacCorquodale, D.W.; Doisy, E.A. The constitution and synthesis of vitamin K1. J. Am. Chem. Soc. 1939, 61, 2558–2559. [Google Scholar] [CrossRef]
- Sadowski, J.A.; Esmon, C.T.; Suttie, J.W. Vitamin K-dependent carboxylase. Requirements of the rat liver microsomal enzyme system. J. Biol. Chem. 1976, 251, 2770–2776. [Google Scholar] [CrossRef] [PubMed]
- Siegfried, C.M. Solubilization of vitamin K epoxide reductase and vitamin K-dependent carboxylase from rat liver microsomes. Biochem. Biophys. Res. Commun. 1978, 83, 1488–1495. [Google Scholar] [CrossRef] [PubMed]
- Uotila, L.; Suttie, J.W. Recent findings in understanding the biological function of vitamin K. Med. Biol. 1982, 60, 16–24. [Google Scholar] [PubMed]
- Campbell, H.A.; Smith, W.K.; Roberts, W.L.; Link, K.P. Studies on the hemorrhagic sweet clover disease: II. the bioassay of hemorrhagic concentrates by following the prothrombin level in the plasma of rabbit blood. J. Biol. Chem. 1941, 138, 1–20. [Google Scholar] [CrossRef]
- Link, K.P. The discovery of dicumarol and its sequels. Circulation 1959, 19, 97–107. [Google Scholar] [CrossRef]
- Shearer, M.J. Role of vitamin K and Gla proteins in the pathophysiology of osteoporosis and vascular calcification. Curr. Opin. Clin. Nutr. Metab. Care 2000, 3, 433–438. [Google Scholar] [CrossRef]
- Shearer, M.J.; Newman, P. Metabolism and cell biology of vitamin K. Thromb. Haemost. 2008, 100, 530–547. [Google Scholar] [PubMed]
- Stock, M.; Schett, G. Vitamin K-Dependent Proteins in Skeletal Development and Disease. Int. J. Mol. Sci. 2021, 22, 9328. [Google Scholar] [CrossRef] [PubMed]
- Elodi, S.; Elodi, P. Surface-governed molecular regulation of blood coagulation. Mol. Asp. Med. 1983, 6, 291–353. [Google Scholar] [CrossRef] [PubMed]
- Kodama, S.; Yoshii, N.; Ota, A.; Takeshita, J.I.; Yoshinari, K.; Ono, A. Association between in vitro nuclear receptor-activating profiles of chemical compounds and their in vivo hepatotoxicity in rats. J. Toxicol. Sci. 2021, 46, 569–587. [Google Scholar] [CrossRef]
- Komai, M.; Shirakawa, H. Vitamin K metabolism. Menaquinone-4 (MK-4) formation from ingested VK analogues and its potent relation to bone function. Clin. Calcium 2007, 17, 1663–1672. [Google Scholar] [PubMed]
- Akbari, S.; Rasouli-Ghahroudi, A.A. Vitamin K and Bone Metabolism: A Review of the Latest Evidence in Preclinical Studies. BioMed Res. Int. 2018, 2018, 4629383. [Google Scholar] [CrossRef] [PubMed]
- Bjorklund, G.; Svanberg, E.; Dadar, M.; Card, D.J.; Chirumbolo, S.; Harrington, D.J.; Aaseth, J. The Role of Matrix Gla Protein (MGP) in Vascular Calcification. Curr. Med. Chem. 2020, 27, 1647–1660. [Google Scholar] [CrossRef] [PubMed]
- Ngo, E.O.; Sun, T.P.; Chang, J.Y.; Wang, C.C.; Chi, K.H.; Cheng, A.L.; Nutter, L.M. Menadione-induced DNA damage in a human tumor cell line. Biochem. Pharmacol. 1991, 42, 1961–1968. [Google Scholar] [CrossRef] [PubMed]
- Morgan, W.A.; Hartley, J.A.; Cohen, G.M. Quinone-induced DNA single strand breaks in rat hepatocytes and human chronic myelogenous leukaemic K562 cells. Biochem. Pharmacol. 1992, 44, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Loor, G.; Kondapalli, J.; Schriewer, J.M.; Chandel, N.S.; Vanden Hoek, T.L.; Schumacker, P.T. Menadione triggers cell death through ROS-dependent mechanisms involving PARP activation without requiring apoptosis. Free Radic. Biol. Med. 2010, 49, 1925–1936. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Smith, B.L.; Fowke, E.A. Haemolytic activity and nephrotoxicity of 2-hydroxy-1,4-naphthoquinone in rats. J. Appl. Toxicol. JAT 1991, 11, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Smith, B.L.; Munday, C.M. Toxicity of 2,3-dialkyl-1,4-naphthoquinones in rats: Comparison with cytotoxicity in vitro. Free Radic. Biol. Med. 1995, 19, 759–765. