
Airway Epithelial Cell Cilia and Obstructive Lung Disease

Abstract
:1. Introduction
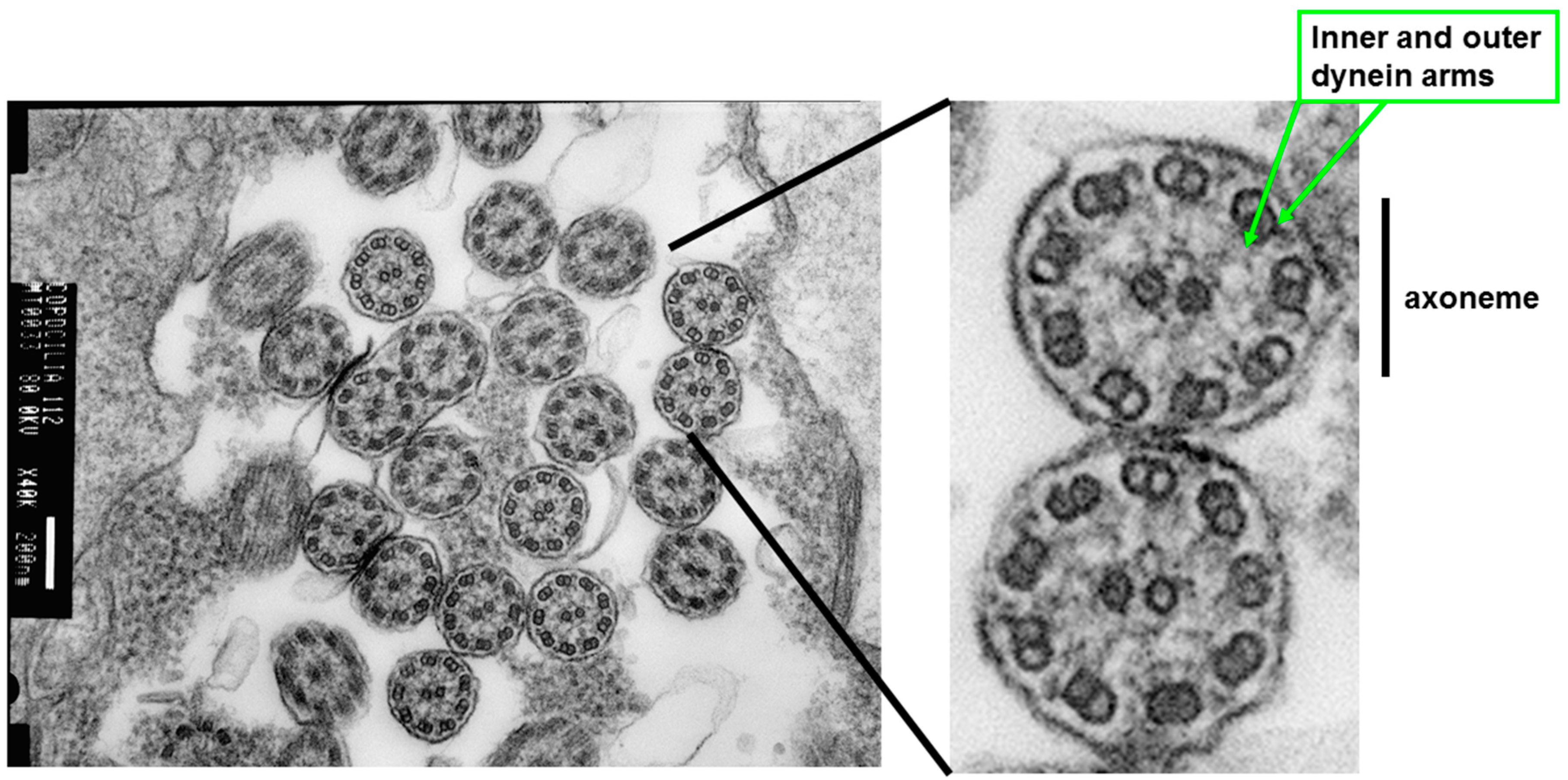
2. Airway Epithelium: A Specialized Physical, Secretory, and Regulatory Barrier
3. What Modulates Airway Cilia Function?
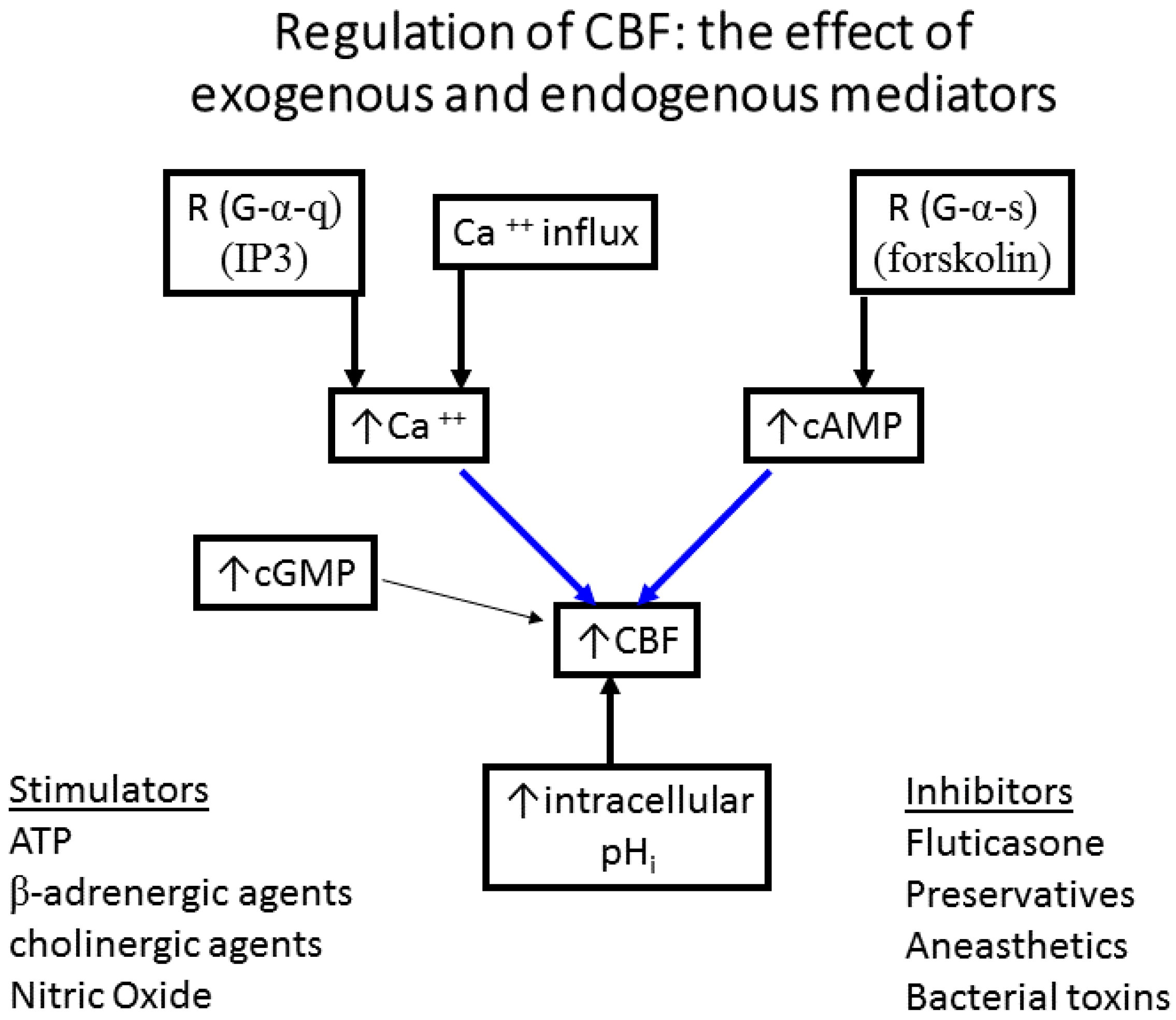
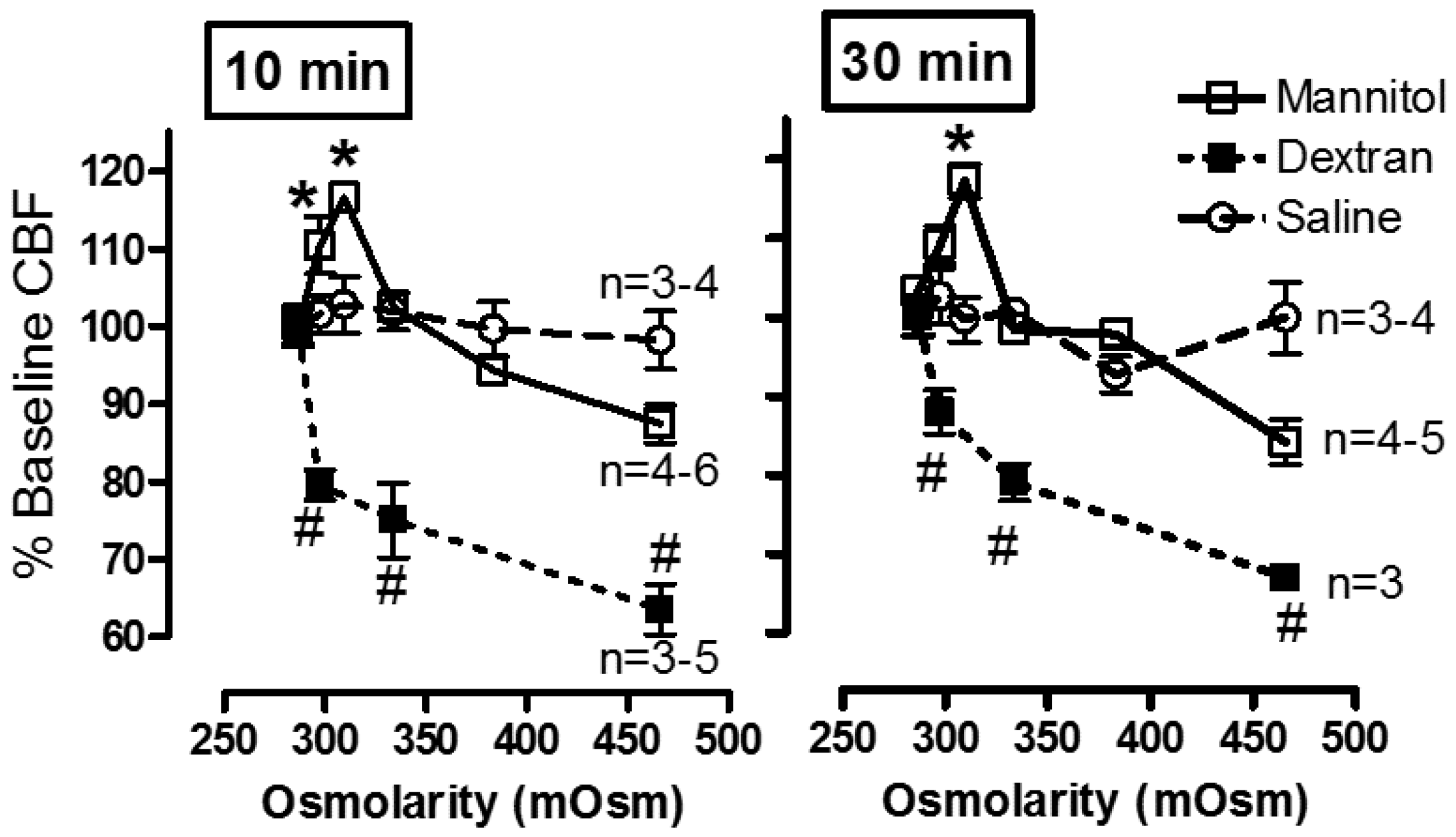
- The effect of exogenous and endogenous mediators on CBF: Airway cilia function can be modified by various endogenous and exogenous exposures (Figure 4) including infections such as rhinitis, rhinosinusitis [6,76,77], and pneumonia [78]; environmental pollutants [79,80,81]; cigarette smoke and oxidative stress [82,83,84,85], and the rheology of the mucus secretions [75,86,87]. Numerous agents and drugs (Table 1) have been shown to modulate CBF [16,17] with efficient mucociliary transport relying on effective regulation of ciliary beating [11,22]. In vitro, beta-adrenergic and cholinergic agents were demonstrated to stimulate CBF, while fluticasone, some preservatives, anaesthetics, and some bacterial toxins inhibited CBF [16,17,18,19]. In addition, the effect of hyperosmolar agents on the muco-ciliary transport apparatus has been documented [38,75]. In a study of the direct effect of mannitol on the CBF of human bronchial epithelial cells (Figure 5), we demonstrated a direct osmolarity-independent cilio-stimulatory effect, a unique mechanism of action for mannitol, compared to dextran and saline, on ciliary beating [53]. A systematic review of the literature that examines the effect of 229 drugs and other substances on CBF of airway epithelial cell cilia has been published [20].
- Relationship of inflammation to reduced or variable CBF: earlier studies in patients with cystic fibrosis have demonstrated a link between airway inflammation and a reduction in CBF [88,89]. Similar links between inflammation and the impaired mucociliary transport rate were investigated with sputum collected from COPD patients [90], and in ciliated nasal epithelial cells from stable bronchiectatic subjects [91]. However, the effects of inflammation on CBF are not always direct and can reflect modifications of epithelial differentiation and proliferation, possibly related to local chronic inflammation. This occurred in nasal airway cells from Rhinitis patients indicating a possible effect of inflammation on epithelial cell integrity and the number of cilia [92]. In addition, a correlation between ciliary activity and ciliary ultrastructure of the nasal mucosa in chronic and recurrent sinusitis was demonstrated [93] and leukotrienes released during inflammation could slow down