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Smith, B.L.; Munday, C.M. Effects of butylated hydroxyanisole and dicoumarol on the toxicity of menadione to rats. Chem. Biol. Interact. 1998, 108, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.L.; Karl, J.P.; Oliverio, A.M.; Fu, X.; Soares, J.W.; Wolfe, B.E.; Hernandez, C.J.; Mason, J.B.; Booth, S.L. Dietary vitamin K is remodeled by gut microbiota and influences community composition. Gut Microbes 2021, 13, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Singla, R.K.; Qin, Q.; Ding, Y.; Liu, L.; Ding, X.; Qu, W.; Huang, C.; Shen, Z.; Shen, B.; et al. Exploring the complex relationship between vitamin K, gut microbiota, and warfarin variability in cardiac surgery patients. Int. J. Surg. 2023, 109, 3861–3871. [Google Scholar] [CrossRef] [PubMed]
- Karl, J.P.; Meydani, M.; Barnett, J.B.; Vanegas, S.M.; Barger, K.; Fu, X.; Goldin, B.; Kane, A.; Rasmussen, H.; Vangay, P.; et al. Fecal concentrations of bacterially derived vitamin K forms are associated with gut microbiota composition but not plasma or fecal cytokine concentrations in healthy adults. Am. J. Clin. Nutr. 2017, 106, 1052–1061. [Google Scholar] [CrossRef]
- Marinova, M.; Lütjohann, D.; Breuer, O.; Kölsch, H.; Westhofen, P.; Watzka, M.; Mengel, M.; Stoffel-Wagner, B.; Hartmann, G.; Coch, C.; et al. VKORC1-dependent pharmacokinetics of intravenous and oral phylloquinone (vitamin K1) mixed micelles formulation. Eur. J. Clin. Pharmacol. 2013, 69, 467–475 . [Google Scholar] [CrossRef]
- Fusaro, M.; Mereu, M.C.; Aghi, A.; Iervasi, G.; Gallieni, M. Vitamin K and bone. Clin. Cases Miner. Bone Metab. Off. J. Ital. Soc. Osteoporos. Miner. Metab. Skelet. Dis. 2017, 14, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Mladěnka, P.; Macáková, K.; Kujovská Krčmová, L.; Javorská, L.; Mrštná, K.; Carazo, A.; Protti, M.; Remião, F.; Nováková, L.; OEMONOM Researchers and Collaborators. Vitamin K—Sources, physiological role, kinetics, deficiency, detection, therapeutic use, and toxicity. Nutr. Rev. 2021, 80, 677–698. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Gallieni, M.; Rizzo, M.A.; Stucchi, A.; Delanaye, P.; Cavalier, E.; Moysés, R.M.A.; Jorgetti, V.; Iervasi, G.; Giannini, S.; et al. Vitamin K plasma levels determination in human health. Clin. Chem. Lab. Med. 2017, 55, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Fusaro, M.; Noale, M.; Viola, V.; Galli, F.; Tripepi, G.; Vajente, N.; Plebani, M.; Zaninotto, M.; Guglielmi, G.; Miotto, D.; et al. Vitamin K, vertebral fractures, vascular calcifications, and mortality: VItamin K Italian (VIKI) dialysis study. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2012, 27, 2271–2278. [Google Scholar] [CrossRef] [PubMed]
- Simes, D.C.; Viegas, C.S.B.; Araujo, N.; Marreiros, C. Vitamin K as a Diet Supplement with Impact in Human Health: Current Evidence in Age-Related Diseases. Nutrients 2020, 12, 138. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef] [PubMed]
- Suhara, Y.; Watanabe, M.; Motoyoshi, S.; Nakagawa, K.; Wada, A.; Takeda, K.; Takahashi, K.; Tokiwa, H.; Okano, T. Synthesis of new vitamin K analogues as steroid and xenobiotic receptor (SXR) agonists: Insights into the biological role of the side chain part of vitamin K. J. Med. Chem. 2011, 54, 4918–4922. [Google Scholar] [CrossRef] [PubMed]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grün, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 regulation of bone homeostasis is mediated by the steroid and xenobiotic receptor SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Fang, Z.Z.; Nagaoka, K.; Okamoto, M.; Qu, A.; Tanaka, N.; Kimura, S.; Gonzalez, F.J. Activation of intestinal human pregnane X receptor protects against azoxymethane/dextran sulfate sodium-induced colon cancer. J. Pharmacol. Exp. Ther. 2014, 351, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Shah, Y.M.; Gonzalez, F.J. Pregnane X receptor as a target for treatment of inflammatory bowel disorders. Trends Pharmacol. Sci. 2012, 33, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Shah, Y.M.; Ma, X.; Pang, X.; Tanaka, T.; Kodama, T.; Krausz, K.W.; Gonzalez, F.J. Therapeutic role of rifaximin in inflammatory bowel disease: Clinical implication of human pregnane X receptor activation. J. Pharmacol. Exp. Ther. 2010, 335, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, N.; Ke, S.; Eagleton, N.; Xie, Y.; Chen, G.; Laffins, B.; Yao, H.; Zhou, B.; Tian, Y. Pregnane X receptor suppresses proliferation and tumourigenicity of colon cancer cells. Br. J. Cancer 2010, 102, 1753–1761. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Sun, Z.; Wang, Q.; Zhang, Y.; Jia, Z. Role of nuclear receptor PXR in immune cells and inflammatory diseases. Front. Immunol. 2022, 13, 969399. [Google Scholar] [CrossRef] [PubMed]
- Theocharis, S.; Giaginis, C.; Gourzi, S.; Alexandrou, P.; Tsourouflis, G.; Sarantis, P.; Danas, E.; Michail, A.; Tsoukalas, N.; Pergaris, A.; et al. High Pregnane X Receptor (PXR) Expression Is Correlated with Poor Prognosis in Invasive Breast Carcinoma. Diagnostics 2021, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Ma, Y.; Xiong, K.; Wang, Y.; Liu, Y.; Sun, Y.; Yang, Y.; Ma, A. Ameliorating Effects of Vitamin K2 on Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Int. J. Mol. Sci. 2023, 24, 2986. [Google Scholar] [CrossRef] [PubMed]
- McGarvey, T.W.; Nguyen, T.; Tomaszewski, J.E.; Monson, F.C.; Malkowicz, S.B. Isolation and characterization of the TERE1 gene, a gene down-regulated in transitional cell carcinoma of the bladder. Oncogene 2001, 20, 1042–1051. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Vitamin K2 induces phosphorylation of protein kinase A and expression of novel target genes in osteoblastic cells. J. Mol. Endocrinol. 2007, 39, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Otsuka, M.; Kato, N.; Ichimura, T.; Abe, S.; Tanaka, Y.; Taniguchi, H.; Hoshida, Y.; Moriyama, M.; Wang, Y.; Shao, R.X.; et al. Vitamin K2 binds 17beta-hydroxysteroid dehydrogenase 4 and modulates estrogen metabolism. Life Sci. 2005, 76, 2473–2482. [Google Scholar] [CrossRef]
- Karasawa, S.; Azuma, M.; Kasama, T.; Sakamoto, S.; Kabe, Y.; Imai, T.; Yamaguchi, Y.; Miyazawa, K.; Handa, H. Vitamin K2 covalently binds to Bak and induces Bak-mediated apoptosis. Mol. Pharmacol. 2013, 83, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.T.; Foley, A.L.; Engelke, J.A.; Suttie, J.W. Conversion of dietary phylloquinone to tissue menaquinone-4 in rats is not dependent on gut bacteria. J. Nutr. 1998, 128, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Ronden, J.E.; Drittij-Reijnders, M.J.; Vermeer, C.; Thijssen, H.H. Intestinal flora is not an intermediate in the phylloquinone-menaquinone-4 conversion in the rat. Biochim. Biophys. Acta 1998, 1379, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Thijssen, H.H.; Drittij-Reijnders, M.J. Vitamin K distribution in rat tissues: Dietary phylloquinone is a source of tissue menaquinone-4. Br. J. Nutr. 1994, 72, 415–425. [Google Scholar] [CrossRef]
- Will, B.H.; Usui, Y.; Suttie, J.W. Comparative metabolism and requirement of vitamin K in chicks and rats. J. Nutr. 1992, 122, 2354–2360. [Google Scholar] [CrossRef] [PubMed]