CBF [94,95]. Bronchial cilia abnormalities worsened with the disease possibly contributing to the impairment of the mucociliary clearance in chronic bronchitis patients [96]. Also infections such as P. aeruginosa and H. influenzae caused a significant decrease in CBF of human nasal cilia, while staphylococcal products did not [97]. In comparison, RSV infection of human bronchial epithelial cells in culture resulted in ciliostasis and loss of cilia from the cell surface [98]. Resolution of infection and/or inflammation could restore normal cilia function several weeks following upper respiratory viral infection [99] or acute bronchiolitis in infancy [100].
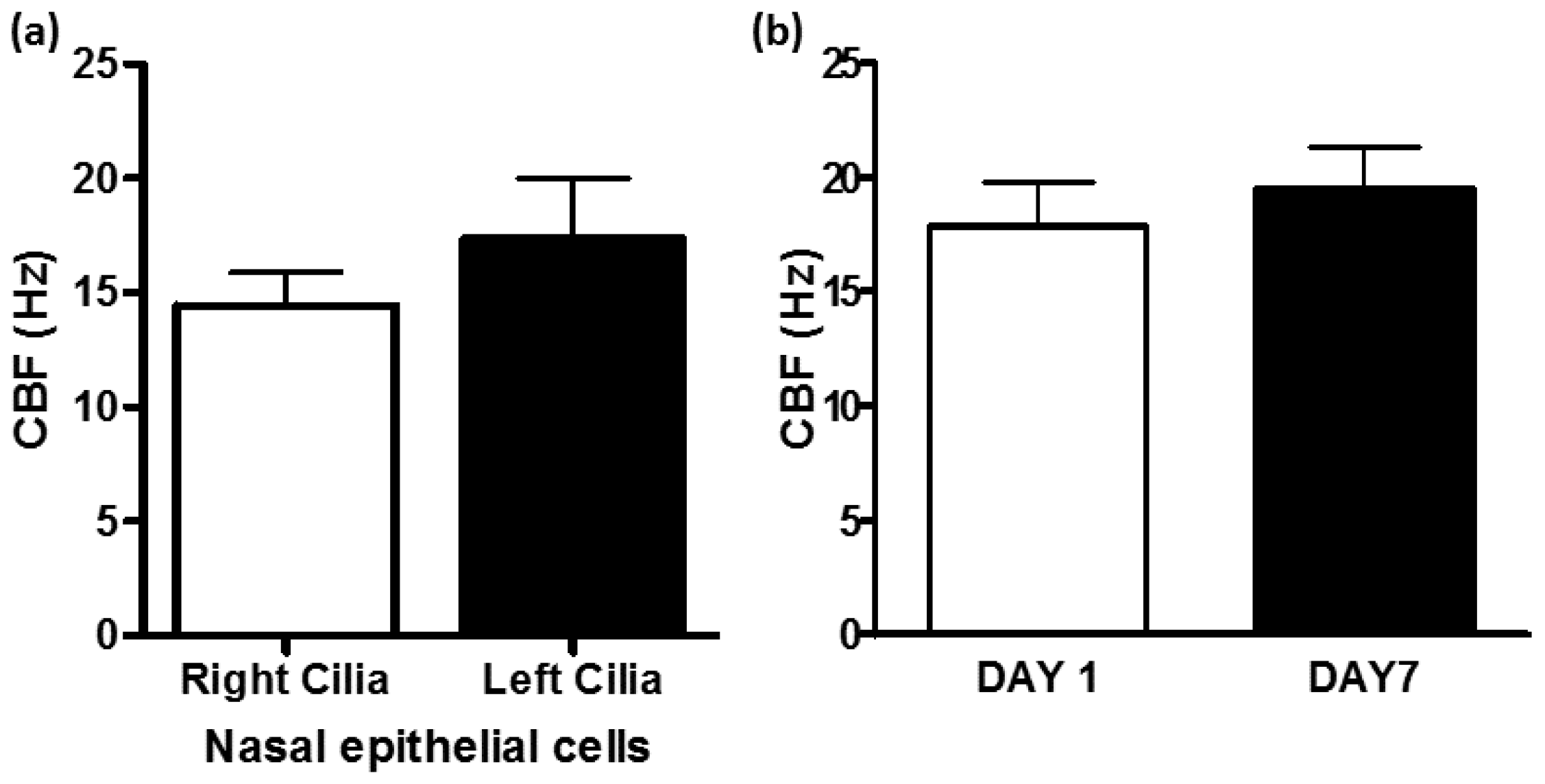
- Nasal ciliary changes can reflect bronchial changes: Respiratory cilia from different locations have been studied. Correlation between nasal, tracheal, and bronchial CBF and between mucociliary clearance and CBF were demonstrated [9,61,101,102,103]. In addition, nasal and bronchial cilia were shown to be comparable in function and structure allowing the use of nasal epithelial cells as surrogates of bronchial epithelial cells in airway inflammation studies [12]. Patients with allergic rhinitis and asthma have stronger nasal responsiveness to cold, dry air compared with patients with rhinitis alone, indicating that upper airway responses could represent lower airway responsiveness [104]. However, the use of nasal cilia as representative of tracheal and bronchial cilia for study of respiratory diseases such as asthma or COPD should be done with caution and preferably in the absence of rhinitis and runny nose due to allergies or active respiratory infections as these directly influence the differentiation and proliferation of nasal cilia and could negate their use as surrogates of tracheal and bronchial cilia. Evidence for this restriction has been demonstrated when intermittent allergic rhinitis was associated with decreased expression of anti-inflammatory genes in nasal fluids obtained from intermittent allergic rhinitis patients [105]. Also, when compared to controls, rhinitis patients exhibited lower percentages of ciliated cells and higher percentages of goblet and basal cells in nasal epithelial brushings related to local chronic inflammation [92]. In addition, respiratory viruses lead to ciliary defects in nasal epithelium of children with recovery occurring within 2–10 weeks after infection [99] and 13–17 weeks following acute bronchiolitis in infancy [100]. All these studies stress the need to study ciliated epithelium specimens in the absence of active respiratory infections.