- Shearer, M.J.; Newman, P. Recent trends in the metabolism and cell biology of vitamin K with special reference to vitamin K cycling and MK-4 biosynthesis. J. Lipid Res. 2014, 55, 345–362. [Google Scholar] [CrossRef] [PubMed]
- Weisshof, R.; Chermesh, I. Micronutrient deficiencies in inflammatory bowel disease. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 576–581. [Google Scholar] [CrossRef]
- Pulakazhi Venu, V.K.; Saifeddine, M.; Mihara, K.; Tsai, Y.C.; Nieves, K.; Alston, L.; Mani, S.; McCoy, K.D.; Hollenberg, M.D.; Hirota, S.A. The pregnane X receptor and its microbiota-derived ligand indole 3-propionic acid regulate endothelium-dependent vasodilation. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E350–E361. [Google Scholar] [CrossRef] [PubMed]
- Venkatesh, M.; Mukherjee, S.; Wang, H.; Li, H.; Sun, K.; Benechet, A.P.; Qiu, Z.; Maher, L.; Redinbo, M.R.; Phillips, R.S.; et al. Symbiotic bacterial metabolites regulate gastrointestinal barrier function via the xenobiotic sensor PXR and Toll-like receptor 4. Immunity 2014, 41, 296–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xiong, L.; Ruan, Z.; Gong, X.; Luo, Y.; Wu, C.; Wang, Y.; Shang, H.; Chen, J. Indole-3-propionic acid alleviates sepsis-associated acute liver injury by activating pregnane X receptor. Mol. Med. 2023, 29, 65. [Google Scholar] [CrossRef] [PubMed]
- Flannigan, K.L.; Nieves, K.M.; Szczepanski, H.E.; Serra, A.; Lee, J.W.; Alston, L.A.; Ramay, H.; Mani, S.; Hirota, S.A. The Pregnane X Receptor and Indole-3-Propionic Acid Shape the Intestinal Mesenchyme to Restrain Inflammation and Fibrosis. Cell. Mol. Gastroenterol. Hepatol. 2023, 15, 765–795. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Cheung, C.; Krausz, K.W.; Shah, Y.M.; Wang, T.; Idle, J.R.; Gonzalez, F.J. A double transgenic mouse model expressing human pregnane X receptor and cytochrome P450 3A4. Drug Metab. Dispos. Biol. Fate Chem. 2008, 36, 2506–2512. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Shah, Y.; Cheung, C.; Guo, G.L.; Feigenbaum, L.; Krausz, K.W.; Idle, J.R.; Gonzalez, F.J. The PREgnane X receptor gene-humanized mouse: A model for investigating drug-drug interactions mediated by cytochromes P450 3A. Drug Metab. Dispos. Biol. Fate Chem. 2007, 35, 194–200. [Google Scholar] [CrossRef]
- Ma, X.; Shah, Y.M.; Guo, G.L.; Wang, T.; Krausz, K.W.; Idle, J.R.; Gonzalez, F.J. Rifaximin is a gut-specific human pregnane X receptor activator. J. Pharmacol. Exp. Ther. 2007, 322, 391–398. [Google Scholar] [CrossRef]
- Shah, Y.M.; Ma, X.; Morimura, K.; Kim, I.; Gonzalez, F.J. Pregnane X receptor activation ameliorates DSS-induced inflammatory bowel disease via inhibition of NF-kappaB target gene expression. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1114–G1122. [Google Scholar] [CrossRef] [PubMed]
- Nieves, K.M.; Hirota, S.A.; Flannigan, K.L. Xenobiotic receptors and the regulation of intestinal homeostasis: Harnessing the chemical output of the intestinal microbiota. Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 322, G268–G281. [Google Scholar] [CrossRef] [PubMed]
- Vyhlídalová, B.; Bartoňková, I.; Jiskrová, E.; Li, H.; Mani, S.; Dvořák, Z. Differential activation of human pregnane X receptor PXR by isomeric mono-methylated indoles in intestinal and hepatic in vitro models. Toxicol. Lett. 2020, 324, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Staudinger, J.L. Clinical applications of small molecule inhibitors of Pregnane X receptor. Mol. Cell. Endocrinol. 2019, 485, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Deuring, J.J.; Li, M.; Cao, W.; Chen, S.; Wang, W.; de Haar, C.; van der Woude, C.J.; Peppelenbosch, M. Pregnane X receptor activation constrains mucosal NF-κB activity in active inflammatory bowel disease. PLoS ONE 2019, 14, e0221924. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Yan, J.; Niu, Y. PXR: A center of transcriptional regulation in cancer. Acta Pharm. Sin. B 2020, 10, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Dvořák, Z.; Li, H.; Mani, S. Microbial Metabolites as Ligands to Xenobiotic Receptors: Chemical Mimicry as Potential Drugs of the Future. Drug Metab. Dispos. Biol. Fate Chem. 2023, 51, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Ledala, N.; Malik, M.; Rezaul, K.; Paveglio, S.; Provatas, A.; Kiel, A.; Caimano, M.; Zhou, Y.; Lindgren, J.; Krasulova, K.; et al. Bacterial Indole as a Multifunctional Regulator of Klebsiella oxytoca Complex Enterotoxicity. mBio 2022, 13, e0375221. [Google Scholar] [CrossRef] [PubMed]
- Little, M.; Dutta, M.; Li, H.; Matson, A.; Shi, X.; Mascarinas, G.; Molla, B.; Weigel, K.; Gu, H.; Mani, S.; et al. Understanding the physiological functions of the host xenobiotic-sensing nuclear receptors PXR and CAR on the gut microbiome using genetically modified mice. Acta Pharm. Sin. B 2022, 12, 801–820. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Ma, P.; Kong, L.; Wang, X.; Wang, Y.; Jiang, L. Vitamin K2 Inhibits Hepatocellular Carcinoma Cell Proliferation by Binding to 17β-Hydroxysteroid Dehydrogenase 4. Front. Oncol. 2021, 11, 757603. [Google Scholar] [CrossRef] [PubMed]
- Okano, T. A New Horizon in Vitamin K Research. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2016, 136, 1141–1159. [Google Scholar] [CrossRef] [PubMed]
- Sultana, H.; Kato, A.; Ohashi, A.; Takashima, R.; Katsurai, T.; Sato, S.; Monma, M.; Ohsaki, Y.; Goto, T.; Komai, M.; et al. Effect of Vitamin K-Mediated PXR Activation on Drug-Metabolizing Gene Expression in Human Intestinal Carcinoma LS180 Cell Line. Nutrients 2021, 13, 1709. [Google Scholar] [CrossRef] [PubMed]
- Pondugula, S.R.; Pavek, P.; Mani, S. Pregnane X Receptor and Cancer: Context-Specificity is Key. Nucl. Recept. Res. 2016, 3, 101198. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gong, W.; Wang, G.; Yu, E.; Tian, J.; Xia, Y.; Li, Z.; Zhang, K.; Xie, J. Role of nuclear pregnane X receptor in Cu-induced lipid metabolism and xenobiotic responses in largemouth bass (Micropterus salmoides). Front. Endocrinol. 2022, 13, 950985. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lu, H.; Wang, W.; Song, L.; Liu, M.; Cao, Y.; Qi, X.; Sun, J.; Gong, L. Prevention of D-GalN/LPS-induced ALI by 18β-glycyrrhetinic acid through PXR-mediated inhibition of autophagy degradation. Cell Death Dis. 2021, 12, 480. [Google Scholar] [CrossRef] [PubMed]
- Larabi, A.; Barnich, N.; Nguyen, H.T.T. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy 2020, 16, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, H.; Dong, M.; Zhu, P.; Cai, Y. The anticancer effects and mechanisms of fucoxanthin combined with other drugs. J. Cancer Res. Clin. Oncol. 2019, 145, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Chen, Z.; Wu, L.; Su, Y.; Wang, X.; Tang, N. Inhibitory effect of PXR on ammonia-induced hepatocyte autophagy via P53. Toxicol. Lett. 2018, 295, 153–161. [Google Scholar] [CrossRef]
- Sugatani, J.; Noguchi, Y.; Hattori, Y.; Yamaguchi, M.; Yamazaki, Y.; Ikari, A. Threonine-408 Regulates the Stability of Human Pregnane X Receptor through Its Phosphorylation and the CHIP/Chaperone-Autophagy Pathway. Drug Metab. Dispos. Biol. Fate Chem. 2016, 44, 137–150. [Google Scholar] [CrossRef]
- Azuma, K.; Casey, S.C.; Ito, M.; Urano, T.; Horie, K.; Ouchi, Y.; Kirchner, S.; Blumberg, B.; Inoue, S. Pregnane X receptor knockout mice display osteopenia with reduced bone formation and enhanced bone resorption. J. Endocrinol. 2010, 207, 257–263. [Google Scholar] [CrossRef]