4. Pathophysiology of COPD
5. Factors That Can Affect CBF in COPD
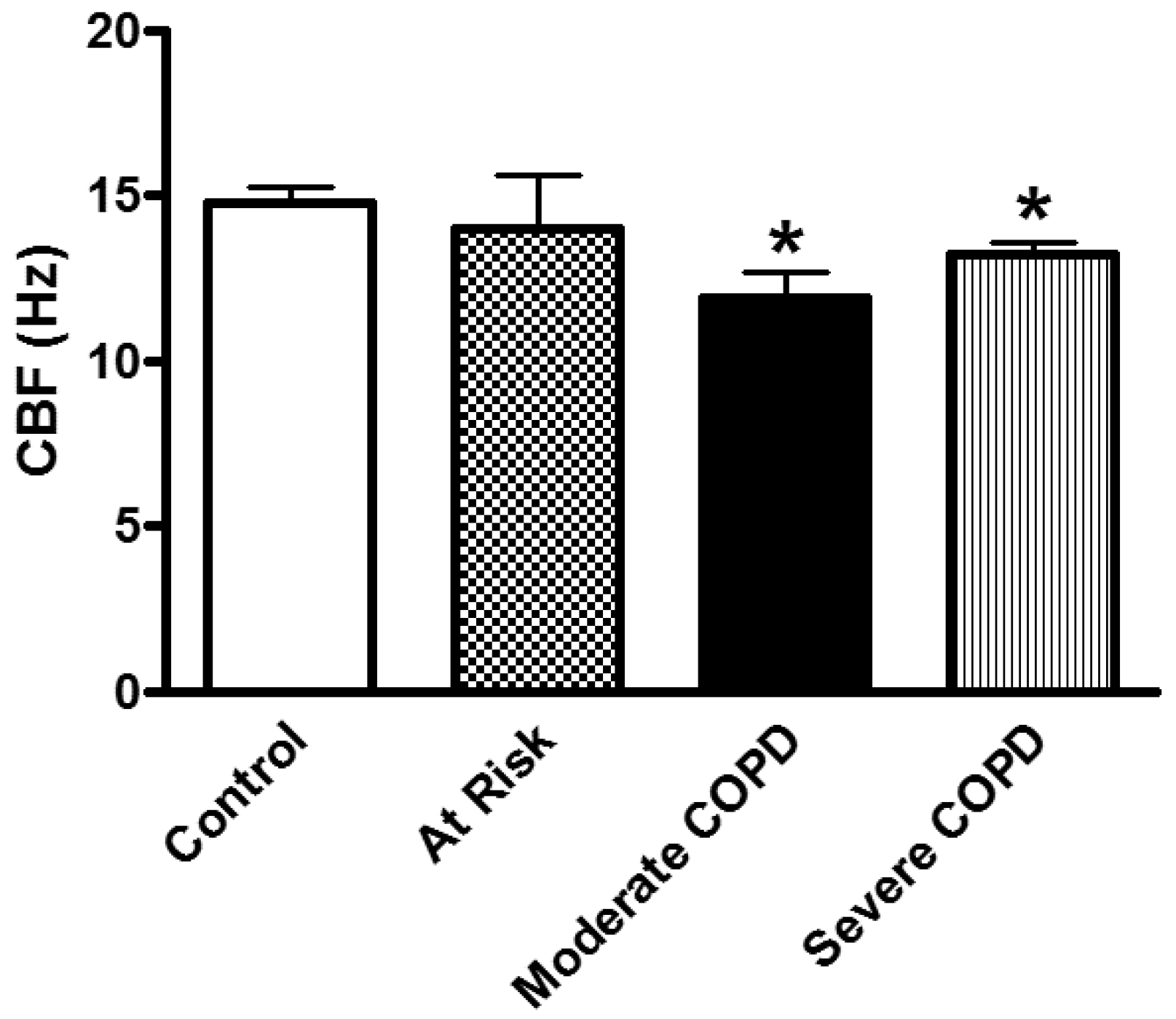
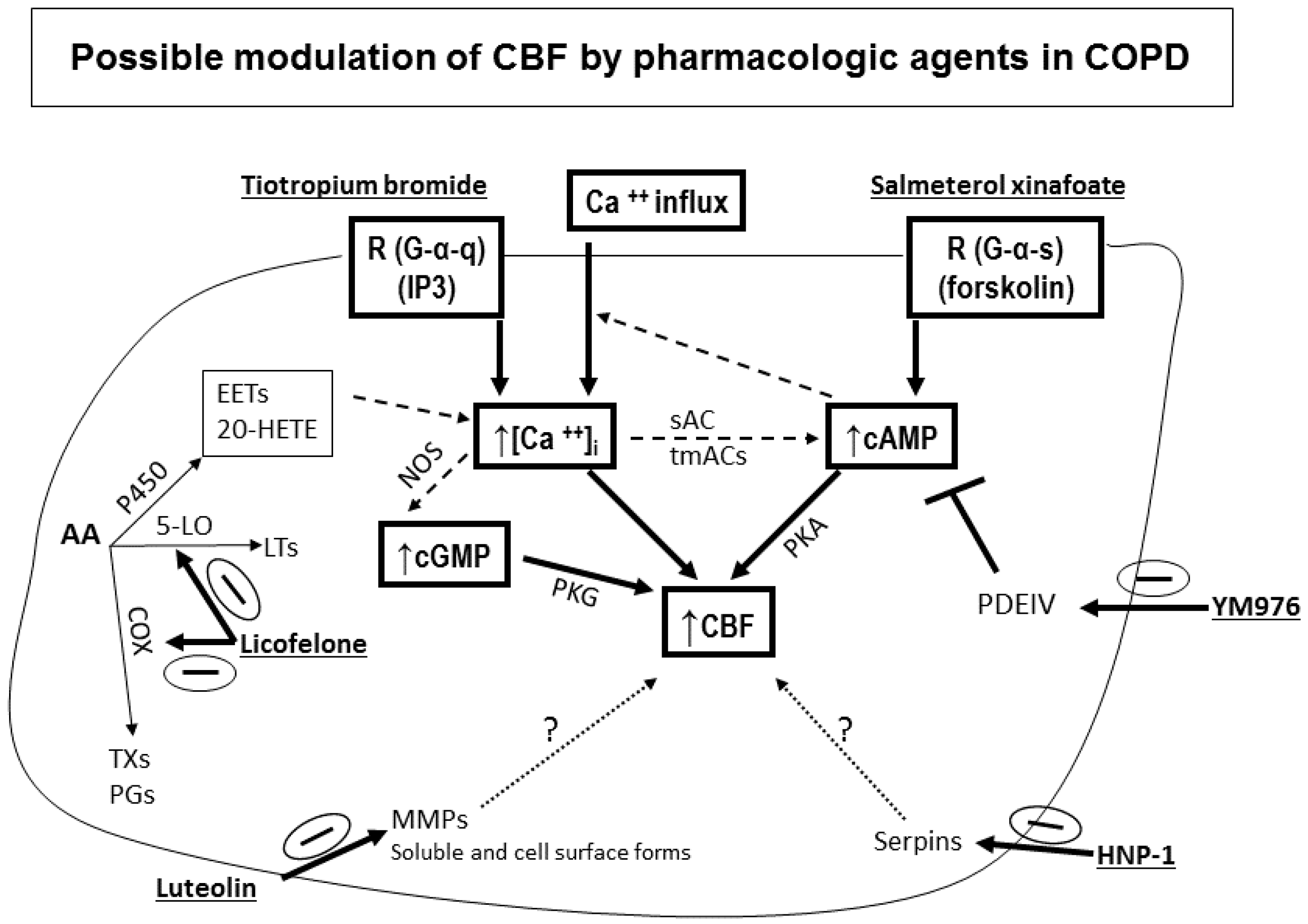
- Effect of smoking: Mucociliary clearance has been demonstrated to be affected in smoking [123,124] and while smoke caused cilia loss in bovine lungs studied in vitro [125], ciliary function has been shown to be normal in smoking asymptomatic subjects [64]. When investigating the ultrastructure (TEM) of bronchial epithelial cell cilia, significant ciliary abnormalities in smokers and ex-smokers compared to non-smokers and controls have been observed with the higher percentage of non-specific ciliary abnormalities associated with chronic tobacco smoke in those with chronic sputum production [126], a characteristic symptom of COPD [109,127]. Earlier studies into the effect of smoking on cilia abundance have yielded conflicting results. Rankin et al. (2007) observed a smoking-related difference in cilia abundance in explanted human bronchial tissue [128] similar to that previously described in bronchial [129] and tracheal explants [130]. Other studies, into cilia abnormalities in asymptomatic smokers and nonsmokers, have yielded conflicting results with no difference between asymptomatic smokers and nonsmokers observed in nasal brushings [7,55]. In contrast, Rutland et al. (1983) and Fox et al. (1983), studied cilia ultrastructure in nasal brushings and reported significantly more abnormal cilia in smokers than in non-smokers [131,132]. In a study of bronchial epithelium from normal smokers and matched nonsmokers, the small airway samples (10th to 12th order) had a higher proportion of ciliated cells compared to samples from large (second to third order) bronchi, and smoking caused a decline in the proportion of ciliated cells at both sites [46]. Another study of bronchial biopsies suggested that smoking-associated shorter airway epithelial cilia could play a role in the pathogenesis of smoking-induced lung disease [48]. Cigarette smoke exposure of C57BL/6 mice for 6–12 month was associated with slowed ciliary motility, decreased ciliated cell numbers, and an impaired ciliary response to beta-agonists [133]. In recent studies, exposure of the human epithelium in vitro to cigarette smoke extracts resulted in a broad suppression of genes involved in ciliogenesis [134], and evaluations of cilia lengths from bronchial biopsies or brushings of healthy smokers and nonsmokers showed that smoking is associated with shorter cilia [48,135]. In the large and small airway epithelium, cilia were significantly shorter in COPD smokers compared to healthy smokers and nonsmokers. However, when investigating the gene expression data of the intraflagellar transport system of cilia (which regulates cilia length), some IFT genes were downregulated in smokers but not in COPD smokers compared to healthy smokers [135]. In our COPD study, nasal CBF was significantly depressed in cilia obtained from moderate and severe COPD subjects compared to At Risk and Control subjects (Figure 7). This significant decline in CBF was not due to a difference in the quality of samples (abundance of cilia), or to cell size, cilia length, or cilia coordination. In addition, since At Risk, and COPD (Moderate, Severe) subjects were smokers/ex-smokers with minimum of 32 pack years, the reduced CBF in COPD cilia only could not be attributed just to the effect of cigarette smoke [52]. In addition, various agents could modulate in vitro cilia function of COPD patients confirming a complicated role for cilia in COPD [52] (Figure 8). We also demonstrated that mannitol has a direct effect on CBF of bronchial cilia [53]. Additionally, airway surface properties (hydration, viscosity) can modulate cilia function in vivo as demonstrated in studies of inhaled mannitol in lung disease patients [38,136,137]. This indicates that further studies of the function and ultrastructure of cilia in COPD are needed.
6. Factors That Can Affect CBF in Asthma
7. Summary and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Powles-Glover, N. Cilia and ciliopathies: Classic examples linking phenotype and genotype-an overview. Reprod. Toxicol. 2014, 48, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Werner, C.; Onnebrink, J.G.; Omran, H. Diagnosis and management of primary ciliary dyskinesia. Cilia 2015, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.R.; Daniels, L.A.; Davis, S.D.; Zariwala, M.A.; Leigh, M.W. Primary ciliary dyskinesia. Recent advances in diagnostics, genetics, and characterization of clinical disease. Am. J. Respir. Crit. Care Med. 2013, 188, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Rossman, C.M.; Forrest, J.B.; Lee, R.M.; Newhouse, A.F.; Newhouse, M.T. The dyskinetic cilia syndrome; abnormal ciliary motility in association with abnormal ciliary ultrastructure. Chest 1981, 80 (Suppl. S6), 860–865. [Google Scholar] [PubMed]
- Leigh, M.W.; Pittman, J.E.; Carson, J.L.; Ferkol, T.W.; Dell, S.D.; Davis, S.D.; Knowles, M.R.; Zariwala, M.A. Clinical and genetic aspects of primary ciliary dyskinesia/Kartagener syndrome. Genet. Med. 2009, 11, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Gudis, D.; Zhao, K.Q.; Cohen, N.A. Acquired cilia dysfunction in chronic rhinosinusitis. Am. J. Rhinol. Allergy 2012, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rossman, C.M.; Lee, R.M.; Forrest, J.B.; Newhouse, M.T. Nasal cilia in normal man, primary ciliary dyskinesia and other respiratory diseases: Analysis of motility and ultrastructure. Eur. J. Respir. Dis. Suppl. 1983, 127, 64–70. [Google Scholar] [PubMed]
- Rutland, J.; Griffin, W.M.; Cole, P.J. Human ciliary beat frequency in epithelium from intrathoracic and extrathoracic airways. Am. Rev. Respir. Dis. 1982, 125, 100–105. [Google Scholar] [PubMed]
- Yager, J.A.; Ellman, H.; Dulfano, M.J. Human ciliary beat frequency at three levels of the tracheobronchial tree. Am. Rev. Respir. Dis. 1980, 121, 661–665. [Google Scholar] [PubMed]
- Wanner, A.; Salathe, M.; O’Riordan, T.G. Mucociliary clearance in the airways. Am. J. Respir. Crit. Care Med. 1996, 154 Pt 1, 1868–1902. [Google Scholar] [CrossRef] [PubMed]
- Braiman, A.; Priel, Z. Efficient mucociliary transport relies on efficient regulation of ciliary beating. Respir. Physiol. Neurobiol. 2008, 163, 202–207. [Google Scholar] [CrossRef] [PubMed]
- McDougall, C.M.; Blaylock, M.G.; Douglas, J.G.; Brooker, R.J.; Helms, P.J.; Walsh, G.M. Nasal Epithelial Cells as Surrogates of Bronchial Epithelial Cells in Airway Inflammation Studies. Am. J. Respir. Cell Mol. Biol. 2008, 39, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Salathe, M. Regulation of mammalian ciliary beating. Annu. Rev. Physiol. 2007, 69, 401–422. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, H.; Marshall, W.F. Mechanobiology of Ciliogenesis. BioScience 2014, 64, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Tilley, A.E.; Walters, M.S.; Shaykhiev, R.; Crystal, R.G. Cilia dysfunction in lung disease. Annu. Rev. Physiol. 2015, 77, 379–406. [Google Scholar] [CrossRef] [PubMed]
- Merkus, P.; Romeijn, S.G.; Verhoef, J.C.; Merkus, F.W.; Schouwenburg, P.F. Classification of cilio-inhibiting effects of nasal drugs. Laryngoscope 2001, 111 Pt 1, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Rusznak, C.; Devalia, J.L.; Lozewicz, S.; Davies, R.J. The assessment of nasal mucociliary clearance and the effect of drugs. Respir. Med. 1994, 88, 89–101. [Google Scholar] [CrossRef]
- Dowling, R.B.; Wilson, R. Bacterial toxins which perturb ciliary function and respiratory epithelium. Symp. Ser. Soc. Appl. Microbiol. 1998, 27, 138S–148S. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, T.; Gugatschga, M.; Koidl, B.; Wolf, G. Influence of preservatives and topical steroids on ciliary beat frequency in vitro. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Workman, A.D.; Cohen, N.A. The effect of drugs and other compounds on the ciliary beat frequency of human respiratory epithelium. Am. J. Rhinol. Allergy 2014, 28, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Salathe, M. Effects of beta-agonists on airway epithelial cells. J. Allergy Clin. Immunol. 2002, 110 (Suppl. S6), S275–S281. [Google Scholar] [CrossRef] [PubMed]
- Stannard, W.; O’Callaghan, C. Ciliary function and the role of cilia in clearance. J. Aerosol Med. 2006, 19, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.F.; Nonaka, S. Cilia: Tuning in to the cell’s antenna. Curr. Biol. 2006, 16, R604–R614. [Google Scholar] [CrossRef] [PubMed]
- Satir, P.; Christensen, S.T. Overview of structure and function of mammalian cilia. Annu. Rev. Physiol 2007, 69, 377–400. [Google Scholar] [CrossRef] [PubMed]
- Chilvers, M.A.; Rutman, A.; O’Callaghan, C. Ciliary beat pattern is associated with specific ultrastructural defects in primary ciliary dyskinesia. J. Allergy Clin. Immunol. 2003, 112, 518–524. [Google Scholar] [CrossRef]
- Willems, T.; Jorissen, M. Correlations between ciliary structure and ciliary function. Acta Otorhinolaryngol. Belg. 2000, 54, 299–308. [Google Scholar] [PubMed]