- Saeki, N.; Itoh, Y.; Kanai, R.; Itoh, S.; Inububishi, T.; Akiyama, S.; Inui-Yamamoto, C.; Abe, M. Pregnane X receptor (PXR) represses osteoblast differentiation through repression of the Hedgehog signaling pathway. Exp. Cell Res. 2022, 416, 113156. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Jiang, C.; Hou, J.; Qian, H.; Chu, F.; Zhang, W.; Ye, M.; Chen, Z.; Liu, J.; Yao, H.; et al. Patchouli Alcohol Modulates the Pregnancy X Receptor/Toll-like Receptor 4/Nuclear Factor Kappa B Axis to Suppress Osteoclastogenesis. Front. Pharmacol. 2021, 12, 684976. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.E.; Wang, Z.; Liao, M.Z.; Lin, Y.S.; Shuhart, M.C.; Schuetz, E.G.; Thummel, K.E. Human PXR-mediated induction of intestinal CYP3A4 attenuates 1α,25-dihydroxyvitamin D₃ function in human colon adenocarcinoma LS180 cells. Biochem. Pharmacol. 2012, 84, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Assem, M.; Tay, J.C.; Watkins, P.B.; Blumberg, B.; Schuetz, E.G.; Thummel, K.E. Steroid and xenobiotic receptor and vitamin D receptor crosstalk mediates CYP24 expression and drug-induced osteomalacia. J. Clin. Investig. 2006, 116, 1703–1712. [Google Scholar] [CrossRef] [PubMed]
- Buchman, C.D.; Chai, S.C.; Chen, T. A current structural perspective on PXR and CAR in drug metabolism. Expert Opin. Drug Metab. Toxicol. 2018, 14, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Chai, S.C.; Wright, W.C.; Chen, T. Strategies for developing pregnane X receptor antagonists: Implications from metabolism to cancer. Med. Res. Rev. 2020, 40, 1061–1083. [Google Scholar] [CrossRef] [PubMed]
- Jabbari Hagh, E.; Mousavi, A.; Hejazian, S.M.; Haghi, M.; Esfahanian, S.; Ahmadian, E.; Zununi Vahed, S.; Ardalan, M. The Impact of Single Nucleotide Polymorphisms on the Pharmacokinetics of Tacrolimus in Kidney Allograft Recipients of Northern-West, Iran. Adv. Pharm. Bull. 2023, 13, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Meyer zu Schwabedissen, H.E.; Kim, R.B. Hepatic OATP1B transporters and nuclear receptors PXR and CAR: Interplay, regulation of drug disposition genes, and single nucleotide polymorphisms. Mol. Pharm. 2009, 6, 1644–1661. [Google Scholar] [CrossRef]
- Rana, M.; Coshic, P.; Goswami, R.; Tyagi, R.K. Influence of a critical single nucleotide polymorphism on nuclear receptor PXR-promoter function. Cell Biol. Int. 2017, 41, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Skandalaki, A.; Sarantis, P.; Theocharis, S. Pregnane X Receptor (PXR) Polymorphisms and Cancer Treatment. Biomolecules 2021, 11, 1142. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Staudinger, J.L.; Mahroke, A.; Patel, G.; Dattel, C.; Reddy, S. Pregnane X Receptor Signaling Pathway and Vitamin K: Molecular Mechanisms and Clinical Relevance in Human Health. Cells 2024, 13, 681. https://doi.org/10.3390/cells13080681
Staudinger JL, Mahroke A, Patel G, Dattel C, Reddy S. Pregnane X Receptor Signaling Pathway and Vitamin K: Molecular Mechanisms and Clinical Relevance in Human Health. Cells. 2024; 13(8):681. https://doi.org/10.3390/cells13080681
Chicago/Turabian StyleStaudinger, Jeff L., Avina Mahroke, Gauri Patel, Cole Dattel, and Sahana Reddy. 2024. "Pregnane X Receptor Signaling Pathway and Vitamin K: Molecular Mechanisms and Clinical Relevance in Human Health" Cells 13, no. 8: 681. https://doi.org/10.3390/cells13080681
APA StyleStaudinger, J. L., Mahroke, A., Patel, G., Dattel, C., & Reddy, S. (2024). Pregnane X Receptor Signaling Pathway and Vitamin K: Molecular Mechanisms and Clinical Relevance in Human Health. Cells, 13(8), 681. https://doi.org/10.3390/cells13080681