- Jorissen, M. Correlations among mucociliary transport, ciliary function, and ciliary structure. Am. J. Rhinol. 1998, 12, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.D.; Ferkol, T.W.; Rosenfeld, M.; Lee, H.S.; Dell, S.D.; Sagel, S.D.; Milla, C.; Zariwala, M.A.; Pittman, J.E.; Shapiro, A.J.; et al. Clinical features of childhood primary ciliary dyskinesia by genotype and ultrastructural phenotype. Am. J. Respir. Crit. Care Med. 2015, 191, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Mullowney, T.; Manson, D.; Kim, R.; Stephens, D.; Shah, V.; Dell, S. Primary ciliary dyskinesia and neonatal respiratory distress. Pediatrics 2014, 134, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.R.; Ostrowski, L.E.; Loges, N.T.; Hurd, T.; Leigh, M.W.; Huang, L.; Wolf, W.E.; Carson, J.L.; Hazucha, M.J.; Yin, W.; et al. Mutations in SPAG1 cause primary ciliary dyskinesia associated with defective outer and inner dynein arms. Am. J. Hum. Genet. 2013, 93, 711–720. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.H.; Hall, A.; Cutz, E.; Knowles, M.R.; Nelligan, K.A.; Nykamp, K.; Zariwala, M.A.; Dell, S.D. The role of molecular genetic analysis in the diagnosis of primary ciliary dyskinesia. Ann. Am. Thorac. Soc. 2014, 11, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Knowles, M.R.; Ostrowski, L.E.; Leigh, M.W.; Sears, P.R.; Davis, S.D.; Wolf, W.E.; Hazucha, M.J.; Carson, J.L.; Olivier, K.N.; Sagel, S.D.; et al. Mutations in RSPH1 cause primary ciliary dyskinesia with a unique clinical and ciliary phenotype. Am. J. Respir. Crit. Care Med. 2014, 189, 707–717. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.J.; Davis, S.D.; Ferkol, T.; Dell, S.D.; Rosenfeld, M.; Olivier, K.N.; Sagel, S.D.; Milla, C.; Zariwala, M.A.; Wolf, W.; et al. Laterality defects other than situs inversus totalis in primary ciliary dyskinesia: Insights into situs ambiguus and heterotaxy. Chest 2014, 146, 1176–1186. [Google Scholar] [CrossRef] [PubMed]
- Fretzayas, A.; Moustaki, M. Clinical spectrum of primary ciliary dyskinesia in childhood. World J. Clin. Pediatr. 2016, 5, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Milla, C.E. The evolving spectrum of ciliopathies and respiratory disease. Curr. Opin. Pediatr. 2016, 28, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.B.; Vasquez, P.A.; Mellnik, J.; McKinley, S.A.; Vose, A.; Mu, F.; Henderson, A.G.; Donaldson, S.H.; Alexis, N.E.; Boucher, R.C.; et al. A biophysical basis for mucus solids concentration as a candidate biomarker for airways disease. PLoS ONE 2014, 9, e87681. [Google Scholar] [CrossRef] [PubMed]
- Button, B.; Boucher, R.C. Role of mechanical stress in regulating airway surface hydration and mucus clearance rates. Respir. Physiol. Neurobiol. 2008, 163, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Rubin, B.K. Effect of inhaled dry powder mannitol on mucus and its clearance. Expert. Rev. Respir. Med. 2013, 7, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Bals, R.; Hiemstra, P.S. Innate immunity in the lung: How epithelial cells fight against respiratory pathogens. Eur. Respir. J. 2004, 23, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, L.M.; Davies, D.E.; Wilson, S.J.; Kimber, I.; Dearman, R.J.; Holgate, S.T. The bronchial epithelium in asthma--much more than a passive barrier. Monaldi Arch. Chest Dis. 2001, 56, 48–54. [Google Scholar] [PubMed]
- Hermans, C.; Bernard, A. Lung epithelium-specific proteins: Characteristics and potential applications as markers. Am. J. Respir. Crit. Care Med. 1999, 159, 646–678. [Google Scholar] [CrossRef] [PubMed]
- Mills, P.R.; Davies, R.J.; Devalia, J.L. Airway epithelial cells, cytokines, and pollutants. Am. J. Respir. Crit. Care Med. 1999, 160 (Suppl. S1), S38–S43. [Google Scholar] [CrossRef] [PubMed]
- Hiemstra, P.S.; van Wetering, S.; Stolk, J. Neutrophil serine proteinases and defensins in chronic obstructive pulmonary disease: Effects on pulmonary epithelium. Eur. Respir. J. 1998, 12, 1200–1208. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Wells, J.M.; Zorn, A.M.; Wert, S.E.; Laubach, V.E.; Fernandez, L.G.; Whitsett, J.A. Transdifferentiation of ciliated cells during repair of the respiratory epithelium. Am. J. Respir. Cell Mol. Biol. 2006, 34, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.S.; Byron, P.R. Inhaling medicines: Delivering drugs to the body through the lungs. Nat. Rev. Drug Discov. 2007, 6, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Harvey, B.G.; Heguy, A.; Leopold, P.L.; Carolan, B.J.; Ferris, B.; Crystal, R.G. Modification of gene expression of the small airway epithelium in response to cigarette smoking. J. Mol. Med. 2007, 85, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Rawlins, E.L.; Hogan, B.L. Ciliated epithelial cell lifespan in the mouse trachea and lung. Am. J. Physiol. Lung Cell. Mol. Physiol. 2008, 295, L231–L234. [Google Scholar] [CrossRef] [PubMed]
- Leopold, P.L.; O’Mahony, M.J.; Lian, X.J.; Tilley, A.E.; Harvey, B.G.; Crystal, R.G. Smoking is associated with shortened airway cilia. PLoS ONE 2009, 4, e8157. [Google Scholar] [CrossRef] [PubMed]
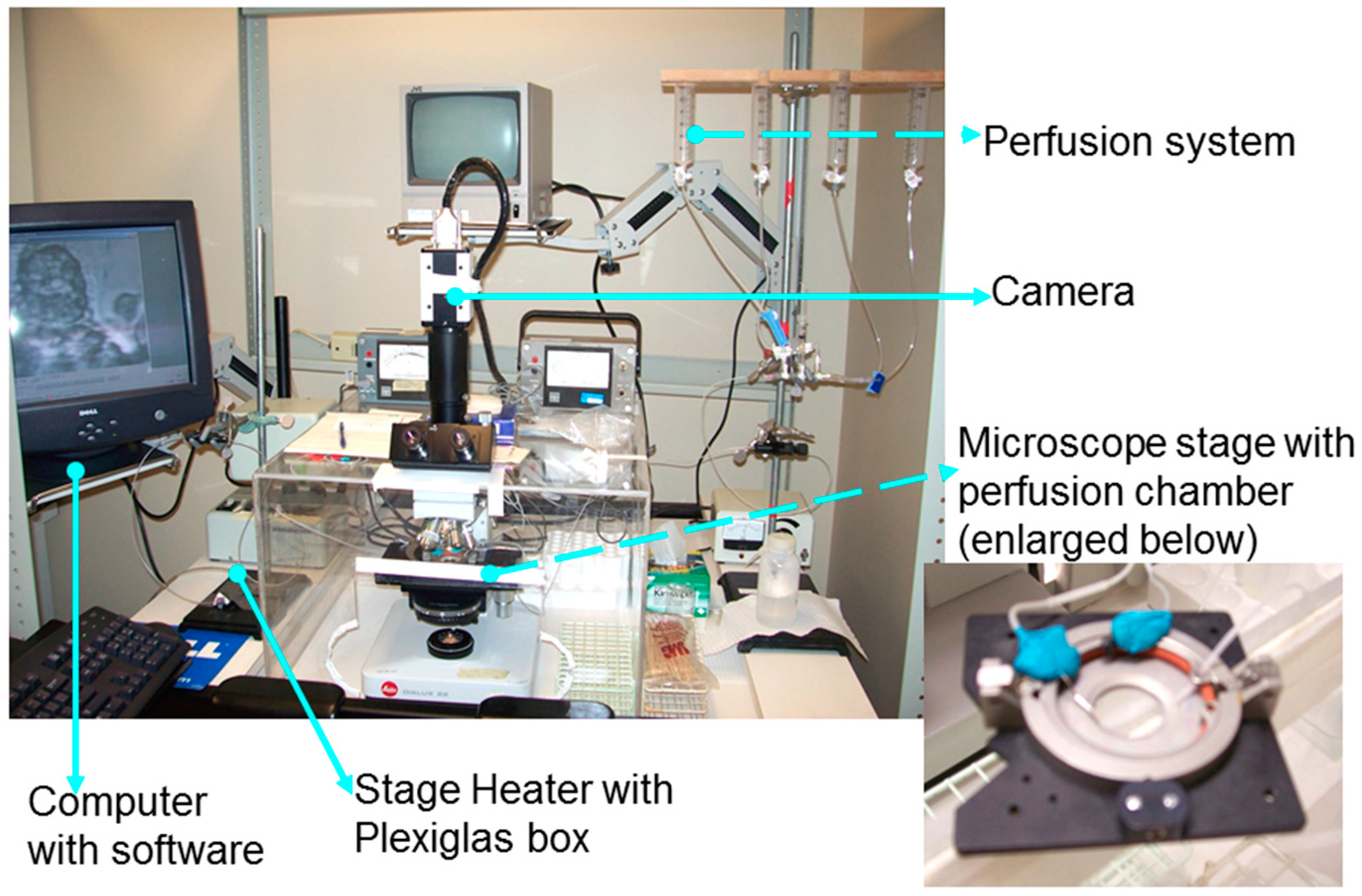
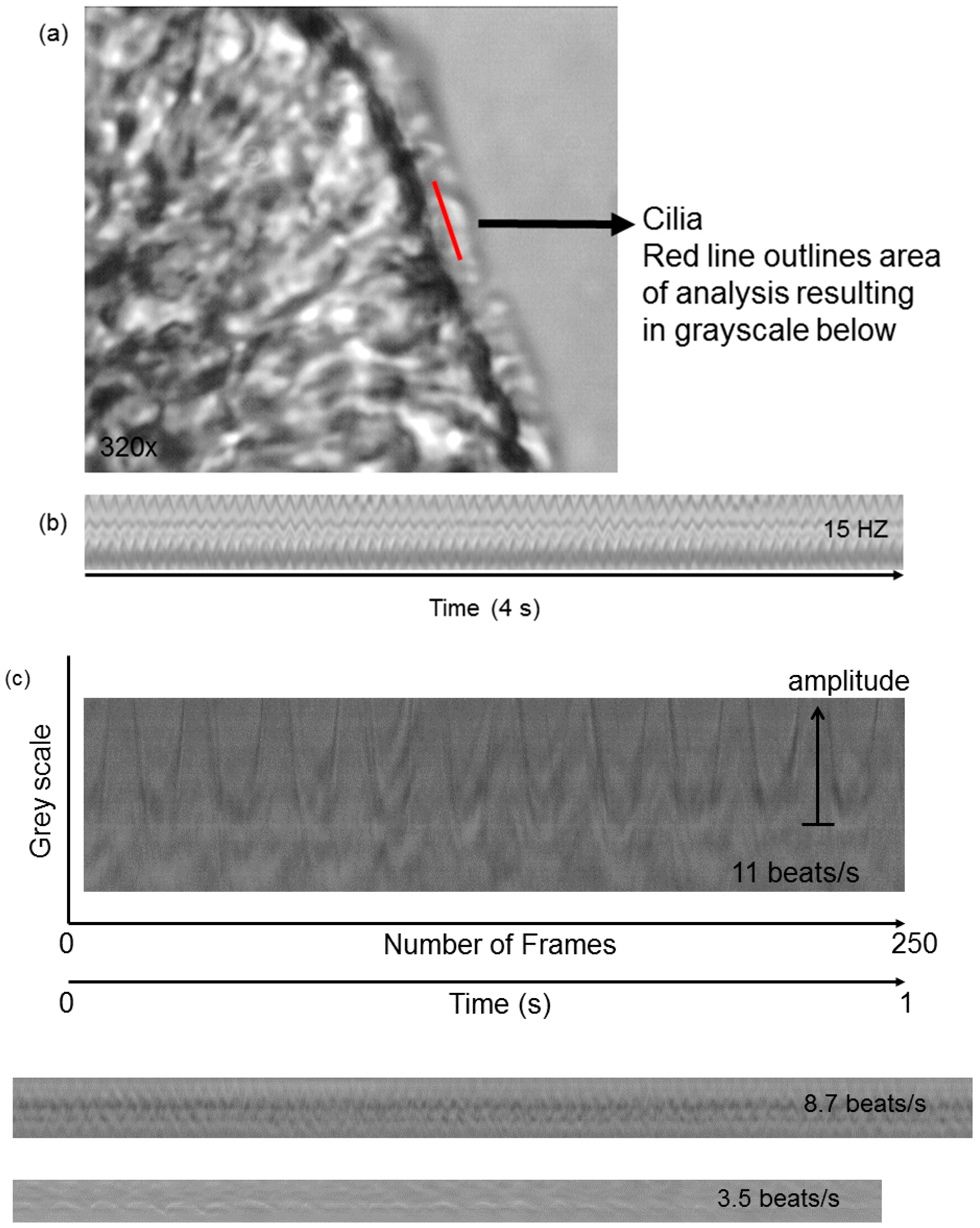
- Yaghi, A.; Dolovich, M. Investigating the Function of Epithelial Cell Cilia Using High Speed Digital Imaging. Am. J. Respir. Crit. Care Med. 2008, 177, A206. [Google Scholar]
- Iida, H.; Matsuura, S.; Shirakami, G.; Tanimoto, K.; Fukuda, K. Differential effects of intravenous anesthetics on ciliary motility in cultured rat tracheal epithelial cells. Can. J. Anaesth. 2006, 53, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Chilvers, M.A.; O’Callaghan, C. Local mucociliary defence mechanisms. Paediatr. Respir. Rev. 2000, 1, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Yaghi, A.; Zaman, A.; Cox, G.; Dolovich, M.B. Ciliary beating is depressed in nasal cilia from chronic obstructive pulmonary disease subjects. Respir. Med. 2012, 106, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Yaghi, A.; Zaman, A.; Dolovich, M.B. The direct effect of hyperosmolar agents on ciliary beating of human bronchial epithelial cells. J. Aerosol Med. Pulm. Drug Deliv. 2012, 25, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Rutland, J.; de Iongh, R.U. Random ciliary orientation. A cause of respiratory tract disease. N. Engl. J. Med. 1990, 323, 1681–1684. [Google Scholar] [CrossRef] [PubMed]
- De Iongh, R.U.; Rutland, J. Ciliary defects in healthy subjects, bronchiectasis, and primary ciliary dyskinesia. Am. J. Respir. Crit. Care Med. 1995, 151, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.; Ben-Shahar, Y.; Moninger, T.O.; Kline, J.N.; Welsh, M.J. Motile Cilia of Human Airway Epithelia Are Chemosensory. Science 2009, 325, 1131–1134. [Google Scholar] [CrossRef] [PubMed]
- Horvath, G.; Sorscher, E.J. Luminal fluid tonicity regulates airway ciliary beating by altering membrane stretch and intracellular calcium. Cell Motil. Cytoskeleton 2008, 65, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Kanthakumar, K.; Taylor, G.W.; Cundell, D.R.; Dowling, R.B.; Johnson, M.; Cole, P.J.; Wilson, R. The effect of bacterial toxins on levels of intracellular adenosine nucleotides and human ciliary beat frequency. Pulm. Pharmacol. 1996, 9, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Agius, A.M.; Wake, M.; Pahor, A.L.; Smallman, A. The effects of in vitro cotitine on nasal ciliary beat frequency. Clin. Otolaryngol. Allied Sci. 1995, 20, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Luk, C.K.; Dulfano, M.J. Effect of pH, viscosity and ionic-strength changes on ciliary beating frequency of human bronchial explants. Clin. Sci. (Lond.) 1983, 64, 449–451. [Google Scholar] [CrossRef] [PubMed]
- Roth, Y.; Aharonson, E.F.; Teichtahl, H.; Baum, G.L.; Priel, Z.; Modan, M. Human in vitro nasal and tracheal ciliary beat frequencies: Comparison of sampling sites, combined effect of medication, and demographic relationships. Ann. Otol. Rhinol. Laryngol. 1991, 100 Pt 1, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Yager, J.; Chen, T.M.; Dulfano, M.J. Measurement of frequency of ciliary beats of human respiratory epithelium. Chest 1978, 73, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Chilvers, M.A.; Rutman, A.; O’Callaghan, C. Functional analysis of cilia and ciliated epithelial ultrastructure in healthy children and young adults. Thorax 2003, 58, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Rossman, C.M.; Lee, R.M.; Forrest, J.B.; Newhouse, M.T. Nasal ciliary ultrastructure and function in patients with primary ciliary dyskinesia compared with that in normal subjects and in subjects with various respiratory diseases. Am. Rev. Respir. Dis. 1984, 129, 161–167. [Google Scholar] [PubMed]
- Satir, P.; Dirksen, E.R. Function-structure correlations in cilia from mammalian respiratory tract. In The Respiratory System Volume 1: Circulation and Nonrespiratory Functions; Weibel, E.R., Ed.; The American Physiological Society: Bethesda, MD, USA, 1985; pp. 473–494. [Google Scholar]
- Pazour, G.J.; Rosenbaum, J.L. Intraflagellar transport and cilia-dependent diseases. Trends Cell Biol. 2002, 12, 551–555. [Google Scholar] [CrossRef]
- Rosenbaum, J.L.; Witman, G.B. Intraflagellar transport. Nat. Rev. Mol. Cell Biol. 2002, 3, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Badano, J.L.; Mitsuma, N.; Beales, P.L.; Katsanis, N. The Ciliopathies: An Emerging Class of Human Genetic Disorders. Annu. Rev. Genomics Hum. Genet. 2006, 7, 125–148. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Forget, M.A.; Adams, J.M.; Sisson, J.H. Both cAMP and cGMP are required for maximal ciliary beat stimulation in a cell-free model of bovine ciliary axonemes. Am. J. Physiol Lung Cell. Mol. Physiol. 2005, 288, L546–L551. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.A.; Spurzem, J.R.; May, K.; Sisson, J.H. Regulation of ciliary beat frequency by both PKA and PKG in bovine airway epithelial cells. Am. J. Physiol 1998, 275 Pt 1, L827–L835. [Google Scholar] [PubMed]
- Zhang, L.; Sanderson, M.J. Oscillations in ciliary beat frequency and intracellular calcium concentration in rabbit tracheal epithelial cells induced by ATP. J. Physiol. 2003, 546 Pt 3, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Sanderson, M.J. The role of cGMP in the regulation of rabbit airway ciliary beat frequency. J. Physiol. 2003, 551 Pt 3, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Schmid, A.; Sutto, Z.; Nlend, M.C.; Horvath, G.; Schmid, N.; Buck, J.; Levin, L.R.; Conner, G.E.; Fregien, N.; Salathe, M. Soluble adenylyl cyclase is localized to cilia and contributes to ciliary beat frequency regulation via production of cAMP. J. Gen. Physiol. 2007, 130, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Sutto, Z.; Conner, G.E.; Salathe, M. Regulation of human airway ciliary beat frequency by intracellular pH. J. Physiol. 2004, 560, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Anderson, S.D. Hyperosmolar agents and clearance of mucus in the diseased airway. J. Aerosol Med. 2006, 19, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Braunstahl, G.J.; Fokkens, W.J.; Overbeek, S.E.; Kleinjan, A.; Hoogsteden, H.C.; Prins, J.B. Mucosal and systemic inflammatory changes in allergic rhinitis and asthma: A comparison between upper and lower airways. Clin. Exp. Allergy 2003, 33, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, B.; Collet, S.; Eloy, P.; Rombaux, P. Secondary ciliary dyskinesia in upper respiratory tract. Acta Otorhinolaryngol. Belg. 2000, 54, 309–316. [Google Scholar] [PubMed]
- Piatti, G.; Ambrosetti, U.; Santus, P.; Allegra, L. Effects of salmeterol on cilia and mucus in COPD and pneumonia patients. Pharmacol. Res. 2005, 51, 165–168. [Google Scholar] [CrossRef] [PubMed]
- BeruBe, K.; Balharry, D.; Sexton, K.; Koshy, L.; Jones, T. Combustion-derived nanoparticles: Mechanisms of pulmonary toxicity. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1044–1050. [Google Scholar] [CrossRef] [PubMed]
- Duffin, R.; Mills, N.L.; Donaldson, K. Nanoparticles-a thoracic toxicology perspective. Yonsei Med. J. 2007, 48, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Monteiller, C.; Tran, L.; MacNee, W.; Faux, S.; Jones, A.; Miller, B.; Donaldson, K. The pro-inflammatory effects of low-toxicity low-solubility particles, nanoparticles and fine particles, on epithelial cells in vitro: The role of surface area. Occup. Environ. Med. 2007, 64, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Pace, E.; Ferraro, M.; Di, V.S.; Gerbino, S.; Bruno, A.; Lanata, L.; Gjomarkaj, M. Oxidative stress and innate immunity responses in cigarette smoke stimulated nasal epithelial cells. Toxicol. In Vitro 2014, 28, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.C.; Cloonan, S.M.; Bhashyam, A.R.; Haspel, J.A.; Singh, A.; Sathirapongsasuti, J.F.; Cervo, M.; Yao, H.; Chung, A.L.; Mizumura, K.; et al. Histone deacetylase 6-mediated selective autophagy regulates COPD-associated cilia dysfunction. J. Clin. Investig. 2013, 123, 5212–5230. [Google Scholar] [CrossRef] [PubMed]
- Pierrou, S.; Broberg, P.; O’donnell, R.A.; Pawlowski, K.; Virtala, R.; Lindqvist, E.; Richter, A.; Wilson, S.J.; Angco, G.; Moller, S.; et al. Expression of genes involved in oxidative stress responses in airway epithelial cells of smokers with chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2007, 175, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Tuder, R.M. Pathobiology of cigarette smoke-induced chronic obstructive pulmonary disease. Physiol. Rev. 2007, 87, 1047–1082. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Anderson, S.D.; Young, I.H. Effect of mannitol and repetitive coughing on the sputum properties in bronchiectasis. Respir. Med. 2010, 104, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Robinson, M.; Anderson, S.D.; Bye, P.T. Osmotic stimuli increase clearance of mucus in patients with mucociliary dysfunction. J. Aerosol Med. 2002, 15, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Newhouse, M.T.; Rossman, C.M.; Dolovich, J.; Dolovich, M.B.; Wilson, W.M. Impairment of mucociliary transport in cystic fibrosis. Mod. Probl. Paediatr. 1976, 19, 190–198. [Google Scholar] [PubMed]
- Rossman, C.; Dolovich, J.; Dolovich, M.; Wilson, W.; Newhouse, M. Cystic fibrosis--related inhibition of mucociliary clearance in vivo in man. J. Pediatr. 1977, 90, 579–584. [Google Scholar] [CrossRef]
- Puchelle, E.; Zahm, J.M. Influence of rheological properties of human bronchial secretions on the ciliary beat frequency. Biorheology 1984, 21, 265–272. [Google Scholar] [PubMed]
- Veale, D.; Rodgers, A.D.; Griffiths, C.J.; Ashcroft, T.; Gibson, G.J. Variability in ciliary beat frequency in normal subjects and in patients with bronchiectasis. Thorax 1993, 48, 1018–1020. [Google Scholar] [CrossRef] [PubMed]
- Chapelin, C.; Coste, A.; Gilain, L.; Poron, F.; Verra, F.; Escudier, E. Modified epithelial cell distribution in chronic airways inflammation. Eur. Respir. J. 1996, 9, 2474–2478. [Google Scholar] [CrossRef] [PubMed]
- Joki, S.; Toskala, E.; Saano, V.; Nuutinen, J. Correlation between ciliary beat frequency and the structure of ciliated epithelia in pathologic human nasal mucosa. Laryngoscope 1998, 108, 426–430. [Google Scholar] [CrossRef] [PubMed]
- Hisamatsu, K.; Nakajima, M. Pranlukast protects leukotriene C4- and D4-induced epithelial cell impairment of the nasal mucosa in vitro. Life Sci. 2000, 67, 2767–2773. [Google Scholar] [CrossRef]
- Sabater, J.R.; Wanner, A.; Abraham, W.M. Montelukast prevents antigen-induced mucociliary dysfunction in sheep. Am. J. Respir. Crit. Care Med. 2002, 166, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Trevisani, L.; Sartori, S.; Pazzi, P.; Potena, A.; Aleotti, A.; Grandi, E.; Stabellini, G. Acquired ciliary defects in bronchial epithelium of patients with chronic bronchitis. Respiration 1990, 57, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.; Roberts, D.; Cole, P. Effect of bacterial products on human ciliary function in vitro. Thorax 1985, 40, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Tristram, D.A.; Hicks, W., Jr.; Hard, R. Respiratory syncytial virus and human bronchial epithelium. Arch. Otolaryngol. Head Neck Surg. 1998, 124, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Carson, J.L.; Collier, A.M.; Hu, S.S. Acquired ciliary defects in nasal epithelium of children with acute viral upper respiratory infections. N. Engl. J. Med. 1985, 312, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.Y.; Rutman, A.; O’Callaghan, C. Recovery of the ciliated epithelium following acute bronchiolitis in infancy. Thorax 2005, 60, 582–587. [Google Scholar] [CrossRef] [PubMed]
- Low, P.M.; Luk, C.K.; Dulfano, M.J.; Finch, P.J. Ciliary beat frequency of human respiratory tract by different sampling techniques. Am. Rev. Respir. Dis. 1984, 130, 497–498. [Google Scholar] [PubMed]
- Boek, W.M.; Graamans, K.; Natzijl, H.; van Rijk, P.P.; Huizing, E.H. Nasal mucociliary transport: New evidence for a key role of ciliary beat frequency. Laryngoscope 2002, 112, 570–573. [Google Scholar] [CrossRef] [PubMed]
- Verra, F.; Fleury-Feith, J.; Boucherat, M.; Pinchon, M.C.; Bignon, J.; Escudier, E. Do nasal ciliary changes reflect bronchial changes? An ultrastructural study. Am. Rev. Respir. Dis. 1993, 147, 908–913. [Google Scholar] [CrossRef] [PubMed]
- Hanes, L.S.; Issa, E.; Proud, D.; Togias, A. Stronger nasal responsiveness to cold air in individuals with rhinitis and asthma, compared with rhinitis alone. Clin. Exp. Allergy 2006, 36, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Benson, M.; Jansson, L.; Adner, M.; Luts, A.; Uddman, R.; Cardell, L.O. Gene profiling reveals decreased expression of uteroglobin and other anti-inflammatory genes in nasal fluid cells from patients with intermittent allergic rhinitis. Clin. Exp. Allergy 2005, 35, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Devalia, J.L.; Sapsford, R.J.; Rusznak, C.; Toumbis, M.J.; Davies, R.J. The effects of salmeterol and salbutamol on ciliary beat frequency of cultured human bronchial epithelial cells, in vitro. Pulm. Pharmacol. 1992, 5, 257–263. [Google Scholar] [CrossRef]
- Hermens, W.A.; Merkus, F.W. The influence of drugs on nasal ciliary movement. Pharm. Res. 1987, 4, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Romeijn, S.G.; Verhoef, J.C.; Marttin, E.; Merkus, F.W. The effect of nasal drug formulations on ciliary beating in vitro. Int. J. Pharm. 1996, 135, 137–145. [Google Scholar] [CrossRef]
- WatermarkedGlobal-Strategy-2016.pdf. From the Global Strategy for the Diagnosis, Management and Prevention of COPD, Global Initiative for Chronic Obstructive Lung Disease (GOLD) 2016. Available online: http://goldcopd.org/ (accessed on 8 November 2016).
- Barnes, P.J.; Stockley, R.A. COPD: Current therapeutic interventions and future approaches. Eur. Respir. J. 2005, 25, 1084–1106. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Emerging pharmacotherapies for COPD. Chest 2008, 134, 1278–1286. [Google Scholar] [CrossRef] [PubMed]
- O’donnell, R.A.; Peebles, C.; Ward, J.A.; Daraker, A.; Angco, G.; Broberg, P.; Pierrou, S.; Lund, J.; Holgate, S.T.; Davies, D.E.; et al. Relationship between peripheral airway dysfunction, airway obstruction, and neutrophilic inflammation in COPD. Thorax 2004, 59, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Lapperre, T.S.; Postma, D.S.; Gosman, M.M.; Snoeck-Stroband, J.B.; ten Hacken, N.H.; Hiemstra, P.S.; Timens, W.; Sterk, P.J.; Mauad, T. Relation between duration of smoking cessation and bronchial inflammation in COPD. Thorax 2006, 61, 115–121. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, R.; Breen, D.; Wilson, S.; Djukanovic, R. Inflammatory cells in the airways in COPD. Thorax 2006, 61, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Hurst, J.R.; Wilkinson, T.M.; Perera, W.R.; Donaldson, G.C.; Wedzicha, J.A. Relationships among bacteria, upper airway, lower airway, and systemic inflammation in COPD. Chest 2005, 127, 1219–1226. [Google Scholar] [CrossRef]
- Tamaoki, J.; Sakai, N.; Kobayashi, K.; Kanemura, T.; Takizawa, T. Stimulation of airway ciliary motility by immunologically activated canine pulmonary macrophages: Role of leukotrienes. Acta Physiol. Scand. 1991, 141, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Devalia, J.L.; Davies, R.J. Human nasal and bronchial epithelial cells in culture: An overview of their characteristics and function. Allergy Proc. 1991, 12, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Taggart, C.C.; Greene, C.M.; Carroll, T.P.; O’neill, S.J.; McElvaney, N.G. Elastolytic proteases: Inflammation resolution and dysregulation in chronic infective lung disease. Am. J. Respir. Crit. Care Med. 2005, 171, 1070–1076. [Google Scholar] [CrossRef] [PubMed]
- Van Wetering, S.; Sterk, P.J.; Rabe, K.F.; Hiemstra, P.S. Defensins: Key players or bystanders in infection, injury, and repair in the lung? J. Allergy Clin. Immunol. 1999, 104, 1131–1138. [Google Scholar] [CrossRef]
- Montuschi, P.; Kharitonov, S.A.; Ciabattoni, G.; Barnes, P.J. Exhaled leukotrienes and prostaglandins in COPD. Thorax 2003, 58, 585–588. [Google Scholar] [CrossRef] [PubMed]
- Rossman, C.M.; Newhouse, M.T. Primary ciliary dyskinesia: Evaluation and management. Pediatr. Pulmonol. 1988, 5, 36–50. [Google Scholar] [CrossRef] [PubMed]
- Dolovich, M.B.; Mahony, J.B.; Chambers, C.; Newhouse, M.T.; Chernesky, M.A. Ciliary function, cell viability, and in vitro effect of ribavirin on nasal epithelial cells in acute rhinorrhea. Chest 1992, 102, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Wolff, R.K. Effects of airborne pollutants on mucociliary clearance. Environ. Health Perspect. 1986, 66, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P.J.; Wilson, R.; Greenstone, M.A.; MacWilliam, L.; Cole, P.J. Effect of cigarette smoking on nasal mucociliary clearance and ciliary beat frequency. Thorax 1986, 41, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Sisson, J.H.; Papi, A.; Beckmann, J.D.; Leise, K.L.; Wisecarver, J.; Brodersen, B.W.; Kelling, C.L.; Spurzem, J.R.; Rennard, S.I. Smoke and viral infection cause cilia loss detectable by bronchoalveolar lavage cytology and dynein ELISA. Am. J. Respir. Crit. Care Med. 1994, 149, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Verra, F.; Escudier, E.; Lebargy, F.; Bernaudin, J.F.; De, C.H.; Bignon, J. Ciliary abnormalities in bronchial epithelium of smokers, ex-smokers, and nonsmokers. Am. J. Respir. Crit. Care Med. 1995, 151 Pt 1, 630–634. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, D.E.; Aaron, S.; Bourbeau, J.; Hernandez, P.; Marciniuk, D.; Balter, M.; Ford, G.; Gervais, A.; Goldstein, R.; Hodder, R.; et al. State of the Art Compendium: Canadian Thoracic Society recommendations for the management of chronic obstructive pulmonary disease. Can. Respir. J. 2004, 11 (Suppl. B), 7B–59B. [Google Scholar] [CrossRef] [PubMed]
- Rankin, H.V.; Moody, A.J.; Moate, R.M.; Macnaughton, P.D.; Rahamim, J.; Smith, M.E.; Sneyd, J.R. Elevated oxygen fraction reduces cilial abundance in explanted human bronchial tissue. Ultrastruct. Pathol. 2007, 31, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, O.; Hammond, E.C.; Garfinkel, L. Bronchial epithelium and cigarette smoking. N. Engl. J. Med. 1979, 300, 1395–1396. [Google Scholar] [PubMed]
- Hobson, J.; Wright, J.; Churg, A. Histochemical evidence for generation of active oxygen species on the apical surface of cigarette-smoke-exposed tracheal explants. Am. J. Pathol. 1991, 139, 573–580. [Google Scholar] [PubMed]
- Rutland, J.; Cox, T.; Dewar, A.; Cole, P. Screening for ciliary dyskinesia - a spectrum of defects of motility and structure. Eur. J. Respir. Dis. Suppl. 1983, 127, 71–77. [Google Scholar] [PubMed]
- Fox, B.; Bull, T.B.; Oliver, T.N. The distribution and assessment of electron-microscopic abnormalities of human cilia. Eur. J. Respir. Dis. Suppl 1983, 127, 11–18. [Google Scholar] [PubMed]
- Wyatt, T.A.; Sisson, J.H.; Pavlik, J.A.; Devasure, J.; Boyer, C.; Liu, X.; Kawasaki, S.; Rennard, S.I. Long-Term Cigarette Smoke Exposure in a Mouse Model of Ciliated Epithelial Cell Function. Am. J. Respir. Crit. Care Med. 2010, 43, 634–640. [Google Scholar]
- Brekman, A.; Walters, M.S.; Tilley, A.E.; Crystal, R.G. FOXJ1 prevents cilia growth inhibition by cigarette smoke in human airway epithelium in vitro. Am. J. Respir. Cell Mol. Biol. 2014, 51, 688–700. [Google Scholar] [CrossRef] [PubMed]
- Hessel, J.; Heldrich, J.; Fuller, J.; Staudt, M.R.; Radisch, S.; Hollmann, C.; Harvey, B.G.; Kaner, R.J.; Salit, J.; Yee-Levin, J.; et al. Intraflagellar transport gene expression associated with short cilia in smoking and COPD. PLoS ONE 2014, 9, e85453. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Anderson, S.D.; Gomes, K.; Briffa, P.; Cochrane, B.; Chan, H.K.; Young, I.H.; Rubin, B.K. Inhaled mannitol for the treatment of mucociliary dysfunction in patients with bronchiectasis: Effect on lung function, health status and sputum. Respirology 2005, 10, 46–56. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Anderson, S.D.; Jaques, A.; Charlton, B. Inhaled Mannitol Improves the Hydration and Surface properties of Sputum in patients with Cystic Fibrosis. Chest 2010, 137, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Cutz, E.; Levison, H.; Cooper, D.M. Ultrastructure of airways in children with asthma. Histopathology 1978, 2, 407–421. [Google Scholar] [CrossRef] [PubMed]
- Dulfano, M.J.; Luk, C.K.; Beckage, M.; Wooten, O. Ciliary inhibitory effects of asthma patients’ sputum. Clin. Sci. (Lond.) 1982, 63, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Dulfano, M.J.; Luk, C.K. Sputum and ciliary inhibition in asthma. Thorax 1982, 37, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Mezey, R.J.; Cohn, M.A.; Fernandez, R.J.; Januszkiewicz, A.J.; Wanner, A. Mucociliary transport in allergic patients with antigen-induced bronchospasm. Am. Rev. Respir. Dis. 1978, 118, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Wanner, A. Allergic mucociliary dysfunction. Laryngoscope 1983, 93, 68–70. [Google Scholar] [CrossRef] [PubMed]
- Wanner, A. Allergic mucociliary dysfunction. J. Allergy Clin. Immunol. 1983, 72, 347–350. [Google Scholar] [CrossRef]
- Wanner, A. The role of mucociliary dysfunction in bronchial asthma. Am. J. Med. 1979, 67, 477–485. [Google Scholar] [CrossRef]
- Daviskas, E.; Anderson, S.D.; Gonda, I.; Eberl, S.; Meikle, S.; Seale, J.P.; Bautovich, G. Inhalation of hypertonic saline aerosol enhances mucociliary clearance in asthmatic and healthy subjects. Eur. Respir. J. 1996, 9, 725–732. [Google Scholar] [CrossRef] [PubMed]
- Daviskas, E.; Anderson, S.D.; Brannan, J.D.; Chan, H.K.; Eberl, S.; Bautovich, G. Inhalation of dry-powder mannitol increases mucociliary clearance. Eur. Respir. J. 1997, 10, 2449–2454. [Google Scholar] [CrossRef] [PubMed]
- Heino, M.; Karjalainen, J.; Ylikoski, J.; Laitinen, A.; Laitinen, L.A. Bronchial ciliogenesis and oral steroid treatment in patients with asthma. Br. J. Dis. Chest 1988, 82, 175–178. [Google Scholar] [CrossRef]
- Agnew, J.E.; Bateman, J.R.; Sheahan, N.F.; Lennard-Jones, A.M.; Pavia, D.; Clarke, S.W. Effect of oral corticosteroids on mucus clearance by cough and mucociliary transport in stable asthma. Bull. Eur. Physiopathol. Respir. 1983, 19, 37–41. [Google Scholar] [PubMed]
- Lundgren, R.; Soderberg, M.; Horstedt, P.; Stenling, R. Morphological studies of bronchial mucosal biopsies from asthmatics before and after ten years of treatment with inhaled steroids. Eur. Respir. J. 1988, 1, 883–889. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agents/Factors that Increase CBF | Agents/Factors that Decrease CBF | Agents/Factors that Have No Effect |
|---|---|---|
|
|
|
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yaghi, A.; Dolovich, M.B. Airway Epithelial Cell Cilia and Obstructive Lung Disease. Cells 2016, 5, 40. https://doi.org/10.3390/cells5040040
Yaghi A, Dolovich MB. Airway Epithelial Cell Cilia and Obstructive Lung Disease. Cells. 2016; 5(4):40. https://doi.org/10.3390/cells5040040
Chicago/Turabian StyleYaghi, Asma, and Myrna B. Dolovich. 2016. "Airway Epithelial Cell Cilia and Obstructive Lung Disease" Cells 5, no. 4: 40. https://doi.org/10.3390/cells5040040
APA StyleYaghi, A., & Dolovich, M. B. (2016). Airway Epithelial Cell Cilia and Obstructive Lung Disease. Cells, 5(4), 40. https://doi.org/10.3390/cells5040040